Simple Summary
The extensive array of applications in 3D culture models closely mimics the precise physiological conditions of in vivo settings, enabling the investigation of anticancer drug resistance, which suggests that, in addition to genes, cell structure significantly impacts toxicity outcomes. Commercial 3D culture systems offer a valuable platform to explore various facets of oral cancer research. These models present an ethical and reproducible alternative to animal studies, facilitating the examination of oral microbial population dynamics, cellular interactions between cancer cells and immune cells, and the role of macrophage cells in oral cancer.
Abstract
The onset and progression of oral cancer are accompanied by a dynamic interaction with the host immune system, and the immune cells within the tumor microenvironment play a pivotal role in the development of the tumor. By exploring the cellular immunity of oral cancer, we can gain insight into the contribution of both tumor cells and immune cells to tumorigenesis. This understanding is crucial for developing effective immunotherapeutic strategies to combat oral cancer. Studies of cancer immunology present unique challenges in terms of modeling due to the extraordinary complexity of the immune system. With its multitude of cellular components, each with distinct subtypes and various activation states, the immune system interacts with cancer cells and other components of the tumor, ultimately shaping the course of the disease. Conventional two-dimensional (2D) culture methods fall short of capturing these intricate cellular interactions. Mouse models enable us to learn about tumor biology in complicated and dynamic physiological systems but have limitations as the murine immune system differs significantly from that of humans. In light of these challenges, three-dimensional (3D) culture systems offer an alternative approach to studying cancer immunology and filling the existing gaps in available models. These 3D culture models provide a means to investigate complex cellular interactions that are difficult to replicate in 2D cultures. The direct study of the interaction between immune cells and cancer cells of human origin offers a more relevant and representative platform compared to mouse models, enabling advancements in our understanding of cancer immunology. This review explores commonly used 3D culture models and highlights their significant contributions to expanding our knowledge of cancer immunology. By harnessing the power of 3D culture systems, we can unlock new insights that pave the way for improved strategies in the battle against oral cancer.
1. Introduction
Head and neck cancers (HNCs), positioned as the seventh most prevalent cancer on a global scale, carry a significant burden of mortality. In 2018 alone, staggeringly, more than 170,000 deaths were attributed to HNCs, which also exhibit a discouraging prognosis characterized by a 5-year relative survival rate of about 68% in developing countries [1,2]. The WHO reports indicating that there are approximately 900,000 annual cases of HNCs, resulting in more than 400,000 deaths [3]. Particularly disheartening is the fact that developing countries experience even lower survival rates in this context [4,5,6,7]. Furthermore, oral cancer, encompassing approximately half of HNC instances, predominantly manifests as oral squamous cell carcinoma (OSCC), accounting for roughly 90% of these cases [8,9].
OSCC occurs within the oral cavity, encompassing vital areas such as the lips, gums, inner linings of the cheeks and lips, the front two-thirds of the tongue, the floor beneath the tongue, and the roof of the mouth [10]. Despite significant strides made in diagnostic techniques and the availability of various treatment approaches, the worldwide 5-year survival rate for OSCC continues to linger below 50%, underscoring the persistent challenges in effectively combatting this disease [11].
OSCC is characterized by an abundance of immune cells infiltrating the tumor, making it a tumor with high immunogenicity [12]. Within the tumor microenvironment (TME), which is a complex landscape consisting of the extracellular matrix (ECM), various stromal cells, and immune cells, there exists a dynamic interplay and coordination with the tumor cells. Examples of these interacting cells include tumor-associated macrophages (TAMs), regulatory T cells (Tregs), cancer-associated fibroblasts (CAFs), and endothelial cells [13]. Furthermore, the innate immune system contributes to the TME through macrophages, dendritic cells (DCs), neutrophils, myeloid-derived suppressor cells (MDSCs), natural killer cells (NKs), and innate lymphoid cells. The adaptive immune response is represented by T cells and B cells [14]. The communication and interaction among these cells, along with the ECM and tumor cells, play a significant role in driving tumor progression [15,16].
The utilization of three-dimensional (3D) models provides a novel avenue for investigating cancer immunology in preclinical studies. These models offer a more realistic and physiologically relevant environment compared to traditional two-dimensional (2D) models. Unlike murine models, 3D models are cost-effective, amenable to high-throughput research requirements, and can incorporate human cancers and immune components. The application of 3D models has significantly advanced various areas of cancer immunology research [17,18]. Among the commonly employed 3D models are spheroids, organoids, and microfluidic chips.
This paper aims to discuss the functions of immune cells related to tumors with a focus on in vitro cell culture technology. The review also provides an overview of the application of 3D models involved in oral microbial pathogenesis, drug discovery, and cell–cell interaction in carcinogenesis. Finally, the potential application of 3D models in the study of TAM functions in oral cancer is discussed.
2. Tumor Immune Microenvironment of Oral Cancer
The tumor immune microenvironment (TIME) of oral cancer is a complicated system composed of immune cells, cytokines, and chemokines that interact with tumor cells to promote or prevent tumor growth [19,20]. It is proven that the immunological microenvironment is vital in developing oral cancer and potentially serves as a target for cancer therapy [21,22].
In most cases, pro-inflammatory cytokines (IFN-γ, IL-2, IL-1α, IL-1β, TNF-α, IL-17, and IL-8), anti-inflammatory cytokines (IL-4 and IL-10), and pro-/anti-inflammatory cytokines (IL-6 and TGF-β) are unbalanced in oral cancer, resulting in an immunosuppressive environment. This immunosuppressive milieu permits cancer cells to avoid detection by the immune system [23,24]. A study showed how an unbalanced quantity of cytokines can act like a two-edged sword. In the study, IL-10, as an anti-inflammatory cytokine, exhibited an elevated expression, while TGF-β experienced a reduction. This situation ultimately led to the suppression of the immune system in the progressive stage of OSCC [25]. Therefore, gaining insights into the markers associated with oral cancer and comprehending the intricate dynamics between cancer cells, stromal cells, and the tumor microenvironment hold paramount significance in the realms of diagnosis and treatment.
2.1. Interaction between the Immune System and Tumor Cells
The immune system has a dual role in tumor growth, either suppressing it or promoting it, known as immunoediting [26]. Initially, the immune system can eliminate tumor cells, a phase called immunosurveillance. During this stage, there are lively molecules and immune cells at work, capable of identifying and eliminating cancerous cells. Furthermore, perilous signals such as Type I interferons (IFNs) are discharged by dying tumor cells or injured tissues as the tumor advances. These signals rouse the immune cells and promote a flexible, tailored anti-tumor immune reaction. Certain molecules such as MHC class I chain-related protein A and B (MICA and MICB) and the histocompatibility 60 (H60), which are plentiful on the surface of tumor cells, bind to activation receptors found on immune cells, setting them into action. The distribution of cluster of differentiation (CD)4+ T and CD8+ T cells facilitates a harmonized and well-balanced activation of both the innate and adaptive immune responses [27].
If the defective cells are not completely removed, the equilibrium phase begins, where tumor cells remain dormant with the help of interleukin (IL)-12, T cells, and IFN-γ [28]. During this phase, the adaptive immune system becomes crucial, leading to two possible outcomes: regression and elimination of the tumor or its progression [29,30]. For example, it was observed that after oral cancer cells in the dormancy stage were exposed to irradiation, M2 TAMs actively induce neovasculogenesis, fostering the resurgence of oral cancer [31]. The escape phase, extensively studied in cancer immunoediting, often occurs due to T-cell exhaustion [32]. This exhaustion involves intrinsic mechanisms of T cells, such as the Programmed death-1 (PD-1)/Programmed Cell Death Ligand 1 (PD-L1) pathway and immunoregulatory receptors such as cytotoxic T-lymphocyte antigen 4(CTLA-4), T-cell immunoglobulin and mucin-domain containing-3 (TIM-3), and lymphocyte activation gene-3 (LAG-3), as well as extrinsic pathways mediated by Tregs and MDSCs. These pathways secrete cytokines such as transforming growth factor beta (TGF-β) [33]. The interaction between hypoxia-inducible factor (HIF)-1α and immune cells contributes to the evasion of the immune system by tumors, including OSCC [34].
2.2. Immune and Non-Immune Markers
One strategy for early detection of oral cancer is the identification of immune and non-immune markers [35,36]. T cells, B cells, and NKs, among many other immune and non-immune cells, are used to diagnose oral cancer [37,38,39]. Non-immune markers used in diagnosis include changes in DNA, RNA, and protein expression levels. These markers can help identify individuals who are most prone to being affected by the condition, allowing for early identification and treatment [40]. Immune and non-immune biomarkers are listed in Table 1.

Table 1.
Immune and non-immune markers in oral cancer.
2.3. Stromal Cell and Extracellular Matrix (ECM) on Cancer Immunity
The intricate makeup of the tumor microenvironment and the interactions between the tumor and its surrounding stroma play a pivotal role in fueling tumor growth and facilitating its spread, resulting in unfavorable clinical outcomes in understanding and defining the disease, as it exerts significant influence over cancer cell invasion, migration, angiogenesis, resistance to drugs [84,85], maintenance of cancer stem cells [86], and evasion of immune surveillance [87,88].
Within the TME, the tumor stroma represents the noncancerous components and consists of an abundant ECM and various supportive cell types [89]. These include CAFs, endothelial cells, pericytes, and immune cells such as lymphocytes, neutrophils, DCs, monocytes, and TAMs, which are the most prevalent cell types. Additionally, there are less common factors such as MDSCs and mesenchymal stromal cells (MSCs) [84,90], as well as platelets [87,91]. These stromal cells actively engage in intricate interactions with tumor cells, with one another, and with the ECM. They achieve this by releasing chemokines, growth factors (GFs), enzymes, extracellular vesicles, and miRNAs that regulate the expression of genes and proteins, thereby influencing metabolic pathways associated with cancer [92]. Consequently, different cell types can either promote or suppress tumor growth depending on the specific cellular context [93]. Current in vitro culture models come with numerous limitations, including a lack of the dynamic interplay between cells and their surrounding environment, and alterations in cell shape, orientation, and proliferation. These disadvantages prompted the development of alternative models that better simulate in vivo conditions. Among these approaches, 3D culture appeared to be a promising method to bridge the gap and led to a significant emphasis on developing accurate models to study and simulate TME interactions both in laboratory settings and in living organisms [94]. Table 2 shows the role and function of stromal cells in oral cancer.
