
The Flt3L/Flt3 Axis in Dendritic Cell Biology and Cancer Immunotherapy



Abstract
:Simple Summary
Abstract
1. Introduction
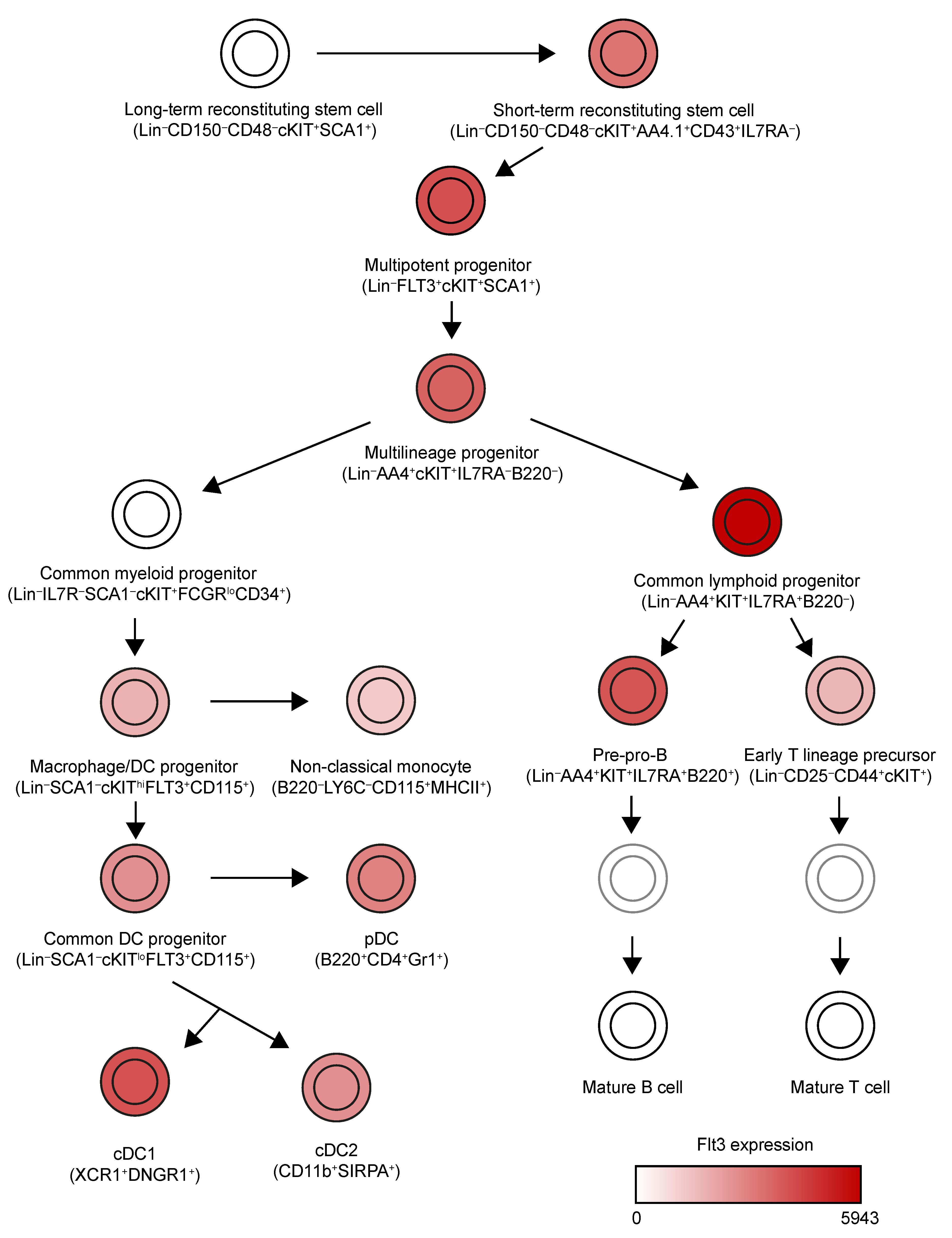
2. Expression Pattern of Flt3 and Flt3L
3. Flt3 Signaling in Response to Its Ligand
4. Expansion of DCs with Flt3L
5. Preclinical Studies Involving Flt3L
6. Flt3L in the Clinic
7. Conclusions and Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guilliams, M.; Ginhoux, F.; Jakubzick, C.; Naik, S.H.; Onai, N.; Schraml, B.U.; Segura, E.; Tussiwand, R.; Yona, S. Dendritic cells, monocytes and macrophages: A unified nomenclature based on ontogeny. Nat. Rev. Immunol. 2014, 14, 571–578. [Google Scholar] [CrossRef]
- Merad, M.; Sathe, P.; Helft, J.; Miller, J.; Mortha, A. The Dendritic Cell Lineage: Ontogeny and Function of Dendritic Cells and Their Subsets in the Steady State and the Inflamed Setting. Annu. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef] [Green Version]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Hawiger, D.; Nussenzweig, M.C. Tolerogenic Dendritic Cells. Annu. Rev. Immunol. 2003, 21, 685–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jojic, V.; Shay, T.; Sylvia, K.; Zuk, O.; Sun, X.; Kang, J.; Regev, A.; Koller, D.; Best, A.J.; Knell, J.; et al. Identification of transcriptional regulators in the mouse immune system. Nat. Immunol. 2013, 14, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Schraml, B.U.; Van Blijswijk, J.; Zelenay, S.; Whitney, P.G.; Filby, A.; Acton, S.E.; Rogers, N.C.; Moncaut, N.; Carvajal, J.J.; Reis, E.; et al. Genetic tracing via DNGR-1 expression history defines dendritic cells as a hematopoietic lineage. Cell 2013, 154, 843–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferris, S.T.; Durai, V.; Wu, R.; Theisen, D.J.; Ward, J.P.; Bern, M.D.; Davidson, J.T.; Bagadia, P.; Liu, T.; Briseño, C.G.; et al. cDC1 prime and are licensed by CD4+ T cells to induce anti-tumour immunity. Nature 2020, 584, 624–629. [Google Scholar] [CrossRef]
- Hildner, K.; Edelson, B.T.; Purtha, W.E.; Diamond, M.; Matsushita, H.; Kohyama, M.; Calderon, B.; Schraml, B.U.; Unanue, E.R.; Diamond, M.S.; et al. Batf3 deficiency reveals a critical role for CD8alpha+ dendritic cells in cytotoxic T cell immunity. Science 2008, 322, 1097–1100. [Google Scholar] [CrossRef] [Green Version]
- Böttcher, J.P.; Reis e Sousa, C. The Role of Type 1 Conventional Dendritic Cells in Cancer Immunity. Trends Cancer 2018, 4, 784–792. [Google Scholar] [CrossRef] [Green Version]
- Villani, A.-C.; Satija, R.; Reynolds, G.; Sarkizova, S.; Shekhar, K.; Fletcher, J.; Griesbeck, M.; Butler, A.; Zheng, S.; Lazo, S.; et al. Single-cell RNA-seq reveals new types of human blood dendritic cells, monocytes, and progenitors. Science 2017, 356, eaah4573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binnewies, M.; Mujal, A.M.; Pollack, J.L.; Combes, A.J.; Hardison, E.A.; Barry, K.C.; Tsui, J.; Ruhland, M.K.; Kersten, K.; Abushawish, M.A.; et al. Unleashing Type-2 Dendritic Cells to Drive Protective Antitumor CD4+ T Cell Immunity. Cell 2019, 177, 556–571.e16. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, J.M.; Ibrahim, J.G.; Sosman, J.A.; Sondak, V.K.; Agarwala, S.S.; Ernstoff, M.S.; Rao, U. High-Dose Interferon Alfa-2b Significantly Prolongs Relapse-Free and Overall Survival Compared with the GM2-KLH/QS-21 Vaccine in Patients With Resected Stage IIB-III Melanoma: Results of Intergroup Trial E1694/S9512/C509801. J. Clin. Oncol. 2001, 19, 2370–2380. [Google Scholar] [CrossRef]
- Tarhini, A.A.; Cherian, J.; Moschos, S.J.; Tawbi, H.A.; Shuai, Y.; Gooding, W.E.; Sander, C.; Kirkwood, J.M. Safety and Efficacy of Combination Immunotherapy With Interferon Alfa-2b and Tremelimumab in Patients with Stage IV Melanoma. J. Clin. Oncol. 2011, 30, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Conrad, C.; Gregorio, J.; Wang, Y.-H.; Ito, T.; Meller, S.; Hanabuchi, S.; Anderson, S.; Atkinson, N.; Ramirez, P.T.; Liu, Y.-J.; et al. Plasmacytoid dendritic cells promote immunosuppression in ovarian cancer via ICOS costimulation of Foxp3(+) T-regulatory cells. Cancer Res. 2012, 72, 5240–5249. [Google Scholar] [CrossRef] [Green Version]
