
Nucleolin Aptamer N6L Reprograms the Translational Machinery and Acts Synergistically with mTORi to Inhibit Pancreatic Cancer Proliferation

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Cell Lines
2.2. Spheroid and Organoid Formation
2.3. Viability Assay (MTS) and Apoptosis Analysis
2.4. Analysis of Synergy/Antagonism from Combination Studies
2.5. Dual-Luciferase Reporter Assay for 5′TOP Constructs
2.6. SUnSET for Global Protein Synthesis, Western Blots
2.7. Polysome Profiling and RNA Extraction and Sequencing
2.8. Statistics
3. Results
3.1. NCL Targeting by N6L Impairs Protein Synthesis and Induces a Translational Reprogramming in mPDAC Cells
3.2. N6L Induces a Global Decrease in mRNA Translation Correlated with a Decrease in EiF3 mRNAs Translation
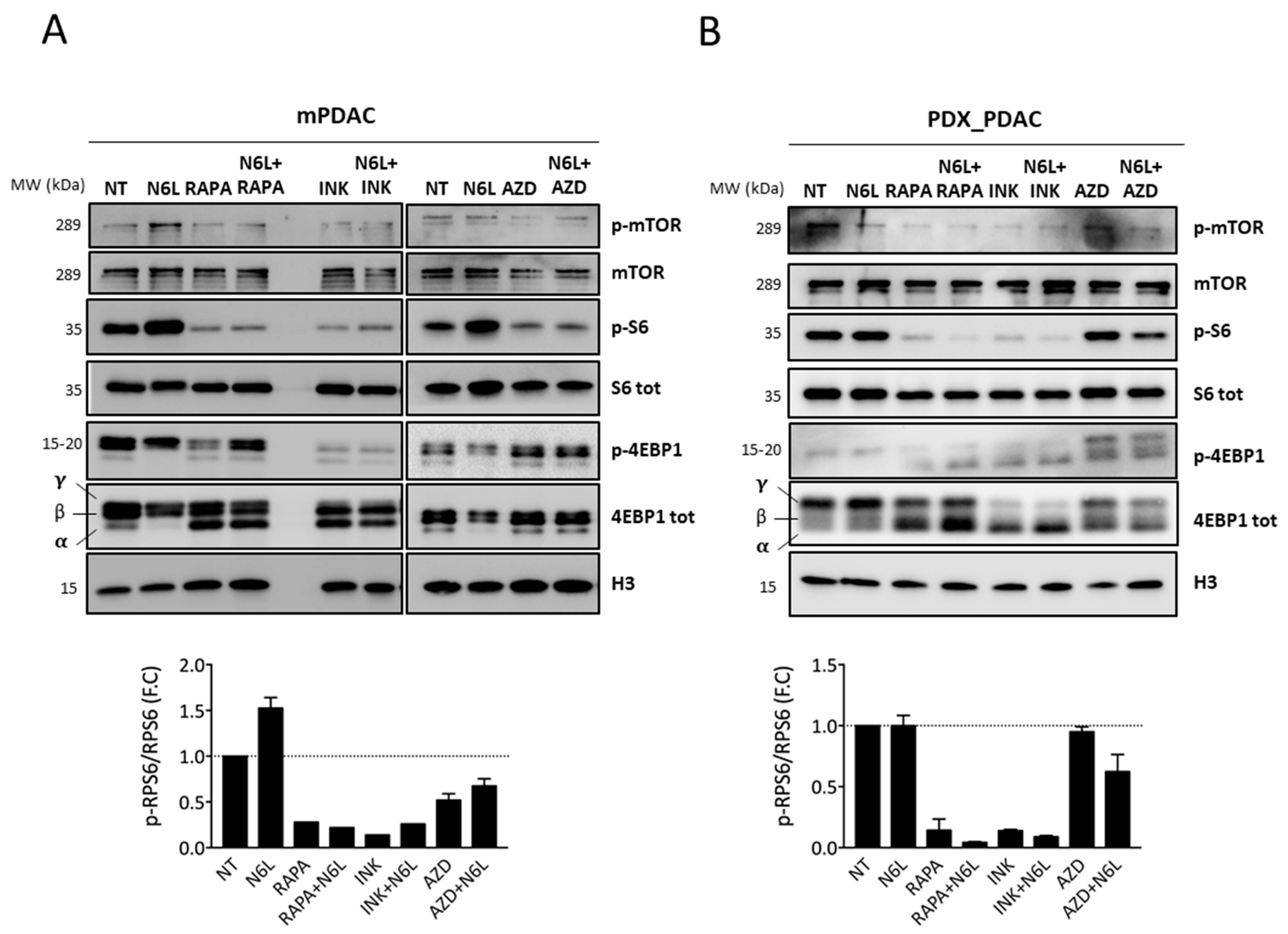
3.3. NCL Targeting by N6L Increases 5′TOP mRNA Translation by Activating the mTOR Pathway
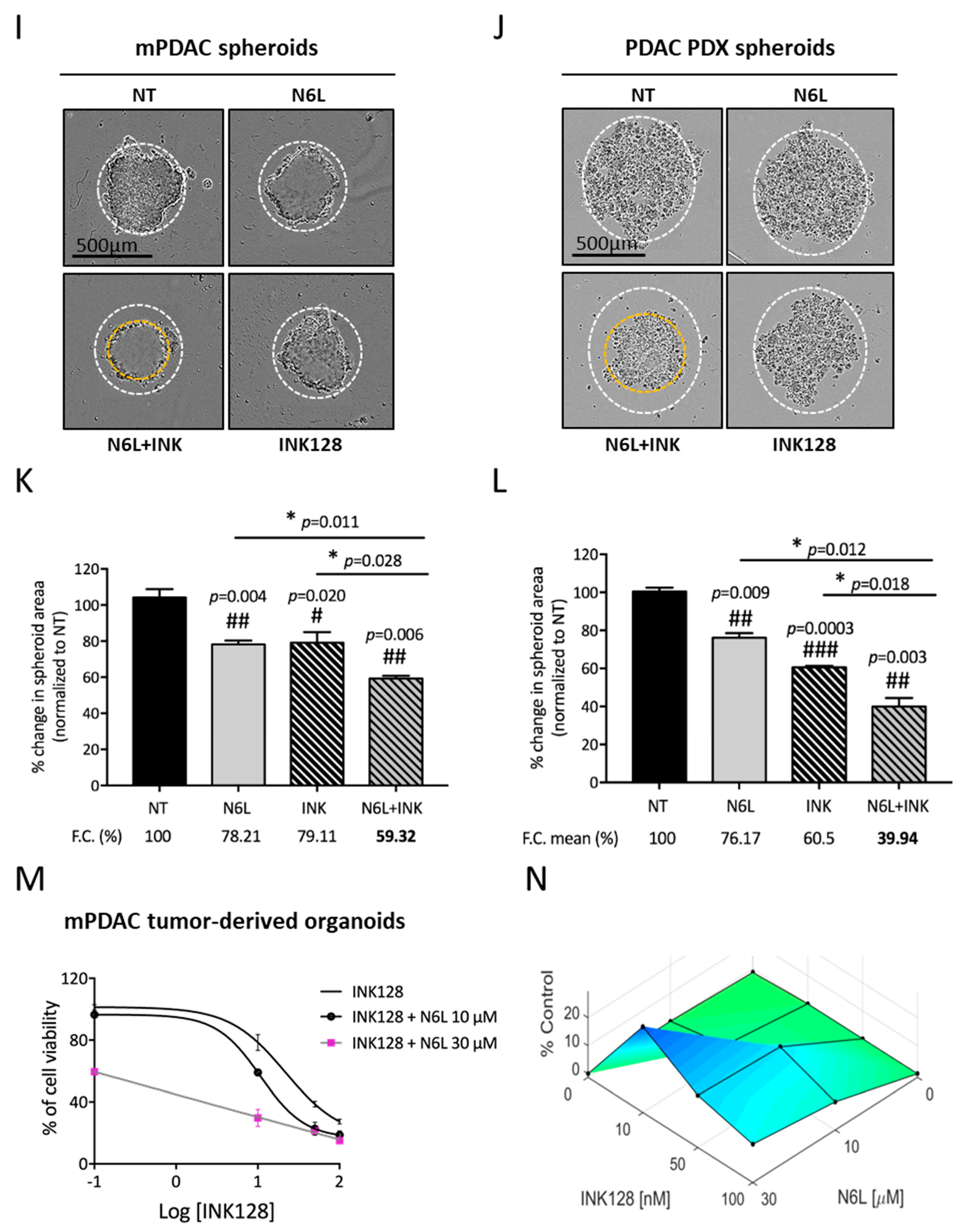
3.4. Combinations of N6L and mTORi Are Synergistic on PDAC Cell Growth and Viability Inhibition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kamisawa, T.; Wood, L.D.; Itoi, T.; Takaori, K. Pancreatic Cancer. Lancet 2016, 388, 73–85. [Google Scholar] [CrossRef]
- Søreide, K.; Primavesi, F.; Labori, K.J.; Watson, M.M.; Stättner, S. Molecular Biology in Pancreatic Ductal Adenocarcinoma: Implications for Future Diagnostics and Therapy. Eur. Surg. 2019, 51, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Bachet, J.-B.; Hammel, P.; Desramé, J.; Meurisse, A.; Chibaudel, B.; André, T.; Debourdeau, P.; Dauba, J.; Lecomte, T.; Seitz, J.-F.; et al. Nab-Paclitaxel plus Either Gemcitabine or Simplified Leucovorin and Fluorouracil as First-Line Therapy for Metastatic Pancreatic Adenocarcinoma (AFUGEM GERCOR): A Non-Comparative, Multicentre, Open-Label, Randomised Phase 2 Trial. Lancet Gastroenterol. Hepatol. 2017, 2, 337–346. [Google Scholar] [CrossRef]
- Gunturu, K.S.; Yao, X.; Cong, X.; Thumar, J.R.; Hochster, H.S.; Stein, S.M.; Lacy, J. FOLFIRINOX for Locally Advanced and Metastatic Pancreatic Cancer: Single Institution Retrospective Review of Efficacy and Toxicity. Med. Oncol. 2013, 30, 361. [Google Scholar] [CrossRef]
