
Novel Protein-Based Vaccine against Self-Antigen Reduces the Formation of Sporadic Colon Adenomas in Mice

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Colonoscopy
2.3. Vaccines
2.4. Animal Treatment
2.5. ELISA Assay for Humoral Response
2.6. ELISpot Assay
2.7. Histopathology
2.8. Immunohistochemistry
2.9. Statistical Analysis
3. Results
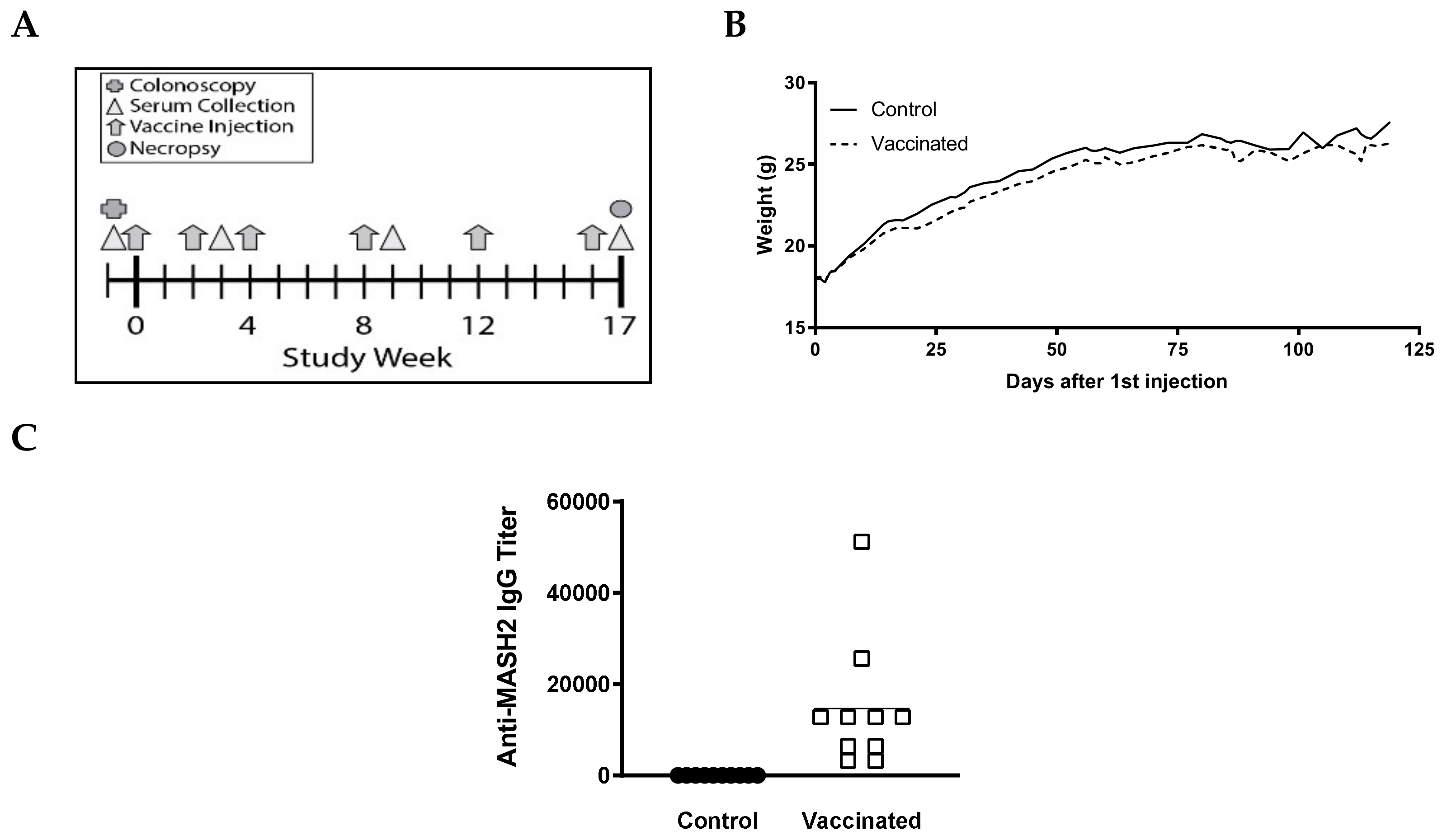
3.1. KISIMA-Mash2 Vaccine Promotes T Cell and Humoral Responses
3.2. KISIMA-Mash2 Vaccine Is Safe and Promotes an Antibody Response in Apc+/Min-FCCC Mice
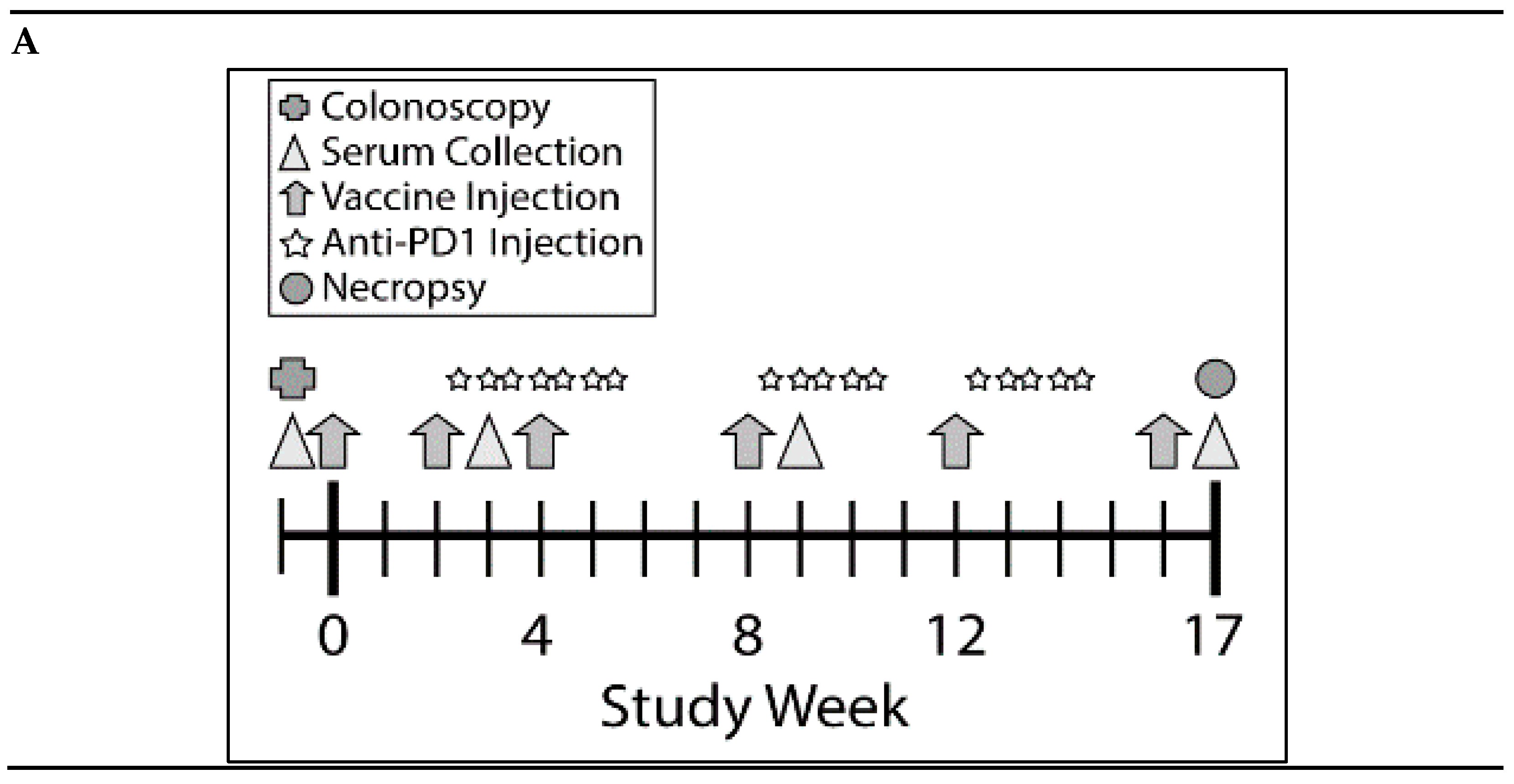
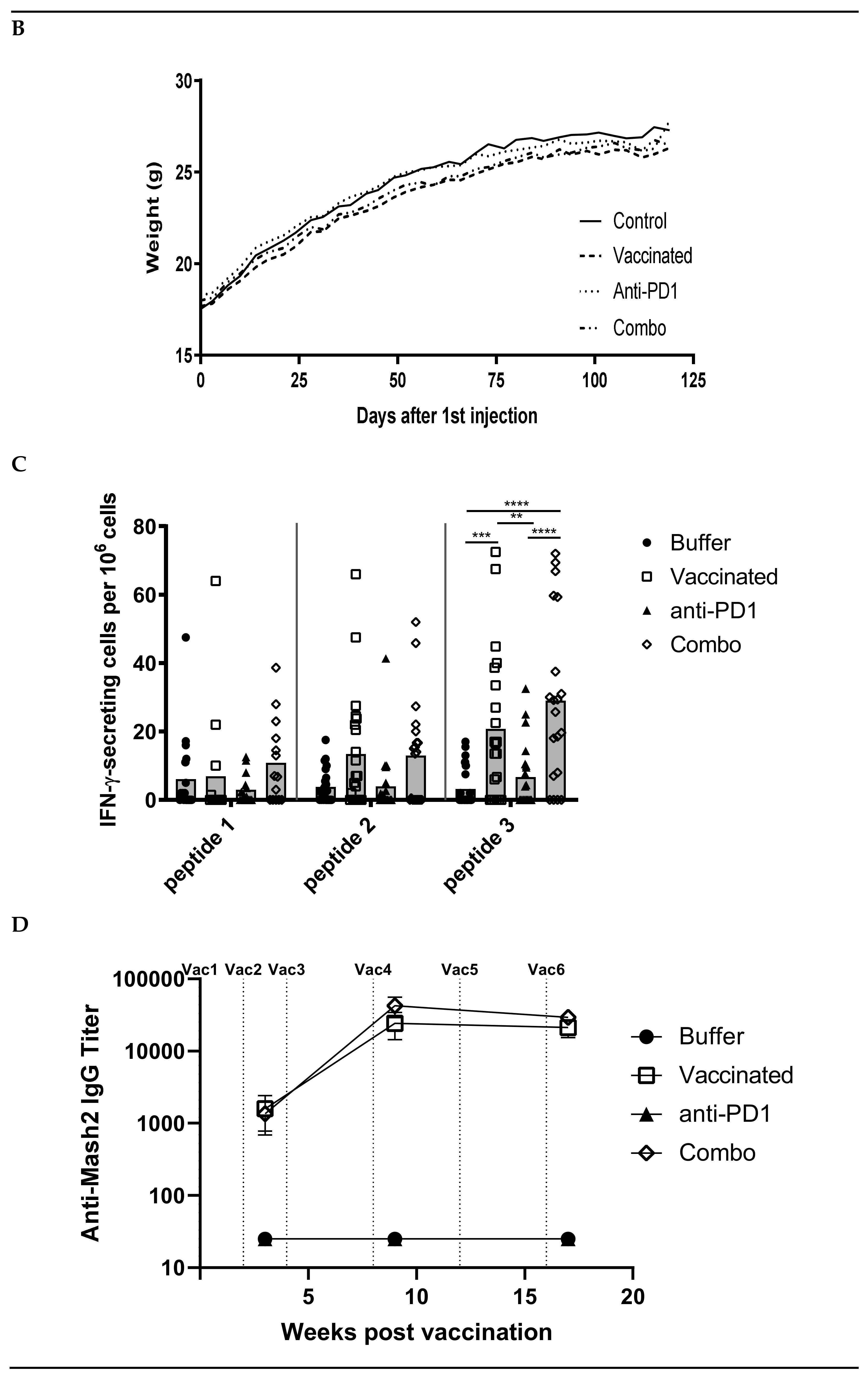
3.3. Combination of KISIMA-Mash2 Vaccine and Anti-PD-1 Treatment Is Safe and Promotes T Cell and Antibody Response in Apc+/Min-FCCC Mice
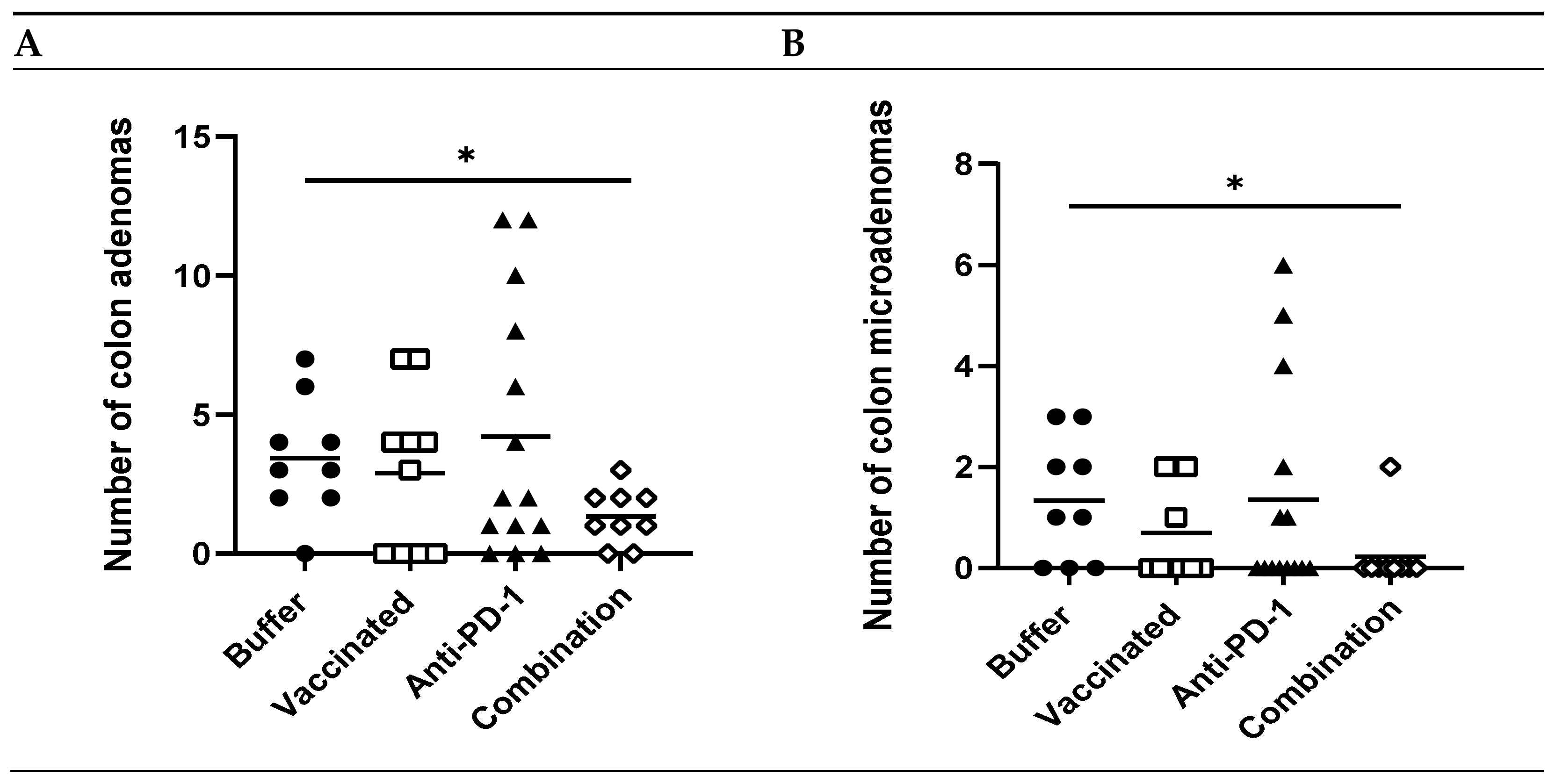
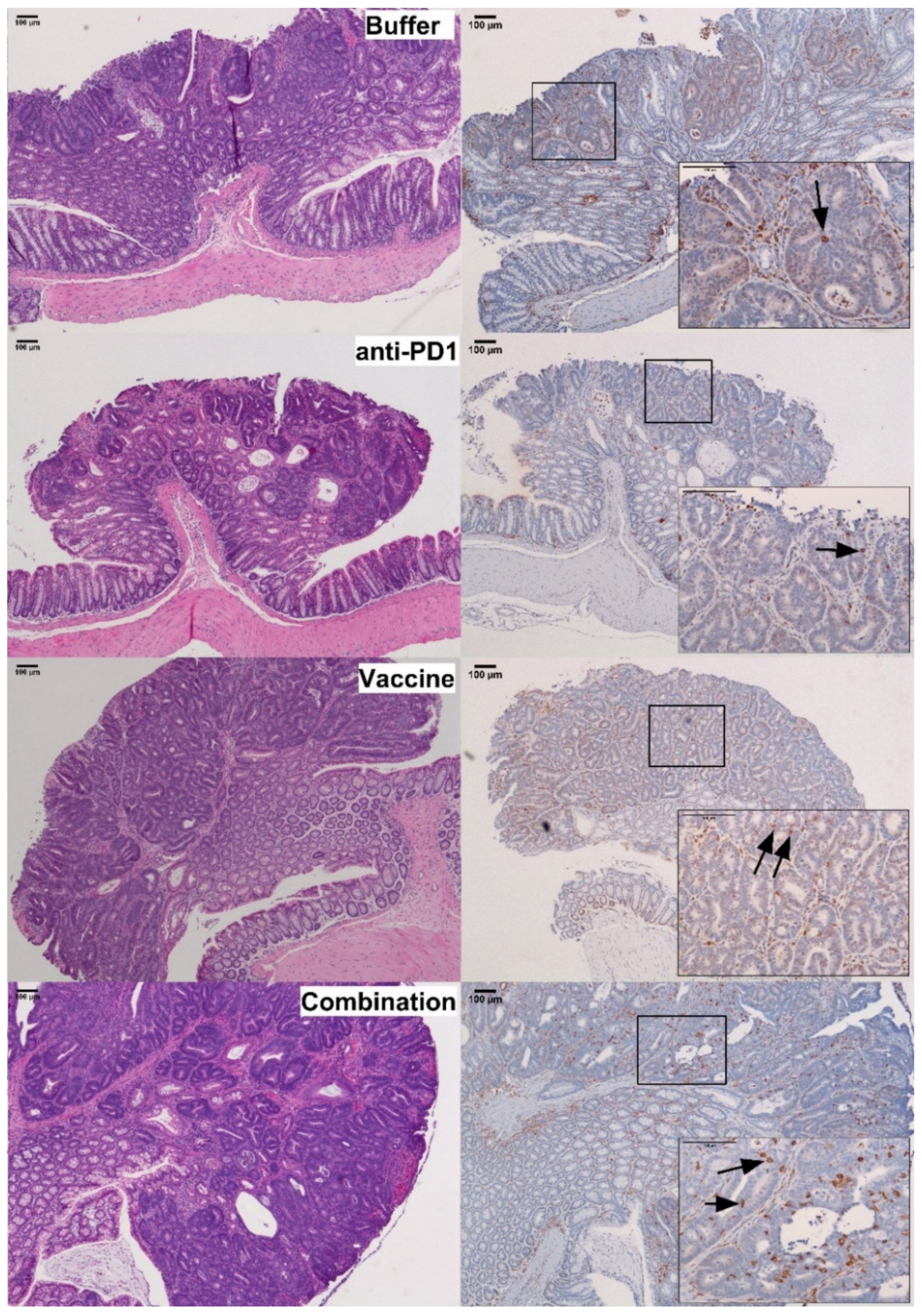
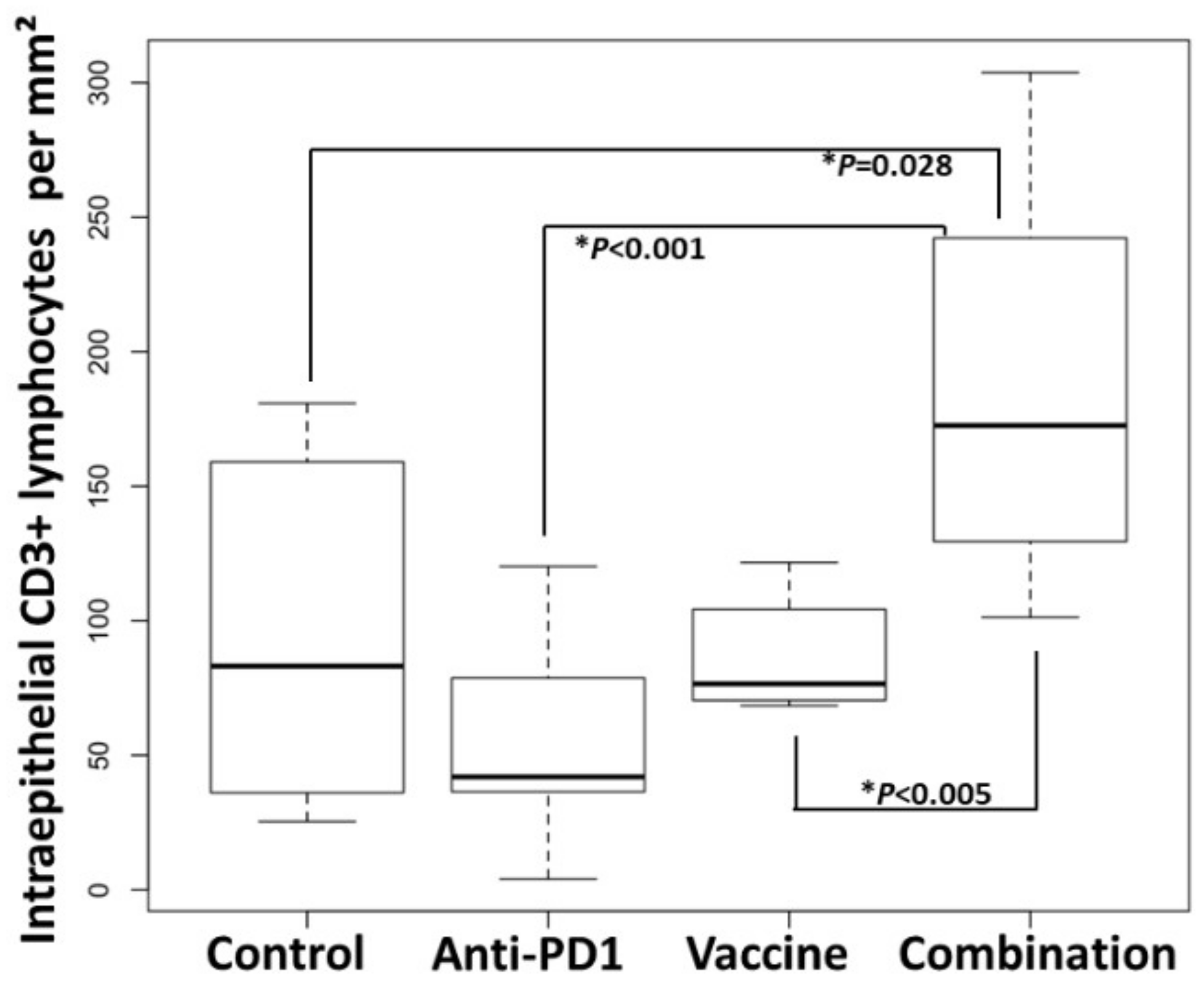
3.4. Combination of KISIMA-Mash2 Vaccine and Anti-PD-1 Treatment Reduces the Formation of Colon Tumors in Apc+/Min-FCCC Mice and Is Associated with Increased T-cell Infiltration into the Intraepithelial Compartment of Colon Adenomas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Wools, A.; Dapper, E.; De Leeuw, J. Colorectal cancer screening participation: A systematic review. Eur. J. Public Health 2015, 26, 158–168. [Google Scholar] [CrossRef]
- Wolf, A.M.D.; Fontham, E.T.; Church, T.R.; Flowers, C.R.; Guerra, C.E.; LaMonte, S.J.; Etzioni, R.; McKenna, M.T.; Oeffinger, K.C.; Shih, Y.-C.T.; et al. Colorectal cancer screening for average-risk adults: 2018 guideline update from the American Cancer Society. CA Cancer J. Clin. 2018, 68, 250–281. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Fedewa, S.A.; Anderson, W.F.; Miller, K.D.; Ma, J.; Rosenberg, P.S.; Jemal, A. Colorectal cancer incidence patterns in the United States, 1974–2013. J. Natl. Cancer Inst. 2017, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasperson, K.W.; Tuohy, T.M.; Neklason, D.W.; Burt, R.W. Hereditary and familial colon cancer. Gastroenterology 2010, 138, 2044–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, S.D.; McMurray, J.J.; Pfeffer, M.A.; Wittes, J.; Fowler, R.; Finn, P.; Anderson, W.F.; Zauber, A.; Hawk, E.; Bertagnolli, M. Cardiovascular