
Integrated Analysis and Identification of Critical RNA-Binding Proteins in Bladder Cancer

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Datasets and Patient Information
2.2. Construction of an Individualized Prognostic Index Based on RBPs
2.3. Gene Set Enrichment Analysis (GSEA)
2.4. Establishment and Assessment of the Nomogram
2.5. Cell Culture and Reagents
2.5.1. Plasmid Transfection
2.5.2. Cell Viability and 5-Ethynyl-20-Deoxyuridine (EdU) Assays
2.5.3. Transwell Migration and Invasion Assay
2.5.4. Wound-Healing Assay
2.5.5. RNA Extraction and Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.5.6. Western Blotting Assay
2.5.7. In Vivo Experiments
2.5.8. Histology and Immunohistochemistry (IHC)
2.6. Statistical Analysis
3. Results
3.1. Identification of DERBPs
3.2. Identification of DERBPs Associated with Survival in Patients with BC
3.3. Application of the Six-RBPs Signature in the Validation Cohort
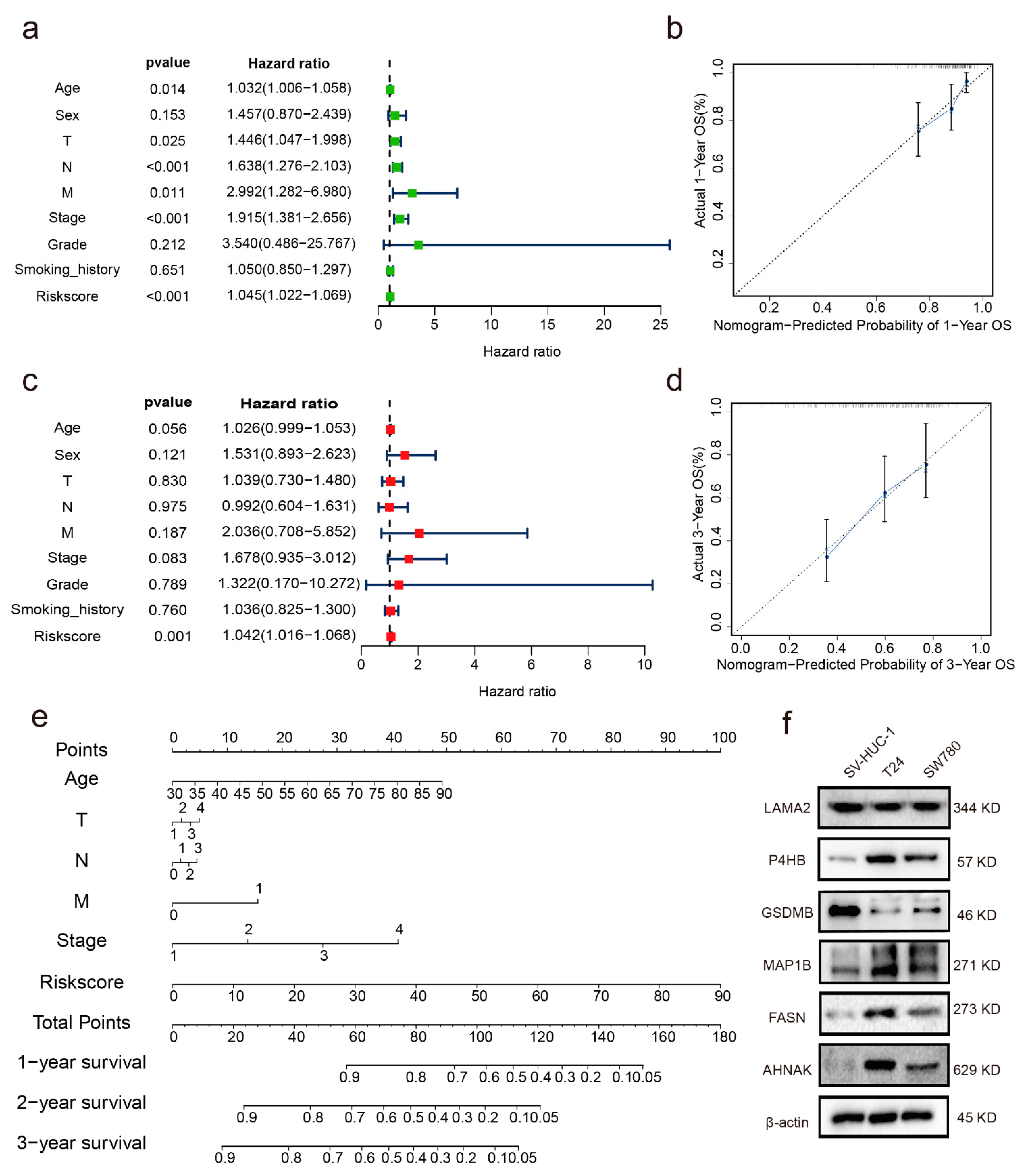
3.4. Independent Prognostic Indicator
3.5. Establishment of a Nomogram
3.6. GSEA
3.7. Verification of the Expression Levels and Prognostic Significance of the Six RBPs
3.8. Assessment of Immune Cell Infiltration
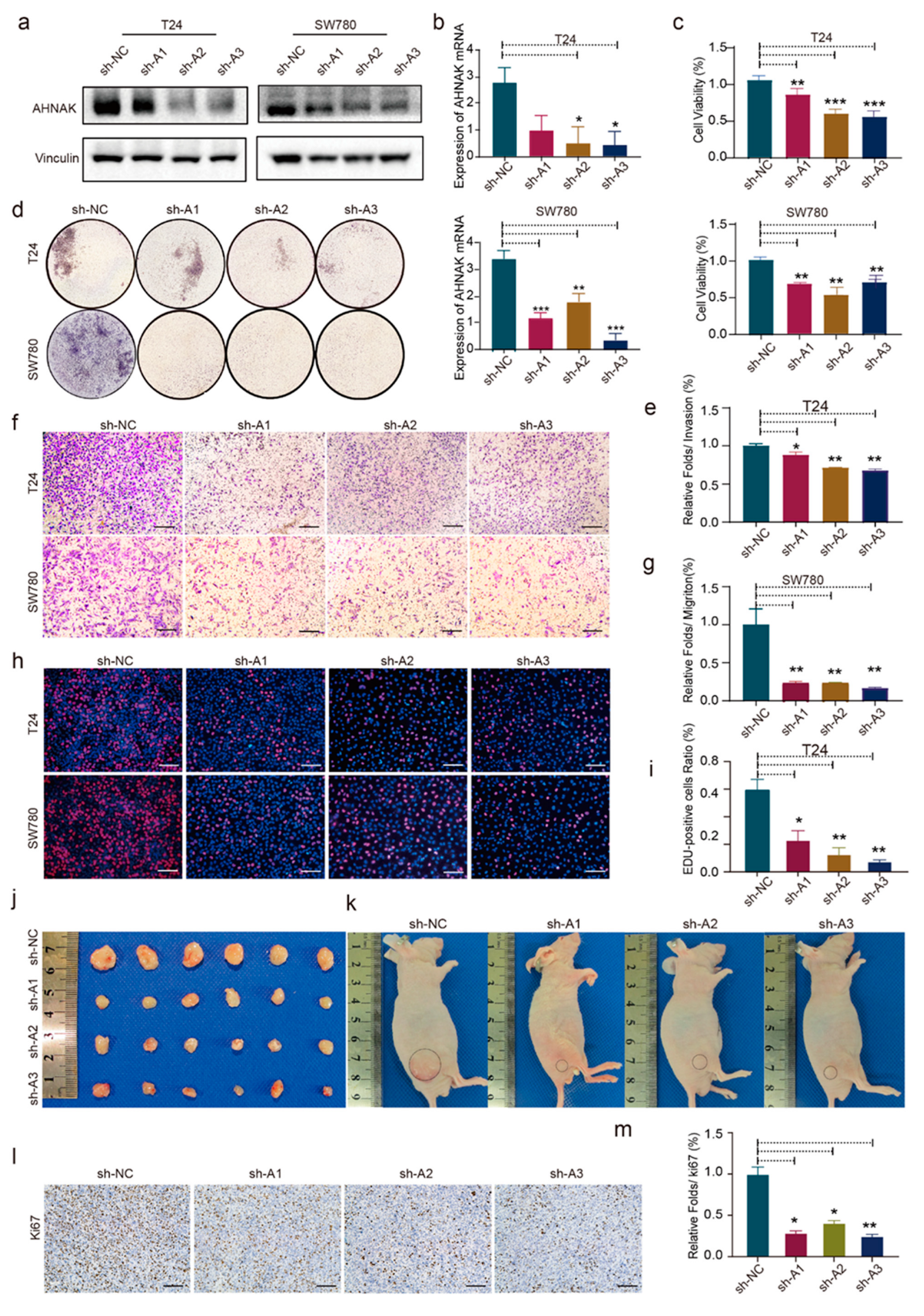
3.9. AHNAK Promotes Cell Proliferation, Migration, and Invasion of BC Cell Lines In Vitro and In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.-S.; Chen, G.; Li, S.-H.; Pang, J.-S.; Cai, K.-T.; Yan, H.-B.; Huang, Z.-G.; He, R.-Q. Identification and validation of an individualized autophagy-clinical prognostic index in bladder cancer patients. OncoTargets Ther. 2019, 12, 3695–3712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moschini, M. From Basic Science to Clinical Research to Develop New Solutions to Improve Diagnoses and Treatment of Bladder Cancer Patients. J. Clin. Med. 2020, 9, 2373. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Transcriptome-wide profiles of circular RNA and RNA-binding protein interactions reveal effects on circular RNA biogenesis and cancer pathway expression. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Li, X.X.; Chung, H.K.; Kalakonda, S.; Cai, J.Z.; Cao, S.; Chen, N.; Liu, Y.; Rao, J.N.; Wang, H.Y.; et al. RNA-Binding Protein HuR Regulates Paneth Cell Function by Altering Membrane Localization of TLR2 via Post-transcriptional Control of CNPY3. Gastroenterology 2019, 157, 731–743. [Google Scholar] [CrossRef]
- Theotoki, E.I.; Pantazopoulou, V.I.; Georgiou, S.; Kakoulidis, P.; Filippa, V.; Stravopodis, D.J.; Anastasiadou, E. Dicing the Disease with Dicer: The Implications of Dicer Ribonuclease in Human Pathologies. Int. J. Mol. Sci. 2020, 21, 7223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Han, B.; Xu, T.; Li, D. The biological function and potential mechanism of long non-coding RNAs in cardiovascular disease. J. Cell Mol. Med. 2020, 24, 12900–12909. [Google Scholar] [CrossRef] [PubMed]
- Schieweck, R.; Ninkovic, J.; Kiebler, M.A. RNA-binding proteins balance brain function in health and disease. Physiol. Rev. 2021, 101, 1309–1370. [Google Scholar] [CrossRef] [PubMed]
- Kechavarzi, B.; Janga, S.C. Dissecting the expression landscape of RNA-binding proteins in human cancers. Genome Biol. 2014, 15, R14. [Google Scholar] [CrossRef] [Green Version]