- Guo, X.; Xiong, L.; Yu, L.; Li, R.; Wang, Z.; Ren, B.; Dong, J.; Li, B.; Wang, D. Increased Level of Nucleolin Confers to Aggressive Tumor Progression and Poor Prognosis in Patients with Hepatocellular Carcinoma after Hepatectomy. Diagn. Pathol. 2014, 9, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcel, V.; Catez, F.; Berger, C.M.; Perrial, E.; Plesa, A.; Thomas, X.; Mattei, E.; Hayette, S.; Saintigny, P.; Bouvet, P.; et al. Expression Profiling of Ribosome Biogenesis Factors Reveals Nucleolin as a Novel Potential Marker to Predict Outcome in AML Patients. PLoS ONE 2017, 12, e0170160. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Huang, Y.; Xue, C.; Chen, Y.; Hou, X.; Guo, Y.; Zhao, L.; Hu, Z.H.; Huang, Y.; Luo, Y.; et al. Prognostic Significance of the Combined Score of Endothelial Expression of Nucleolin and CD31 in Surgically Resected Non-Small Cell Lung Cancer. PLoS ONE 2013, 8, e54674. [Google Scholar] [CrossRef] [Green Version]
- Qiu, W.; Zhou, F.; Zhang, Q.; Sun, X.; Shi, X.; Liang, Y.; Wang, X.; Yue, L. Overexpression of Nucleolin and Different Expression Sites Both Related to the Prognosis of Gastric Cancer. APMIS 2013, 121, 919–925. [Google Scholar] [CrossRef]
- Gilles, M.-E.; Maione, F.; Cossutta, M.; Carpentier, G.; Caruana, L.; Di Maria, S.; Houppe, C.; Destouches, D.; Shchors, K.; Prochasson, C.; et al. Nucleolin Targeting Impairs the Progression of Pancreatic Cancer and Promotes the Normalization of Tumor Vasculature. Cancer Res. 2016, 76, 7181–7193. [Google Scholar] [CrossRef] [Green Version]
- Ugrinova, I.; Petrova, M.; Chalabi-Dchar, M.; Bouvet, P. Multifaceted Nucleolin Protein and Its Molecular Partners in Oncogenesis. In Advances in Protein Chemistry and Structural Biology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 111, pp. 133–164. ISBN 978-0-12-814342-1. [Google Scholar]
- Hovanessian, A.G.; Puvion-Dutilleul, F.; Nisole, S.; Svab, J.; Perret, E.; Deng, J.-S.; Krust, B. The Cell-Surface-Expressed Nucleolin Is Associated with the Actin Cytoskeleton. Exp. Cell Res. 2000, 261, 312–328. [Google Scholar] [CrossRef]
- Koutsioumpa, M.; Papadimitriou, E. Cell Surface Nucleolin as a Target for Anti-Cancer Therapies. Recent Pat. Anticancer Drug Discov. 2014, 9, 137–152. [Google Scholar] [CrossRef]
- Farin, K.; Schokoroy, S.; Haklai, R.; Cohen-Or, I.; Elad-Sfadia, G.; Reyes-Reyes, M.E.; Bates, P.J.; Cox, A.D.; Kloog, Y.; Pinkas-Kramarski, R. Oncogenic Synergism between ErbB1, Nucleolin, and Mutant Ras. Cancer Res. 2011, 71, 2140–2151. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Gomez, E.C.; Chalabi-Dchar, M.; Rong, C.; Das, S.; Ugrinova, I.; Gaume, X.; Monier, K.; Mongelard, F.; Bouvet, P. Integrated Analysis of MRNA and MiRNA Expression in HeLa Cells Expressing Low Levels of Nucleolin. Sci. Rep. 2017, 7, 9017. [Google Scholar] [CrossRef] [Green Version]