risk associated with celecoxib in a clinical trial for colorectal adenoma Prevention. N. Engl. J. Med. 2005, 352, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Coyle, C.; Cafferty, F.H.; Langley, R.E. Aspirin and colorectal cancer prevention and treatment: Is it for everyone? Curr. Color Cancer Rep. 2016, 12, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Finn, O.J. The dawn of vaccines for cancer prevention. Nat. Rev. Immunol. 2018, 18, 183–194. [Google Scholar] [CrossRef]
- Palladini, A.; Landuzzi, L.; Lollini, P.-L.; Nanni, P. Cancer immunoprevention: From mice to early clinical trials. BMC Immunol. 2018, 19, 1–6. [Google Scholar] [CrossRef]
- Kimura, T.; McKolanis, J.R.; Dzubinski, L.A.; Islam, K.; Potter, D.M.; Salazar, A.M.; Schoen, R.E.; Finn, O.J. MUC1 Vaccine for individuals with advanced adenoma of the colon: A cancer immunoprevention feasibility study. Cancer Prev. Res. 2013, 6, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Markowitz, S.D.; Bertagnolli, M.M. Molecular basis of colorectal cancer. N. Engl. J. Med. 2009, 361, 2449–2460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuijers, J.; Junker, J.P.; Mokry, M.; Hatzis, P.; Koo, B.-K.; Sasselli, V.; Van Der Flier, L.G.; Cuppen, E.; Van Oudenaarden, A.; Clevers, H. Ascl2 Acts as an R-spondin/Wnt-responsive switch to control stemness in intestinal crypts. Cell Stem Cell 2015, 16, 158–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jubb, A.M.; Chalasani, S.; Frantz, G.D.; Smits, R.; Grabsch, H.I.; Kavi, V.; Maughan, N.J.; Hillan, K.J.; Quirke, P.; Koeppen, H. Achaete-scute like 2 (ascl2) is a target of Wnt signalling and is upregulated in intestinal neoplasia. Oncogene 2006, 25, 3445–3457. [Google Scholar] [CrossRef] [Green Version]
- Broussard, E.K.; Kim, R.; Wiley, J.C.; Marquez, J.P.; Annis, J.E.; Pritchard, D.; Disis, M.L. Identification of Putative immunologic targets for colon cancer prevention based on conserved gene upregulation from preinvasive to malignant lesions. Cancer Prev. Res. 2013, 6, 666–674. [Google Scholar] [CrossRef] [Green Version]
