
Understanding Mechanisms of RKIP Regulation to Improve the Development of New Diagnostic Tools



Abstract
Simple Summary
Abstract
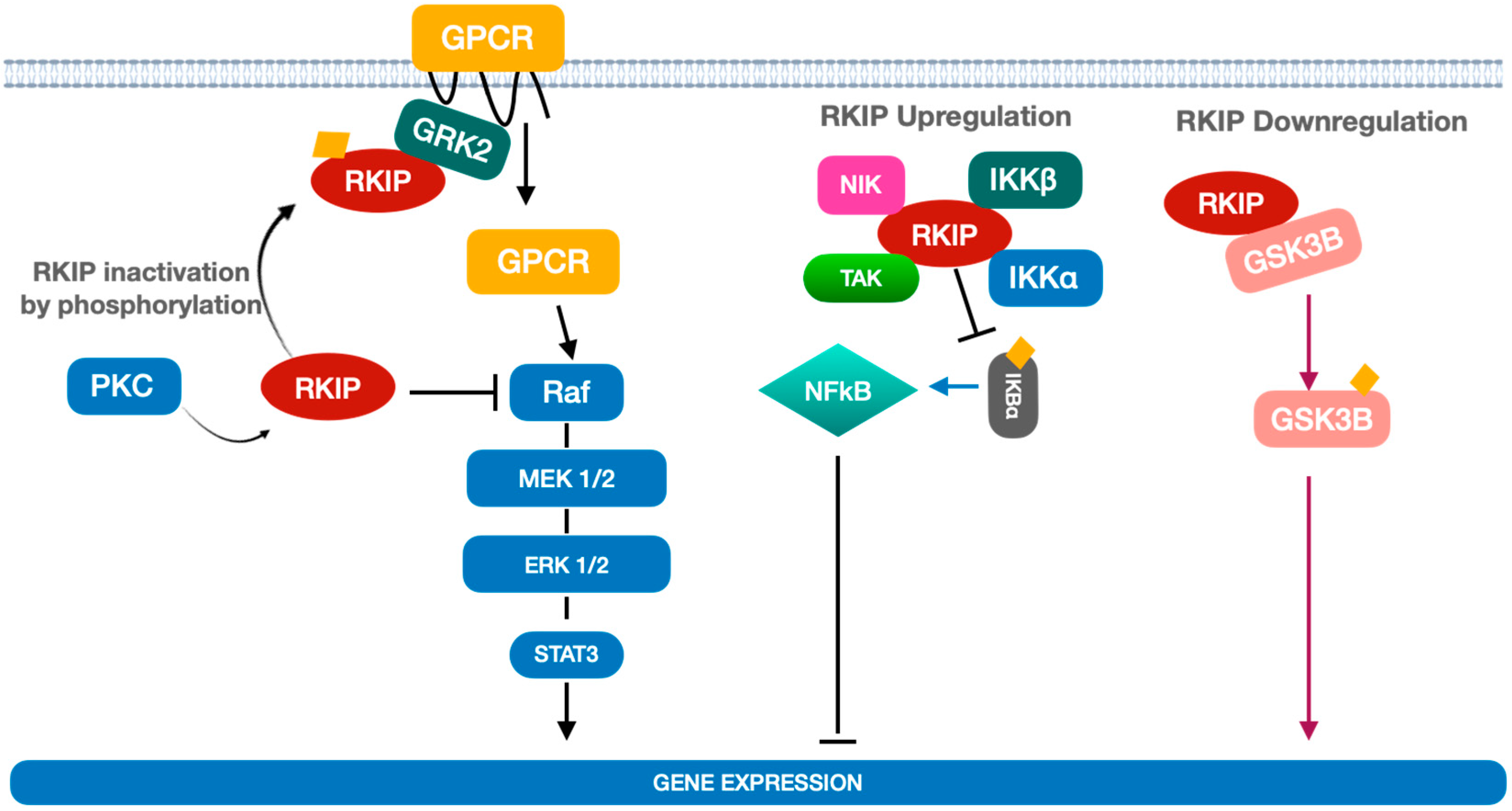
1. Overview on Raf Kinase Inhibitor Protein (RKIP) and Its Biological Functions
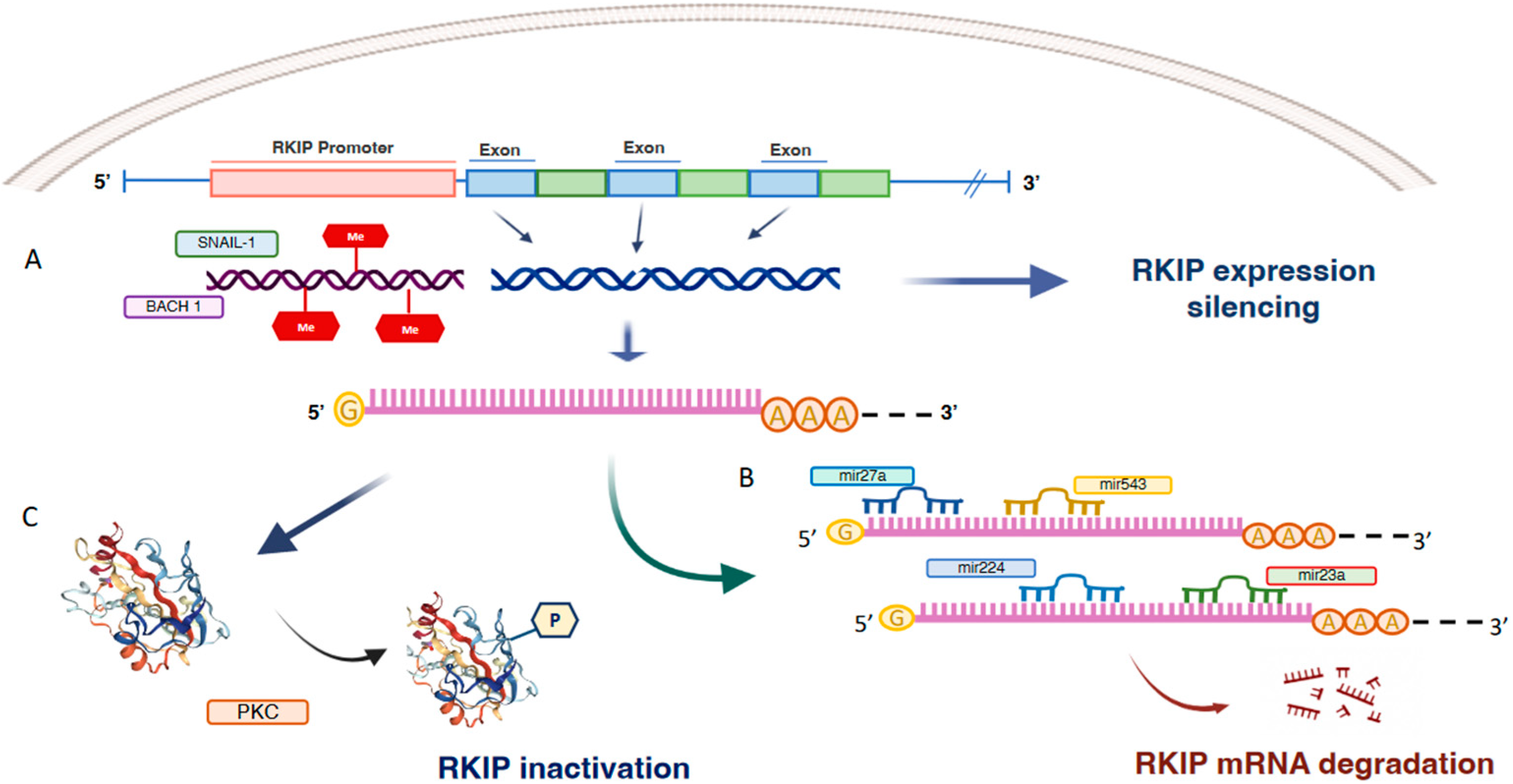
2. Mechanisms of RKIP Regulation
3. RKIP and Cancer
3.1. Lung Cancer
3.2. Colon Cancer
3.3. Breast Cancer
3.4. Myeloid Neoplasms (MNs) and Multiple Myeloma
3.5. Melanoma
3.6. Clear Cells Renal Cell Carcinoma

{kind=link}
{kind=link}
| Human Cancers | RKIP Dysregulation | Ref. |
|---|---|---|
| Lung Cancer |
| [42] |
| [43] | |
| [45] | |
| [48] | |
| [52,53] | |
| Colon Cancer |
| [56] |
| [31] | |
| [57,59] | |
| Breast Cancer |
| [62] |
| [30,61] | |
| [62] | |
| Myeloid neoplasms (MNs) and Multiple Myeloma |
| [65] |
| [67] | |
| [66] | |
| Melanoma |
| [68] |
| [69] | |
| [70] | |
| [72] | |
| clear cells Renal Cell Carcinoma |
| [73] |
| [73,75] | |
| [73] | |
| [32] |
4. RKIP in Biofluids
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, W.; Liu, H.T. MAPK Signal Pathways in the Regulation of Cell Proliferation in Mammalian Cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Keller, E.T.; Fu, Z.; Brennan, M. The Role of Raf Kinase Inhibitor Protein (RKIP) in Health and Disease. Biochem. Pharm. 2004, 68, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Klysik, J.; Theroux, S.J.; Sedivy, J.M.; Moffit, J.S.; Boekelheide, K. Signaling Crossroads: The Function of Raf Kinase Inhibitory Protein in Cancer, the Central Nervous System and Reproduction. Cell Signal 2008, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, K.; Lohse, M.J.; Quitterer, U. Protein Kinase C Switches the Raf Kinase Inhibitor from Raf-1 to GRK-2. Nature 2003, 426, 574–579. [Google Scholar] [CrossRef]
- Zhao, J.; Wenzel, S. Interactions of RKIP with Inflammatory Signaling Pathways. Crit. Rev. Oncog. 2014, 19, 497–504. [Google Scholar] [CrossRef]
- Yeung, K.C.; Rose, D.W.; Dhillon, A.S.; Yaros, D.; Gustafsson, M.; Chatterjee, D.; McFerran, B.; Wyche, J.; Kolch, W.; Sedivy, J.M. Raf Kinase Inhibitor Protein Interacts with NF-ΚB-Inducing Kinase and TAK1 and Inhibits NF-ΚB Activation. Mol. Cell Biol. 2001, 21, 7207–7217. [Google Scholar] [CrossRef] [PubMed]
- Touboul, R.; Baritaki, S.; Zaravinos, A.; Bonavida, B. RKIP Pleiotropic Activities in Cancer and Inflammatory Diseases: Role in Immunity. Cancers 2021, 13, 6247. [Google Scholar] [CrossRef]
- Gabriela-Freitas, M.; Pinheiro, J.; Raquel-Cunha, A.; Cardoso-Carneiro, D.; Martinho, O. RKIP as an Inflammatory and Immune System Modulator: Implications in Cancer. Biomolecules 2019, 9, 769. [Google Scholar] [CrossRef]