- Krust, B.; El Khoury, D.; Nondier, I.; Soundaramourty, C.; Hovanessian, A.G. Targeting Surface Nucleolin with Multivalent HB-19 and Related Nucant Pseudopeptides Results in Distinct Inhibitory Mechanisms Depending on the Malignant Tumor Cell Type. BMC Cancer 2011, 11, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Destouches, D.; El Khoury, D.; Hamma-Kourbali, Y.; Krust, B.; Albanese, P.; Katsoris, P.; Guichard, G.; Briand, J.P.; Courty, J.; Hovanessian, A.G. Suppression of Tumor Growth and Angiogenesis by a Specific Antagonist of the Cell-Surface Expressed Nucleolin. PLoS ONE 2008, 3, e2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Destouches, D.; Page, N.; Hamma-Kourbali, Y.; Machi, V.; Chaloin, O.; Frechault, S.; Birmpas, C.; Katsoris, P.; Beyrath, J.; Albanese, P.; et al. A Simple Approach to Cancer Therapy Afforded by Multivalent Pseudopeptides That Target Cell-Surface Nucleoproteins. Cancer Res. 2011, 71, 3296–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birmpas, C.; Briand, J.P.; Courty, J.; Katsoris, P. Nucleolin Mediates the Antiangiogenesis Effect of the Pseudopeptide N6L. BMC Cell Biol. 2012, 13, 32. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, E.; Antonosante, A.; d’Angelo, M.; Cristiano, L.; Galzio, R.; Destouches, D.; Florio, T.M.; Dhez, A.C.; Astarita, C.; Cinque, B.; et al. Nucleolin Antagonist Triggers Autophagic Cell Death in Human Glioblastoma Primary Cells and Decreased in Vivo Tumor Growth in Orthotopic Brain Tumor Model. Oncotarget 2015, 6, 42091. [Google Scholar] [CrossRef] [Green Version]
- Ramos, K.S.; Moore, S.; Runge, I.; Tavera-Garcia, M.A.; Cascone, I.; Courty, J.; Reyes-Reyes, E.M. The Nucleolin Antagonist N6L Inhibits LINE1 Retrotransposon Activity in Non-Small Cell Lung Carcinoma Cells. J. Cancer 2020, 11, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Sanhaji, M.; Göring, J.; Couleaud, P.; Aires, A.; Cortajarena, A.L.; Courty, J.; Prina-Mello, A.; Stapf, M.; Ludwig, R.; Volkov, Y.; et al. The Phenotype of Target Pancreatic Cancer Cells Influences Cell Death by Magnetic Hyperthermia with Nanoparticles Carrying Gemicitabine and the Pseudo-Peptide NucAnt. Nanomed. Nanotechnol. Biol. Med. 2019, 20, 101983. [Google Scholar] [CrossRef]
- Boj, S.F.; Hwang, C.-I.; Baker, L.A.; Chio, I.I.C.; Engle, D.D.; Corbo, V.; Jager, M.; Ponz-Sarvise, M.; Tiriac, H.; Spector, M.S.; et al. Organoid Models of Human and Mouse Ductal Pancreatic Cancer. Cell 2015, 160, 324–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Veroli, G.Y.; Fornari, C.; Wang, D.; Mollard, S.; Bramhall, J.L.; Richards, F.M.; Jodrell, D.I. Combenefit: An Interactive Platform for the Analysis and Visualization of Drug Combinations. Bioinformatics 2016, 32, 2866–2868. [Google Scholar] [CrossRef] [PubMed]