Table 2.
Mechanism and function of stromal cells in OSCC.
3. In Vitro Models in Oral Cancer
At present, the most common evaluation platform for drug development in oral cancer is conventional 2D in vitro models to their low cost, high reproducibility, and potential co-culture capability [134]. For example, cell co-culture in a 2D model helped in gaining a deeper comprehension of how CAFs interact with cancer cells within the microenvironment, revealing the potential for CAFs to regulate cancer cells as a means of therapeutic mediator [135]. However, 2D in vitro models are unable to mimic the physical geometry of the tumor, avoid the cross-contamination of culture media in multicellular models, and mimic the oxygen deprivation and irregular irrigation of the hypoxia region, which are key factors in the evaluation of tumor progression, chemoresistance, and treatment response [136,137]. Advanced in vitro systems, including spheroids, 3D scaffolds, and microfluidic devices, have thus been developed to overcome these barriers [138]. Although the application of these culture platforms to model the oral cancer microenvironment and its drug discovery is still in its infancy, recent research on oral cancer has used 3D in vitro models to advance the growing need for these systems for clinical translation, which is categorized into two forms, scaffold-free and scaffold-based strategies (Figure 1, Table 3).
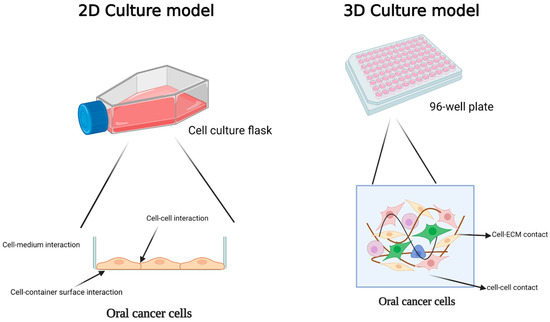
Figure 1.
Comparison of 2D and 3D models for oral cancer studies and exploring cellular interactions in these conditions. In a 2D culture, cells establish connections with neighboring cells, the container’s surface, and the surrounding environment on one side. In 3D culture, the cell maintains efficient communication with neighboring cells and the extracellular matrix. Created with BioRender.com.
Table 3.
Comparison among 2D model, 3D model, and animal model.
3.1. Two-Dimensional Models
In the realm of in vitro cancer research, the traditional way of cultivating cells in a flat, 2D environment using culture flasks or Petri dishes has long been favored due to its cost-effectiveness, simplicity, and ability to yield consistent results. The application of 2D cell culture models is accepted as a means to evaluate how cells respond to potential drugs. While the acceptance and uses of 2D cell culture have undoubtedly contributed to the advancement of comprehending the mechanisms of drug action, it is not without its constraints [139,140]. Therefore, this straightforward approach falls short of capturing the intricate, diverse, and ever-changing nature of the TME due to a range of limitations. These drawbacks encompass insufficient cell differentiation, unrealistic cell growth patterns, diminished drug resistance, and inaccurate reactions to mechanical or chemical signals [141]. Consequently, employing this 2D culture technique creates notable discrepancies when compared to the dynamic conditions present in living organisms, encompassing factors such as tissue development, specialized cell functions, cell division, cellular migration, gene and protein expression, signal transduction, and responsiveness to stimuli [142,143].
Relying solely on the 2D pre-clinical model is far from ideal and can generate misleading research results, primarily because cells exhibit distinct behaviors when cultured in a 2D setting compared to when they are organized in a 3D structure [144]. The earliest indication of this phenomenon dates back to 1985 when Miller and colleagues made a noteworthy discovery: tumor cells cultivated as multicellular spheroids within a collagen gel displayed heightened resistance to drugs compared to cells grown as a single layer [145]. Recent studies exploring the response to chemotherapy and radiation treatment in both 2D and 3D spheroids have yielded similar findings. These investigations revealed that oral cancer cells cultured in 3D tumor spheroids exhibited enhanced resistance to radiation and greater viability, even when exposed to higher doses of cisplatin [146,147,148].
3.2. Three-Dimensional Models
In a bid to overcome the constraints inherent in flat cell cultures, scientists have ventured into the realm of creating advanced in vitro cancer models that embody 3D structures. It has been scientifically established that the essential functions of the ECM and its significance as a vital biomaterial in tissue engineering cannot be understated. When it comes to obtaining the ECM for tissue engineering, two distinct strategies come into play: the employment of scaffolds or the scaffold-free approach. Each approach brings forth its own set of advantages and disadvantages; thus, the purpose of the research is decisive in choosing a type of strategy. The scaffold-based approach entails the utilization of an external material as a substitute for the natural ECM. This method grants the ability to manipulate the mechanical and chemical aspects of the cell’s ECM, while concurrently providing enhanced structural integrity for the regeneration of larger tissues [149]. Conversely, the scaffold-free approach facilitates tissue formation by compelling cells to aggregate into small spheres, aptly named “spheroids”, which possess the capacity to develop an inherent ECM. This approach offers several benefits, including a high initial cell density, swift formation, and the creation of a self-organized tissue-like structure [150] (Figure 2).
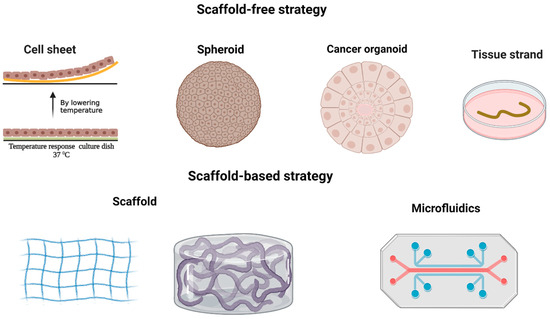
Figure 2.
In cancer research, two distinct approaches are employed for 3D modeling. The first is the scaffold-free strategy, which encompasses cell sheet, spheroid, cancer organoid, and tissue strand techniques. The second is the scaffold-based strategy, involving the use of scaffolds and microfluidics. Created with BioRender.com.
3.2.1. Scaffold-Free Strategy
The scaffold-free method takes a unique “bottom-up” approach to constructing tissues. Instead of using scaffolds, it relies on small building blocks in three forms: spheroid, cell sheet, or tissue strands. These building blocks naturally combine and form larger structures. Unlike the scaffold-based technique, the scaffold-free method does not heavily rely on cell growth and movement, making the tissue construction process faster. One significant advantage of this approach is its capability to create complex tissue and organ structures by using diverse building blocks made up of different types of cells [150].
In a study, a cell sheet was made up of patient-specific epithelial and sub-epithelial cells to create a co-culture system for head and neck squamous cell carcinoma (HNSCC) patient-derived explants. The findings revealed that this co-culture system significantly enhanced the survival and longevity of the explants compared to other non-matrix models [151]. Moreover, the system also demonstrated a decrease in viable cancer cells following standard-of-care treatments. However, despite the potential of cell sheet cultural techniques, they are not easily accessible, challenging to establish, and expensive, which limits their widespread adoption [152]. In addition, there is no original study about tissue strand technology and oral cancer. Thus, most studies in oral cancer using the scaffold-free method focus on spheroids and their advanced form, the organoid, which is addressed in the following sections.
Spheroid
Tumor cells, when cultivated as spheroids, take on a 3D structure that emphasizes cell-to-cell connections rather than interactions with the culture substrate. These spheroids, unlike conventional 2D cell cultures, provide numerous benefits. They foster enhanced cell-to-cell communication and facilitate improved diffusion of vital substances and molecules. Multicellular tumor spheroids bear a striking resemblance to avascular tumor masses found in large solid tumors. They exhibit comparable characteristics in terms of appearance, growth dynamics, cell division, drug permeability, nutrient gradients, oxygen levels, and the formation of a hypoxic core [153].
As in the case of in vivo tumors, deep within the spheroid, cells encounter limited diffusion of nutrients, causing the development of a necrotic core. Additionally, the accumulation of metabolic waste and decreased nutrient availability leads to a notable decrease in the microenvironment’s pH. This pH gradient has the potential to affect drug release and influence the activity of genes related to multiple drug resistance, such as Multiple drug resistance (MDR). Consequently, therapeutics targeting hypoxic regions may face reduced uptake [154]. On the other hand, cells residing in the outer boundary of the spheroid (around 100–300 μm from the surface) enjoy better access to oxygen and nutrients, leading to heightened rates of proliferation [155].
Numerous investigations have showcased the potential of oral cancer spheroids as a valuable tool for replicating different aspects of tumor tissue, particularly its growth state and hypoxic conditions [156,157,158]. However, the utilization of this system is limited due to the spheroids’ constrained ability to self-renew and differentiate. Furthermore, spheroids can only mimic small-scale microenvironments since controlling their growth becomes challenging during culturing, resulting in central necrosis if they exceed a certain size [159]. Moreover, spheroids lack the intricate complexity found in actual tumor tissues, primarily because they are often composed of a single cell line and have minimal ECM deposition [153]. In response to these limitations, researchers have devised spheroids that incorporate diverse stromal components, such as CAFs, alongside epithelial HNSCC cancer cells. The aim is to establish a more representative tumor model that accounts for the crucial role of stromal cells in tumorigenesis [160].
Organoid
An organoid represents a more advanced version of a cellular spheroid that remarkably imitates the physicochemical conditions of a specific tissue. In contrast to spheroids, organoids possess a more organized cellular architecture when grown on specific matrices, resulting in a structured arrangement of cells [161]. Organoids can display various biological characteristics akin to living tissues, such as tissue organization, regenerative capacity, drug responses, and the retention of tumor heterogeneity [162]. Notably, Driehuis et al. [163] successfully generated human mucosal organoids from 31 patient-derived HNSCC samples originating from diverse head and neck regions. Through immunohistochemical and genetic analyses, it was revealed that these organoids preserved the histological traits and molecular abnormalities observed in the original tumor specimens. Functionally, the organoids exhibited distinct sensitivity to a range of drugs, including cisplatin, carboplatin, cetuximab, and radiotherapy, highlighting their potential for drug screening in HNSCC. Furthermore, when transplanted beneath the skin of mice, the organoids displayed atypical features, tripolar mitotic figures, nuclear pleomorphism, and invasion into muscle tissue, showcasing their tumorigenic abilities in vivo. These findings indicate that HNSCC organoids faithfully replicate the genetic, histological, and functional aspects of HNSCC and hold promise as a platform for personalized cancer therapy.