- Al-Mulla, F.; Bitar, M.S.; Al-Maghrebi, M.; Behbehani, A.I.; Al-Ali, W.; Rath, O.; Doyle, B.; Tan, K.Y.; Pitt, A.; Kolch, W. Raf Kinase Inhibitor Protein RKIP Enhances Signaling by Glycogen Synthase Kinase-3β. Cancer Res. 2011, 71, 1334–1343. [Google Scholar] [CrossRef]
- Yesilkanal, A.; Rosner, M. Targeting Raf Kinase Inhibitory Protein Regulation and Function. Cancers 2018, 10, 306. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Song, Y.; Fu, Z.; Yu, W. MiR-27a Regulates Cisplatin Resistance and Metastasis by Targeting RKIP in Human Lung Adenocarcinoma Cells. Mol. Cancer 2014, 13, 193. [Google Scholar] [CrossRef]
- Minoo, P. Extensive DNA Methylation in Normal Colorectal Mucosa in Hyperplastic Polyposis. Gut 2006, 55, 1467–1474. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.E.; Kim, N.I.; Lee, J.S.; Park, M.H.; Yoon, J.H. Reduced RKIP Expression Is Associated with Breast Neoplastic Progression and Is Correlated With Poor Outcomes and Aberrant Methylation in Breast Carcinoma. Appl. Immunohistochem. Mol. Morphol. 2017, 25, 467–474. [Google Scholar] [CrossRef]
- Wei, H.; Liu, Z.; She, H.; Liu, B.; Gu, J.; Wei, D.; Zhang, X.; Wang, J.; Qi, S.; Ping, F. Promoter Methylation and Expression of Raf Kinase Inhibitory Protein in Esophageal Squamous Cell Carcinoma. Oncol. Lett. 2017, 13, 1866–1872. [Google Scholar] [CrossRef] [PubMed]
- Zaravinos, A.; Bonavida, B.; Chatzaki, E.; Baritaki, S. RKIP: A Key Regulator in Tumor Metastasis Initiation and Resistance to Apoptosis: Therapeutic Targeting and Impact. Cancers 2018, 10, 287. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, O.; Qin, J.; Liu, S.; Sun, S.; Liu, H.; Kuang, J.; Jiang, G.; Zhang, W. Cis-Acting Elements and Trans-Acting Factors in the Transcriptional Regulation of Raf Kinase Inhibitory Protein Expression. PLoS ONE 2013, 8, e83097. [Google Scholar] [CrossRef]
- Feldman, B.J.; Feldman, D. The Development of Androgen-Independent Prostate Cancer. Nat. Rev. Cancer 2001, 1, 34–45. [Google Scholar] [CrossRef]
- Beach, S.; Tang, H.; Park, S.; Dhillon, A.S.; Keller, E.T.; Kolch, W.; Yeung, K.C. Snail Is a Repressor of RKIP Transcription in Metastatic Prostate Cancer Cells. Oncogene 2008, 27, 2243–2248. [Google Scholar] [CrossRef]
- Lee, J.J.; Lee, J.J.; Farquhar, K.S.; Yun, J.; Frankenberger, C.A.; Bevilacqua, E.; Yeung, K.; Kim, E.-J.; Balázsi, G.; Rosner, M.R. Network of Mutually Repressive Metastasis Regulators Can Promote Cell Heterogeneity and Metastatic Transitions. Proc. Natl. Acad. Sci. USA 2014, 111, E364–E373. [Google Scholar] [CrossRef]
