
Tumor Innervation: History, Methodologies, and Significance




Abstract
:Simple Summary
Abstract
1. Introduction
2. Historical Aspects
3. Ambiguous Evidence of Tumor Innervation
4. Methodological and Conceptual Clarifications
5. Negative Evidence of Tumor Innervation
”According to Herzog, the nerve elements of spontaneous growths are mostly to be understood as preserved, preformed structures. There is said to be a noticeable resistance of the nerve elements to the tumoral proliferation, even if they occasionally show signs of degeneration. Proliferative processes, as described by some authors, have nothing to do with the tumor itself, but are merely phenomena of regeneration as a result of the abnormal elements leading to the development of experimental tumors. Oertel described a nervous plexus of the same arrangement as the nerve connections supplying the normal organ. Adventitial plexuses attached to the great blood vessels and arranged spirally around the blood capillaries can be seen. Martynkov (sic) did not find any morphological changes in the nervous elements running along the blood vessels in the tissues immediately surrounding the tumor mass. Cailliau believes that the tissue of malignant tumors possesses a special nervous system which is intimately related to the stage of development of the tumor and should be regarded as nervous stroma (neurostroma). The nervous system of the vessel walls is said to be quite different from the normal nervous supply. The innervation of the tumor tissue is more plentiful than in normal similar territories. The more malignant the neoplasm, the greater the wealth of nervous elements. With Bordallo (1948) I have described the changes in the autonomic terminal formation in rectal cancer. Between the cancer cells the nervous syncytial elements have completely perished. Changes in form, structure, and color of nervous cords occur in the immediate vicinity of the tumor. These are alterations that are not exclusively dependent on tumor proliferation but also—and perhaps mainly—on inflammatory infiltration”.
6. Positive Evidence of Tumor Innervation
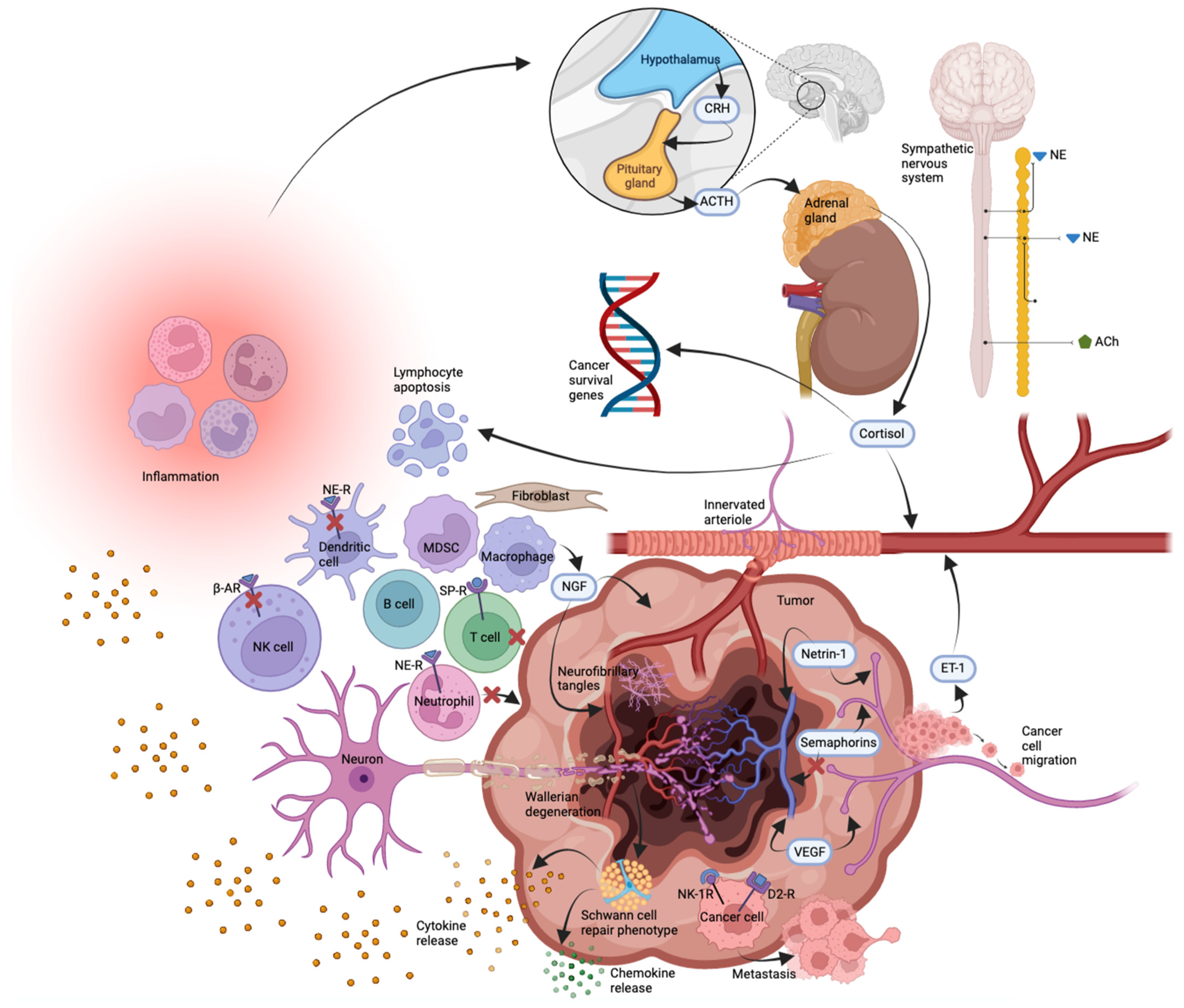
7. Neuroimmunooncology
8. Regulatory Mechanisms for Cancer-Related Neurogenesis
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Valtorta, F.; Leoni, C. Molecular mechanisms of neurite extension. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1999, 354, 387–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobling, P.; Pundavela, J.; Oliveira, S.M.R.; Roselli, S.; Walker, M.M.; Hondermarck, H. Nerve-Cancer Cell Cross-talk: A Novel Promoter of Tumor Progression. Cancer Res. 2015, 75, 1777–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, H.H. On the Presence of Nerves in Tumors and of Other Structures in Them as Revealed by a Modification of Ehrlich’s Method of “Vital Staining” with Methylene Blue. J. Exp. Med. 1897, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martynow, W. Verhalten der peripheren Nerven zum Plattenepthelkrebs des Menschen. Virchows. Arch. Pathol. Anat. Physiol. Klin. Med. 1930, 278, 498–517. [Google Scholar] [CrossRef]
- Argaud, M.R. Sur les terminaisons nerveuses dans les cancers humains. Comptes R. Acad. Hebd. Seances Acad. Sci. D 1925, T180, 551–554. [Google Scholar]
- Itchikawa, K. Sur l’existence d’une relation entre les nerfs periphériques et le développement du cancer. Bull. Assoc. Franç. Étude Canc. 1928, 18, 455–489. [Google Scholar]
- Oertel, H. Innervation and tumour growth: A preliminary report. Can. Med. Assoc. 1928, 18, 135–139. [Google Scholar]
- Shapiro, D.M.; Warren, S. Cancer Innervation. Cancer Res. 1949, 9, 707–711. [Google Scholar]
- Zhou, M.; Patel, A.; Rubin, M.A. Prevalence and location of peripheral nerve found on prostate needle biopsy. Am. J. Clin. Pathol. 2001, 115, 39–43. [Google Scholar] [CrossRef]
- Seifert, P.; Spitznas, M. Tumours May Be Innervated. Virchows Arch. 2001, 438, 228–231. [Google Scholar] [CrossRef]
- Cain, D.M.; Wacnik, P.W.; Turner, M.; Wendelschafer-Crabb, G.; Kennedy, W.R.; Wilcox, G.L.; Simone, D.A. Functional Interactions between Tumor and Peripheral Nerve: Changes in Excitability and Morphology of Primary Afferent Fibers in a Murine Model of Cancer Pain. J. Neurosci. 2001, 21, 9367–9376. [Google Scholar] [CrossRef] [PubMed]
- Wacnik, P.W.; Eikmeier, L.J.; Ruggles, T.R.; Ramnaraine, M.L.; Walcheck, B.K.; Beitz, A.J.; Wilcox, G.L. Functional Interactions between Tumor and Peripheral Nerve: Morphology, Algogen Identification, and Behavioral Characterization of a New Murine Model of Cancer Pain. J. Neurosci. 2001, 21, 9355–9366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, G.E.; Wheeler, T.M.; Shine, H.D.; Schmelz, M.; Frolov, A.; Chakraborty, S.; Rowley, D. In vitro dorsal root ganglia and human prostate cell line interaction: Redefining perineural invasion in prostate cancer. Prostate 2001, 49, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Seifert, P.; Benedic, M.; Effert, P. Nerve Fibers in Tumors of the Human Urinary Bladder. Virchows Arch. 2002, 440, 291–297. [Google Scholar] [CrossRef]