- Bauer, M.; Vaxevanis, C.; Heimer, N.; Al-Ali, H.K.; Jaekel, N.; Bachmann, M.; Wickenhauser, C.; Seliger, B. Expression, Regulation, and Function of microRNA as Important Players in the Transition of MDS to Secondary AML and Their Cross Talk to RNA-Binding Proteins. Int. J. Mol. Sci. 2020, 21, 7140. [Google Scholar] [CrossRef]
- Correa, B.R.; Araujo, P.R.; Qiao, M.; Burns, S.C.; Chen, C.; Schlegel, R.; Agarwal, S.; Galante, P.A.; Penalva, L.O. Functional genomics analyses of RNA-binding proteins reveal the splicing regulator SNRPB as an oncogenic candidate in glioblastoma. Genome Biol. 2016, 17, 125. [Google Scholar] [CrossRef] [Green Version]
- Neelamraju, Y.; Perez, A.; Nakshatra, P.; Nakshatra, H.; Janga, S.C. Mutational landscape of RNA-binding proteins in human cancers. RNA Biol. 2018, 15, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Zhang, Y.L. Novel Long Non-coding RNA Markers for Prognostic Prediction of Patients with Bladder Cancer. Chin. Med. Sci. J. 2020, 35, 239–247. [Google Scholar] [PubMed]
- Okholm, T.L.H.; Sathe, S.; Park, S.S.; Kamstrup, A.B.; Rasmussen, A.M.; Shankar, A.; Chua, Z.M.; Fristrup, N.; Nielsen, M.M.; Vang, S.; et al. Transcriptome-wide profiles of circular RNA and RNA-binding protein interactions reveal effects on circular RNA biogenesis and cancer pathway expression. Genome Med. 2020, 12, 112. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, Y.; He, A. Identification of the Six-RNA-Binding Protein Signature for Prognosis Prediction in Bladder Cancer. Front. Genet. 2020, 11, 992. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, Z.; Wei, X.; Feng, H.; Hu, B.; Liu, B.; Luan, Y.; Ruan, Y.; Liu, X.; Liu, Z.; et al. Identifying the Functions and Prognostic Values of RNA Binding Proteins in Bladder Cancer. Front. Genet. 2021, 12, 574196. [Google Scholar] [CrossRef]