- Juven-Gershon, T.; Kadonaga, J.T. Regulation of Gene Expression via the Core Promoter and the Basal Transcriptional Machinery. Dev. Biol. 2010, 339, 225–229. [Google Scholar] [CrossRef]
- Datar, I.; Tegegne, H.; Qin, K.; Al-Mulla, F.; Bitar, M.S.; Trumbly, R.J.; Yeung, K.C. Genetic and Epigenetic Control of RKIP Transcription. Crit. Rev. Oncog. 2014, 19, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Frankenberger, C.A.; Kuo, W.L.; Boelens, M.C.; Eves, E.M.; Cheng, N.; Liang, H.; Li, W.H.; Ishwaran, H.; Minn, A.J.; et al. Signalling Pathway for RKIP and Let-7 Regulates and Predicts Metastatic Breast Cancer. EMBO J. 2011, 30, 4500–4514. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Park, S.; Sun, S.C.; Trumbly, R.; Ren, G.; Tsung, E.; Yeung, K.C. RKIP Inhibits NF-ΚB in Cancer Cells by Regulating Upstream Signaling Components of the IκB Kinase Complex. FEBS Lett. 2010, 584, 662–668. [Google Scholar] [CrossRef]
- Dangi-Garimella, S.; Yun, J.; Eves, E.M.; Newman, M.; Erkeland, S.J.; Hammond, S.M.; Minn, A.J.; Rosner, M.R. Raf Kinase Inhibitory Protein Suppresses a Metastasis Signalling Cascade Involving LIN28 and Let-7. EMBO J. 2009, 28, 347–358. [Google Scholar] [CrossRef]
- Yook, J.I.; Li, X.Y.; Ota, I.; Hu, C.; Kim, H.S.; Kim, N.H.; Cha, S.Y.; Ryu, J.K.; Choi, Y.J.; Kim, J.; et al. A Wnt–Axin2–GSK3β Cascade Regulates Snail1 Activity in Breast Cancer Cells. Nat. Cell Biol. 2006, 8, 1398–1406. [Google Scholar] [CrossRef]
- Bonavida, B.; Baritaki, S. Dual Role of NO Donors in the Reversal of Tumor Cell Resistance and EMT: Downregulation of the NF-ΚB/Snail/YY1/RKIP Circuitry. Nitric Oxide 2011, 24, 1–7. [Google Scholar] [CrossRef]
- Das, S.K.; Bhutia, S.K.; Sokhi, U.K.; Azab, B.; Su, Z.; Boukerche, H.; Anwar, T.; Moen, E.L.; Chatterjee, D.; Pellecchia, M.; et al. Raf Kinase Inhibitor RKIP Inhibits MDA-9/Syntenin-Mediated Metastasis in Melanoma. Cancer Res. 2012, 72, 6217–6226. [Google Scholar] [CrossRef]
- Du, Y.; Liu, X.; Zhu, H.; Wang, L.; Ning, J.; Xiao, C. MiR-543 Promotes Proliferation and Epithelial-Mesenchymal Transition in Prostate Cancer via Targeting RKIP. Cell. Physiol. Biochem. 2017, 41, 1135–1146. [Google Scholar] [CrossRef]
- Du, Y.; Weng, X.D.; Wang, L.; Liu, X.H.; Zhu, H.C.; Guo, J.; Ning, J.Z.; Xiao, C.C. LncRNA XIST Acts as a Tumor Suppressor in Prostate Cancer through Sponging MiR-23a to Modulate RKIP Expression. Oncotarget 2017, 8, 94358–94370. [Google Scholar] [CrossRef]
- Huang, L.; Dai, T.; Lin, X.; Zhao, X.; Chen, X.; Wang, C.; Li, X.; Shen, H.; Wang, X. MicroRNA-224 Targets RKIP to Control Cell Invasion and Expression of Metastasis Genes in Human Breast Cancer Cells. Biochem. Biophys. Res. Commun. 2012, 425, 127–133. [Google Scholar] [CrossRef]