- Jang, B.G.; Kim, H.S.; Kim, K.J.; Rhee, Y.-Y.; Kim, W.H.; Kang, G.H. Distribution of intestinal stem cell markers in colorectal precancerous lesions. Histopathology 2015, 68, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Rioux, C.R.; Clapper, M.L.; Cooper, H.S.; Michaud, J.; Amant, N.S.; Koohsari, H.; Workman, L.; Kaunga, E.; Hensley, H.; Pilorget, A.; et al. Self-antigen MASH2 combined with the AS15 immunostimulant induces tumor protection in colorectal cancer mouse models. PLoS ONE 2019, 14, e0210261. [Google Scholar] [CrossRef]
- Belnoue, E.; Mayol, J.-F.; Carboni, S.; Besson, W.D.B.; Dupuychaffray, E.; Nelde, A.; Stevanovic, S.; Santiago-Raber, M.-L.; Walker, P.R.; Derouazi, M. Targeting self- and neoepitopes with a modular self-adjuvanting cancer vaccine. JCI Insight 2019, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Derouazi, M.S.; Di Berardino-Besson, W.; Belnoue, E.; Hoepner, S.; Walther, R.; Benkhoucha, M.; Teta, P.; Dufour, Y.J.; Maroun, C.Y.; Salazar, A.M.; et al. Novel cell-penetrating peptide-based vaccine induces robust CD4+ and CD8+ T cell–mediated antitumor immunity. Cancer Res. 2015, 75, 3020–3031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belnoue, E.; Di Berardino-Besson, W.; Gaertner, H.; Carboni, S.; Dunand-Sauthier, I.; Cerini, F.; Suso-Inderberg, E.-M.; Wälchli, S.; König, S.; Salazar, A.M.; et al. Enhancing Antitumor immune responses by optimized combinations of cell-penetrating peptide-based vaccines and adjuvants. Mol. Ther. 2016, 24, 1675–1685. [Google Scholar] [CrossRef] [Green Version]
- Cooper, H.S.; Chang, W.-C.L.; Coudry, R.; Gary, M.A.; Everley, L.; Spittle, C.S.; Wang, H.; Litwin, S.; Clapper, M.L. Generation of a unique strain of multiple intestinal neoplasia (Apc+/Min-FCCC) mice with significantly increased numbers of colorectal adenomas. Mol. Carcinog. 2005, 44, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Hensley, H.H.; Merkel, C.E.; Chang, W.-C.L.; Devarajan, K.; Cooper, H.S.; Clapper, M.L. Endoscopic imaging and size estimation of colorectal adenomas in the multiple intestinal neoplasia mouse. Gastrointest. Endosc. 2009, 69, 742–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.; Taggart, M.W.; Reyes-Uribe, L.; Borras, E.; Riquelme, E.; Barnett, R.M.; Leoni, G.; Lucas, F.A.S.; Catanese, M.T.; Mori, F.; et al. Immune profiling of premalignant lesions in patients with lynch syndrome. JAMA Oncol. 