- Aspord, C.; Leccia, M.-T.; Charles, J.; Plumas, J. Plasmacytoid Dendritic Cells Support Melanoma Progression by Promoting Th2 and Regulatory Immunity through OX40L and ICOSL. Cancer Immunol. Res. 2013, 1, 402–415. [Google Scholar] [CrossRef] [Green Version]
- Combes, A.; Camosseto, V.; N’Guessan, P.; Argüello, R.J.; Mussard, J.; Caux, C.; Bendriss-Vermare, N.; Pierre, P.; Gatti, E. BAD-LAMP controls TLR9 trafficking and signalling in human plasmacytoid dendritic cells. Nat. Commun. 2017, 8, 913. [Google Scholar] [CrossRef] [Green Version]
- Demoulin, S.; Herfs, M.; Delvenne, P.; Hubert, P. Tumor microenvironment converts plasmacytoid dendritic cells into immunosuppressive/tolerogenic cells: Insight into the molecular mechanisms. J. Leukoc. Biol. 2013, 93, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Matthews, W.; Jordan, C.T.; Wiegand, G.W.; Pardoll, D.; Lemischka, I.R. A receptor tyrosine kinase specific to hematopoietic stem and progenitor cell-enriched populations. Cell 1991, 65, 1143–1152. [Google Scholar] [CrossRef]
- Waskow, C.; Liu, K.; Darrasse-Jèze, G.; Guermonprez, P.; Ginhoux, F.; Merad, M.; Shengelia, T.; Yao, K.; Nussenzweig, M. The receptor tyrosine kinase Flt3 is required for dendritic cell development in peripheral lymphoid tissues. Nat. Immunol. 2008, 9, 676–683. [Google Scholar] [CrossRef]
- Ginhoux, F.; Liu, K.; Helft, J.; Bogunovic, M.; Greter, M.; Hashimoto, D.; Price, J.; Yin, N.; Bromberg, J.; Lira, S.A.; et al. The origin and development of nonlymphoid tissue CD103 + DCs. J. Exp. Med. 2009, 206, 3115–3130. [Google Scholar] [CrossRef] [Green Version]
- Ray, R.J.; Paige, C.J.; Furlonger, C.; Lyman, S.D.; Rottapel, R. Flt3 ligand supports the differentiation of early B cell progenitors in the presence of interleukin-11 and interleukin-7. Eur. J. Immunol. 1996, 26, 1504–1510. [Google Scholar] [CrossRef] [PubMed]
- Svensson, M.N.D.; Andersson, K.M.E.; Wasén, C.; Erlandsson, M.C.; Nurkkala-Karlsson, M.; Jonsson, I.M.; Brisslert, M.; Bemark, M.; Bokarewa, M.I. Murine germinal center B cells require functional fms-like tyrosine kinase 3 signaling for IgG1 class-switch recombination. Proc. Natl. Acad. Sci. USA 2015, 112, E6644–E6653. [Google Scholar] [CrossRef] [Green Version]
- Baerenwaldt, A.; von Burg, N.; Kreuzaler, M.; Sitte, S.; Horvath, E.; Peter, A.; Voehringer, D.; Rolink, A.G.; Finke, D. Flt3 Ligand Regulates the Development of Innate Lymphoid Cells in Fetal and Adult Mice. J. Immunol. 2016, 196, 2561–2571. [Google Scholar] [CrossRef]
- Parigi, S.M.; Czarnewski, P.; Das, S.; Steeg, C.; Brockmann, L.; Fernandez-Gaitero, S.; Yman, V.; Forkel, M.; Höög, C.; Mjösberg, J.; et al. Flt3 ligand expands bona fide innate lymphoid cell precursors in vivo. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Agnès, F.; Shamoon, B.; Dina, C.; Rosnet, O.; Birnbaum, D.; Galibert, F. Genomic structure of the downstream part of the human FLT3 gene: Exon/intron structure conservation among genes encoding receptor tyrosine kinases (RTK) of subclass III. Gene 1994, 145, 283–288. [Google Scholar] [CrossRef]
- Rosnet, O.; Birnbaum, D. Hematopoietic receptors of class III receptor-type tyrosine kinases. Crit. Rev. Oncog. 1993, 4, 595–613. [Google Scholar] [PubMed]
- Gary Gilliland, D.; Griffin, J.D. The roles of FLT3 in hematopoiesis and leukemia. Blood 2002, 100, 1532–1542. [Google Scholar] [CrossRef] [Green Version]
- Lyman, S.D.; James, L.; Bos, T.V.; de Vries, P.; Brasel, K.; Gliniak, B.; Hollingsworth, L.T.; Picha, K.S.; McKenna, H.J.; Splett, R.R.; et al. Molecular cloning of a ligand for the flt3 flk-2 tyrosine kinase receptor: A proliferative factor for primitive hematopoietic cells. Cell 1993, 75, 1157–1167. [Google Scholar] [CrossRef]
- Lyman, S.D.; James, L.; Johnson, L.; Brasel, K.; de Vries, P.; Escobar, S.S.; Downey, H.; Splett, R.R.; Beckmann, M.P.; McKenna, H.J. Cloning of the human homologue of the murine flt3 ligand: A growth factor for early hematopoietic progenitor cells. Blood 1994, 83, 2795–2801. [Google Scholar] [CrossRef] [PubMed]