- Cross-Knorr, S.; Lu, S.; Perez, K.; Guevara, S.; Brilliant, K.; Pisano, C.; Quesenberry, P.J.; Resnick, M.B.; Chatterjee, D. RKIP Phosphorylation and STAT3 Activation Is Inhibited by Oxaliplatin and Camptothecin and Are Associated with Poor Prognosis in Stage II Colon Cancer Patients. BMC Cancer 2013, 13, 463. [Google Scholar] [CrossRef] [PubMed]
- Papale, M.; Vocino, G.; Lucarelli, G.; Rutigliano, M.; Gigante, M.; Rocchetti, M.T.; Pesce, F.; Sanguedolce, F.; Bufo, P.; Battaglia, M.; et al. Urinary RKIP/p-RKIP Is a Potential Diagnostic and Prognostic Marker of Clear Cell Renal Cell Carcinoma. Oncotarget 2017, 8, 40412–40424. [Google Scholar] [CrossRef] [PubMed]
- Schoentgen, F.; Jonic, S. PEBP1/RKIP Behavior: A Mirror of Actin-Membrane Organization. Cell. Mol. Life Sci. 2020, 77, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Schoentgen, F. Insight on the Role of RKIP in Cancer through Key Protein Partners and Cellular Protrusions. In Prognostic and Therapeutic Applications of RKIP in Cancer; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–35. [Google Scholar]
- Netti, G.S.; Lucarelli, G.; Spadaccino, F.; Castellano, G.; Gigante, M.; Divella, C.; Rocchetti, M.T.; Rascio, F.; Mancini, V.; Stallone, G.; et al. PTX3 Modulates the Immunoflogosis in Tumor Microenvironment and Is a Prognostic Factor for Patients with Clear Cell Renal Cell Carcinoma. Aging 2020, 12, 7585. [Google Scholar] [CrossRef] [PubMed]
- Rascio, F.; Spadaccino, F.; Rocchetti, M.T.; Castellano, G.; Stallone, G.; Netti, G.S.; Ranieri, E. The Pathogenic Role of PI3K/AKT Pathway in Cancer Onset and Drug Resistance: An Updated Review. Cancers 2021, 13, 3949. [Google Scholar] [CrossRef] [PubMed]
- Stallone, G.; Netti, G.S.; Cormio, L.; Castellano, G.; Infante, B.; Pontrelli, P.; Divella, C.; Selvaggio, O.; Spadaccino, F.; Ranieri, E.; et al. Modulation of Complement Activation by Pentraxin-3 in Prostate Cancer. Sci. Rep. 2020, 10, 18400. [Google Scholar] [CrossRef] [PubMed]
- Stallone, G.; Cormio, L.; Netti, G.S.; Infante, B.; Selvaggio, O.; di Fino, G.; Ranieri, E.; Bruno, F.; Prattichizzo, C.; Sanguedolce, F.; et al. Pentraxin 3: A Novel Biomarker for Predicting Progression from Prostatic Inflammation to Prostate Cancer. Cancer Res. 2014, 74, 4230–4238. [Google Scholar] [CrossRef]
- Deleonardis, A.; Papale, M. Methods to Study Posttranslational Modification Patterns in Cytotoxic T-Cells and Cancer. In Cytotoxic T-Cells. Methods in Molecular Biology; Gigante, M., Ranieri, E., Eds.; Humana: New York, NY, USA, 2020; Volume 2325. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef]