- Lü, S.-H.; Zhou, Y.; Que, H.-P.; Liu, S.-J. Peptidergic Innervation of Human Esophageal and Cardiac Carcinoma. World J. Gastroenterol. 2003, 9, 399–403. [Google Scholar] [CrossRef]
- Wacnik, P.W.; Baker, C.M.; Herron, M.J.; Kren, B.T.; Blazar, B.R.; Wilcox, G.L.; Hordinsky, M.K.; Beitz, A.J.; Ericson, M.E. Tumor-induced mechanical hyperalgesia involves CGRP receptors and altered innervation and vascularization of DsRed2 fluorescent hindpaw tumors. Pain 2005, 115, 95–106. [Google Scholar] [CrossRef]
- Lindsay, T.H.; Jonas, B.M.; Sevcik, M.A.; Kubota, K.; Halvorson, K.G.; Ghilardi, J.R.; Kuskowski, M.A.; Stelow, E.B.; Mukherjee, P.; Gendler, S.J.; et al. Pancreatic cancer pain and its correlation with changes in tumor vasculature, macrophage infiltration, neuronal innervation, body weight and disease progression. Pain 2005, 119, 233–246. [Google Scholar] [CrossRef]
- Ayala, G.E.; Dai, H.; Powell, M.; Li, R.; Ding, Y.; Wheeler, T.M.; Shine, D.; Kadmon, D.; Thompson, T.; Miles, B.J.; et al. Cancer-related axonogenesis and neurogenesis in prostate cancer. Clin. Cancer Res. 2008, 14, 7593–7603. [Google Scholar] [CrossRef] [Green Version]
- Albo, D.; Akay, C.L.; Marshall, C.L.; Wilks, J.A.; Verstovsek, G.; Liu, H.; Agarwal, N.; Berger, D.H.; Ayala, G.E. Neurogenesis in colorectal cancer is a marker of aggressive tumor behavior and poor outcomes. Cancer 2011, 117, 4834–4845. [Google Scholar] [CrossRef]
- Sone, Y.; Takatori, S.; Ochi, E.; Zamani, Y.; Matsuyama, A.; Fukuhara, S.; Goda, M.; Kitamura, Y.; Kawasaki, H. Nerve Growth Factor Facilitates the Innervation of Perivascular Nerves in Tumor-Derived Neovasculature in the Mouse Cornea. Pharmacology. 2017, 99, 57–66. [Google Scholar] [CrossRef]
- Wang, W.; Li, L.; Chen, N.; Niu, C.; Li, Z.; Hu, J.; Cui, J. Nerves in the Tumor Microenvironment: Origin and Effects. Front. Cell. Dev. Biol. 2020, 8, 601738. [Google Scholar] [CrossRef] [PubMed]
- Magnon, C.; Hall, S.J.; Lin, J.; Xue, X.; Gerber, L.; Freedland, S.J.; Frenette, P.S. Autonomic nerve development contributes to prostate cancer progression. Science 2013, 341, 1236361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnon, C. Role of the autonomic nervous system in tumorigenesis and metastasis. Mol. Cell. Oncol. 2015, 2, e975643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, S.W.; Nagaraja, A.S.; Lutgendorf, S.K.; Green, P.A.; Sood, A.K. Sympathetic nervous system regulation of the tumour microenvironment. Nat. Rev. Cancer. 2015, 15, 563–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saloman, J.L.; Albers, K.M.; Rhim, A.D.; Davis, B.M. Can Stopping Nerves, Stop Cancer? Trends Neurosci. 2016, 39, 880–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuol, N.; Stojanovska, L.; Apostolopoulos, V.; Nurgali, K. Role of the nervous system in cancer metastasis. J. Exp. Clin. Cancer Res. 2018, 37, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulkner, S.; Jobling, P.; March, B.; Jiang, C.C.; Hondermarck, H. Tumor Neurobiology and the War of Nerves in Cancer. Cancer Discov. 2019, 9, 702–710. [Google Scholar] [CrossRef] [Green Version]
- Herzog, E. Beitrag zur Frage der Innervation der Geschwülste. Virchows Arch. Pathol. Anat. 1928, 268, 536–565. [Google Scholar] [CrossRef]
- Ryrie, G.M. On the significance of nerve fibres in human malignant neoplasms. J. Pathol. Bacteriol. 1933, 36, 13–18. [Google Scholar] [CrossRef]
- Jabonero, V. Mikroskopische Studien über die Morphologie und die Morphopatholigie der vegetativen Innervation der menschlichen Haut (II). Acta Neurovegetativa. 1958, 18, 354–387. [Google Scholar] [CrossRef]
- Mitchell, B.S.; Schumacher, U.; Kaiserling, E. Are tumours innervated? Immunohistological investigations using antibodies against the neuronal marker protein gene product 9.5 (PGP 9.5) in benign, malignant and experimental tumours. Tumour Biol. 1994, 15, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.S.; Schumacher, U.; Stauber, V.V.; Kaiserling, E. Are breast tumours innervated? Immunohistological investigations using antibodies against the neuronal marker protein gene product 9.5 (PGP 9.5) in benign and malignant breast lesions. Euro. J. Cancer 1994, 30, 1100–1103. [Google Scholar] [CrossRef]
- Terada, T.; Matsunaga, Y. S-100-positive nerve fibers in hepatocellular carcinoma and intrahepatic cholangiocarcinoma: An immunohistochemical study. Pathol. Int. 2001, 51, 89–93. [Google Scholar] [CrossRef]
- Habash, F.S.; Hantash, R.O.A.; Yunis, M.A. Assessment of the innervation pattern of oral squamous cell carcinoma using neural protein gene product (9.5)-An immunocytochemical study. J. Oral Maxillofac. Pathol. 2012, 16, 16–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, T. Localization of nerve fibers in colonic polyps, adenomas, and adenocarcinomas by immunocytochemical staining for PGP 9.5. Dig. Dis. Sci. 2012, 57, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Lücke, G.A.; Zahn, F.W. Chirurgie der Geschwülste; 1. Theil (sic): Allgemeine Geschwulstlehre; Verlag von Ferdinand Enke: Stuttgart, Germany, 1896. [Google Scholar]