- Oya, Y.; Hayakawa, Y.; Koike, K. Tumor microenvironment in gastric cancers. Cancer Sci. 2020, 111, 2696–2707. [Google Scholar] [CrossRef]
- Jiang, Y.; Xie, J.; Huang, W.; Chen, H.; Xi, S.; Han, Z.; Huang, L.; Lin, T.; Zhao, L.Y.; Hu, Y.F.; et al. Tumor Immune Microenvironment and Chemosensitivity Signature for Predicting Response to Chemotherapy in Gastric Cancer. Cancer Immunol. Res. 2019, 7, 2065–2073. [Google Scholar] [CrossRef]
- Bi, K.-W.; Wei, X.-G.; Qin, X.-X.; Li, B. BTK Has Potential to Be a Prognostic Factor for Lung Adenocarcinoma and an Indicator for Tumor Microenvironment Remodeling: A Study Based on TCGA Data Mining. Front. Oncol. 2020, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- TCGA. Available online: https://portal.gdc.cancer.gov/ (accessed on 25 September 2021).
- GEO. Available online: https://www.ncbi.nlm.nih.gov/geo/ (accessed on 25 September 2021).
- Gerstberger, S.; Hafner, M.; Tuschl, T. A census of human RNA-binding proteins. Nat. Rev. Genet. 2014, 15, 829–845. [Google Scholar] [CrossRef]
- KEGG. Available online: https://www.kegg.jp/kegg/pathway.html/ (accessed on 25 September 2021).
- GO. Available online: http://geneontology.org/ (accessed on 25 September 2021).
- HPA. Available online: http://www.proteinatlas.org/ (accessed on 25 September 2021).
- Lanczky, A.; Gyorffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef] [Green Version]
- TIMER2.0. Available online: http://timer.comp-genomics.org/ (accessed on 25 September 2021).