- Black, R.C.; Khurshid, H. NSCLC: An Update of Driver Mutations, Their Role in Pathogenesis and Clinical Significance. Rhode Isl. Med. J. 2015, 98, 25–28. [Google Scholar]
- Wang, Q.; Wu, X.; Wu, T.; Li, G.; Shi, Y. Clinical Significance of RKIP MRNA Expression in Non-Small Cell Lung Cancer. Tumor Biol. 2014, 35, 4377–4380. [Google Scholar] [CrossRef]
- Wang, A.; Duan, G.; Zhao, C.; Gao, Y.; Liu, X.; Wang, Z.; Li, W.; Wang, K.; Wang, W. Reduced RKIP Expression Levels Are Associated with Frequent Non-Small Cell Lung Cancer Metastasis and STAT3 Phosphorylation and Activation. Oncol. Lett. 2017, 13, 3039–3045. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Ji, R.; Laber, D.A.; Yan, X.; Xu, X. Association of Low RKIP Expression with Poor Prognosis in Non-Small Cell Lung Cancer. J. Clin. Oncol. 2021, 39, 3068. [Google Scholar] [CrossRef]
- Xie, S.Y.; Li, G.; Han, C.; Yu, Y.Y.; Li, N. RKIP Reduction Enhances Radioresistance by Activating the Shh Signaling Pathway in Non-Small-Cell Lung Cancer. Onco Targets Ther. 2017, 10, 5605–5619. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Zhang, Z.; Zhang, Z.; Li, J.Y.; Cui, J.; Shi, W.P.; Dong, X.W.; Yuan, L.; Lin, P.; Chen, Z.N.; et al. Aberrant Expression of MiR-362 Promotes Lung Cancer Metastasis through Downregulation of Sema3A. J. Immunol. Res. 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- Wang, S.; Jiang, W.; Zhang, X.; Lu, Z.; Geng, Q.; Wang, W.; Li, N.; Cai, X. LINC-PINT Alleviates Lung Cancer Progression via Sponging MiR-543 and Inducing PTEN. Cancer Med. 2020, 9, 1999–2009. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhang, X.; Lin, Z.; Xiong, A.; Xie, S.; Liang, J.; Zhang, W. SFTA1P, LINC00968, GATA6-AS1, TBX5-AS1, and FEZF1-AS1 Are Crucial Long Non-coding RNAs Associated with the Prognosis of Lung Squamous Cell Carcinoma. Oncol. Lett. 2019, 18, 3985–3993. [Google Scholar] [CrossRef]
- Gong, Z.; Chen, X.; Zhang, Y.; Liu, C.; Wang, Z.; Xu, X.; Zhu, J.; Xue, T. LncRNA GATA6-AS1 Inhibits the Progression of Non-Small Cell Lung Cancer via Repressing MicroRNA-543 to Up-Regulating RKIP. Cancer Manag. Res. 2020, 12, 9327–9338. [Google Scholar] [CrossRef]
- Li, H.; Ouyang, R.; Wang, Z.; Zhou, W.; Chen, H.; Jiang, Y.; Zhang, Y.; Li, H.; Liao, M.; Wang, W.; et al. MiR-150 Promotes Cellular Metastasis in Non-Small Cell Lung Cancer by Targeting FOXO4. Sci. Rep. 2016, 6, 39001. [Google Scholar] [CrossRef]
- Raquel-Cunha, A.; Cardoso-Carneiro, D.; Reis, R.M.; Martinho, O. Current Status of Raf Kinase Inhibitor Protein (RKIP) in Lung Cancer: Behind RTK Signaling. Cells 2019, 8, 442. [Google Scholar] [CrossRef]