- Oertel, H. On the mechanism of cancer development. Can. Med. Assoc. 1930, 23, 183–189. [Google Scholar]
- Raisman, G. An urge to explain the incomprehensible: Geoffrey Harris and the discovery of the neural control of the pituitary gland. Annu. Rev. Neurosci. 1997, 20, 533–566. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.A. The cognitive revolution: A historical perspective. Trends Cogn. Sci. 2003, 7, 141–144. [Google Scholar] [CrossRef]
- Stanford Encyclopedia of Philosophy. Cognitive Science. Available online: https://plato.stanford.edu/entries/cognitive-science/ (accessed on 20 January 2022).
- Kowal, S.J. Emotions as a cause of cancer; 18th and 19th century contributions. Psychoanal. Rev. 1955, 42, 217–227. [Google Scholar]
- Reznikoff, M. Psychological Factors in Breast Cancer: A Preliminary Study of Some Personality Trends in Patients with Cancer of the Breast. Psychosom. Med. 1955, 27, 96–108. [Google Scholar] [CrossRef]
- LeShan, L. A Basic Psychological Orientation Apparently Associated with Malignant Disease. Psychiatr. Q. 1961, 35, 314–330. [Google Scholar] [CrossRef] [PubMed]
- LeShan, L. An Emotional Life-History Pattern Associated with Neoplastic Disease. Ann. N. Y. Acad. Sci. 1966, 125, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Hitzer, B.; León-Sanz, P. The Feeling Body and Its Diseases: How Cancer Went Psychosomatic in Twentieth-Century Germany. Osiris. 2016, 31, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Ader, R.; Cohen, N. Behaviorally conditioned immunosuppression. Psychosom. Med. 1975, 37, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Renoux, G.; Biziere, K. Neocortex Lateralization of Immune Function and of the Activities of Imuthiol, a T-cell Immunopotentiator. In Psychoneuroimmunology, 2nd ed.; Ader, R., Felten, D.L., Cohen, N., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1991; pp. 127–128. [Google Scholar]
- Ramirez, A.J. Life Events and Cancer: Conceptual and Methodological Issues. In Psychosocial Oncology; Watson, M., Greer, S., Thomas, C., Eds.; Pergamon: Oxford, UK, 1988; pp. 53–60. [Google Scholar]
- Ramirez, A.J.; Craig, T.K.J.; Watson, J.P.; Fentiman, I.S.; North, W.R.S.; Rubens, R.D. Stress and relapse of breast cancer. BMJ 1989, 298, 291–293. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.B. Do hormones cause breast cancer? Cancer 1984, 53, 595–604. [Google Scholar] [CrossRef]
- Bosman, F.T.; Blankenstein, M.; Daxenbichler, G.; Falkmer, S.; Heitz, P.U.; Kracht, J. What’s new in endocrine factors of tumor growth? Pathol. Res. Pract. 1985, 180, 81–92. [Google Scholar] [CrossRef]
- Vihko, R.; Apter, D. Endogenous steroids in the pathophysiology of breast cancer. Crit. Rev. Oncol. Hematol. 1989, 9, 1–16. [Google Scholar] [CrossRef]
- Olsson, H. Reproductive events, occurring in adolescence at the time of development of reproductive organs and at the time of tumour initiation, have a bearing on growth characteristics and reproductive hormone regulation in normal and tumour tissue investigated decades later—A hypothesis. Med. Hypotheses 1989, 28, 93–97. [Google Scholar] [CrossRef]
- Morris, S.A. Origin of mutation in neoplasia. Med. Hypotheses 1988, 27, 209–214. [Google Scholar] [CrossRef]
- Clarke, R.; Dickson, R.B.; Brünner, N. The process of malignant progression in human breast cancer. Ann. Oncol. 1990, 1, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Hulka, B.S. Hormone-replacement therapy and the risk of breast cancer. CA Cancer J. Clin. 1990, 40, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.M.; Herberman, R.B.; Maluish, A.M.; Schlien, B.; Lippman, M. Prognostic risk assessment in primary breast cancer by behavioral and immunological parameters. Health Psychol. 1985, 4, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.; Herberman, R.; Lippman, M.; d’Angelo, T. Correlation of stress factors with sustained depression of natural killer cell activity and predicted prognosis in patients with breast cancer. J. Clin. Oncol. 1987, 5, 348–353. [Google Scholar] [CrossRef]
- Bovbjerg, D.H. Psychoneuroimmunology. Implications for oncology? Cancer 1991, 67, 828–832. [Google Scholar] [CrossRef]
- Cohen, S.; Rabin, B.S. Psychologic stress, immunity, and cancer. J. Natl. Cancer Inst. 1998, 90, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Heffner, K.L.; Loving, T.J.; Robles, T.F.; Kiecolt-Glaser, J.K. Examining psychosocial factors related to cancer incidence and progression: In search of the silver lining. Brain Behav. Immun. 2003, 17, S109–S111. [Google Scholar] [CrossRef]
- Costanzo, E.S.; Sood, A.K.; Lutgendorf, S.K. Biobehavioral influences on cancer progression. Immunol. Allergy Clin. N. Am. 2011, 31, 109–132. [Google Scholar] [CrossRef] [Green Version]
- Antoni, M.H.; Dhabhar, F.S. The impact of psychosocial stress and stress management on immune responses in patients with cancer. Cancer 2019, 125, 1417–1431. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Hanke, M.L.; Corona, A.W.; Powell, N.D.; Stiner, L.M.; Bailey, M.T.; Nelson, R.J.; Godbout, J.P.; Sheridan, J.F. β-Adrenergic receptor antagonism prevents anxiety-like behavior and microglial reactivity induced by repeated social defeat. J. Neurosci. 2011, 31, 6277–6288. [Google Scholar] [CrossRef] [Green Version]