- Hannum, C.; Culpepper, J.; Campbell, D.; McClanahan, T.; Zurawski, S.; Kastelein, R.; Bazan, J.F.; Hudak, S.; Wagner, J.; Mattson, J.; et al. Ligand for FLT3/FLK2 receptor tyrosine kinase regulates growth of haematopoietic stem cells and is encoded by variant RNAs. Nature 1994, 368, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Lisovsky, M.; Braun, S.E.; Ge, Y.; Takahira, H.; Lu, L.; Savchenko, V.G.; Lyman, S.D.; Broxmeyer, H.E. Flt3-ligand production by human bone marrow stromal cells. Leukemia 1996, 10, 1012–1018. [Google Scholar]
- Chklovskaia, E.; Nissen, C.; Landmann, L.; Rahner, C.; Pfister, O.; Wodnar-Filipowicz, A. Cell-surface trafficking and release of flt3 ligand from T lymphocytes is induced by common cytokine receptor gamma-chain signaling and inhibited by cyclosporin A. Blood 2001, 97, 1027–1034. [Google Scholar] [CrossRef]
- Barry, K.C.; Hsu, J.; Broz, M.L.; Cueto, F.J.; Binnewies, M.; Combes, A.J.; Nelson, A.E.; Loo, K.; Kumar, R.; Rosenblum, M.D.; et al. A natural killer–dendritic cell axis defines checkpoint therapy–responsive tumor microenvironments. Nat. Med. 2018, 24, 1178–1191. [Google Scholar] [CrossRef]
- Wodnar-Filipowicz, A.; Lyman, S.D.; Gratwohl, A.; Tichelli, A.; Speck, B.; Nissen, C. Flt3 ligand level reflects hematopoietic progenitor cell function in aplastic anemia and chemotherapy-induced bone marrow aplasia. Blood 1996, 88, 4493–4499. [Google Scholar] [CrossRef] [Green Version]
- Balog, R.P.; Bacher, R.; Chang, P.; Greenstein, M.; Jammalamadaka, S.; Javitz, H.; Knox, S.J.; Lee, S.; Lin, H.; Shaler, T.; et al. Development of a biodosimeter for radiation triage using novel blood protein biomarker panels in humans and non-human primates. Int. J. Radiat. Biol. 2019, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lyman, S.; James, L.; Escobar, S.; Downey, H.; de Vries, P.; Brasel, K.; Stocking, K.; Beckmann, M.; Copeland, N.; Cleveland, L. Identification of soluble and membrane-bound isoforms of the murine flt3 ligand generated by alternative splicing of mRNAs. Oncogene 1995, 10, 149–157. [Google Scholar] [PubMed]
- Horiuchi, K.; Morioka, H.; Takaishi, H.; Akiyama, H.; Blobel, C.P.; Toyama, Y. Ectodomain Shedding of FLT3 Ligand Is Mediated by TNF-α Converting Enzyme. J. Immunol. 2009, 182, 7408–7414. [Google Scholar] [CrossRef] [Green Version]
- Kingston, D.; Schmid, M.A.; Onai, N.; Obata-Onai, A.; Baumjohann, D.; Manz, M.G. The concerted action of GM-CSF and Flt3-ligand on in vivo dendritic cell homeostasis. Blood 2009, 114, 835–843. [Google Scholar] [CrossRef]
- Durai, V.; Bagadia, P.; Briseño, C.G.; Theisen, D.J.; Iwata, A.; Davidson, J.T.; Gargaro, M.; Fremont, D.H.; Murphy, T.L.; Murphy, K.M. Altered compensatory cytokine signaling underlies the discrepancy between Flt3−/− and Flt3l−/− mice. J. Exp. Med. 2018, 215, 1417–1435. [Google Scholar] [CrossRef] [Green Version]
- Wculek, S.K.; Amores-Iniesta, J.; Conde-Garrosa, R.; Khouili, S.C.; Melero, I.; Sancho, D. Effective cancer immunotherapy by natural mouse conventional type-1 dendritic cells bearing dead tumor antigen. J. Immunother. Cancer 2019, 7, 100. [Google Scholar] [CrossRef] [PubMed]
- Broz, M.L.; Binnewies, M.; Boldajipour, B.; Nelson, A.E.; Pollack, J.L.; Erle, D.J.; Barczak, A.; Rosenblum, M.D.; Daud, A.; Barber, D.L.; et al. Dissecting the Tumor Myeloid Compartment Reveals Rare Activating Antigen-Presenting Cells Critical for T Cell Immunity. Cancer Cell 2014, 26, 638–652. [Google Scholar] [CrossRef] [Green Version]
- Solheim, J.C.; Reber, A.J.; Ashour, A.E.; Robinson, S.; Futakuchi, M.; Kurz, S.G.; Hood, K.; Fields, R.R.; Shafer, L.R.; Cornell, D.; et al. Spleen but not tumor infiltration by dendritic and T cells is increased by intravenous adenovirus-Flt3 ligand injection. Cancer Gene. 2007, 14, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furumoto, K.; Soares, L.; Engleman, E.G.; Merad, M. Induction of potent antitumor immunity by in situ targeting of intratumoral DCs. J. Clin. Investig. 2004, 113, 774–783. [Google Scholar] [CrossRef] [Green Version]