Nevertheless, despite the benefits of organoids, there remain various limitations that necessitate attention. Tumor organoids usually recreate tumors originating from a single organ and fail to capture the intricate nature of cancer metastasis that involves multiple organs. Additionally, organoid cultures do not faithfully reproduce the spatial arrangement of cellular and non-cellular constituents within the tumor microenvironment [162]. Consequently, there are discrepancies in organoid complexity, size, morphology, and 3D structure, posing challenges in the standardization of organoid culture protocols [164].
3.2.2. Scaffold-Based Strategy
Scaffolds crafted from natural substances such as collagen, Matrigel, and silk, as well as synthetic materials including polyethylene glycol (PEG) and poly(lactic-co-glycolic acid) (PLGA), or a combination thereof, offer robust support for cell development and can emulate numerous mechanical and biochemical characteristics of the original ECM [165]. The approach of utilizing scaffolds has been extensively employed to engineer diverse oral tissues, facilitating the exploration of underlying cancer mechanisms. For instance, scientists have ingeniously created human-tissue-engineered oral mucosal models (TEOMMs) by seeding keratinocytes onto matrices populated with fibroblasts and culturing them at the interface between air and liquid [166]. TEOMMs have enabled researchers to capture distinct stages of OSCC, encompassing oral dysplasia, early invasive OSCC, and the neoplastic transformation associated with stromal fibroblasts [167,168,169]. As an illustration, Sawant et al. [170] developed a TEOMM representing normal, dysplastic, and malignant tongue tissues to investigate the neoplastic progression throughout various stages of tongue tumorigenesis. Histomorphometry, immunohistochemistry, and electron microscopy analyses performed on the three types of models demonstrated comparable stratified growth, cell proliferation, and differentiation between co-cultures and their respective native tissues. These findings suggest that crucial steps in the development of oral tumors can be replicated in vitro using scaffold-based engineered tissues.
Limitation and Improvement of 3D Model
However, it is crucial to acknowledge that the distinctions between traditional 3D models and the authentic TME go beyond mere dimensional variations. The development of cancer involves a multifaceted process influenced by numerous cues that synergistically manifest cancer hallmarks. Consequently, to capture in vivo scenarios with greater fidelity, it is imperative to devise biomimetic in vitro alternatives. In general, the conventional in vitro 3D models face notable limitations, including the constrained potential for vascularization and the absence of well-arranged spatial distribution of tumor cells and ECM compositions. These factors are pivotal aspects of the native TME. The 3D in vitro models incorporate co-culture of cancer cells with several types of stromal cells to mimic tumor characteristics in vivo. In vitro, 3D culture revealed more aggressive behaviors and resistance to anticancer medicines than 2D culture, underlining the potential use of chemotherapy screening [134]. Advanced cancer screening models have the potential to reduce drug development costs by reducing the number of animals required for clinical trials [171].
To tackle this obstacle, the utilization of microfluidic platforms presents a promising solution to mimic complex and dynamic tumor microenvironments. These platforms, intricately crafted at a microscopic scale, consist of interconnected chambers, membranes, and grooves that facilitate the controlled manipulation of small fluid volumes. They have found extensive applications in the field of in vitro modeling, including the development of organ-on-a-chip models [172,173,174,175] and point-of-care systems [176]. Within the realm of cancer research, microfluidic platforms are predominantly constructed using advanced techniques such as lithography and surface micromachining, employing materials such as polydimethylsiloxane, silicon, glass, polycarbonate, and polymethylmethacrylate [172,174,177,178].
4. Application of 3D Model in Oral Cancer
4.1. Oral Microbiota Study
OSCCs, which refer to malignant growths in the epithelial lining of the lip and oral cavity, contribute significantly to global mortality and morbidity [1]. The primary culprits behind OSCC are believed to be tobacco and alcohol consumption. However, recent research suggests that factors such as inflammation and the oral microbiome also play a role in developing oral cancer. The presence of bacterial dysbiosis in surface lesion samples of OSCC is characterized by significant alterations in bacterial composition and bacterial gene functions compared to control samples. Specifically, a notable enrichment of periodontitis-associated taxa within the OSCC samples was observed, including Fusobacterium, Dialister, Peptostreptococcus, Filifactor, Peptococcus, Catonella, and Parvimonas [179]. Several inflammatory mediators, salivary proteins, and oral microbiota are shared between inflammatory conditions of the oral cavity (IL-1, IL-6, IL-8, TNF-α), such as periodontitis, and oral cancer [180,181].
To imitate the interactions between a host and microorganisms in the mouth, an ideal host model should possess the ability to support a viable microbial community. It should facilitate the exchange of metabolites and nutrients with the microbes, interpret microbial signals, respond accordingly, and eventually achieve a state of balance within a specific timeframe [182].
Due to their increased physiological relevance, 3D host models are becoming increasingly valuable for both fundamental research and clinical applications. In the current field of oral (dental) research, animal models and 2D cell models continue to be heavily relied upon for mechanistic studies and pre-clinical testing of oral hygiene products and medications. These studies encompass various areas, such as the treatment of salivary secretory disorders, oral wound healing, and oral cancers [183,184,185]. Since hard tissues contain relatively few organic components, studies on corresponding host–microbe interactions have predominantly focused on biofilms formed on inorganic surfaces, extensively exploring their metabolism, composition, and structures [186,187,188].
The earliest organotypic models of the host involved using tissue explants embedded in or cultured on various types of 3D gels to create a structure resembling an organ [189]. In the case of oral 3D models, the tissue is obtained by directly removing a section of oral mucosa from a human or animal [190]. However, due to ethical concerns and the limited availability of human oral mucosa, along with significant variations between donors [191], researchers began developing in vitro reconstructed organotypic models as a scalable and reproducible alternative. These models were designed to replicate important biological characteristics of the oral epithelium or oral mucosa (gingiva), including the 3D tissue architecture and the communication between different types of host cells and ECM. Organotypic models of oral cancer have proven their worth in the discovery of novel drug targets, evaluating the effectiveness of drug delivery systems, and confirming the biosafety and compatibility of graft materials for oral-maxillofacial surgery. Extensive reviews of these models can be found elsewhere [192,193,194] (Figure 3A). However, despite the significant impact of the oral microbiota on host physiology and pathology, it has not been widely incorporated into these models. In the studies that have begun to address this, researchers have employed various representative microorganisms to imitate host–microbe interactions in different oral niches.
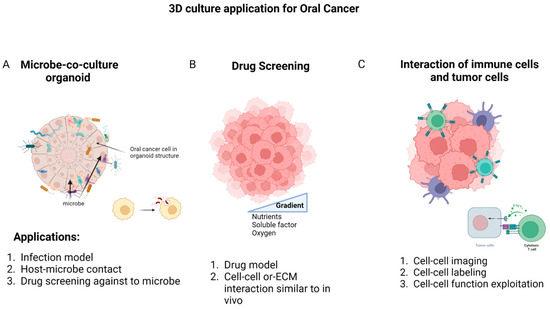
Figure 3.
The most common application of 3D culture in oral cancer is related to examining the microbial population in oral cancer, drug screening, and the interaction between immune cells and cancer cells. (A) The first application can be a suitable model for investigating infection; it facilitates the study of the relationship between cells and microbes, and is ideal for the screening of effective drugs for infection control in oral cancer. (B) The second approach offers a solution to explore drug resistance mechanisms and examine crucial cellular interactions more effectively. (C) Finally, the investigation of interactions between oral cancer cells and immune cells involves labeling and imaging cell contacts to identify cell functions. Created with BioRender.com.
Various types of cells were used as host cell sources in 3D oral host–microbe interaction models, including cancer cell lines [195,196], immortalized cell lines [197,198], and primary cells [186,199]. Studies utilizing these different cell sources showed similar outcomes when exposed to specific microbes, such as Candida albicans, in terms of tissue histology, Lactate dehydrogenase (LDH release), Human beta-defensin-2 (hBD2) expression, and secretions of TNF-α, IL-1β, and CXCL-8 [200].
When exposed to commensal microbial species, reconstructed human mucosal (RHM) models using either primary gingival cells or immortalized telomerase reverse transcriptase (TERT) cell lines exhibited similar responses. They remained viable and showed protective responses, such as activation of toll-like receptor (TLR) signaling pathways, increased expression of antimicrobial peptides (Elafin, hBD-2, hBD-3), and release of cytokines (IL-6, CXCL8, CXCL1, and C-C Motif Chemokine Ligand (CCL) 20) (Shang et al., 2018, 2019, 2020). Following exposure to a pathogenic biofilm composed of five species, two RHM models constructed with different keratinocyte cell lines demonstrated comparable levels of tissue damage, which were lower than those observed in a commercial reconstructed human epidermis (RHE) model after the same biofilm exposure [195]. Another study suggested a slightly higher invasion of Porphyromonas gingivalis occurred in 3D models using the H357 cell line (Human oral squamous cell carcinoma) compared to primary keratinocytes, but the difference was not significant [201].
4.2. Drug Discovery
Currently, the most commonly used method to evaluate drug development for HNCs is the conventional 2D in vitro model. These models are favored due to their low cost, high reproducibility, and potential for co-culture capabilities [134]. However, there are certain limitations with 2D models that need to be addressed. Firstly, they are unable to replicate the physical structure of tumors. Secondly, they cannot prevent the mixing of culture media in multicellular models, which can lead to cross-contamination. Lastly, they fail to simulate the oxygen deprivation and irregular blood flow in the hypoxia region, which are crucial factors in assessing tumor progression, chemoresistance, and treatment response [136,137]. To overcome these challenges, advanced in vitro systems such as spheroids, 3D scaffolds, and microfluidic devices have been developed [138]. Although the use of these culture platforms to model the HNCs and oral cancer microenvironment, and facilitate drug discovery, is still in its early stages, recent research in HNCs has shown promising results using 3D in vitro models. These advancements are essential to meet the increasing demand for improved systems for clinical translation (Table 4, Figure 3B).
Table 4.
3D models for drug discovery in OSCC and HNCs.
4.3. Cell–Cell Interactions
Understanding the progression of cancer heavily relies on comprehending the intricate microenvironment in which malignant tumor cells thrive [219]. The physical and biochemical attributes of this environment play a pivotal role in regulating various aspects of cancer, including cell differentiation, proliferation, invasion, and metastasis [220]. Thus, it is imperative to gain insights into the interactions and communication between cancer cells and their surrounding tissue, referred to as the tumor stroma, as this interplay profoundly influences the advancement of the disease. In order to mimic the conditions of TME, 3D in vitro models have become widely employed [221,222] (Figure 3C).