- Aldea, M.; Craciun, L.; Tomuleasa, C.; Crivii, C. The role of depression and neuroimmune axis in the prognosis of cancer patients. J. BUON 2014, 19, 5–14. [Google Scholar] [PubMed]
- Antoni, M.H.; Lutgendorf, S.K.; Cole, S.W.; Dhabhar, F.S.; Sephton, S.E.; McDonald, P.G.; Stefanek, M.; Sood, A.K. The influence of bio-behavioral factors on tumour biology: Pathways and mechanisms. Nat. Rev. Cancer 2006, 6, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Raffa, R.B.; Jacoby, H.I. Endothelin-1, -2 and -3 directly and big-endothelin-1 indirectly elicit an abdominal constriction response in mice. Life Sci. 1991, 48, L85–L90. [Google Scholar] [CrossRef]
- Raffa, R.B.; Schupsky, J.J.; Jacoby, H.I. Endothelin-induced nociception in mice: Mediation by ETA and ETB receptors. J. Pharmacol. Exp. Ther. 1996, 276, 647–651. [Google Scholar] [PubMed]
- Davar, G.; Hans, G.; Fareed, M.U.; Sinnott, C.; Strichartz, G. Behavioral signs of acute pain produced by application of endothelin-1 to rat sciatic nerve. Neuroreport 1998, 9, 2279–2283. [Google Scholar] [CrossRef]
- Fareed, M.U.; Hans, G.H.; Atanda, A.; Strichartz, G.R.; Davar, G. Pharmacological characterization of acute pain behavior produced by the application of endothelin-1 to the rat sciatic nerve. J. Pain. 2000, 1, 46–53. [Google Scholar] [CrossRef]
- Piovezan, A.P.; D’Orleans-Juste, P.; Souza, G.E.; Rae, G.A. Endothelin-1-induced ET(A) receptor-mediated nociception, hyperalgesia and oedema in the mouse hind-paw: Modulation by simultaneous ET(B) receptor activation. Br. J. Pharmacol. 2000, 129, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Pomonis, J.D.; Rogers, S.D.; Peters, C.M.; Ghilardi, J.R.; Mantyh, P.W. Expression and localization of endothelin receptors: Implications for the involvement of peripheral glia in nociception. J. Neurosci. 2001, 21, 999–1006. [Google Scholar] [CrossRef]
- Gokin, A.P.; Fareed, M.U.; Pan, H.-L.; Hans, G.; Strichartz, G.R.; Davar, G. Local injection of endothelin-1 produces pain-like behavior and excitation of nociceptors in rats. J. Neurosci. 2001, 21, 5358–5366. [Google Scholar] [CrossRef] [Green Version]
- Nelson, J.B.; Hedican, S.P.; George, D.J.; Reddi, A.H.; Piantadosi, S.; Eisenberger, M.A.; Simons, J.W. Identification of endothelin-1 in the pathophysiology of metastatic adenocarcinoma of the prostate. Nat. Med. 1995, 1, 944–949. [Google Scholar] [CrossRef]
- Oaklander, A.L. The density of remaining nerve endings in human skin with and without postherpetic neuralgia after shingles. Pain 2001, 92, 139–145. [Google Scholar] [CrossRef]
- Ayala, G.E.; Dai, H.; Ittmann, M.; Li, R.; Powell, M.; Frolov, A.; Wheeler, T.M.; Thompson, T.C.; Rowley, D. Growth and survival mechanisms associated with perineural invasion in prostate cancer. Cancer Res. 2004, 64, 6082–6090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisbie, J.H.; Binard, J. Low prevalence of prostatic cancer among myelopathy patients. J. Am. Paraplegia Soc. 1994, 17, 148–149. [Google Scholar] [CrossRef]
- Frisbie, J.H. Cancer of the prostate in myelopathy patients: Lower risk with higher levels of paralysis. J. Spinal Cord Med. 2001, 24, 92–94; discussion 95. [Google Scholar] [CrossRef] [PubMed]
- Oiseth, S.J.; Aziz, M.S. Cancer immunotherapy: A brief review of the history, possibilities, and challenges ahead. J. Cancer Metastasis Treat. 2017, 3, 250–261. [Google Scholar] [CrossRef]
- Dobosz, P.; Dzieciątkowski, T. The Intriguing History of Cancer Immunotherapy. Front. Immunol. 2019, 10, 2965. [Google Scholar] [CrossRef] [Green Version]
- Busch, W. Aus der Sitzung der medicinischen Section vom 13 November 1867. Berlin Klin. Wochenschr. 1868, 5, 137. [Google Scholar]
- Fehleisen, F. Ueber die Züchtung der Erysipelkokken auf künstlichem Nährboden und ihre übertragbarkeit auf den Menschen. Dtsch. Med. Wochenschr. 1882, 8, 553–554. [Google Scholar]
- McCarthy, E.F. The toxins of William B. Coley and the treatment of bone and soft-tissue sarcomas. Iowa Orthop. J. 2006, 26, 154–158. [Google Scholar]
- Parish, C.R. Cancer immunotherapy: The past, the present and the future. Immunol. Cell Biol. 2003, 81, 106–113. [Google Scholar] [CrossRef]
- Old, L.J.; Clarke, D.A.; Benacerraf, B. Effect of Bacillus Calmette-Guérin infection on transplanted tumours in the mouse. Nature 1959, 184, 291–292. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; Eidinger, D.; Bruce, A.W. Intracavitary Bacillus Calmette-guerin in the Treatment of Superficial Bladder Tumors. J. Urol. 1976, 116, 180–183. [Google Scholar] [CrossRef]