2018, 4, 1085–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Riddell, R.H.; Goldman, H.; Ransohoff, D.F.; Appelman, H.D.; Fenoglio, C.M.; Haggitt, R.C.; Hren, C.; Correa, P.; Hamilton, S.R.; Morson, B.C.; et al. Dysplasia in inflammatory bowel disease: Standardized classification with provisional clinical applications. Hum. Pathol. 1983, 14, 931–968. [Google Scholar] [CrossRef]
- Odze, R.D.; Goldblum, J.R.; Crawford, J.M.; Renshaw, A. Surgical pathology of the gi tract, liver, biliary tree, and pancreas. Adv. Anat. Pathol. 2005, 12, 35. [Google Scholar] [CrossRef]
- Hendry, S.; Salgado, R.; Gevaert, T.; Russell, P.A.; John, T.; Thapa, B.; Christie, M.; van de Vijver, K.; Estrada, M.; Gonzalez-Ericsson, P.I.; et al. Assessing Tumor-infiltrating lymphocytes in solid tumors: A practical review for pathologists and proposal for a standardized method from the international immuno-oncology biomarkers working group: Part 2: TILs in melanoma, gastrointestinal tract carcinomas, non–small cell lung carcinoma and mesothelioma, endometrial and ovarian carcinomas, squamous cell carcinoma of the head and neck, genitourinary carcinomas, and primary brain tumors. Adv. Anat. Pathol. 2017, 24, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Belnoue, E.; Leystra, A.A.; Clapper, M.L.; Derouazi, M. KISIMA-Mash2 Vaccine Reduces the Number of Gross Colon Tumors in Male Apc+/Min-FCCC Mice; Material Not Intended for Publication; AMAL Therapeutics: Geneva, Switzerland; Fox Chase Cancer Center: Philadelphia, PA, USA, 2021. [Google Scholar]
- Sahin, U.; Oehm, P.; Derhovanessian, E.; Jabulowsky, R.A.; Vormehr, M.; Gold, M.; Maurus, D.; Schwarck-Kokarakis, D.; Kuhn, A.N.; Omokoko, T.; et al. An RNA vaccine drives immunity in checkpoint-inhibitor-treated melanoma. Nature 2020, 585, 107–112. [Google Scholar] [CrossRef]
- Kallert, S.M.; Darbre, S.; Bonilla, W.V.; Kreutzfeldt, M.; Page, N.; Müller, P.; Kreuzaler, M.; Lu, M.; Favre, S.; Kreppel, F.; et al. Replicating viral vector platform exploits alarmin signals for potent CD8+ T cell-mediated tumour immunotherapy. Nat. Commun. 2017, 8, 15327. [Google Scholar] [CrossRef]