- Stirewalt, D.L.; Radich, J.P. The role of FLT3 in haematopoietic malignancies. Nat. Rev. Cancer 2003, 3, 650–665. [Google Scholar] [CrossRef] [PubMed]
- Verstraete, K.; Vandriessche, G.; Januar, M.; Elegheert, J.; Shkumatov, A.V.; Desfosses, A.; Van Craenenbroeck, K.; Svergun, D.I.; Gutsche, I.; Vergauwen, B.; et al. Structural insights into the extracellular assembly of the hematopoietic Flt3 signaling complex. Blood 2011, 118, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.M.; Lin, N.L.; Issarachai, S.; Lyman, S.D.; Broudy, V.C. FLT3 receptor expression on the surface of normal and malignant human hematopoietic cells. Blood 1996, 88, 3383–3390. [Google Scholar] [CrossRef]
- Zhang, S.; Broxmeyer, H.E. Flt3 Ligand Induces Tyrosine Phosphorylation of Gab1 and Gab2 and Their Association with Shp-2, Grb2, and PI3 Kinase. Biochem. Biophys. Res. Commun. 2000, 277, 195–199. [Google Scholar] [CrossRef]
- Dosil, M.; Wang, S.; Lemischka, I.R. Mitogenic signalling and substrate specificity of the Flk2/Flt3 receptor tyrosine kinase in fibroblasts and interleukin 3-dependent hematopoietic cells. Mol. Cell. Biol. 1993, 13, 6572–6585. [Google Scholar] [CrossRef] [Green Version]
- Rottapel, R.; Turck, C.W.; Casteran, N.; Liu, X.; Birnbaum, D.; Pawson, T.; Dubreuil, P. Substrate specificities and identification of a putative binding site for P13K in the carboxy tail of the murine Flt3 receptor tyrosine kinase. Oncogene 1994, 9, 1755–1765. [Google Scholar] [PubMed]
- Marchetto, S.; Fournier, E.; Beslu, N.; Aurran-Schleinitz, T.; Dubreuil, P.; Borg, J.P.; Birnbaum, D.; Rosnet, O. SHC and SHIP phosphorylation and interaction in response to activation of the FLT3 receptor. Leukemia 1999, 13, 1374–1382. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Fukuda, S.; Lee, Y.; Hangoc, G.; Cooper, S.; Spolski, R.; Leonard, W.J.; Broxmeyer, H.E. Essential Role of Signal Transducer and Activator of Transcription (Stat)5a but Not Stat5b for Flt3-Dependent Signaling. J. Exp. Med. 2000, 192, 719–728. [Google Scholar] [CrossRef]
- Laouar, Y.; Welte, T.; Fu, X.-Y.; Flavell, R.A. STAT3 Is Required for Flt3L-Dependent Dendritic Cell Differentiation. Immunity 2003, 19, 903–912. [Google Scholar] [CrossRef] [Green Version]
- Esashi, E.; Wang, Y.-H.; Perng, O.; Qin, X.-F.; Liu, Y.-J.; Watowich, S.S. The Signal Transducer STAT5 Inhibits Plasmacytoid Dendritic Cell Development by Suppressing Transcription Factor IRF8. Immunity 2008, 28, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.S.; Yang, C.Y.; Nallaparaju, K.C.; Zhang, H.; Liu, Y.J.; Goldrath, A.W.; Watowich, S.S. The signal transducers STAT5 and STAT3 control expression of Id2 and E2-2 during dendritic cell development. Blood 2012, 120, 4363–4373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathaliyawala, T.; O’Gorman, W.E.; Greter, M.; Bogunovic, M.; Konjufca, V.; Hou, Z.E.; Nolan, G.P.; Miller, M.J.; Merad, M.; Reizis, B. Mammalian Target of Rapamycin Controls Dendritic Cell Development Downstream of Flt3 Ligand Signaling. Immunity 2010, 33, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Brasel, K.; McKenna, H.J.; Morrissey, P.J.; Charrier, K.; Morris, A.E.; Lee, C.C.; Williams, D.E.; Lyman, S.D. Hematologic effects of flt3 ligand in vivo in mice. Blood 1996, 88, 2004–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraskovsky, E.; Brasel, K.; Teepe, M.; Roux, E.R.; Lyman, S.D.; Shortman, K.; McKenna, H.J. Dramatic increase in the numbers of functionally mature dendritic cells in Flt3 ligand-treated mice: Multiple dendritic cell subpopulations identified. J. Exp. Med. 1996, 184, 1953–1962. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.T.; Kharazi, S.; Boiers, C.; Cheng, M.; Lubking, A.; Sitnicka, E.; Jacobsen, S.E.W. FLT3 ligand and not TSLP is the key regulator of IL-7-independent B-1 and B-2 B lymphopoiesis. Blood 2008, 112, 2297–2304. [Google Scholar] [CrossRef] [Green Version]