- Vajdic, C.M.; McDonald, S.P.; McCredie, M.R.E.; van Leeuwen, M.T.; Stewart, J.H.; Law, M.; Chapman, J.R.; Webster, A.C.; Kaldor, J.M.; Grulich, A.E. Cancer incidence before and after kidney transplantation. JAMA 2006, 296, 2823–2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimmer, C.D.; Rentsch, M.; Crispin, A.; Illner, W.D.; Arbogast, H.; Graeb, C.; Jauch, K.-W.; Guba, M. The janus face of immunosuppression—De novo malignancy after renal transplantation: The experience of the Transplantation Center Munich. Kidney Int. 2007, 71, 1271–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraneveld, A.D.; de Theije, C.G.M.; van Heesch, F.; Borre, Y.; de Kivit, S.; Oliver, B.; Korte, M.; Garssen, J. The neuro-immune axis: Prospect for novel treatments for mental disorders. Basic Clin. Pharmocol. Toxicol. 2014, 114, 128–136. [Google Scholar] [CrossRef]
- Brimberg, L.; Mader, S.; Fujieda, Y.; Arinuma, Y.; Kowal, C.; Volpe, B.T.; Diamond, B. Antibodies as Mediators of Brain Pathology. Trends Immunol. 2015, 36, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune attack: The role of inflammation in Alzheimer disease. Nat. Rev. Neurosci. 2015, 16, 358–372. [Google Scholar] [CrossRef]
- Hodes, G.E.; Kana, V.; Menard, C.; Merad, M.; Russo, S.J. Neuroimmune mechanisms of depression. Nat. Neurosci. 2015, 18, 1386–1393. [Google Scholar] [CrossRef]
- Li, H.; Fan, X.; Houghton, J. Tumor microenvironment: The role of the tumor stroma in cancer. J. Cell. Biochem. 2007, 101, 805–815. [Google Scholar] [CrossRef]
- Shurin, M.R.; Shurin, G.V.; Zlotnikov, S.B.; Bunimovich, Y.L. The Neuroimmune Axis in the Tumor Microenvironment. J. Immunol. 2020, 204, 280–285. [Google Scholar] [CrossRef]
- Lang, K.; Drell, T.L.; Niggemann, B.; Zänker, K.S.; Entschladen, F. Neurotransmitters regulate the migration and cytotoxicity in natural killer cells. Immunol. Lett. 2003, 90, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.-H.; Guo, S.-Y.; Xu, S.; Yin, Q.-Z.; Ohshita, Y.; Naitoh, M.; Horibe, Y.; Hisamitsu, T. Sympathetic nervous system mediates cold stress-induced suppression of natural killer cytotoxicity in rats. Neurosci. Lett. 2004, 358, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Maestroni, G.J.M. Adrenergic modulation of dendritic cells function: Relevance for the immune homeostasis. Curr. Neurovasc. Res. 2005, 2, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Bastian, P.; Posch, B.; Lang, K.; Niggemann, B.; Zaenker, K.S.; Hatt, H.; Entschladen, F. Phosphatidylinositol 3-kinase in the G protein-coupled receptor-induced chemokinesis and chemotaxis of MDA-MB-468 breast carcinoma cells: A comparison with leukocytes. Mol. Cancer Res. 2006, 4, 411–421. [Google Scholar] [CrossRef] [Green Version]
- Levite, M. Nerve-driven immunity. The direct effects of neurotransmitters on T-cell function. Ann. N. Y. Acad. Sci. 2000, 917, 307–321. [Google Scholar] [CrossRef]
- Reiche, E.M.V.; Nunes, S.O.V.; Morimoto, H.K. Stress, depression, the immune system, and cancer. Lancet Oncol. 2004, 5, 617–625. [Google Scholar] [CrossRef]
- Ahmad, M.H.; Rizvi, M.A.; Fatima, M.; Mondal, A.C. Pathophysiological implications of neuroinflammation mediated HPA axis dysregulation in the prognosis of cancer and depression. Mol. Cell. Endocrinol. 2021, 520, 111093. [Google Scholar] [CrossRef]
- Gasparini, G.; Longo, R.; Toi, M.; Ferrara, N. Angiogenic inhibitors: A new therapeutic strategy in oncology. Nat. Clin. Pract. Oncol. 2005, 2, 562–577. [Google Scholar] [CrossRef]
- Entschladen, F.; Palm, D.; Lang, K.; Drell IV, T.L.; Zaenker, K.S. Neoneurogenesis: Tumors may initiate their own innervation by the release of neurotrophic factors in analogy to lymphangiogenesis and neoangiogenesis. Med. Hypotheses 2006, 67, 33–35. [Google Scholar] [CrossRef]
- Entschladen, F.; Palm, D.; Niggemann, B.; Zaenker, K.S. The cancer’s nervous tooth: Considering the neuronal crosstalk within tumors. Semin. Cancer Biol. 2008, 18, 171–175. [Google Scholar] [CrossRef]
- Strell, C.; Entschladen, F. Extravasation of leukocytes in comparison to tumor cells. Cell. Commun. Signal. 2008, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Schuller, H.M. Is cancer triggered by altered signalling of nicotinic acetylcholine receptors? Nat. Rev. Cancer. 2009, 9, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Voss, M.J.; Niggemann, B.; Zänker, K.S.; Entschladen, F. PC-3 prostate carcinoma cells release signal substances that influence the migratory activity of cells in the tumor’s microenvironment. Cell Commun. Signal. 2010, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Patani, N.; Jiang, W.G.; Mokbel, K. Brain-derived neurotrophic factor expression predicts adverse pathological & clinical outcomes in human breast cancer. Cancer Cell Int. 2011, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Demir, I.E.; Friess, H.; Ceyhan, G.O. Nerve-cancer interactions in the stromal biology of pancreatic cancer. Front. Physiol. 2012, 3, 97. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Sun, Y.; Gao, D. Role of the nervous system in cancer metastasis. Oncol. Lett. 2013, 5, 1101–1111. [Google Scholar] [CrossRef] [Green Version]
- Fink, D.M.; Connor, A.L.; Kelley, P.M.; Steele, M.M.; Hollingsworth, M.A.; Tempero, R.M. Nerve growth factor regulates neurolymphatic remodeling during corneal inflammation and resolution. PLoS ONE 2014, 9, e112737. [Google Scholar] [CrossRef] [Green Version]