- Huerta-Yepez, S.; Yoon, N.K.; Hernandez-Cueto, A.; Mah, V.; Rivera-Pazos, C.M.; Chatterjee, D.; Vega, M.I.; Maresh, E.L.; Horvath, S.; Chia, D.; et al. Expression of Phosphorylated Raf Kinase Inhibitor Protein (PRKIP) Is a Predictor of Lung Cancer Survival. BMC Cancer 2011, 11, 259. [Google Scholar] [CrossRef]
- Albano, G.D.; Bonanno, A.; Moscato, M.; Anzalone, G.; di Sano, C.; Riccobono, L.; Wenzel, S.E.; Profita, M. Crosstalk between MAChRM3 and Β2AR, via Acetylcholine PI3/PKC/PBEP1/Raf-1 MEK1/2/ERK1/2 Pathway Activation, in Human Bronchial Epithelial Cells after Long-Term Cigarette Smoke Exposure. Life Sci. 2018, 192, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Minoo, P.; Zlobec, I.; Baker, K.; Tornillo, L.; Terracciano, L.; Jass, J.R.; Lugli, A. Loss of Raf-1 Kinase Inhibitor Protein Expression Is Associated with Tumor Progression and Metastasis in Colorectal Cancer. Am. J. Clin. Pathol. 2007, 127, 820–827. [Google Scholar] [CrossRef]
- Al-Mulla, F.; Hagan, S.; Behbehani, A.I.; Bitar, M.S.; George, S.S.; Going, J.J.; García, J.J.C.; Scott, L.; Fyfe, N.; Murray, G.I.; et al. Raf Kinase Inhibitor Protein Expression in a Survival Analysis of Colorectal Cancer Patients. J. Clin. Oncol. 2006, 24, 5672–5679. [Google Scholar] [CrossRef] [PubMed]
- Nie, F.; Cao, J.; Tong, J.; Zhu, M.; Gao, Y.; Ran, Z. Role of Raf-Kinase Inhibitor Protein in Colorectal Cancer and Its Regulation by Hydroxycamptothecine. J. Biomed. Sci. 2015, 22, 56. [Google Scholar] [CrossRef] [PubMed]
- Oberg, A.L.; French, A.J.; Sarver, A.L.; Subramanian, S.; Morlan, B.W.; Riska, S.M.; Borralho, P.M.; Cunningham, J.M.; Boardman, L.A.; Wang, L.; et al. MiRNA Expression in Colon Polyps Provides Evidence for a Multihit Model of Colon Cancer. PLoS ONE 2011, 6, e20465. [Google Scholar] [CrossRef]
- Bartley, A.N.; Yao, H.; Barkoh, B.A.; Ivan, C.; Mishra, B.M.; Rashid, A.; Calin, G.A.; Luthra, R.; Hamilton, S.R. Complex Patterns of Altered MicroRNA Expression during the Adenoma-Adenocarcinoma Sequence for Microsatellite-Stable Colorectal Cancer. Clin. Cancer Res. 2011, 17, 7283–7293. [Google Scholar] [CrossRef]
- Shirjang, S.; Mansoori, B.; Mohammadi, A.; Shajari, N.; Duijf, P.H.; Najafi, S.; Abedi Gaballu, F.; Nofouzi, K.; Baradaran, B. MiR-330 Regulates Colorectal Cancer Oncogenesis by Targeting BACH1. Adv. Pharm. Bull. 2020, 10, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Baritaki, S.; Marathe, H.; Feng, J.; Park, S.; Beach, S.; Bazeley, P.S.; Beshir, A.B.; Fenteany, G.; Mehra, R.; et al. Polycomb Protein EZH2 Regulates Tumor Invasion via the Transcriptional Repression of the Metastasis Suppressor RKIP in Breast and Prostate Cancer. Cancer Res. 2012, 72, 3091–3104. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Wu, H.; Fu, F.; Yi, W.; Pei, L.; Zhou, M. RKIP Suppresses the Proliferation and Metastasis of Breast Cancer Cell Lines through Up-Regulation of MiR-185 Targeting HMGA2. Arch. Biochem. Biophys. 2016, 610, 25–32. [Google Scholar] [CrossRef]