- Zemp, F.; Rajwani, J.; Mahoney, D.J. Rhabdoviruses as vaccine platforms for infectious disease and cancer. Biotechnol. Genet. Eng. Rev. 2018, 34, 122–138. [Google Scholar] [CrossRef]
- Snook, A.E.; Baybutt, T.R.; Xiang, B.; Abraham, T.S.; Flickinger, J.C.; Hyslop, T.; Zhan, T.; Kraft, W.K.; Sato, T.; Waldman, S.A. Split tolerance permits safe Ad5-GUCY2C-PADRE vaccine-induced T-cell responses in colon cancer patients. J. Immunother. Cancer 2019, 7, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuhara, H.; Ino, Y.; Todo, T. Oncolytic virus therapy: A new era of cancer treatment at dawn. Cancer Sci. 2016, 107, 1373–1379. [Google Scholar] [CrossRef]
- Lichty, B.D.; Breitbach, C.J.; Stojdl, D.F.; Bell, J.C. Going viral with cancer immunotherapy. Nat. Rev. Cancer 2014, 14, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Bissa, M.; Illiano, E.; Pacchioni, S.; Paolini, F.; Zanotto, C.; Morghen, C.D.G.; Massa, S.; Franconi, R.; Radaelli, A.; Venuti, A. A prime/boost strategy using DNA/fowlpox recombinants expressing the genetically attenuated E6 protein as a putative vaccine against HPV-16-associated cancers. J. Transl. Med. 2015, 13, 80. [Google Scholar] [CrossRef] [Green Version]
- Swadling, L.; Capone, S.; Antrobus, R.D.; Brown, A.; Richardson, R.; Newell, E.W.; Halliday, J.; Kelly, C.; Bowen, D.; Fergusson, J.; et al. A human vaccine strategy based on chimpanzee adenoviral and MVA vectors that primes, boosts, and sustains functional HCV-specific T cell memory. Sci. Transl. Med. 2014, 6, 261ra153. [Google Scholar] [CrossRef] [Green Version]
- Aitken, A.S.; Roy, D.G.; Martin, N.T.; Sad, S.; Bell, J.C.; Bourgeois-Daigneault, M.-C. Brief communication; A heterologous oncolytic bacteria-virus prime-boost approach for anticancer vaccination in mice. J. Immunother. 2018, 41, 125–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, L.; Schrump, D.S.; Gildersleeve, J.C. Whole-cell cancer vaccines induce large antibody responses to carbohydrates and glycoproteins. Cell Chem. Biol. 2016, 23, 1515–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atanackovic, D.; Altorki, N.K.; Stockert, E.; Williamson, B.; Jungbluth, A.A.; Ritter, E.; Santiago, D.; Ferrara, C.A.; Matsuo, M.; Selvakumar, A.; et al. Vaccine-Induced CD4+T Cell Responses to MAGE-3 Protein in lung cancer patients. J. Immunol. 