- Bai, J.-Y.; Jin, B.; Ma, J.-B.; Liu, T.-J.; Yang, C.; Chong, Y.; Wang, X.; He, D.; Guo, P. HOTAIR and androgen receptor synergistically increase GLI2 transcription to promote tumor angiogenesis and cancer stemness in renal cell carcinoma. Cancer Lett. 2021, 498, 70–79. [Google Scholar] [CrossRef] [PubMed]
- GEMICCL. Available online: https://www.kobic.kr/GEMICCL/browse (accessed on 28 September 2021).
- Mathlin, J.; Pera, L.; Colombo, T. A Census and Categorization Method of Epitranscriptomic Marks. Int. J. Mol. Sci. 2020, 21, 4684. [Google Scholar] [CrossRef] [PubMed]
- Castello, A.; Fischer, B.; Frese, C.K.; Horos, R.; Alleaume, A.M.; Foehr, S.; Curk, T.; Krijgsveld, J.; Hentze, M.W. Comprehensive Identification of RNA-Binding Domains in Human Cells. Mol. Cell 2016, 63, 696–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quattrone, A.; Dassi, E. The Architecture of the Human RNA-Binding Protein Regulatory Network. iScience 2019, 21, 706–719. [Google Scholar] [CrossRef] [Green Version]
- Zekavati, A.; Nasir, A.; Alcaraz, A.; Aldrovandi, M.; Marsh, P.; Norton, J.D.; Murphy, J.J. Post-Transcriptional Regulation of BCL2 mRNA by the RNA-Binding Protein ZFP36L1 in Malignant B Cells. PLoS ONE 2014, 9, e102625. [Google Scholar] [CrossRef] [Green Version]
- Saini, Y.; Chen, J.; Patial, S. The Tristetraprolin Family of RNA-Binding Proteins in Cancer: Progress and Future Prospects. Cancers 2020, 12, 1539. [Google Scholar] [CrossRef]
- Rounbehler, R.J.; Fallahi, M.; Yang, C.Y.; Steeves, M.A.; Li, W.M.; Doherty, J.R.; Schaub, F.X.; Sanduja, S.; Dixon, D.A.; Blackshear, P.J.; et al. Tristetraprolin Impairs Myc-Induced Lymphoma and Abolishes the Malignant State. Cell 2012, 150, 563–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rounbehler, R.J.; Berglund, A.E.; Gerke, T.; Takhar, M.M.; Awasthi, S.; Li, W.M.; Davicioni, E.; Erho, N.G.; Ross, A.E.; Schaeffer, E.M.; et al. Tristetraprolin Is a Prognostic Biomarker for Poor Outcomes among Patients with Low-Grade Prostate Cancer. Cancer Epidemiol. Biomark. Prev. 2018, 27, 1376–1383. [Google Scholar] [CrossRef] [Green Version]
- Schubert, T.; Renninger, M.; Schmid, M.A.; Hassan, F.N.; Sokolakis, I.; Fahmy, O.; Hatzichristodoulou, G.; Stenzl, A.; Gakis, G. Prognostic impact of tumor-associated immune cell infiltrates at radical cystectomy for bladder cancer. Urol. Oncol. 2020, 38, e7–e15. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, L.; Zhou, Q.; Xiong, Y.; Wang, G.; Liu, X.; Xiao, Y.; Ju, L.; Wang, X. Identification of a prognostic gene signature based on an immunogenomic landscape analysis of bladder cancer29. J. Cell Mol. Med. 2020, 24, 13370–13382. [Google Scholar] [CrossRef]
- Qing, L.; Li, Q.; Yang, Y.; Xu, W.; Dong, Z. A prognosis marker MUC1 correlates with metabolism and drug resistance in bladder cancer: A bioinformatics research. BMC Urol. 2022, 22, 114. [Google Scholar] [CrossRef] [PubMed]