- Al-Mulla, F.; Bitar, M.S.; Thiery, J.P.; Zea, T.T.; Chatterjee, D.; Bennett, L.; Park, S.; Edwards, J.; Yeung, K.C. Clinical Implications for Loss or Diminution of Expression of Raf-1 Kinase Inhibitory Protein and Its Phosphorylated Form in Ductal Breast Cancer. Am. J. Cancer Res. 2013, 3, 446–464. [Google Scholar]
- Zebisch, A.; Caraffini, V.; Sill, H. RAF Kinase Inhibitor Protein in Myeloid Leukemogenesis. Int. J. Mol. Sci. 2019, 20, 5756. [Google Scholar] [CrossRef] [PubMed]
- Caraffini, V.; Perfler, B.; Berg, J.L.; Uhl, B.; Schauer, S.; Kashofer, K.; Ghaffari-Tabrizi-Wizsy, N.; Strobl, H.; Wölfler, A.; Hoefler, G.; et al. Loss of RKIP Is a Frequent Event in Myeloid Sarcoma and Promotes Leukemic Tissue Infiltration. Blood 2018, 131, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Hatzl, S.; Geiger, O.; Kuepper, M.K.; Caraffini, V.; Seime, T.; Furlan, T.; Nussbaumer, E.; Wieser, R.; Pichler, M.; Scheideler, M.; et al. Increased Expression of MiR-23a Mediates a Loss of Expression in the RAF Kinase Inhibitor Protein RKIP. Cancer Res. 2016, 76, 3644–3654. [Google Scholar] [CrossRef] [PubMed]
- Baritaki, S.; Huerta-Yepez, S.; da Cabrava-Haimandez, M.L.; Sensi, M.; Canevari, S.; Libra, M.; Penichet, M.; Chen, H.; Berenson, J.R.; Bonavida, B. Unique Pattern of Overexpression of Raf-1 Kinase Inhibitory Protein in Its Inactivated Phosphorylated Form in Human Multiple Myeloma. Immunopathol. Dis. Ther. 2011, 2, 179–188. [Google Scholar] [CrossRef]
- Huerta-Yepez, S.; Liu, H.; Baritaki, S.; del Lourdes Cebrera-Munoz, M.; Rivera-Pazos, C.; Maldonado-Valenzuela, A.; Valencia-Hipolito, A.; Vega, M.I.; Chen, H.; Berenson, J.R.; et al. Overexpression of Yin Yang 1 in Bone Marrow-Derived Human Multiple Myeloma and Its Clinical Significance. Int. J. Oncol. 2014, 45, 1184–1192. [Google Scholar] [CrossRef]
- Penas, C.; Apraiz, A.; Muñoa, I.; Arroyo-Berdugo, Y.; Rasero, J.; Ezkurra, P.A.; Velasco, V.; Subiran, N.; Bosserhoff, A.K.; Alonso, S.; et al. RKIP Regulates Differentiation-Related Features in Melanocytic Cells. Cancers 2020, 12, 1451. [Google Scholar] [CrossRef] [PubMed]
- Cardile, V.; Malaponte, G.; Loreto, C.; Libra, M.; Caggia, S.; Trovato, F.M.; Musumeci, G. Raf Kinase Inhibitor Protein (RKIP) and Phospho-RKIP Expression in Melanomas. Acta Histochem. 2013, 115, 795–802. [Google Scholar] [CrossRef]