2004, 172, 3289–3296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, D.Y.; Kwek, S.S.; Raju, S.S.; Li, T.; McCarthy, E.; Chow, E.; Aran, D.; Ilano, A.; Pai, C.-C.S.; Rancan, C.; et al. intratumoral CD4+ T cells mediate anti-tumor cytotoxicity in human bladder cancer. Cell 2020, 181, 1612–1625. [Google Scholar] [CrossRef] [PubMed]
- Eyvazi, S.; Farajnia, S.; Dastmalchi, S.; Kanipour, F.; Zarredar, H.; Bandehpour, M.; Bandehpour, M. antibody based EpCAM targeted therapy of cancer, review and update. Curr. Cancer Drug Targets 2018, 18, 857–868. [Google Scholar] [CrossRef]
- Zheng, C.; Feng, J.; Lu, D.; Wang, P.; Xing, S.; Coll, J.-L.; Yang, D.; Yan, X. A novel anti-CEACAM5 monoclonal antibody, CC4, suppresses colorectal tumor growth and enhances NK Cells-mediated tumor immunity. PLoS ONE 2011, 6, e21146. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.-H.; Rao, J.; Yang, J.; Wu, F.; Tan, J.; Xu, S.-L.; Ding, Y.; Zhan, N.; Hu, X.-G.; Cui, Y.-H.; et al. SEMA3F prevents metastasis of colorectal cancer by PI3K-AKT-dependent down-regulation of the ASCL2-CXCR4 axis. J. Pathol. 2015, 236, 467–478. [Google Scholar] [CrossRef]
- Agudo, J.; Park, E.S.; Rose, S.A.; Alibo, E.; Sweeney, R.; Dhainaut, M.; Kobayashi, K.S.; Sachidanandam, R.; Baccarini, A.; Merad, M.; et al. Quiescent tissue stem cells evade immune surveillance. Immunity 2018, 48, 271–285.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metcalfe, C.; Kljavin, N.M.; Ybarra, R.; De Sauvage, F.J. Lgr5+ Stem cells are indispensable for radiation-induced intestinal regeneration. Cell Stem Cell 2014, 14, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.C.; Guerra, G.R.; Pham, T.; Mitchell, C.; Lynch, A.C.; Warrier, S.K.; Ramsay, R.G.; Heriot, A.G. Prognostic impact of tumor-infiltrating lymphocytes in primary and metastatic colorectal cancer: A systematic review and meta-analysis. Dis. Colon Rectum 2019, 62, 498–508. [Google Scholar] [CrossRef]