- David, A.; Dolan, B.P.; Hickman, H.D.; Knowlton, J.J.; Clavarino, G.; Pierre, P.; Bennink, J.R.; Yewdell, J.W. Nuclear Translation Visualized by Ribosome-Bound Nascent Chain Puromycylation. J. Cell Biol. 2012, 197, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Bash-Imam, Z.; Thérizols, G.; Vincent, A.; Lafôrets, F.; Espinoza, M.P.; Pion, N.; Macari, F.; Pannequin, J.; David, A.; Saurin, J.-C.; et al. Translational Reprogramming of Colorectal Cancer Cells Induced by 5-Fluorouracil through a MiRNA-Dependent Mechanism. Oncotarget 2017, 8, 46219–46233. [Google Scholar] [CrossRef]
- Jourdren, L.; Bernard, M.; Dillies, M.-A.; Le Crom, S. Eoulsan: A Cloud Computing-Based Framework Facilitating High Throughput Sequencing Analyses. Bioinformatics 2012, 28, 1542–1543. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonenberg, N.; Hinnebusch, A.G. Regulation of Translation Initiation in Eukaryotes: Mechanisms and Biological Targets. Cell 2009, 136, 731–745. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Diaz, J.E.L.; Flamholz, Z.N.; Keenan, A.B.; Lachmann, A.; Wojciechowicz, M.L.; Cagan, R.L.; Ma’ayan, A. ModEnrichr: A Suite of Gene Set Enrichment Analysis Tools for Model Organisms. Nucleic Acids Res. 2019, 47, W183–W190. [Google Scholar] [CrossRef]
- Gentilella, A.; Morón-Duran, F.D.; Fuentes, P.; Zweig-Rocha, G.; Riaño-Canalias, F.; Pelletier, J.; Ruiz, M.; Turón, G.; Castaño, J.; Tauler, A.; et al. Autogenous Control of 5′TOP MRNA Stability by 40S Ribosomes. Mol. Cell 2017, 67, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Meyuhas, O.; Kahan, T. The Race to Decipher the Top Secrets of TOP MRNAs. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2015, 1849, 801–811. [Google Scholar] [CrossRef]
- Thoreen, C.C.; Chantranupong, L.; Keys, H.R.; Wang, T.; Gray, N.S.; Sabatini, D.M. A Unifying Model for MTORC1-Mediated Regulation of MRNA Translation. Nature 2012, 485, 109–113. [Google Scholar] [CrossRef]
- Aoki, K.; Adachi, S.; Homoto, M.; Kusano, H.; Koike, K.; Natsume, T. LARP1 Specifically Recognizes the 3′ Terminus of Poly(A) MRNA. FEBS Lett. 2013, 587, 2173–2178. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, B.D.; Lahr, R.M.; Damgaard, C.K.; Alain, T.; Berman, A.J. LARP1 on TOP of Ribosome Production. Wiley Interdiscip. Rev. RNA 2018, 9, e1480. [Google Scholar] [CrossRef]