- Molofsky, A.V.; Pardal, R.; Morrison, S.J. Diverse Mechanisms Regulate Stem Cell Self-Renewal. Curr. Opin. Cell Biol. 2004, 16, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Ohnishi, Y.; Inoue, H.; Wato, M.; Tanaka, A.; Kakudo, K.; Nozaki, M. NANOG Expression Correlates with Differentiation, Metastasis and Resistance to Preoperative Adjuvant Therapy in Oral Squamous Cell Carcinoma. Oncol. Lett. 2014, 7, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Sun, L.; Jiang, K.; Gao, D.M.; Kang, X.N.; Wang, C.; Zhang, S.; Huang, S.; Qin, X.; Li, Y.; et al. NANOG Promotes Liver Cancer Cell Invasion by Inducing Epithelial–Mesenchymal Transition through NODAL/SMAD3 Signaling Pathway. Int. J. Biochem. Cell Biol. 2013, 45, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Hill, B.; de Melo, J.; Yan, J.; Kapoor, A.; He, L.; Cutz, J.C.; Feng, X.; Bakhtyar, N.; Tang, D. Common Reduction of the Raf Kinase Inhibitory Protein in Clear Cell Renal Cell Carcinoma. Oncotarget 2014, 5, 7406–7419. [Google Scholar] [CrossRef] [PubMed]
- Neumann, H.P.H.; Zbar, B. Renal Cysts, Renal Cancer and von Hippel-Lindau Disease. Kidney Int. 1997, 51, 16–26. [Google Scholar] [CrossRef]
- Choyke, P.L.; Glenn, G.M.; Walther, M.M.; Zbar, B.; Linehan, W.M. Hereditary Renal Cancers. Radiology 2003, 226, 33–46. [Google Scholar] [CrossRef]
- Bedri, S.K.; Nilsson, O.B.; Fink, K.; Månberg, A.; Hamsten, C.; Ayoglu, B.; Manouchehrinia, A.; Nilsson, P.; Olsson, T.; Hillert, J.; et al. Plasma Protein Profiling Reveals Candidate Biomarkers for Multiple Sclerosis Treatment. PLoS ONE 2019, 14, e0217208. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papale, M.; Netti, G.S.; Stallone, G.; Ranieri, E. Understanding Mechanisms of RKIP Regulation to Improve the Development of New Diagnostic Tools. Cancers 2022, 14, 5070. https://doi.org/10.3390/cancers14205070
Papale M, Netti GS, Stallone G, Ranieri E. Understanding Mechanisms of RKIP Regulation to Improve the Development of New Diagnostic Tools. Cancers. 2022; 14(20):5070. https://doi.org/10.3390/cancers14205070
Chicago/Turabian StylePapale, Massimo, Giuseppe Stefano Netti, Giovanni Stallone, and Elena Ranieri. 2022. "Understanding Mechanisms of RKIP Regulation to Improve the Development of New Diagnostic Tools" Cancers 14, no. 20: 5070. https://doi.org/10.3390/cancers14205070
APA StylePapale, M., Netti, G. S., Stallone, G., & Ranieri, E. (2022). Understanding Mechanisms of RKIP Regulation to Improve the Development of New Diagnostic Tools. Cancers, 14(20), 5070. https://doi.org/10.3390/cancers14205070