The use of 3D organoid models in co-culture systems has emerged as a prominent area of research, particularly in the evaluation of anticancer properties. These models aim to mimic the complex interactions occurring within the tumor microenvironment [223]. Likewise, the exchange of signals between stromal cells and cancer cells through paracrine signaling has been recognized to promote both cell proliferation and the development of drug resistance. In Table 5, various model systems employed in studies related to HNCs are presented. These models offer the advantage of customization according to the specific parameters being investigated, surpassing traditional techniques. For example, to understand the role of monocytes, they can be cultured alongside HNC cells [224,225,226,227]. Similarly, fibroblasts and peripheral blood mononuclear cells (PBMCs) can be co-cultured with HNC cells for epidermal growth factor receptor (EGFR)-related investigations [228] and antibody testing [229], respectively.
Several previous studies have provided evidence regarding the involvement of TAMs and Human Dermal Fibroblasts (HDFs) in cancer stemness and invasion, respectively [230,231]. The response to drugs targeting EGFR has been investigated with CAFs, revealing variations [214]. Various organoid models of HNCs have been established to investigate signaling pathways, such as Extracellular signal-regulated kinase 1/2 (ERK1/2) and Nanog [232], as well as their interactions with Human herpes simplex-1 (HSV1) and papillomavirus type 16 (HPV16) [163], the invasiveness of cancer [233], drug screening [234], and other defining characteristics [235]. An innovative microfluidics platform called the Hydrodynamic Shuttling Chip (HSC) has been developed to separate and co-culture single-cell SCC with lymphatic endothelial cells, enabling the observation of cell motility and intercellular communication [236].
Table 5.
Interactions of oral cancer cells and immune cells.
Table 5.
Interactions of oral cancer cells and immune cells.
| 3D Model | Aim | Result | References |
|---|---|---|---|
| Co-culture of monocytes with spheroids originating Malignant/benign HNC | The connection between the response of this cytokine co-culture and the prediction of outcomes. | The secretion of IL-6 during in vitro co-culture with monocytes and BF 1-spheroids serves as a prognostic indicator for recurrence and overall prognosis, whereas co-culture with monocytes and MF 2-spheroids predicts the likelihood of recurrence. | [227] |
| Co-culture of HNC cell line with fibroblasts in spheroid form | Generation of a spheroid model of EGFR-expressing HNC. | The upregulation of chemokine expression by anti-EGFR mAb 3 promotes the infiltration of leukocytes into tumor spheroids. This unique mechanism of action of anti-EGFR mAb could potentially enhance the anti-tumor effects of the antibody in living organisms. | [228] |
| Spheroid form of HNC cell line culture with leukocytes from PBMC 4 | The evaluation of utilizing a 3D tumor cell culture model, specifically spheroids, as a suitable representation of micro-metastases. | The utilization of the spheroid model demonstrates the manifestation of pathophysiological traits, intricacy, and heterogeneity of tumor tissue observed in vivo, which significantly impacts the effectiveness of therapeutic interventions. | [229] |
| Co culture of HNC spheroids with TAMs | The signaling of CD44, influenced by TAMs, has the potential to facilitate stemness through the PI3K-4EBP1-SOX2 pathway. This effect may occur by regulating the availability of HA 5, which is the primary ligand for CD44. | The results establish a mechanistic connection between CD44 in tumor cells, TAMs, and the properties of CSCs 6 at the interface between tumor and stroma. This connection highlights a crucial area for targeting and discovering drugs. | [231] |
| Co-culture of HNC cell line with HDFs | The understanding of how cancer cells, fibroblasts, and the surrounding collagen matrices interact and promote cancer cell invasion in different environments with varying concentrations of collagen. | The presence of HDFs played a crucial role in facilitating the invasion of HNC cells into the surrounding extracellular matrix characterized by high collagen concentration, elevated storage modulus, and narrow pore sizes. | [230] |
| Co-culture of HNC cell line with CAFS | Assessing the impact of CAFs on the treatment response and migratory behavior of HNC. | The presence of CAFs resulted in enhanced cell proliferation within the tumor spheroids, which was accompanied by elevated EGFR expression. Notably, spheroids exhibiting heightened EGFR expression displayed an augmented response to cetuximab treatment. | [210] |
| HNC spheroids | Role of ERK1/2-Nanog pathway in tumorigenesis in HNC. | HNSCCs sustain a population of CSCs by utilizing the ERK1/2 signaling pathway and Nanog. | [232] |
| Oral mucosal Organoids and HNC patient-derived tumoroids | In vitro 3D model for HNC. | Drug screening for both existing and experimental therapeutic treatments for HNC. | [163] |
| HNC spheroids | The correlation of CD44 and HIF-1α expression. | By focusing on HIF-1α, the impact of NOTCH1-induced stemness, which controls the reaction to chemotherapy or radiotherapy as well as the malignancy in CD44+ HNSCCs, was reduced. Targeting the signaling of HIF-1α/NOTCH1 could potentially serve as a therapeutic approach for the treatment of HNSCC. | [237] |
| Co-culture of OSCC cell line with CAFS | Role of stromal NNMT 7 in TME. | The harmful cancer-promoting effects caused by stromal NNMT were reduced when fibroblasts were treated with inhibitors targeting collagen production, such as losartan, tranilast, and halofuginone. | [238] |
1 benign, 2 malignant, 3 monoclonal antibody, 4 peripheral blood mononuclear cells, 5 hyaluronic acid, 6 cancer stem cells, 7 Nicotinamide N-methyltransferase.
5. Potential Application of 3D Model in Studying TAM Functions in Oral Cancer
Within the TME, immune cell components, notably TAMs [239,240], play a dominant role in suppressing the adaptive immune system through various pathways [241,242].
TAMs employ multiple strategies to hinder the activity of immune cells. They express receptor molecules for immune checkpoint proteins, including PD-1 and CTLA-4, while also releasing cytokines and chemokines that trigger Treg-mediated suppression pathways [243]. This interplay between TAMs and other immune elements is bidirectional.
It is crucial to utilize 3D in vitro tumor models to investigate the interaction between tumor cells and their microenvironment in a biologically relevant manner. Organotypic co-cultures have been previously employed to examine the malignant growth of human SCC cell lines in vitro, where the cell lines are cultivated on a collagen-I gel containing fibroblasts to mimic the stromal environment [244]. Recognizing the significant role played by macrophages in tumor progression, this model was further developed by incorporating macrophages into the collagen gel of these organotypic tumor co-cultures. This approach was established as both a murine and a human system for studying skin SCCs. The impact of macrophages on tumor progression relies on their polarization state. It was demonstrated that in organotypic co-cultures, the polarization of macrophages can be directed towards either an M1 or an M2 phenotype by introducing recombinant IFN-γ and lipopolysaccharide (LPS) or IL-4, respectively, into the growth medium. Stimulation of macrophages with IL-4 in these cultures led to an intensified invasion of tumor cells, as evidenced by basement membrane degradation, increased collagenolytic activity, and elevated levels of MMP-2 and MMP-9. Interestingly, prolonged co-culture with tumor cells for three weeks resulted in spontaneous M2 polarization of macrophages without the need for IL-4 treatment. Thus, the integration of macrophages into organotypic co-cultures of murine or human skin SCCs has proven to be successful, providing a valuable model for investigating the activation of macrophages towards a phenotype that supports tumor growth [245].
The influence of different macrophage phenotypes on cancer cells was examined, revealing that activated macrophages of the M2 phenotype had a substantial impact on promoting the proliferation, migration, and invasion abilities of cancer cells. To mimic the tumor microenvironment, co-cultures of M2 macrophages and cancer cells were established within agarose hydrogels. It was confirmed that the expression of CXCL2, a chemokine, was notably higher in the co-culture system. Functional analysis data indicated that the addition of recombinant human CXCL2 significantly enhanced the migration and invasion abilities of cancer cells while impairing their adhesion ability. Notably, the effects of CXCL2 on cancer cells were effectively diminished by neutralizing CXCL2 or blocking its receptor, CXCR2. This observation suggests that CXCL2 may play a critical role in facilitating metastasis [246].
TAMs exhibit a mixed phenotype combining characteristics of both M1 and M2 macrophages, and their distribution within the tumor microenvironment is believed to be dynamic and evolving throughout tumor progression [247,248,249]. However, the understanding of how TAMs specifically influence the tumor microenvironment is limited by the absence of suitable 3D in vitro models that can capture the intricate dynamics between cells with high spatial and temporal resolution. To address this, a novel microphysiological “tumor-on-a-chip” (TOC) device was employed to investigate the impact of distinct subsets of macrophages on tumor behavior. The TOC device features three interconnected chambers, with tumor cells loaded in both the upper and lower chambers, while the central chamber contains a living microvascular network that can be perfused. When human pancreatic or colorectal cancer cells were introduced alongside M1-polarized macrophages, a significant inhibition of tumor growth and tumor-induced angiogenesis was observed. Detailed protein analysis and neutralization studies using specific antibodies confirmed that these effects were mediated through the production of C-X-C motif chemokines (CXCL9), CXCL10, and CXCL11 by the M1 macrophages. In contrast, M2 macrophages facilitated increased migration of tumor cells into the vascularized chamber, without inhibiting tumor growth or angiogenesis. This innovative TOC model provides valuable insights into the complex interactions between TAMs and tumor cells, shedding light on the contrasting roles played by M1 and M2 macrophages in the tumor microenvironment [250].
Macrophages, commonly found within the TME, actively collaborate with cancer cells during the invasion process [251,252], coinciding with the presence of elevated interstitial flow (IF) levels [253,254]. Although studies have extensively explored the biochemical mechanisms driving pro-tumor macrophage functions, IF within the TME has often been neglected [239,255]. The concept of a 3D microfluidic model revolves around understanding the dynamics of fluid movement within micro-sized channels and the advanced techniques involved in fabricating miniature devices comprising chambers and tunnels that serve as pathways or confinement spaces for fluid flow [256]. Therefore, a novel 3D microfluidic model was devised to investigate how IF impacts macrophage migration and its potential contribution to cancer invasion when combined with tumor cells [257]. Interestingly, the presence of either tumor cells or IF individually heightened macrophage migration in terms of directionality and speed. However, when tumor cells and IF were simultaneously present, no additional effect on macrophage migration directionality and speed was observed [257].