- Gadoth, A.; Kryzer, T.J.; Fryer, J.; McKeon, A.; Lennon, V.A.; Pittock, S.J. Microtubule-associated protein 1B: Novel paraneoplastic biomarker. Ann. Neurol. 2017, 81, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Byrne, F.L.; Yang, L.; Phillips, P.A.; Hansford, L.M.; Fletcher, J.I.; Ormandy, C.J.; McCarroll, J.A.; Kavallaris, M. RNAi-mediated stathmin suppression reduces lung metastasis in an orthotopic neuroblastoma mouse model. Oncogene 2014, 3, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wang, J.; Zhuang, J.; Ma, X.; Zheng, N.; Song, Y.; Xia, W. P4HB modulates epithelial-mesenchymal transition and the beta-catenin/Snail pathway influencing chemoresistance in liver cancer cells. Oncol. Lett. 2020, 20, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Peng, Y.; Guan, B. P4HB: A novel diagnostic and prognostic biomarker for bladder carcinoma. Oncol. Lett. 2021, 21, 95. [Google Scholar] [CrossRef]
- Tao, T.; Su, Q.L.; Xu, S.M.; Deng, J.; Zhou, S.C.; Zhuang, Y.; Huang, Y.J.; He, C.M.; He, S.P.; Peng, M.; et al. Down-regulation of PKM2 decreases FASN expression in bladder cancer cells through AKT/mTOR/SREBP-1c axis. J. Cell Physiol. 2019, 234, 3088–3104. [Google Scholar] [CrossRef]
- Komiyama, H.; Aoki, A.; Tanaka, S.; Maekawa, H.; Kato, Y.; Wada, R.; Maekawa, T.; Tamura, M.; Shiroishi, T. Alu-derived cis-element regulates tumorigenesis-dependent gastric expression of GASDERMIN B (GSDMB). Genes Genet. Syst. 2010, 85, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Zhou, Z.; Chai, Y.; Zhang, Y. Upregulated GSDMB in Clear Cell Renal Cell Carcinoma Is Associated with Immune Infiltrates and Poor Prognosis. J. Immunol. Res. 2021, 2021, 1–22. [Google Scholar] [CrossRef]
- Wang, R.-Q.; Lan, Y.-L.; Lou, J.-C.; Lyu, Y.-Z.; Hao, Y.-C.; Su, Q.-F.; Ma, B.-B.; Yuan, Z.-B.; Yu, Z.-K.; Zhang, H.-Q.; et al. Expression and methylation status of LAMA2 are associated with the invasiveness of nonfunctioning PitNET. Ther. Adv. Endocrinol. 2019, 10, 2042018818821296. [Google Scholar] [CrossRef]
- Shen, E.; Wang, X.; Liu, X.; Lv, M.; Zhang, L.; Zhu, G.; Sun, Z. MicroRNA-93-5p promotes epithelial-mesenchymal transition in gastric cancer by repressing tumor suppressor AHNAK expression. Cancer Cell Int. 2020, 20, 76. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Mo, X.; Fu, L.; Xiao, B.; Guo, J. Molecular mechanisms of long noncoding RNAs on gastric cancer. Oncotarget 2016, 7, 8601–8612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiek, D.M.; Erdogdu, B.; Razaghi, R.; Jin, L.; Sadowski, N.; Alamillo-Ferrer, C.; Hogg, J.R.; Haddad, B.R.; Drewry, D.H.; Wells, C.I.; et al. Temozolomide-induced guanine mutations create exploitable vulnerabilities of guanine-rich DNA and RNA regions in drug-resistant gliomas. Sci. Adv. 2022, 8, eabn3471. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Malu, S.; McKenzie, J.A.; Andrews, M.C.; Talukder, A.H.; Tieu, T.; Karpinets, T.V.; Haymaker, C.; Forget, M.-A.; Williams, L.J.; et al. The RNA-binding Protein MEX3B Mediates Resistance to Cancer Immunotherapy by Downregulating HLA-A Expression. Clin. Cancer Res. 2018, 24, 3366–3376. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | TCGA | GEO (GSE32548) | GEO (GSE31684) | |