- Galon, J.; Costes, A.; Sanchez-Cabo, F.; Kirilovsky, A.; Mlecnik, B.; Lagorce-Pagès, C.; Tosolini, M.; Camus, M.; Berger, A.; Wind, P.; et al. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science 2006, 313, 1960–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagès, F.; Mlecnik, B.; Marliot, F.; Bindea, G.; Ou, F.-S.; Bifulco, C.; Lugli, A.; Zlobec, I.; Rau, T.T.; Berger, M.D.; et al. International validation of the consensus Immunoscore for the classification of colon cancer: A prognostic and accuracy study. Lancet 2018, 391, 2128–2139. [Google Scholar] [CrossRef]
- Xiao, Q.; Wu, J.; Wang, W.-J.; Chen, S.; Zheng, Y.; Yu, X.; Meeth, K.; Sahraei, M.; Bothwell, A.L.M.; Chen, L.; et al. DKK2 imparts tumor immunity evasion through β-catenin-independent suppression of cytotoxic immune-cell activation. Nat. Med. 2018, 24, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, D.; Lyerly, H.K.; Morse, M.A. Deficient mismatch repair and the role of immunotherapy in metastatic colorectal cancer. Curr. Treat. Options Oncol. 2016, 17, 1–16. [Google Scholar] [CrossRef]
- Yarchoan, M.; Huang, C.; Zhu, Q.; Ferguson, A.K.; Durham, J.N.; Anders, R.A.; Thompson, E.D.; Rozich, N.S.; Ii, D.L.T.; Nauroth, J.M.; et al. A phase 2 study of GVAX colon vaccine with cyclophosphamide and pembrolizumab in patients with mismatch repair proficient advanced colorectal cancer. Cancer Med. 2019, 9, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.; Pereira, L.; Roth, S.; Galletta, L.; Link, E.; Akhurst, T.; Solomon, B.; Michael, M.; Darcy, P.; Sampurno, S.; et al. First-in-human phase I clinical trial of a combined immune modulatory approach using TetMYB vaccine and Anti-PD-1 antibody in patients with advanced solid cancer including colorectal or adenoid cystic carcinoma: The MYPHISMO study protocol (NCT03287427). Contemp. Clin. Trials Commun. 2019, 16, 100409. [Google Scholar] [CrossRef]
- Moehler, M.; Delic, M.; Goepfert, K.; Aust, D.; Grabsch, H.I.; Halama, N.; Heinrich, B.; Julié, C.; Lordick, F.; Lutz, M.P.; et al. Immunotherapy in gastrointestinal cancer: Recent results, current studies and future perspectives. Eur. J. Cancer 2016, 59, 160–170. [Google Scholar] [CrossRef]