6. Conclusions and Future Perspective
Over the past decade, remarkable progress has been made in unraveling the mysteries of the immune system’s fight against cancer. These advancements have spanned a wide range, from fundamental scientific research to practical clinical studies. As our understanding of the underlying biology becomes more intricate, it is essential to employ models that can capture this complexity. Three-dimensional culture systems have emerged as invaluable tools that offer both complexity and interpretability.
Geometry, multicellularity, and continuous irrigation are important characteristics for generating oral-cancer-specific in vitro models for drug screening and development. Organotypic multicellular spheroid and organoid cultures are useful for simulating cancer-specific TME by reflecting required shape and cell–cell/–ECM interactions found in tumor tissues. Organotypic models can be integrated with microfluidic devices to assess cell-to-cell communication and obstacles to mass transport of oxygen, nutrients, and medicinal therapies. The use of a tumor-on-a-chip technology is intended to reduce the requirement for animal models and the likelihood of clinical trial failures in translational research.
The application of 3D culture systems has revolutionized cancer research, shedding light on various aspects such as the distribution of immunotherapy, the infiltration of immune cells, and the suppression of the immune system caused by cancer. These systems have provided new insights and deepened our understanding in these areas and beyond. However, there are still untapped areas in the field of cancer immunology where 3D culture systems have yet to make their mark.
For instance, understanding the migration and function of Treg cells, unraveling the biology of myeloid-derived suppressor cells, and advancing vaccine development are promising avenues that could benefit from the application of 3D culture systems. As the cost of 3D technology decreases and accessibility improves, and as we explore the conceptual possibilities of these systems, it is highly likely that their popularity will continue to soar. In fact, they may even replace 2D culture methods altogether, marking a new era in cancer research and therapy.
Hence, 3D culture systems present an immense prospect for investigating cancer immunology; however, they do come with certain limitations. The heightened intricacy of these systems can pose challenges when it comes to reproducing results both within and between experiments. Additionally, compared to 2D culture systems, 3D culture systems are costlier and less readily accessible. Some microscopy techniques may struggle to capture images of 3D cultures due to their depth and limited transparency.
When it comes to utilizing 3D culture systems for studying cancer immunology, maintaining primary immune cells in culture for extended periods can be difficult, regardless of whether it is in a 2D or 3D environment. Moreover, the extraordinary complexity of the immune system, with its multiple steps and interactions between various cell types, may restrict the applicability of simple heterotypic or multicellular culture methods.
However, there is good news on the horizon. As technology continues to advance, the accessibility, versatility, and relevance of 3D models are expected to expand concurrently. This suggests that the limitations currently associated with 3D culture systems can be overcome with further developments, opening up even more possibilities for exploring cancer immunology in the future.
Author Contributions
X.H. conceptualized and designed the study. E.D.A. wrote and finalized the manuscript. X.H. supervised the project. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data will be made available on request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Patterson, R.H.; Fischman, V.G.; Wasserman, I.; Siu, J.; Shrime, M.G.; Fagan, J.J.; Koch, W.; Alkire, B.C. Global Burden of Head and Neck Cancer: Economic Consequences, Health, and the Role of Surgery. Otolaryngol. Head Neck Surg. 2020, 162, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Today; International Agency for Research on Cancer: Lyon, France, 2018; Volume 3, p. 2019.
- Chaturvedi, A.K.; Anderson, W.F.; Lortet-Tieulent, J.; Curado, M.P.; Ferlay, J.; Franceschi, S.; Rosenberg, P.S.; Bray, F.; Gillison, M.L. Worldwide trends in incidence rates for oral cavity and oropharyngeal cancers. J. Clin. Oncol. 2013, 31, 4550–4559. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Ghani, W.M.N.; Ramanathan, A.; Prime, S.S.; Yang, Y.H.; Razak, I.A.; Rahman, Z.A.A.; Abraham, M.T.; Mustafa, W.M.W.; Tay, K.K.; Kallarakkal, T.G.; et al. Survival of Oral Cancer Patients in Different Ethnicities. Cancer Investig. 2019, 37, 275–287. [Google Scholar] [CrossRef]
- Jou, A.; Hess, J. Epidemiology and Molecular Biology of Head and Neck Cancer. Oncol. Res. Treat. 2017, 40, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Tomita, H.; Hirata, A.; Ishida, K.; Hisamatsu, K.; Hatano, Y.; Kanayama, T.; Niwa, A.; Noguchi, K.; Kato, K.; et al. Promotion of cell proliferation by the proto-oncogene DEK enhances oral squamous cell carcinogenesis through field cancerization. Cancer Med. 2017, 6, 2424–2439. [Google Scholar] [CrossRef] [PubMed]
- Vigneswaran, N.; Williams, M.D. Epidemiologic trends in head and neck cancer and aids in diagnosis. Oral Maxillofac. Surg. Clin. N. Am. 2014, 26, 123–141. [Google Scholar] [CrossRef]
- Daraei, P.; Moore, C.E. Racial Disparity Among the Head and Neck Cancer Population. J. Cancer Educ. 2015, 30, 546–551. [Google Scholar] [CrossRef]
- Sathiasekar, A.C.; Mathew, D.G.; Lal, M.S.J.; Prakash, A.A.A.; Kumar, K.U.G. Oral Field Cancerization and Its Clinical Implications in the Management in Potentially Malignant Disorders. J. Pharm. Bioallied Sci. 2017, 9 (Suppl. S1), S23–S25. [Google Scholar] [PubMed]
- Hadler-Olsen, E.; Wirsing, A.M. Tissue-infiltrating immune cells as prognostic markers in oral squamous cell carcinoma: A systematic review and meta-analysis. Br. J. Cancer 2019, 120, 714–727. [Google Scholar] [CrossRef]
- Freeman, P.; Mielgo, A. Cancer-Associated Fibroblast Mediated Inhibition of CD8+ Cytotoxic T Cell Accumulation in Tumours: Mechanisms and Therapeutic Opportunities. Cancers 2020, 12, 2687. [Google Scholar] [CrossRef]
- Hinshaw, D.C.; Shevde, L.A. The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res. 2019, 79, 4557–4566. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, H.; Hagerling, C.; Werb, Z. Roles of the immune system in cancer: From tumor initiation to metastatic progression. Genes Dev. 2018, 32, 1267–1284. [Google Scholar] [CrossRef] [PubMed]
- Nazemi, M.; Rainero, E. Cross-talk between the tumor microenvironment, extracellular matrix, and cell metabolism in cancer. Front. Oncol. 2020, 10, 239. [Google Scholar] [CrossRef]
- Diskin, B.; Adam, S.; Cassini, M.F.; Sanchez, G.; Liria, M.; Aykut, B.; Buttar, C.; Li, E.; Sundberg, B.; Salas, R.D.; et al. PD-L1 engagement on T cells promotes self-tolerance and suppression of neighboring macrophages and effector T cells in cancer. Nat. Immunol. 2020, 21, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.V.; Chisamore, M.J.; Chaney, M.F.; Maradeo, M.E.; Anderson, J.; Baltus, G.A.; Pinheiro, E.M.; Uebele, V.N. Assessment of Clinical Activity of PD-1 Checkpoint Inhibitor Combination Therapies Reported in Clinical Trials. JAMA Netw. Open 2020, 3, e1920833. [Google Scholar] [CrossRef]
- Arner, E.N.; Rathmell, J.C. Metabolic programming and immune suppression in the tumor microenvironment. Cancer Cell 2023, 41, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Bule, P.; Aguiar, S.I.; Aires-Da-Silva, F.; Dias, J.N.R. Chemokine-Directed Tumor Microenvironment Modulation in Cancer Immunotherapy. Int. J. Mol. Sci. 2021, 22, 9804. [Google Scholar] [CrossRef] [PubMed]
- Curry, J.M.; Sprandio, J.; Cognetti, D.; Luginbuhl, A.; Barad, V.; Pribitkin, E.; Tuluc, M. Tumor microenvironment in head and neck squamous cell carcinoma. Semin. Oncol. 2014, 41, 217–234. [Google Scholar] [CrossRef]
- Liu, C.; Wang, M.; Zhang, H.; Li, C.; Zhang, T.; Liu, H.; Zhu, S.; Chen, J. Tumor microenvironment and immunotherapy of oral cancer. Eur. J. Med. Res. 2022, 27, 198. [Google Scholar] [CrossRef]
- Kondoh, N.; Mizuno-Kamiya, M. The Role of Immune Modulatory Cytokines in the Tumor Microenvironments of Head and Neck Squamous Cell Carcinomas. Cancers 2022, 14, 2884. [Google Scholar] [CrossRef]