- Karsunky, H.; Merad, M.; Cozzio, A.; Weissman, I.L.; Manz, M.G. Flt3 Ligand Regulates Dendritic Cell Development from Flt3 + Lymphoid and Myeloid-committed Progenitors to Flt3 + Dendritic Cells In Vivo. J. Exp. Med. 2003, 198, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Guimond, M.; Freud, A.G.; Mao, H.C.; Yu, J.; Blaser, B.W.; Leong, J.W.; Vandeusen, J.B.; Dorrance, A.; Zhang, J.; Mackall, C.L.; et al. In Vivo Role of Flt3 Ligand and Dendritic Cells in NK Cell Homeostasis. J. Immunol. 2010, 184, 2769–2775. [Google Scholar] [CrossRef] [Green Version]
- Maraskovsky, E.; Daro, E.; Roux, E.; Teepe, M.; Maliszewski, C.R.; Hoek, J.; Caron, D.; Lebsack, M.E.; McKenna, H.J. In vivo generation of human dendritic cell subsets by Flt3 ligand. Blood 2000, 96, 878–884. [Google Scholar] [CrossRef]
- Breton, G.; Lee, J.; Zhou, Y.J.; Schreiber, J.J.; Keler, T.; Puhr, S.; Anandasabapathy, N.; Schlesinger, S.; Caskey, M.; Liu, K.; et al. Circulating precursors of human CD1c + and CD141 + dendritic cells. J. Exp. Med. 2015, 212, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Hawley, T.S.; Fong, A.Z.C.; Griesser, H.; Lyman, S.D.; Hawley, R.G. Leukemic Predisposition of Mice Transplanted With Gene-Modified Hematopoietic Precursors Expressing flt3 Ligand. Blood 1998, 92, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Brasel, K.; De Smedt, T.; Smith, J.L.; Maliszewski, C.R. Generation of murine dendritic cells from flt3-ligand-supplemented bone marrow cultures. Blood 2000, 96, 3029–3039. [Google Scholar] [CrossRef]
- Gilliet, M.; Boonstra, A.; Paturel, C.; Antonenko, S.; Xu, X.-L.; Trinchieri, G.; O’Garra, A.; Liu, Y.-J. The Development of Murine Plasmacytoid Dendritic Cell Precursors Is Differentially Regulated by FLT3-ligand and Granulocyte/Macrophage Colony-Stimulating Factor. J. Exp. Med. 2002, 195, 953–958. [Google Scholar] [CrossRef]
- Helft, J.; Böttcher, J.; Chakravarty, P.; Zelenay, S.; Huotari, J.; Schraml, B.U.; Goubau, D.; Reise Sousa, C. GM-CSF Mouse Bone Marrow Cultures Comprise a Heterogeneous Population of CD11c+MHCII+ Macrophages and Dendritic Cells. Immunity 2015, 42, 1197–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkling, M.E.; Cytlak, U.; Lau, C.M.; Lewis, K.L.; Resteu, A.; Khodadadi-Jamayran, A.; Siebel, C.W.; Salmon, H.; Merad, M.; Tsirigos, A.; et al. Notch Signaling Facilitates In Vitro Generation of Cross-Presenting Classical Dendritic Cells. Cell Rep. 2018, 23, 3658–3672.e6. [Google Scholar] [CrossRef] [PubMed]
- Balan, S.; Arnold-Schrauf, C.; Abbas, A.; Couespel, N.; Savoret, J.; Imperatore, F.; Villani, A.C.; Vu Manh, T.P.; Bhardwaj, N.; Dalod, M. Large-Scale Human Dendritic Cell Differentiation Revealing Notch-Dependent Lineage Bifurcation and Heterogeneity. Cell Rep. 2018, 24, 1902–1915.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, C.T.; Ghorbani, P.; Nandan, A.; Dudek, M.; Arnold-Schrauf, C.; Hesse, C.; Berod, L.; Stüve, P.; Puttur, F.; Merad, M.; et al. Selective and efficient generation of functional Batf3-dependent CD103+ dendritic cells from mouse bone marrow. Blood 2014, 124, 3081–3091. [Google Scholar] [CrossRef]
- Salmon, H.; Idoyaga, J.; Rahman, A.; Leboeuf, M.; Remark, R.; Jordan, S.; Casanova-Acebes, M.; Khudoynazarova, M.; Agudo, J.; Tung, N.; et al. Expansion and Activation of CD103+ Dendritic Cell Progenitors at the Tumor Site Enhances Tumor Responses to Therapeutic PD-L1 and BRAF Inhibition. Immunity 2016, 44, 924–938. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Paulete, A.R.; Cueto, F.J.; Martinez-Lopez, M.; Labiano, S.; Morales-Kastresana, A.; Rodriguez-Ruiz, M.E.; Jure-Kunkel, M.; Azpilikueta, A.; Aznar, M.A.; Quetglas, J.I.; et al. Cancer immunotherapy with immunomodulatory anti-CD137 and anti–PD-1 monoclonal antibodies requires BATF3-dependent dendritic cells. Cancer Discov. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Theisen, D.J.; Ferris, S.T.; Briseño, C.G.; Kretzer, N.; Iwata, A.; Murphy, K.M.; Murphy, T.L. Batf3-dependent genes control tumor rejection induced by dendritic cells independently of cross-presentation. Cancer Immunol. Res. 2019, 7, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, D.H.; Andreasen, A.; Maraskovsky, E.; Whitmore, J.; Miller, R.E.; Schuh, J.C.L. Flt3 ligand induces tumor regression and antitumor immune responses in vivo. Nat. Med. 1997, 3, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Braun, S.; Lyman, S.; Fan, Y.; Traycoff, C.M.; Wiebke, E.A.; Gaddy, J.; Sledge, G.; Broxmeyer, H.E.; Cornetta, K. Antitumor activity and immunotherapeutic properties of Flt3-ligand in a murine breast cancer model. Cancer Res. 1997, 57, 3511–3516. [Google Scholar]