- Pike, K.G.; Malagu, K.; Hummersone, M.G.; Menear, K.A.; Duggan, H.M.E.; Gomez, S.; Martin, N.M.B.; Ruston, L.; Pass, S.L.; Pass, M. Optimization of Potent and Selective Dual MTORC1 and MTORC2 Inhibitors: The Discovery of AZD8055 and AZD2014. Bioorg. Med. Chem. Lett. 2013, 23, 1212–1216. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, A.C.; Liu, Y.; Edlind, M.P.; Ingolia, N.T.; Janes, M.R.; Sher, A.; Shi, E.Y.; Stumpf, C.R.; Christensen, C.; Bonham, M.J.; et al. The Translational Landscape of MTOR Signalling Steers Cancer Initiation and Metastasis. Nature 2012, 485, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ware, M.J.; Colbert, K.; Keshishian, V.; Ho, J.; Corr, S.J.; Curley, S.A.; Godin, B. Generation of Homogenous Three-Dimensional Pancreatic Cancer Cell Spheroids Using an Improved Hanging Drop Technique. Tissue Eng. Part C Methods 2016, 22, 312–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiriac, H.; Belleau, P.; Engle, D.D.; Plenker, D.; Deschênes, A.; Somerville, T.D.D.; Froeling, F.E.M.; Burkhart, R.A.; Denroche, R.E.; Jang, G.-H.; et al. Organoid Profiling Identifies Common Responders to Chemotherapy in Pancreatic Cancer. Cancer Discov. 2018, 8, 1112–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, R.; Suzuki, Y.; Takeuchi, N.; Wakaguri, H.; Ueda, T.; Sugano, S.; Nakai, K. Comprehensive Detection of Human Terminal Oligo-Pyrimidine (TOP) Genes and Analysis of Their Characteristics. Nucleic Acids Res. 2008, 36, 3707–3715. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.; Avni, D.; Hariharan, N.; Perry, R.P.; Meyuhas, O. Oligopyrimidine Tract at the 5′ End of Mammalian Ribosomal Protein MRNAs Is Required for Their Translational Control. Proc. Natl. Acad. Sci. USA 1991, 88, 3319–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabatini, D.M. MTOR and Cancer: Insights into a Complex Relationship. Nat. Rev. Cancer 2006, 6, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Philippe, L.; van den Elzen, A.M.G.; Watson, M.J.; Thoreen, C.C. Global Analysis of LARP1 Translation Targets Reveals Tunable and Dynamic Features of 5′TOP Motifs. Proc. Natl. Acad. Sci. USA 2020, 117, 5319–5328. [Google Scholar] [CrossRef]
- Patursky-Polischuk, I.; Kasir, J.; Miloslavski, R.; Hayouka, Z.; Hausner-Hanochi, M.; Stolovich-Rain, M.; Tsukerman, P.; Biton, M.; Mudhasani, R.; Jones, S.N.; et al. Reassessment of the Role of TSC, MTORC1 and MicroRNAs in Amino Acids-Meditated Translational Control of TOP MRNAs. PLoS ONE 2014, 9, e109410. [Google Scholar] [CrossRef] [Green Version]