- Niklander, S.E. Inflammatory Mediators in Oral Cancer: Pathogenic Mechanisms and Diagnostic Potential. Front. Oral Health 2021, 2, 642238. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, N.; Guan, X.; Wu, H.; Sun, Z.; Zeng, H. Immunosuppression Induced by Chronic Inflammation and the Progression to Oral Squamous Cell Carcinoma. Mediat. Inflamm. 2016, 2016, 5715719. [Google Scholar] [CrossRef]
- Kim, R.; Emi, M.; Tanabe, K. Cancer immunoediting from immune surveillance to immune escape. Immunology 2007, 121, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer immunoediting: Integrating immunity’s roles in cancer suppression and promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef]
- Tavakoli, F.; Sartakhti, J.S.; Manshaei, M.H.; Basanta, D. Cancer immunoediting: A game theoretical approach. In Silico Biol. 2021, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Aragon-Sanabria, V.; Kim, G.B.; Dong, C. From Cancer Immunoediting to New Strategies in Cancer Immunotherapy: The Roles of Immune Cells and Mechanics in Oncology. Adv. Exp. Med. Biol. 2018, 1092, 113–138. [Google Scholar] [CrossRef]
- Kubick, B.J.; Fan, X.; Crouch, A.; McCarthy, R.; Roop, D.R. Tracing the Equilibrium Phase of Cancer Immunoediting in Epidermal Neoplasms via Longitudinal Intravital Imaging. J. Investig. Dermatol. 2020, 140, 891–900.e810. [Google Scholar] [CrossRef] [PubMed]
- Okubo, M.; Kioi, M.; Nakashima, H.; Sugiura, K.; Mitsudo, K.; Aoki, I.; Taniguchi, H.; Tohnai, I. M2-polarized macrophages contribute to neovasculogenesis, leading to relapse of oral cancer following radiation. Sci. Rep. 2016, 6, 27548. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sun, Q.; Ren, X. Novel strategies for cancer immunotherapy: Counter-immunoediting therapy. J. Hematol. Oncol. 2023, 16, 38. [Google Scholar] [CrossRef] [PubMed]
- Perri, F.; Ionna, F.; Longo, F.; Della Vittoria Scarpati, G.; De Angelis, C.; Ottaiano, A.; Botti, G.; Caponigro, F. Immune Response Against Head and Neck Cancer: Biological Mechanisms and Implication on Therapy. Transl. Oncol. 2020, 13, 262–274. [Google Scholar] [CrossRef]
- Suárez-Sánchez, F.J.; Lequerica-Fernández, P.; Suárez-Canto, J.; Rodrigo, J.P.; Rodriguez-Santamarta, T.; Domínguez-Iglesias, F.; García-Pedrero, J.M.; de Vicente, J.C. Macrophages in Oral Carcinomas: Relationship with Cancer Stem Cell Markers and PD-L1 Expression. Cancers 2020, 12, 1764. [Google Scholar] [CrossRef] [PubMed]
- Almangush, A.; Heikkinen, I.; Mäkitie, A.A.; Coletta, R.D.; Läärä, E.; Leivo, I.; Salo, T. Prognostic biomarkers for oral tongue squamous cell carcinoma: A systematic review and meta-analysis. Br. J. Cancer 2017, 117, 856–866. [Google Scholar] [CrossRef]
- Rivera, C.; Oliveira, A.K.; Costa, R.A.P.; De Rossi, T.; Leme, A.F.P. Prognostic biomarkers in oral squamous cell carcinoma: A systematic review. Oral Oncol. 2017, 72, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Karmakar, T.; Arora, N.; Mukherjee, G. Immune and genomic signatures in oral (head and neck) cancer. Heliyon 2018, 4, e00880. [Google Scholar] [CrossRef]
- Dikova, V.; Jantus-Lewintre, E.; Bagan, J. Potential Non-Invasive Biomarkers for Early Diagnosis of Oral Squamous Cell Carcinoma. J. Clin. Med. 2021, 10, 1658. [Google Scholar] [CrossRef]
- Grimm, M.; Feyen, O.; Hofmann, H.; Teriete, P.; Biegner, T.; Munz, A.; Reinert, S. Immunophenotyping of patients with oral squamous cell carcinoma in peripheral blood and associated tumor tissue. Tumour Biol. 2016, 37, 3807–3816. [Google Scholar] [CrossRef]
- Németh, Z.; Velich, N.; Bogdan, S.; Ujpál, M.; Szabó, G.; Suba, Z.S. The prognostic role of clinical, morphological and molecular markers in oral squamous cell tumors. Neoplasma 2005, 52, 95–102. [Google Scholar] [PubMed]
- Gustafson, M.P.; Lin, Y.; Bleeker, J.S.; Warad, D.; Tollefson, M.K.; Crispen, P.L.; Bulur, P.A.; Harrington, S.M.; Laborde, R.R.; Gastineau, D.A.; et al. Intratumoral CD14+ Cells and Circulating CD14+HLA-DRlo/neg Monocytes Correlate with Decreased Survival in Patients with Clear Cell Renal Cell Carcinoma. Clin. Cancer Res. 2015, 21, 4224–4233. [Google Scholar] [CrossRef]
- Motomura, Y.; Kanno, S.; Asano, K.; Tanaka, M.; Hasegawa, Y.; Katagiri, H.; Saito, T.; Hara, H.; Nishio, H.; Hara, T.; et al. Identification of Pathogenic Cardiac CD11c+ Macrophages in Nod1-Mediated Acute Coronary Arteritis. Arter. Thromb. Vasc. Biol. 2015, 35, 1423–1433. [Google Scholar] [CrossRef]
- Weber, M.; Büttner-Herold, M.; Hyckel, P.; Moebius, P.; Distel, L.; Ries, J.; Amann, K.; Neukam, F.W.; Wehrhan, F. Small oral squamous cell carcinomas with nodal lymphogenic metastasis show increased infiltration of M2 polarized macrophages—An immunohistochemical analysis. J. Craniomaxillofac. Surg. 2014, 42, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Haque, A.; Moriyama, M.; Kubota, K.; Ishiguro, N.; Sakamoto, M.; Chinju, A.; Mochizuki, K.; Sakamoto, T.; Kaneko, N.; Munemura, R.; et al. CD206+ tumor-associated macrophages promote proliferation and invasion in oral squamous cell carcinoma via EGF production. Sci. Rep. 2019, 9, 14611. [Google Scholar] [CrossRef]
- Tan-Garcia, A.; Wai, L.E.; Zheng, D.; Ceccarello, E.; Jo, J.; Banu, N.; Khakpoor, A.; Chia, A.; Tham, C.Y.L.; Tan, A.T.; et al. Intrahepatic CD206+ macrophages contribute to inflammation in advanced viral-related liver disease. J. Hepatol. 2017, 67, 490–500. [Google Scholar] [CrossRef]
- Yamagata, Y.; Tomioka, H.; Sakamoto, K.; Sato, K.; Harada, H.; Ikeda, T.; Kayamori, K. CD163-Positive Macrophages Within the Tumor Stroma Are Associated With Lymphangiogenesis and Lymph Node Metastasis in Oral Squamous Cell Carcinoma. J. Oral Maxillofac. Surg. 2017, 75, 2144–2153. [Google Scholar] [CrossRef]
- Jardim, J.F.; Gondak, R.; Galvis, M.M.; Pinto, C.A.L.; Kowalski, L.P. A decreased peritumoral CD1a+ cell number predicts a worse prognosis in oral squamous cell carcinoma. Histopathology 2018, 72, 905–913. [Google Scholar] [CrossRef]
- Peña-Cardelles, J.F.; Pozo-Kreilinger, J.J.; Roncador, G.; Esteban-Hernández, J.; Cebrián-Carretero, J.L.; Moro-Rodríguez, J.E. Expression of clec9a in the oral cancer microenvironment. A preliminary immunohistochemical pilot study. Med. Oral Patol. Oral Cir. Bucal 2021, 26, e651–e660. [Google Scholar] [CrossRef] [PubMed]
- Perez, L.; Shurin, M.R.; Collins, B.; Kogan, D.; Tourkova, I.L.; Shurin, G.V. Comparative analysis of CD1a, S-100, CD83, and CD11c human dendritic cells in normal, premalignant, and malignant tissues. Histol. Histopathol. 2005, 20, 1165–1172. [Google Scholar] [PubMed]
- Agarwal, R.; Chaudhary, M.; Bohra, S.; Bajaj, S. Evaluation of natural killer cell (CD57) as a prognostic marker in oral squamous cell carcinoma: An immunohistochemistry study. J. Oral Maxillofac. Pathol. 2016, 20, 173–177. [Google Scholar] [CrossRef]
- Mele, D.; Pessino, G.; Trisolini, G.; Luchena, A.; Benazzo, M.; Morbini, P.; Mantovani, S.; Oliviero, B.; Mondelli, M.U.; Varchetta, S. Impaired intratumoral natural killer cell function in head and neck carcinoma. Front. Immunol. 2022, 13, 997806. [Google Scholar] [CrossRef]
- Taghavi, N.; Bagheri, S.; Akbarzadeh, A. Prognostic implication of CD57, CD16, and TGF-β expression in oral squamous cell carcinoma. J. Oral Pathol. Med. 2016, 45, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, G.; Bag, S.; Chakraborty, P.; Dey, D.; Roy, S.; Jain, P.; Roy, P.; Soong, R.; Majumder, P.P.; Dutt, S. Density of CD3+ and CD8+ cells in gingivo-buccal oral squamous cell carcinoma is associated with lymph node metastases and survival. PLoS ONE 2020, 15, e0242058. [Google Scholar] [CrossRef]
- Spanier, G.; Ugele, I.; Nieberle, F.; Symeou, L.; Schmidhofer, S.; Brand, A.; Meier, J.; Spoerl, S.; Krupar, R.; Rümmele, P.; et al. The predictive power of CD3+ T cell infiltration of oral squamous cell tumors is limited to non-diabetic patients. Cancer Lett. 2021, 499, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.L.; Korzinkin, M.; Zhavoronkov, A.; Ozerov, I.V.; Walker, M.T.; Higgins, K.; Lingen, M.W.; Izumchenko, E.P. A Savage Effector T cell responses unleashed by regulatory T cell ablation exacerbate oral squamous cell carcinoma. Cell Rep. Med. 2021, 2, 100399. [Google Scholar] [CrossRef]
- Kondo, Y.; Suzuki, S.; Takahara, T.; Ono, S.; Goto, M.; Miyabe, S.; Sugita, Y.; Ogawa, T.; Ito, H.; Satou, A.; et al. Improving function of cytotoxic T-lymphocytes by transforming growth factor-β inhibitor in oral squamous cell carcinoma. Cancer Sci. 2021, 112, 4037–4049. [Google Scholar] [CrossRef] [PubMed]