- Colucci, R.; Moretti, S. The role of stress and beta-adrenergic system in melanoma: Current knowledge and possible therapeutic options. J. Cancer Res. Clin. Oncol. 2016, 142, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Nogueira, P.; Bragado, P.; Almendro, V.; Ametller, E.; Rios, J.; Choudhury, S.; Mancino, M.; Gascón, P. Differential expression of neurogenes among breast cancer subtypes identifies high risk patients. Oncotarget 2016, 7, 5313–5326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lolas, G.; Bianchi, A.; Syrigos, K.N. Tumour-induced neoneurogenesis and perineural tumour growth: A mathematical approach. Sci. Rep. 2016, 6, 20684. [Google Scholar] [CrossRef] [PubMed]
- Saloman, J.L.; Albers, K.M.; Li, D.; Hartman, D.J.; Crawford, H.C.; Muha, E.A.; Rhim, A.D.; Davis, B.M. Ablation of sensory neurons in a genetic model of pancreatic ductal adenocarcinoma slows initiation and progression of cancer. Proc. Natl. Acad. Sci. USA 2016, 113, 3078–3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y. The Oncogenic Functions of Nicotinic Acetylcholine Receptors. J. Oncol. 2016, 2016, 9650481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeo, M.; Colbert, P.L.; Vermeer, D.W.; Lucido, C.T.; Cain, J.T.; Vichaya, E.G.; Grossberg, A.J.; Muirhead, D.; Rickel, A.P.; Hong, Z.; et al. Cancer exosomes induce tumor innervation. Nat. Commun. 2018, 9, 4284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Zhang, G.N.; Shi, Y.; Cui, L.; Leng, X.F.; Huang, J.M. Perineural invasion in cervical cancer: Pay attention to the indications of nerve-sparing radical hysterectomy. Ann. Transl. Med. 2019, 7, 203. [Google Scholar] [CrossRef] [PubMed]
- Demidov, V.; Matveev, L.A.; Demidova, O.; Matveyev, A.L.; Zaitsev, V.Y.; Flueraru, C.; Vitkin, I.A. Analysis of low-scattering regions in optical coherence tomography: Applications to neurography and lymphangiography. Biomed. Opt. Express. 2019, 10, 4207–4219. [Google Scholar] [CrossRef]
- Jiang, S.H.; Hu, L.P.; Wang, X.; Li, J.; Zhang, Z.G. Neurotransmitters: Emerging targets in cancer. Oncogene 2020, 39, 503–515. [Google Scholar] [CrossRef]
- Chen, L.; Lin, J.; Chen, L.Z.; Chen, Y.; Wang, X.J.; Guo, Z.Q.; Yu, J.M. Perineural Invasion and Postoperative Complications are Independent Predictors of Early Recurrence and Survival Following Curative Resection of Gastric Cancer. Cancer Manag. Res. 2020, 12, 7601–7610. [Google Scholar] [CrossRef]
- McCallum, G.A.; Shiralkar, J.; Suciu, D.; Covarrubias, G.; Yu, J.S.; Karathanasis, E.; Durand, D.M. Chronic neural activity recorded within breast tumors. Sci. Rep. 2020, 10, 14824. [Google Scholar] [CrossRef]
- Hodo, T.W.; de Aquino, M.T.P.; Shimamoto, A.; Shanker, A. Critical Neurotransmitters in the Neuroimmune Network. Front. Immunol. 2020, 11, 1869. [Google Scholar] [CrossRef]
- Mehedințeanu, A.M.; Sfredel, V.; Stovicek, P.O.; Schenker, M.; Târtea, G.C.; Istrătoaie, O.; Ciurea, A.M.; Vere, C.C. Assessment of Epinephrine and Norepinephrine in Gastric Carcinoma. Int. J. Mol. Sci. 2021, 22, 2042. [Google Scholar] [CrossRef]
- Dlamini, Z.; Mathabe, K.; Padayachy, L.; Marima, R.; Evangelou, G.; Syrigos, K.N.; Bianchi, A.; Lolas, G.; Hull, R. Many Voices in a Choir: Tumor-Induced Neurogenesis and Neuronal Driven Alternative Splicing Sound Like Suspects in Tumor Growth and Dissemination. Cancers 2021, 13, 2138. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, H.; Gan, Y.; Tu, H. Shedding Light on the Role of Neurotransmitters in the Microenvironment of Pancreatic Cancer. Front. Cell Dev. Biol. 2021, 9, 688953. [Google Scholar] [CrossRef]
- Wakiya, T.; Ishido, K.; Kimura, N.; Nagase, H.; Yoshizawa, T.; Morohashi, S.; Fujita, H.; Kanda, T.; Tatara, Y.; Saruwatari, J.; et al. Eukaryotic initiation factor 2 signaling behind neural invasion linked with lymphatic and vascular invasion in pancreatic cancer. Sci. Rep. 2021, 11, 21197. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.K.; Thomas, J.R.; Lamps, L.W.; Smoller, B.R.; Folpe, A.L. Protein Gene Product 9.5 (PGP 9.5) Is Not a Specific Marker of Neural and Nerve Sheath Tumors: An Immunohistochemical Study of 95 Mesenchymal Neoplasms. Mod. Pathol. 2003, 16, 963–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dollé, J.-P.; Rezvan, A.; Allen, F.D.; Lazarovici, P.; Lelkes, P.I. Nerve growth factor-induced migration of endothelial cells. J. Pharmacol. Exp. Ther. 2005, 315, 1220–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.W.; Crouse, D.; Lee, M.; Karnik, S.K.; Sorensen, L.K.; Murphy, K.J.; Kuo, C.J.; Li, D.Y. The axonal attractant Netrin-1 is an angiogenic factor. Proc. Natl. Acad. Sci. USA 2004, 101, 16210–16215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Vutskits, L.; Pepper, M.S.; Kiss, J.Z. VEGF is a chemoattractant for FGF-2-stimulated neural progenitors. J. Cell Biol. 2003, 163, 1375–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Yang, C.; Zhang, Y.; Han, C.; Zhang, G. Vascular Endothelial Growth Factor Mediates the Sprouted Axonogenesis of Breast Cancer in Rat. Am. J. Pathol. 2021, 191, 515–526. [Google Scholar] [CrossRef]