- Al-Ashtal, H.A.; Rubottom, C.M.; Leeper, T.C.; Berman, A.J. The LARP1 La-Module Recognizes Both Ends of TOP MRNAs. RNA Biol. 2021, 18, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.; Freeberg, M.A.; Han, T.; Kamath, A.; Yao, Y.; Fukuda, T.; Suzuki, T.; Kim, J.K.; Inoki, K. LARP1 Functions as a Molecular Switch for MTORC1-Mediated Translation of an Essential Class of MRNAs. eLife 2017, 6, e25237. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Li, X.; Zhang, J. MTOR Signaling in Cancer and MTOR Inhibitors in Solid Tumor Targeting Therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef] [Green Version]
- Bellizzi, A.M.; Bloomston, M.; Zhou, X.-P.; Iwenofu, O.H.; Frankel, W.L. The MTOR Pathway Is Frequently Activated in Pancreatic Ductal Adenocarcinoma and Chronic Pancreatitis. Appl. Immunohistochem. Mol. Morphol. 2010, 18, 442–447. [Google Scholar] [CrossRef]
- Hassan, Z.; Schneeweis, C.; Wirth, M.; Veltkamp, C.; Dantes, Z.; Feuerecker, B.; Ceyhan, G.O.; Knauer, S.K.; Weichert, W.; Schmid, R.M.; et al. MTOR Inhibitor-Based Combination Therapies for Pancreatic Cancer. Br. J. Cancer 2018, 118, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Guri, Y.; Hall, M.N. MTOR Signaling Confers Resistance to Targeted Cancer Drugs. Trends Cancer 2016, 2, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Clinical Trials. Available online: https://clinicaltrials.gov/ct2/results?cond=mTOR+inhibitors&term=&cntry=&state=&city=&dist= (accessed on 15 September 2021).






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chalabi-Dchar, M.; Cruz, E.; Mertani, H.C.; Diaz, J.-J.; Courty, J.; Cascone, I.; Bouvet, P. Nucleolin Aptamer N6L Reprograms the Translational Machinery and Acts Synergistically with mTORi to Inhibit Pancreatic Cancer Proliferation. Cancers 2021, 13, 4957. https://doi.org/10.3390/cancers13194957
Chalabi-Dchar M, Cruz E, Mertani HC, Diaz J-J, Courty J, Cascone I, Bouvet P. Nucleolin Aptamer N6L Reprograms the Translational Machinery and Acts Synergistically with mTORi to Inhibit Pancreatic Cancer Proliferation. Cancers. 2021; 13(19):4957. https://doi.org/10.3390/cancers13194957
Chicago/Turabian StyleChalabi-Dchar, Mounira, Elisabeth Cruz, Hichem C. Mertani, Jean-Jacques Diaz, José Courty, Ilaria Cascone, and Philippe Bouvet. 2021. "Nucleolin Aptamer N6L Reprograms the Translational Machinery and Acts Synergistically with mTORi to Inhibit Pancreatic Cancer Proliferation" Cancers 13, no. 19: 4957. https://doi.org/10.3390/cancers13194957