- Maggioni, D.; Pignataro, L.; Garavello, W. T-helper and T-regulatory cells modulation in head and neck squamous cell carcinoma. Oncoimmunology 2017, 6, e1325066. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Y.; Xie, F. T-regulatory cell/T helper 17 cell imbalance functions as prognostic biomarker of oral squamous cell carcinoma—CONSORT. Medicine 2020, 99, e23145. [Google Scholar] [CrossRef]
- Lechner, A.; Schlößer, H.A.; Thelen, M.; Wennhold, K.; Rothschild, S.I.; Gilles, R.; Quaas, A.; Siefer, O.G.; Huebbers, C.U.; Cukuroglu, E.; et al. Tumor-associated B cells and humoral immune response in head and neck squamous cell carcinoma. Oncoimmunology 2019, 8, 1535293. [Google Scholar] [CrossRef]
- Suárez-Sánchez, F.J.; Lequerica-Fernández, P.; Rodrigo, J.P.; Hermida-Prado, F.; Suárez-Canto, J.; Rodríguez-Santamarta, T.; Domínguez-Iglesias, F.; García-Pedrero, J.M.; de Vicente, J.C. Tumor-Infiltrating CD20+ B Lymphocytes: Significance and Prognostic Implications in Oral Cancer Microenvironment. Cancers 2021, 13, 395. [Google Scholar] [CrossRef]
- Hao, Z.; Li, R.; Wang, Y.; Li, S.; Hong, Z.; Han, Z. Landscape of Myeloid-derived Suppressor Cell in Tumor Immunotherapy. Biomark. Res. 2021, 9, 77. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, C.; Wang, Y.; Zhang, W.; Liu, L.; Cheng, J. Prognostic role of CD11b+ myeloid-derived suppressor cells in oral squamous cell carcinoma. Arch. Med. Sci. 2023, 19, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Fan, H.Y.; Tang, Y.L.; Wang, S.S.; Cao, M.X.; Wang, H.F.; Dai, L.L.; Wang, K.; Yu, X.H.; Wu, J.B.; et al. Myeloid derived suppressor cells contribute to the malignant progression of oral squamous cell carcinoma. PLoS ONE 2020, 15, e0229089. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, P.; Xiao, Y.; Zhang, Y.; Liu, J.; Xie, D.; Cai, M.; Zhang, X. Overexpression of α-sma-positive fibroblasts (CAFs) in Nasopharyngeal Carcinoma Predicts Poor Prognosis. J. Cancer 2017, 8, 3897–3902. [Google Scholar] [CrossRef]
- Lao, X.M.; Liang, Y.J.; Su, Y.X.; Zhang, S.E.; Zhou, X.I.; Liao, G.Q. Distribution and significance of interstitial fibrosis and stroma-infiltrating B cells in tongue squamous cell carcinoma. Oncol. Lett. 2016, 11, 2027–2034. [Google Scholar] [CrossRef]
- Patel, A.K.; Vipparthi, K.; Thatikonda, V.; Arun, I.; Bhattacharjee, S.; Sharan, R.; Arun, P.; Singh, S. A subtype of cancer-associated fibroblasts with lower expression of alpha-smooth muscle actin suppresses stemness through BMP4 in oral carcinoma. Oncogenesis 2018, 7, 78. [Google Scholar] [CrossRef]
- Gaur, P.; Qadir, G.A.; Upadhyay, S.; Singh, A.K.; Shukla, N.K.; Das, S.N. Skewed immunological balance between Th17 (CD4+IL17A+) and Treg (CD4+CD25+FOXP3+) cells in human oral squamous cell carcinoma. Cell. Oncol. 2012, 35, 335–343. [Google Scholar] [CrossRef]
- Lim, K.P.; Chun, N.A.; Ismail, S.M.; Abraham, M.T.; Yusoff, M.N.; Zain, R.B.; Ngeow, W.C.; Ponniah, S.; Cheong, S.C. CD4+CD25hiCD127low regulatory T cells are increased in oral squamous cell carcinoma patients. PLoS ONE 2014, 9, e103975. [Google Scholar] [CrossRef] [PubMed]
- Kukreja, I.; Kapoor, P.; Deshmukh, R.; Kulkarni, V. VEGF and CD34: A correlation between tumor angiogenesis and microvessel density—An immunohistochemical study. J. Oral Maxillofac. Pathol. 2013, 17, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Maqsood, A.; Ali, A.; Zaffar, Z.; Mokeem, S.; Mokeem, S.S.; Ahmed, N.; Al-Hamoudi, N.; Vohra, F.; Javed, F.; Abduljabbar, T. Expression of CD34 and α-SMA Markers in Oral Squamous Cell Carcinoma Differentiation. A Histological and Histo-Chemical Study. Int. J. Environ. Res. Public. Health 2020, 18, 192. [Google Scholar] [CrossRef]
- Teófilo, C.R.; Junior, A.E.C.F.; Batista, A.C.; Jamacaru, F.V.F.; Sousa, F.B.; Mota, M.R.L.; Silva, M.F.E.; Silva, P.G.B.; Alves, A. Mast Cells and Blood Vessels Profile in Oral Carcinogenesis: An Immunohistochemistry Study. Asian Pac. J. Cancer Prev. 2020, 21, 1097–1102. [Google Scholar] [CrossRef]
- Panneerselvam, K.; Ishikawa, S.; Krishnan, R.; Sugimoto, M. Salivary Metabolomics for Oral Cancer Detection: A Narrative Review. Metabolites 2022, 12, 436. [Google Scholar] [CrossRef] [PubMed]
- Santosh, A.B.; Jones, T.; Harvey, J. A review on oral cancer biomarkers: Understanding the past and learning from the present. J. Cancer Res. Ther. 2016, 12, 486–492. [Google Scholar] [CrossRef]
- Wang, Q.; Gao, P.; Wang, X.; Duan, Y. The early diagnosis and monitoring of squamous cell carcinoma via saliva metabolomics. Sci. Rep. 2014, 4, 6802. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Yen, C.Y.; Chen, C.H.; Tsai, J.H.; Tang, J.Y.; Chang, Y.T.; Kao, Y.H.; Wang, Y.Y.; Yuan, S.F.; Lee, S.Y. Evaluation of the mRNA expression levels of integrins α3, α5, β1 and β6 as tumor biomarkers of oral squamous cell carcinoma. Oncol. Lett. 2018, 16, 4773–4781. [Google Scholar] [CrossRef]
- Kurokawa, A.; Nagata, M.; Kitamura, N.; Noman, A.A.; Ohnishi, M.; Ohyama, T.; Kobayashi, T.; Shingaki, S.; Takagi, R. Diagnostic value of integrin α3, β4, and β5 gene expression levels for the clinical outcome of tongue squamous cell carcinoma. Cancer 2008, 112, 1272–1281. [Google Scholar] [CrossRef]
- Nagata, M.; Noman, A.A.; Suzuki, K.; Kurita, H.; Ohnishi, M.; Ohyama, T.; Kitamura, N.; Kobayashi, T.; Uematsu, K.; Takahashi, K.; et al. ITGA3 and ITGB4 expression biomarkers estimate the risks of locoregional and hematogenous dissemination of oral squamous cell carcinoma. BMC Cancer 2013, 13, 410. [Google Scholar] [CrossRef] [PubMed]
- Basheer, H.A.; Pakanavicius, E.; Cooper, P.A.; Shnyder, S.D.; Martin, L.; Hunter, K.D.; Vinader, V.; Afarinkia, K. Hypoxia modulates CCR7 expression in head and neck cancers. Oral Oncol. 2018, 80, 64–73. [Google Scholar] [CrossRef]
- Guo, N.; Liu, F.; Yang, L.; Huang, J.; Ding, X.; Sun, C. Chemokine receptor 7 enhances cell chemotaxis and migration of metastatic squamous cell carcinoma of head and neck through activation of matrix metalloproteinase-9. Oncol. Rep. 2014, 32, 794–800. [Google Scholar] [CrossRef][Green Version]
- Shang, Z.J.; Liu, K.; Shao, Z. Expression of chemokine receptor CCR7 is associated with cervical lymph node metastasis of oral squamous cell carcinoma. Oral Oncol. 2009, 45, 480–485. [Google Scholar] [CrossRef]
- Ohmura, G.; Tsujikawa, T.; Yaguchi, T.; Kawamura, N.; Mikami, S.; Sugiyama, J.; Nakamura, K.; Kobayashi, A.; Iwata, T.; Nakano, H.; et al. Aberrant Myosin 1b Expression Promotes Cell Migration and Lymph Node Metastasis of HNSCC. Mol. Cancer Res. 2015, 13, 721–731. [Google Scholar] [CrossRef]
- Sun, Z.; Guo, X.; Chen, H.; Ling, J.; Zhao, H.; Chang, A.; Zhuo, X. MYO1B as a prognostic biomarker and a therapeutic target in Arecoline-associated oral carcinoma. Mol. Carcinogen. 2023, 62, 920–939. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Okato, A.; Idichi, T.; Arai, T.; Sugawara, S.; Katada, K.; Okamoto, Y.; et al. Passenger strand of miR-145-3p acts as a tumor-suppressor by targeting MYO1B in head and neck squamous cell carcinoma. Int. J. Oncol. 2018, 52, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Maman, S.; Witz, I.P. A history of exploring cancer in context. Nat. Rev. Cancer 2018, 18, 359–376. [Google Scholar] [CrossRef] [PubMed]
- McMillin, D.W.; Negri, J.M.; Mitsiades, C.S. The role of tumour-stromal interactions in modifying drug response: Challenges and opportunities. Nat. Rev. Drug Discov. 2013, 12, 217–228. [Google Scholar] [CrossRef]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef]
- De Palma, M.; Biziato, D.; Petrova, T.V. Microenvironmental regulation of tumour angiogenesis. Nat. Rev. Cancer 2017, 17, 457–474. [Google Scholar] [CrossRef]
- Valkenburg, K.C.; de Groot, A.E.; Pienta, K.J. Targeting the tumour stroma to improve cancer therapy. Nat. Rev. Clin. Oncol. 2018, 15, 366–381. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Jain, R.K.; Langer, R. Engineering and physical sciences in oncology: Challenges and opportunities. Nat. Rev. Cancer 2017, 17, 659–675. [Google Scholar] [CrossRef]
- Li, M.; He, L.; Zhu, J.; Zhang, P.; Liang, S. Targeting tumor-associated macrophages for cancer treatment. Cell Biosci. 2022, 12, 85. [Google Scholar] [CrossRef]
- Yan, M.; Jurasz, P. The role of platelets in the tumor microenvironment: From solid tumors to leukemia. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2016, 1863, 392–400. [Google Scholar] [CrossRef]
- Ramamonjisoa, N.; Ackerstaff, E. Characterization of the Tumor Microenvironment and Tumor-Stroma Interaction by Non-Invasive Preclinical Imaging. Front. Oncol. 2017, 7, 3. [Google Scholar] [CrossRef]
- Bellomo, C.; Caja, L.; Moustakas, A. Transforming growth factor β as regulator of cancer stemness and metastasis. Br. J. Cancer 2016, 115, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D cell cultures—A comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef]