- de Wit, J.; Verhaagen, J. Role of semaphorins in the adult nervous system. Prog. Neurobiol. 2003, 7, 249–267. [Google Scholar] [CrossRef]
- Kessler, O.; Shraga-Heled, N.; Lange, T.; Gutmann-Raviv, N.; Sabo, E.; Baruch, L.; Machluf, M.; Neufeld, G. Semaphorin-3F is an inhibitor of tumor angiogenesis. Cancer Res. 2004, 64, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neufeld, G.; Sabag, A.D.; Rabinovicz, N.; Kessler, O. Semaphorins in Angiogenesis and Tumor Progression. Cold Spring Harb. Perspect. Med. 2012, 2, a006718. [Google Scholar] [CrossRef]
- Masur, K.; Niggeman, B.; Zanker, K.S.; Entschladen, F. Norepinephrine-induced migration of SW 480 colon carcinoma cells is inhibited by beta-blockers. Cancer Res. 2001, 61, 2866–2869. [Google Scholar]
- Drell, T.L., 4th; Joseph, J.; Lang, K.; Niggemann, B.; Zaenker, K.S.; Entschladen, F. Effects of neurotransmitters on the chemokinesis and chemotaxis of MDA-MB-468 human breast carcinoma cells. Breast Cancer Res. Treat. 2003, 80, 63–70. [Google Scholar] [CrossRef]
- Lang, K.; Drell, T.L., 4th; Lindecke, A.; Niggemann, B.; Kaltschmidt, C.; Zaenker, K.S.; Entschladen, F. Induction of a metastatogenic tumor cell type by neurotransmitters and its pharmacological inhibition by established drugs. Int. J. Cancer. 2004, 112, 231–238. [Google Scholar] [CrossRef]
- Palm, D.; Lang, K.; Niggemann, B.; Drell, T.L., 4th; Masur, K.; Zaenker, K.S.; Entschladen, F. The norepinephrine-driven metastasis development of PC-3 human prostate cancer cells in BALB/c nude mice is inhibited by beta-blockers. Int. J. Cancer. 2006, 118, 2744–2749. [Google Scholar] [CrossRef]
- Bunimovich, Y.L.; Keskinov, A.A.; Shurin, G.V.; Shurin, M.R. Schwann cells: A new player in the tumor microenvironment. Cancer Immunol. Immunother. 2017, 66, 959–968. [Google Scholar] [CrossRef]
- Bunge, R.P. Tissue culture observations relevant to the study of axon-Schwann cell interactions during peripheral nerve development and repair. J. Exp. Biol. 1987, 132, 21–34. [Google Scholar] [CrossRef]
- Armati, P.J.; Mathey, E.K. An update on Schwann cell biology--immunomodulation, neural regulation, and other surprises. J. Neurol. Sci. 2013, 333, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Whalley, K. Glia: Schwann cells provide life support for axons. Nat. Rev. Neurosci. 2014, 15, 698–699. [Google Scholar] [CrossRef]
- George, D.; Ahrens, P.; Lambert, S. Satellite glial cells represent a population of developmentally arrested Schwann cells. Glia. 2018, 66, 1496–1506. [Google Scholar] [CrossRef]
- Stassart, R.M.; Möbius, W.; Nave, K.-A.; Edgar, J.M. The Axon-Myelin Unit in Development and Degenerative Disease. Front. Neurosci. 2018, 12, 467. [Google Scholar] [CrossRef] [Green Version]
- Bolino, A. Myelin Biology. Neurotherapeutics 2021, 18, 2169–2184. [Google Scholar] [CrossRef]
- Conforti, L.; Gilley, J.; Coleman, M.P. Wallerian degeneration: An emerging axon death pathway linking injury and disease. Nat. Rev. Neurosci. 2014, 15, 394–409. [Google Scholar] [CrossRef]
- Jessen, K.R.; Mirsky, R. The repair Schwann cell and its function in regenerating nerves. J. Physiol. 2016, 594, 3521–3531. [Google Scholar] [CrossRef] [Green Version]
- Vaquié, A.; Sauvain, A.; Duman, M.; Nocera, G.; Egger, B.; Meyenhofer, F.; Falquet, L.; Bartesaghi, L.; Chrast, R.; Lamy, C.M.; et al. Injured Axons Instruct Schwann Cells to Build Constricting Actin Spheres to Accelerate Axonal Disintegration. Cell Rep. 2019, 27, 3152–3166.e7. [Google Scholar] [CrossRef] [Green Version]
- Harrison, B.M.; Gledhill, R.F.; McDonald, W.J. Remyelination after transient compression of the spinal cord. Proc. Aust. Assoc. Neurol. 1975, 12, 117–122. [Google Scholar] [CrossRef]
- Sroka, I.C.; Chopra, H.; Das, L.; Gard, J.M.C.; Nagle, R.B.; Cress, A.E. Schwann Cells Increase Prostate and Pancreatic Tumor Cell Invasion Using Laminin Binding A6 Integrin. J. Cell. Biochem. 2016, 117, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Deborde, S.; Omelchenko, T.; Lyubchik, A.; Zhou, Y.; He, S.; McNamara, W.F.; Chernichenko, N.; Lee, S.-Y.; Barajas, F.; Chen, C.-H.; et al. Schwann cells induce cancer cell dispersion and invasion. J. Clin. Investig. 2016, 126, 1538–1554. [Google Scholar] [CrossRef] [Green Version]
- Demir, I.E.; Boldis, A.; Pfitzinger, P.L.; Teller, S.; Brunner, E.; Klose, N.; Kehl, T.; Maak, M.; Lesina, M.; Laschinger, M.; et al. Investigation of Schwann cells at neoplastic cell sites before the onset of cancer invasion. J. Natl. Cancer Inst. 2014, 106, dju184. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Shurin, G.V.; Zhong, H.; Bunimovich, Y.L.; Han, B.; Shurin, M.R. Schwann Cells Augment Cell Spreading and Metastasis of Lung Cancer. Cancer Res. 2018, 78, 5927–5939. [Google Scholar] [CrossRef] [Green Version]
- Martyn, G.V.; Shurin, G.V.; Keskinov, A.A.; Bunimovich, Y.L.; Shurin, M.R. Schwann cells shape the neuro-immune environs and control cancer progression. Cancer Immunol. Immunother. 2019, 68, 1819–1829. [Google Scholar] [CrossRef]