|---|---|---|---|---|
| Age | <60 | 88 | 32 | 19 |
| >60 | 324 | 114 | 74 | |
| Sex | Female | 108 | 34 | 25 |
| Male | 304 | 112 | 68 | |
| Stage | Ta | 0 | 49 | 8 |
| T1 | 11 | 53 | 19 | |
| Tis | 0 | 1 | 55 | |
| Tx | 6 | 1 | 10 | |
| ≥T2 | 395 | 42 | 1 | |
| 0 | 0 | 1 | 0 | |
| Grade | G1 | 3 | 19 | 0 |
| G2 | 23 | 46 | 6 | |
| G3 | 386 | 80 | 87 | |
| Survival status | dead | 112 | 119 | 65 |
| living | 217 | 26 | 28 | |
| unknown | 83 | 1 | 0 | |
| Primer Name | Sequence 5′–3′ | |
|---|---|---|
| AHNAK | Forward | CTCGTCGCCGCCAGTAG |
| Reverse | TCTTTGCAGGATTCCGCTCA | |
| MAP1B | Forward | AATTCCTGGGCAAACTGGTCT |
| Reverse | AGAGCCGGACTGGAGAATGA | |
| LAMA2 | Forward | GGCTTCCGTTGTCAGCAATC |
| Reverse | CAAGTTTCTCAGCGTTGGCA | |
| P4HB | Forward | TCATCGCCAAGATGGACTCG |
| Reverse | CCACCGCTCTCCAGGAATTT | |
| FASN | Forward | ACCTCCGTGCAGTTCTTGAG |
| Reverse | GTTCAGGATGGTGGCGTACA | |
| GSDMB | Forward | AGACGATGAGAAAGTCTTTGGGT |
| Reverse | TAGCTCCCCGGAAATCAGGA | |
| ID | coef | HR | HR.95L | HR.95H | p Value |
|---|---|---|---|---|---|
| AHNAK | 0.005486 | 1.005501 | 1.00313 | 1.007878 | 5.26 × 10−6 |
| MAP1B | 0.028034 | 1.028431 | 1.004654 | 1.05277 | 0.018821 |
| LAMA2 | 0.053639 | 1.055104 | 0.983493 | 1.131929 | 0.134704 |
| P4HB | 0.002214 | 1.002217 | 1.000971 | 1.003465 | 0.000487 |
| FASN | 0.003685 | 1.003692 | 1.000466 | 1.006929 | 0.024866 |
| GSDMB | −0.05084 | 0.950431 | 0.926115 | 0.975385 | 0.000121 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, L.; Chen, Y.; Li, X.; Mei, Y.; Zhou, J.; Ma, J.; Zhang, M.; Hou, T.; He, D.; Zeng, J. Integrated Analysis and Identification of Critical RNA-Binding Proteins in Bladder Cancer. Cancers 2022, 14, 3739. https://doi.org/10.3390/cancers14153739
Gu L, Chen Y, Li X, Mei Y, Zhou J, Ma J, Zhang M, Hou T, He D, Zeng J. Integrated Analysis and Identification of Critical RNA-Binding Proteins in Bladder Cancer. Cancers. 2022; 14(15):3739. https://doi.org/10.3390/cancers14153739
Chicago/Turabian StyleGu, Lijiang, Yuhang Chen, Xing Li, Yibo Mei, Jinlai Zhou, Jianbin Ma, Mengzhao Zhang, Tao Hou, Dalin He, and Jin Zeng. 2022. "Integrated Analysis and Identification of Critical RNA-Binding Proteins in Bladder Cancer" Cancers 14, no. 15: 3739. https://doi.org/10.3390/cancers14153739
APA StyleGu, L., Chen, Y., Li, X., Mei, Y., Zhou, J., Ma, J., Zhang, M., Hou, T., He, D., & Zeng, J. (2022). Integrated Analysis and Identification of Critical RNA-Binding Proteins in Bladder Cancer. Cancers, 14(15), 3739. https://doi.org/10.3390/cancers14153739