- Clapper, M.L.; Chang, W.-C.L.; Cooper, H.S. Dysplastic aberrant crypt foci: Biomarkers of early colorectal neoplasia and response to preventive intervention. Cancer Prev. Res. 2020, 13, 229–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahadova, A.; Gallon, R.; Gebert, J.; Ballhausen, A.; Endris, V.; Kirchner, M.; Stenzinger, A.; Burn, J.; Doeberitz, M.V.K.; Bläker, H.; et al. Three molecular pathways model colorectal carcinogenesis in Lynch syndrome. Int. J. Cancer 2018, 143, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucherlapati, M.H.; Lee, K.; Nguyen, A.A.; Clark, A.B.; Hou, H.; Rosulek, A.; Li, H.; Yang, K.; Fan, K.; Lipkin, M.; et al. An Msh2 conditional knockout mouse for studying intestinal cancer and testing anticancer agents. Gastroenterology 2010, 138, 993–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelincik, O.; Ibrahim, H.; Ozkan, M.; Ahadova, A.; Sei, S.; Shoemaker, R.; Kloor, M.; Doeberitz, M.V.K.; Lipkin, S.M. Abstract 2732: Frameshift neoantigen vaccination prevent Lynch syndrome mouse model intestinal cancer. Prev. Early Detect. Intercept. 2019, 79, 2732. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belnoue, E.; Leystra, A.A.; Carboni, S.; Cooper, H.S.; Macedo, R.T.; Harvey, K.N.; Colby, K.B.; Campbell, K.S.; Vanderveer, L.A.; Clapper, M.L.; et al. Novel Protein-Based Vaccine against Self-Antigen Reduces the Formation of Sporadic Colon Adenomas in Mice. Cancers 2021, 13, 845. https://doi.org/10.3390/cancers13040845
Belnoue E, Leystra AA, Carboni S, Cooper HS, Macedo RT, Harvey KN, Colby KB, Campbell KS, Vanderveer LA, Clapper ML, et al. Novel Protein-Based Vaccine against Self-Antigen Reduces the Formation of Sporadic Colon Adenomas in Mice. Cancers. 2021; 13(4):845. https://doi.org/10.3390/cancers13040845
Chicago/Turabian StyleBelnoue, Elodie, Alyssa A. Leystra, Susanna Carboni, Harry S. Cooper, Rodrigo T. Macedo, Kristen N. Harvey, Kimberly B. Colby, Kerry S. Campbell, Lisa A. Vanderveer, Margie L. Clapper, and et al. 2021. "Novel Protein-Based Vaccine against Self-Antigen Reduces the Formation of Sporadic Colon Adenomas in Mice" Cancers 13, no. 4: 845. https://doi.org/10.3390/cancers13040845
APA StyleBelnoue, E., Leystra, A. A., Carboni, S., Cooper, H. S., Macedo, R. T., Harvey, K. N., Colby, K. B., Campbell, K. S., Vanderveer, L. A., Clapper, M. L., & Derouazi, M. (2021). Novel Protein-Based Vaccine against Self-Antigen Reduces the Formation of Sporadic Colon Adenomas in Mice. Cancers, 13(4), 845. https://doi.org/10.3390/cancers13040845