- Shurin, G.V.; Kruglov, O.; Ding, F.; Lin, Y.; Hao, X.; Keskinov, A.A.; You, Z.; Lokshin, A.E.; LaFramboise, W.A.; Falo, L.D., Jr.; et al. Melanoma-Induced Reprogramming of Schwann Cell Signaling Aids Tumor Growth. Cancer Res. 2019, 79, 2736–2747. [Google Scholar] [CrossRef] [Green Version]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef]
- Saussez, S.; Laumbacher, B.; Chantrain, G.; Rodriguez, A.; Gu, S.; Wank, R.; Levite, M. Towards neuroimmunotherapy for cancer: The neurotransmitters glutamate, dopamine and GnRH-II augment substantially the ability of T cells of few head and neck cancer patients to perform spontaneous migration, chemotactic migration and migration towards the autologous tumor, and also elevate markedly the expression of CD3zeta and CD3epsilon TCR-associated chains. J. Neural Transm. 2014, 121, 1007–1027. [Google Scholar] [CrossRef]


{kind=link}
| Cancer | Species | Method | Year | Reference |
|---|---|---|---|---|
| Carcinoma, epithelioma, sarcoma, myxoma, lymphoma, neuroma | Human | Methylene blue staining | 1897 | [3] |
| Cervical tumor at vagus nerve | Human | Methylene blue staining | 1911 | [4] |
| Not specified | Rabbit | Tumor implantation & sympathetic transection | 1923 | [4] |
| Various | Human, rabbit, mouse | Golgi staining | 1925 | [4] |
| Epithelioma | Rabbit | Unilateral resection of the sympathetic nervous system | 1925 | [4] |
| Epithelial meningioblastoma | Human | Apathy’s medium | 1925 | [5] |
| Squamous cell cancer | Human | Methylene blue staining | 1926 | [4] |
| Benign growth | Human | Golgi staining | 1926 | [4] |
| Not specified | Mouse, rabbit | Nerve transection | 1926 | [4] |
| Malignant neoplasm & papilloma | Mouse | Experimental coal tar | 1927 | [4] |
| Sarcoma | Rat | Sciatic nerve transplantation into transplanted sarcoma | 1927 | [4] |
| Not specified | Rat | Sympathetic nervous system resection | 1927 | [4] |
| Benign & malignant tumors | Not specified | Bielschowsky stain | 1928 | [6] |
| Malignant, cancerous, & sarcomatous tumors | Human | Silver nitrate staining | 1928 | [7] |
| Carcinoma, mesothelioma | Rabbit, mouse | Electrical stimulation | 1949 | [8] |
| Prostatic adenocarcinoma | Human | Immunohistochemistry (H&E, S-100) | 2001 | [9] |
| Adenoma | Human | Uranyl acetate & lead citrate staining, electron microscopy | 2001 | [10] |
| Fibrosarcoma | Mouse | Electrophysiological recording, immunohistochemistry (PGP 9.5) | 2001 | [11] |
| Fibrosarcoma | Mouse | Neuroendocrinology (endothelin-1) | 2001 | [12] |
| Prostatic carcinoma | Human | Dark-phase microscopy, image analysis (Optimas 6.1) | 2001 | [13] |
| Urothelial bladder carcinoma | Human | Uranyl acetate & lead citrate staining, electron microscopy, immunohistochemistry (PGP 9.5, VIP) | 2002 | [14] |
| Esophageal & cardiac carcinoma | Human | Immunohistochemistry (CGRP, GAL, SP, NT, SOM, CCK, L-ENK, Dyn, NPY, M-ENK) | 2003 | [15] |
| Fibrosarcoma | Mouse | Sleeping Beauty transposon fluorescent transfection, immunohistochemistry (PGP 9.5, CGRP) | 2005 | [16] |
| Pancreatic adenocarcinoma | Mouse | Immunohistochemistry (CGRP, neurofilament 200, tyrosine hydroxylase) | 2005 | [17] |
| Prostatic carcinoma | Human | Immunohistochemistry (S-100) | 2008 | [18] |
| Colonic carcinoma | Human | Immunohistochemistry (PGP 9.5) | 2011 | [19] |
| Prostatic carcinoma & fibrosarcoma | Mouse | Transplantation with NGF administration | 2017 | [20] |
| Cancer | Species | Method | Year | Reference |
|---|---|---|---|---|
| Various | Human | Bielschowsky stain | 1910 | [4] |
| Malignant growth | Human | Golgi staining | 1926 | [4] |
| Benign & malignant tumors | Not specified | Not specified | 1928 | [28] |
| Malignant neoplasms | Human | Neurotrophin assessment | 1933 | [29] |
| Glomus tumor | Human | Bielschowsky stain | 1958 | [30] |
| Hemangioma, carcinoma, experimental transplanted tumor | Mouse | Indirect immunohistology (PGP 9.5) | 1994 | [31] |
| Chronic mastopathy, fibroadenoma, breast cancer | Mouse | Indirect immunohistology (PGP 9.5) | 1994 | [32] |
| Hepatocellular carcinoma, intrahepatic cholangiocarcinoma | Human | Immunohistochemistry (S-100) | 2001 | [33] |
| Oral squamous cell carcinoma | Human | Immunohistochemistry (PGP 9.5) | 2012 | [34] |
| Colonic carcinoma | Human | Immunohistochemistry (PGP 9.5) | 2012 | [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baraldi, J.H.; Martyn, G.V.; Shurin, G.V.; Shurin, M.R. Tumor Innervation: History, Methodologies, and Significance. Cancers 2022, 14, 1979. https://doi.org/10.3390/cancers14081979
Baraldi JH, Martyn GV, Shurin GV, Shurin MR. Tumor Innervation: History, Methodologies, and Significance. Cancers. 2022; 14(8):1979. https://doi.org/10.3390/cancers14081979
Chicago/Turabian StyleBaraldi, James H., German V. Martyn, Galina V. Shurin, and Michael R. Shurin. 2022. "Tumor Innervation: History, Methodologies, and Significance" Cancers 14, no. 8: 1979. https://doi.org/10.3390/cancers14081979
APA StyleBaraldi, J. H., Martyn, G. V., Shurin, G. V., & Shurin, M. R. (2022). Tumor Innervation: History, Methodologies, and Significance. Cancers, 14(8), 1979. https://doi.org/10.3390/cancers14081979