- Esche, C.; Subbotin, V.M.; Maliszewski, C.; Lotze, M.T.; Shurin, M.R. FLT3 ligand administration inhibits tumor growth in murine melanoma and lymphoma. Cancer Res. 1998, 58, 380–383. [Google Scholar] [PubMed]
- Lai, J.; Mardiana, S.; House, I.G.; Sek, K.; Henderson, M.A.; Giuffrida, L.; Chen, A.X.Y.; Todd, K.L.; Petley, E.V.; Chan, J.D.; et al. Adoptive cellular therapy with T cells expressing the dendritic cell growth factor Flt3L drives epitope spreading and antitumor immunity. Nat. Immunol. 2020, 21, 914–926. [Google Scholar] [CrossRef]
- Frei, E.; Holland, J.F.; Schneidermann, M.A.; Pinkel, D.; Selkirk, G.; Freireich, E.J.; Silver, R.T.; Gold, G.L.; Regelson, W. A Comparative Study of Two Regimens of Combination Chemotherapy in Acute Leukemia. Blood 1958, 13, 1126–1148. [Google Scholar] [CrossRef] [Green Version]
- Hailemichael, Y.; Woods, A.; Fu, T.; He, Q.; Nielsen, M.C.; Hasan, F.; Roszik, J.; Xiao, Z.; Vianden, C.; Khong, H.; et al. Cancer vaccine formulation dictates synergy with CTLA-4 and PD-L1 checkpoint blockade therapy. J. Clin. Investig. 2018, 128, 1338–1354. [Google Scholar] [CrossRef]
- Chakravarty, P.K.; Alfieri, A.; Thomas, E.K.; Beri, V.; Tanaka, K.E.; Vikram, B.; Guha, C. Flt3-ligand administration after radiation therapy prolongs survival in a murine model of metastatic lung cancer. Cancer Res. 1999, 59, 6028–6032. [Google Scholar]
- Rodriguez-Ruiz, M.E.; Rodriguez, I.; Garasa, S.; Barbes, B.; Solorzano, J.L.; Perez-Gracia, J.L.; Labiano, S.; Sanmamed, M.F.; Azpilikueta, A.; Bolaños, E.; et al. Abscopal effects of radiotherapy are enhanced by combined immunostimulatory mAbs and are dependent on CD8 T cells and crosspriming. Cancer Res. 2016, 76, 5994–6005. [Google Scholar] [CrossRef] [Green Version]
- Merad, M.; Sugie, T.; Engleman, E.G.; Fong, L. In vivo manipulation of dendritic cells to induce therapeutic immunity. Blood 2002, 99, 1676–1682. [Google Scholar] [CrossRef] [Green Version]
- Spranger, S.; Bao, R.; Gajewski, T.F. Melanoma-intrinsic β-catenin signalling prevents anti-tumour immunity. Nature 2015, 523, 231–235. [Google Scholar] [CrossRef]
- Curran, M.A.; Allison, J.P. Tumor Vaccines Expressing Flt3 Ligand Synergize with CTLA-4 Blockade to Reject Preimplanted Tumors. Cancer Res. 2009, 69, 7747–7755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffell, B.; Chang-Strachan, D.; Chan, V.; Rosenbusch, A.; Ho, C.M.T.; Pryer, N.; Daniel, D.; Hwang, E.S.; Rugo, H.S.; Coussens, L.M. Macrophage IL-10 Blocks CD8+ T Cell-Dependent Responses to Chemotherapy by Suppressing IL-12 Expression in Intratumoral Dendritic Cells. Cancer Cell 2014, 26, 623–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garris, C.S.; Arlauckas, S.P.; Kohler, R.H.; Trefny, M.P.; Garren, S.; Piot, C.; Engblom, C.; Pfirschke, C.; Siwicki, M.; Gungabeesoon, J.; et al. Successful Anti-PD-1 Cancer Immunotherapy Requires T Cell-Dendritic Cell Crosstalk Involving the Cytokines IFN-γ and IL-12. Immunity 2018, 49, 1148–1161.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerich, L.; Marron, T.U.; Upadhyay, R.; Svensson-Arvelund, J.; Dhainaut, M.; Hussein, S.; Zhan, Y.; Ostrowski, D.; Yellin, M.; Marsh, H.; et al. Systemic clinical tumor regressions and potentiation of PD1 blockade with in situ vaccination. Nat. Med. 2019, 25, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Gilfillan, C.B.; Kuhn, S.; Baey, C.; Hyde, E.J.; Yang, J.; Ruedl, C.; Ronchese, F. Clec9A + Dendritic Cells Are Not Essential for Antitumor CD8 + T Cell Responses Induced by Poly I:C Immunotherapy. J. Immunol. 2018, 200, 2978–2986. [Google Scholar] [CrossRef] [Green Version]
- Enamorado, M.; Iborra, S.; Priego, E.; Cueto, F.J.; Quintana, J.A.; Martínez-Cano, S.; Mejías-Pérez, E.; Esteban, M.; Melero, I.; Hidalgo, A.A.A.; et al. Enhanced anti-tumour immunity requires the interplay between resident and circulating memory CD8+ T cells. Nat. Commun. 2017, 8, 16073. [Google Scholar] [CrossRef]
- Spranger, S.; Dai, D.; Horton, B.; Gajewski, T.F. Tumor-Residing Batf3 Dendritic Cells Are Required for Effector T Cell Trafficking and Adoptive T Cell Therapy. Cancer Cell 2017, 31, 711–723.e4. [Google Scholar] [CrossRef] [Green Version]
- Morse, M.A.; Nair, S.; Fernandez-Casal, M.; Deng, Y.; St Peter, M.; Williams, R.; Hobeika, A.; Mosca, P.; Clay, T.; Cumming, R.I.; et al. Preoperative Mobilization of Circulating Dendritic Cells by Flt3 Ligand Administration to Patients with Metastatic Colon Cancer. J. Clin. Oncol. 2000, 18, 3883–3893. [Google Scholar] [CrossRef]
- Fong, L.; Hou, Y.; Rivas, A.; Benike, C.; Yuen, A.; Fisher, G.A.; Davis, M.M.; Engleman, E.G. Altered peptide ligand vaccination with Flt3 ligand expanded dendritic cells for tumor immunotherapy. Proc. Natl. Acad. Sci. USA 2001, 98, 8809–8814. [Google Scholar] [CrossRef] [Green Version]
- Disis, M.L.; Rinn, K.; Knutson, K.L.; Davis, D.; Caron, D.; dela Rosa, C.; Schiffman, K. Flt3 ligand as a vaccine adjuvant in association with HER-2/neu peptide-based vaccines in patients with HER-2/neu-overexpressing cancers. Blood 2002, 99, 2845–2850. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Braun, S.E.; Sonpavde, G.; Cornetta, K. Antileukemic activity of Flt3 ligand in murine leukemia. Cancer Res. 2000, 60, 1895–1900. [Google Scholar]
- Bhardwaj, N.; Friedlander, P.A.; Pavlick, A.C.; Ernstoff, M.S.; Gastman, B.R.; Hanks, B.A.; Curti, B.D.; Albertini, M.R.; Luke, J.J.; Blazquez, A.B.; et al. Flt3 ligand augments immune responses to anti-DEC-205-NY-ESO-1 vaccine through expansion of dendritic cell subsets. Nat. Cancer 2020, 1, 1204–1217. [Google Scholar] [CrossRef]
- He, S.; Chu, J.; Vasu, S.; Deng, Y.; Yuan, S.; Zhang, J.; Fan, Z.; Hofmeister, C.C.; He, X.; Marsh, H.C.; et al. FLT3L and plerixafor combination increases hematopoietic stem cell mobilization and leads to improved transplantation outcome. Biol. Blood Marrow Transpl. 2014, 20, 309–313. [Google Scholar] [CrossRef] [Green Version]
- Cohen, P.A.; Koski, G.K.; Czerniecki, B.J.; Bunting, K.D.; Fu, X.-Y.; Wang, Z.; Zhang, W.-J.; Carter, C.S.; Awad, M.; Distel, C.A.; et al. STAT3- and STAT5-dependent pathways competitively regulate the pan-differentiation of CD34pos cells into tumor-competent dendritic cells. Blood 2008, 112, 1832–1843. [Google Scholar] [CrossRef] [PubMed]
- Nefedova, Y.; Huang, M.; Kusmartsev, S.; Bhattacharya, R.; Cheng, P.; Salup, R.; Jove, R.; Gabrilovich, D. Hyperactivation of STAT3 is involved in abnormal differentiation of dendritic cells in cancer. J. Immunol. 2004, 172, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Zelenay, S.; van der Veen, A.G.; Böttcher, J.P.; Snelgrove, K.J.; Rogers, N.; Acton, S.E.; Chakravarty, P.; Girotti, M.R.; Marais, R.; Quezada, S.A.; et al. Cyclooxygenase-Dependent Tumor Growth through Evasion of Immunity. Cell 2015, 162, 1257–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Identifier | Title | Indication | Therapeutic Strategy | Accrual | Clinical Trial Phase |
|---|---|---|---|---|---|
| Status: Completed | |||||
| NCT00006223 | Flt3L in Treating Patients With Acute Myeloid Leukemia | Acute myeloid leukemia in remission | S.c. recombinant FLT3L vs. observation alone | 139 * | III |
| NCT00003431 | Flt3L in Treating Patients With Metastatic Colorectal Cancer | Metastatic colorectal cancer | S.c. recombinant FLT3L before resection of hepatic metastases | 12 * | I |
| NCT00019396 | Flt3L With or Without Vaccine Therapy in Treating Patients With Metastatic Melanoma or Renal Cell Cancer | Stage IV melanoma, stage IV renal cell cancer, recurrent renal cell cancer and recurrent melanoma | S.c. recombinant FLT3L alone or with melanoma-associated peptides | 54–96 * | II |
| NCT00020540 | Biological Therapy in Treating Patients With Metastatic Melanoma or Metastatic Kidney Cancer | Metastatic skin melanoma and metastatic kidney cancer | S.c. recombinant FLT3L with s.c. recombinant CD40L | 5 * | I |
| NCT01465139 | A Study to Evaluate CDX-301 (rhuFlt3L) in Healthy Volunteers | Healthy volunteers | Escalating doses of s.c. recombinant FLT3L (CDX-301) | 30 | I |
| NCT01484470 | Umbilical Cord Transplantation for the Elderly Population | Multiple hematologic malignancies | Biological: StemEx | 18 | II |
| NCT02139267 | Dose-finding, Safety Study of Plasmid DNA Therapeutic Vaccine to Treat Cervical Intraepithelial Neoplasia | Cervical intraepithelial neoplasia | Electroporation of DNA vaccine encoding for FLT3L and shuffled E6 and E7 genes of HPV type 16/18 (GX-188E) | 72 | II |
| Status: Active, not Recruiting | |||||
| NCT02839265 | FLT3 Ligand Immunotherapy and Stereotactic Radiotherapy for Advanced Non-small Cell Lung Cancer | Advanced non-small cell lung cancer | S.c. recombinant FLT3L (CDX-301) with stereotactic body radiotherapy | 29 | II |
| NCT01811992 | Combined Cytotoxic and Immune-Stimulatory Therapy for Glioma | Malignant glioma and glioblastoma multiforme | Dose escalation of adenovirus gene transfer that drives direct tumor killing and FLT3L expression | 19 | I |
| NCT02129075 | CDX-1401 and Poly-ICLC Vaccine Therapy With or Without CDX-301 in Treating Patients With Stage IIB-IV Melanoma | Stage IIB-IV melanoma | S.c. recombinant FLT3L (CDX-301), s.c. or i.d. DEC-205/NY-ESO-1 fusion protein (CDX-1401) and s.c. poly-ICLC | 60 | II |
| Status: Recruiting | |||||
| NCT03789097 | Vaccination With Flt3L, Radiation, and Poly-ICLC | Non-Hodgkin’s lymphoma, metastatic breast cancer and head and neck squamous cell carcinoma | In situ recombinant FLT3L, radiation and Poly ICLC with pembrolizumab | 56 * | I/II |
| NCT01976585 | In Situ Vaccine for Low-Grade Lymphoma: Combination of Intratumoral Flt3L and Poly-ICLC With Low-Dose Radiotherapy | Low-grade B-cell lymphoma | In situ recombinant FLT3L (CDX-301) and poly-ICLC | 30 * | I/II |
| NCT03329950 | A Study of CDX-1140 (CD40) as Monotherapy or in Combination in Patients With Advanced Malignancies | Multiple cancer types | CD40 agonist antibody (CDX-1140) alone vs. combination with recombinant FLT3L (CDX-301) vs. combination with pembrolizumab vs. combination with chemotherapy | 260 * | I |
| Status: not yet Recruiting | |||||
| NCT04491084 | FLT3 Ligand, CD40 Agonist Antibody, and Stereotactic Radiotherapy | Non-small cell lung cancer | FLT3L (CDX-301) with CD40 agonist antibody (CDX-1140) and stereotactic radiotherapy vs. stereotactic radiotherapy alone | 46 * | I/II |
| NCT04616248 | Radio-immunotherapy (CDX-301, Radiotherapy, CDX-1140 and Poly-ICLC) for the Treatment of Unresectable or Metastatic Breast Cancer Patients | Unresectable and metastatic breast cancer | In situ FLT3L, CD40 agonist antibody (CDX-1140), poly ICLC and radiation therapy vs. addition of i.v. CDX-1140 | 36 * | I |
| Status: Unknown | |||||
| NCT03206138 | Safety and Efficacy of GX-188E Administered Via EP Plus GX-I7 or Imiquimod. | Cervical intraepithelial neoplasia 3 | GX-188E with GX-I7 vs. GX-188E with imiquimod | 50 * | |
| NCT02411019 | Safety and Efficacy of GX-188E DNA Therapeutic Vaccine Administered by Electroporation After Observation | Cervical intraepithelial neoplasia 3 | GX-188E | 72 | II |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cueto, F.J.; Sancho, D. The Flt3L/Flt3 Axis in Dendritic Cell Biology and Cancer Immunotherapy. Cancers 2021, 13, 1525. https://doi.org/10.3390/cancers13071525
Cueto FJ, Sancho D. The Flt3L/Flt3 Axis in Dendritic Cell Biology and Cancer Immunotherapy. Cancers. 2021; 13(7):1525. https://doi.org/10.3390/cancers13071525
Chicago/Turabian StyleCueto, Francisco J., and David Sancho. 2021. "The Flt3L/Flt3 Axis in Dendritic Cell Biology and Cancer Immunotherapy" Cancers 13, no. 7: 1525. https://doi.org/10.3390/cancers13071525
APA StyleCueto, F. J., & Sancho, D. (2021). The Flt3L/Flt3 Axis in Dendritic Cell Biology and Cancer Immunotherapy. Cancers, 13(7), 1525. https://doi.org/10.3390/cancers13071525