- Sapudom, J.; Müller, C.D.; Nguyen, K.-T.; Martin, S.; Anderegg, U.; Pompe, T. Matrix Remodeling and Hyaluronan Production by Myofibroblasts and Cancer-Associated Fibroblasts in 3D Collagen Matrices. Gels 2020, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tao, D.; Zhang, P.; Liu, X.; Zhang, Y.; Cheng, J.; Yuan, H.; Liu, L.; Jiang, H. Hyaluronan synthase 2 expressed by cancer-associated fibroblasts promotes oral cancer invasion. J. Exp. Clin. Cancer Res. 2016, 35, 181. [Google Scholar] [CrossRef]
- Elmusrati, A.A.; Pilborough, A.E.; Khurram, S.A.; Lambert, D.W. Cancer-associated fibroblasts promote bone invasion in oral squamous cell carcinoma. Br. J. Cancer 2017, 117, 867–875. [Google Scholar] [CrossRef]
- Lin, N.N.; Wang, P.; Zhao, D.; Zhang, F.J.; Yang, K.; Chen, R. Significance of oral cancer-associated fibroblasts in angiogenesis, lymphangiogenesis, and tumor invasion in oral squamous cell carcinoma. J. Oral Pathol. Med. 2017, 46, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Russmueller, G.; Moser, D.; Würger, T.; Wrba, F.; Christopoulos, P.; Kostakis, G.; Seemann, R.; Stadler, V.; Wimmer, G.; Kornek, G.; et al. Upregulation of osteoprotegerin expression correlates with bone invasion and predicts poor clinical outcome in oral cancer. Oral Oncol. 2015, 51, 247–253. [Google Scholar] [CrossRef]
- Sambandam, Y.; Ethiraj, P.; Hathaway-Schrader, J.D.; Novince, C.M.; Panneerselvam, E.; Sundaram, K.; Reddy, S.V. Autoregulation of RANK ligand in oral squamous cell carcinoma tumor cells. J. Cell. Physiol. 2018, 233, 6125–6134. [Google Scholar] [CrossRef]
- Bae, J.Y.; Kim, E.K.; Yang, D.H.; Zhang, X.; Park, Y.J.; Lee, D.Y.; Che, C.M.; Kim, J. Reciprocal interaction between carcinoma-associated fibroblasts and squamous carcinoma cells through interleukin-1α induces cancer progression. Neoplasia 2014, 16, 928–938. [Google Scholar] [CrossRef]
- Korbecki, J.; Bosiacki, M.; Barczak, K.; Łagocka, R.; Chlubek, D.; Baranowska-Bosiacka, I. The Clinical Significance and Role of CXCL1 Chemokine in Gastrointestinal Cancers. Cells 2023, 12, 1406. [Google Scholar] [CrossRef]
- Shishido, K.; Kuroishi, T.; Sugawara, S. P2 purinergic receptor signaling and interleukin-1 synergistically induce interleukin-6 production in a human oral squamous carcinoma cell line. J. Oral Biosci. 2021, 63, 80–90. [Google Scholar] [CrossRef]
- Brady, G.; Crean, S.J.; Naik, P.; Kapas, S. Upregulation of IGF-2 and IGF-1 receptor expression in oral cancer cell lines. Int. J. Oncol. 2007, 31, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Cacheux, W.; Lièvre, A.; Richon, S.; Vacher, S.; El Alam, E.; Briaux, A.; El Botty, R.; Mariani, P.; Buecher, B.; Schnitzler, A.; et al. Dangles-Marie Interaction between IGF2-PI3K axis and cancer-associated-fibroblasts promotes anal squamous carcinogenesis. Int. J. Cancer 2019, 145, 1852–1859. [Google Scholar] [CrossRef] [PubMed]
- Ferreira Mendes, J.M.; de Faro Valverde, L.; Vidal, M.T.A.; Paredes, B.D.; Coelho, P.; Allahdadi, K.J.; Coletta, R.D.; Souza, B.S.F.; Rocha, C.A.G. Effects of IGF-1 on Proliferation, Angiogenesis, Tumor Stem Cell Populations and Activation of AKT and Hedgehog Pathways in Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2020, 21, 6487. [Google Scholar] [CrossRef] [PubMed]
- Kayamori, K.; Katsube, K.; Sakamoto, K.; Ohyama, Y.; Hirai, H.; Yukimori, A.; Ohata, Y.; Akashi, T.; Saitoh, M.; Harada, K.; et al. NOTCH3 Is Induced in Cancer-Associated Fibroblasts and Promotes Angiogenesis in Oral Squamous Cell Carcinoma. PLoS ONE 2016, 11, e0154112. [Google Scholar] [CrossRef]
- Porcheri, C.; Meisel, C.T.; Mitsiadis, T. Multifactorial Contribution of Notch Signaling in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2019, 20, 1520. [Google Scholar] [CrossRef]
- Sun, W.; Gaykalova, D.A.; Ochs, M.F.; Mambo, E.; Arnaoutakis, D.; Liu, Y.; Loyo, M.; Agrawal, N.; Howard, J.; Li, R.; et al. Activation of the NOTCH pathway in head and neck cancer. Cancer Res. 2014, 74, 1091–1104. [Google Scholar] [CrossRef]
- Karakasheva, T.A.; Lin, E.W.; Tang, Q.; Qiao, E.; Waldron, T.J.; Soni, M.; Klein-Szanto, A.J.; Sahu, V.; Basu, D.; Ohashi, S.; et al. IL-6 Mediates Cross-Talk between Tumor Cells and Activated Fibroblasts in the Tumor Microenvironment. Cancer Res. 2018, 78, 4957–4970. [Google Scholar] [CrossRef]
- Mirkeshavarz, M.; Ganjibakhsh, M.; Aminishakib, P.; Farzaneh, P.; Mahdavi, N.; Vakhshiteh, F.; Karimi, A.; Gohari, N.S.; Kamali, F.; Kharazifard, M.J.; et al. Interleukin-6 secreted by oral cancer- associated fibroblast accelerated VEGF expression in tumor and stroma cells. Cell. Mol. Biol. 2017, 63, 131–136. [Google Scholar] [CrossRef]
- Qin, X.; Yan, M.; Wang, X.; Xu, Q.; Wang, X.; Zhu, X.; Shi, J.; Li, Z.; Zhang, J.; Chen, W. Cancer-associated Fibroblast-derived IL-6 Promotes Head and Neck Cancer Progression via the Osteopontin-NF-κB Signaling Pathway. Theranostics 2018, 8, 921–940. [Google Scholar] [CrossRef]
- Li, X.; Bu, W.; Meng, L.; Liu, X.; Wang, S.; Jiang, L.; Ren, M.; Fan, Y.; Sun, H. CXCL12/CXCR4 pathway orchestrates CSC-like properties by CAF recruited tumor associated macrophage in OSCC. Exp. Cell Res. 2019, 378, 131–138. [Google Scholar] [CrossRef]
- Wu, M.H.; Hong, H.C.; Hong, T.M.; Chiang, W.F.; Jin, Y.T.; Chen, Y.L. Targeting galectin-1 in carcinoma-associated fibroblasts inhibits oral squamous cell carcinoma metastasis by downregulating MCP-1/CCL2 expression. Clin. Cancer Res. 2011, 17, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Chiu, J.W.; Hanafi, Z.B.; Chew, L.C.Y.; Mei, Y.; Liu, H. IL-1α Processing, Signaling and Its Role in Cancer Progression. Cells 2021, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.-W.; Che, Z.M.; Kim, J.; Kim, K.; Kim, K.-Y.; Williams, D.; Kim, J. Tumor-stromal crosstalk in invasion of oral squamous cell carcinoma: A pivotal role of CCL7. Int. J. Cancer 2010, 127, 332–344. [Google Scholar] [CrossRef]
- Sergi, B. Remodeling the tumor microenvironment to overcome treatment resistance in HPV-negative head and neck cancer. Cancer Drug Resist. 2023, 6, 291–313. [Google Scholar] [CrossRef]
- Takahashi, H.; Sakakura, K.; Kudo, T.; Toyoda, M.; Kaira, K.; Oyama, T.; Chikamatsu, K. Cancer-associated fibroblasts promote an immunosuppressive microenvironment through the induction and accumulation of protumoral macrophages. Oncotarget 2017, 8, 8633–8647. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Moriyama, M.; Furukawa, S.; Rafiul, H.; Maruse, Y.; Jinno, T.; Tanaka, A.; Ohta, M.; Ishiguro, N.; Yamauchi, M.; et al. CD163+CD204+ tumor-associated macrophages contribute to T cell regulation via interleukin-10 and PD-L1 production in oral squamous cell carcinoma. Sci. Rep. 2017, 7, 1755. [Google Scholar] [CrossRef]
- Wu, T.; Tang, C.; Tao, R.; Yong, X.; Jiang, Q.; Feng, C. PD-L1-Mediated Immunosuppression in Oral Squamous Cell Carcinoma: Relationship With Macrophage Infiltration and Epithelial to Mesenchymal Transition Markers. Front. Immunol. 2021, 12, 693881. [Google Scholar] [CrossRef]
- O’Sullivan, C.; Lewis, C.E.; Harris, A.L.; McGee, J.O. Secretion of epidermal growth factor by macrophages associated with breast carcinoma. Lancet 1993, 342, 148–149. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.D.; Haridas, N.; Patel, J.B.; Shah, F.D.; Shukla, S.N.; Shah, P.M.; Patel, P.S. Matrix metalloproteinases and their inhibitors: Correlation with invasion and metastasis in oral cancer. Indian J. Clin. Biochem. 2010, 25, 250–259. [Google Scholar] [CrossRef]
- Alves, A.; Diel, L.; Ramos, G.; Pinto, A.; Bernardi, L.; Yates, J., 3rd; Lamers, M. Tumor microenvironment and Oral Squamous Cell Carcinoma: A crosstalk between the inflammatory state and tumor cell migration. Oral Oncol. 2021, 112, 105038. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.X.; Wang, S.; Zhao, H.; Liu, N.; Chen, D.; Sun, M.; Zheng, J.H. Sonic hedgehog signaling may promote invasion and metastasis of oral squamous cell carcinoma by activating MMP-9 and E-cadherin expression. Med. Oncol. 2014, 31, 41. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; He, M.Y.; Zhu, L.F.; Yang, C.C.; Zhou, M.L.; Wang, Q.; Zhang, W.; Zheng, Y.Y.; Wang, D.M.; Xu, Z.Q.; et al. Liu Tumor-associated macrophages correlate with the clinicopathological features and poor outcomes via inducing epithelial to mesenchymal transition in oral squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2016, 35, 12. [Google Scholar] [CrossRef]
- Petty, A.J.; Li, A.; Wang, X.; Dai, R.; Heyman, B.; Hsu, D.; Huang, X.; Yang, Y. Hedgehog signaling promotes tumor-associated macrophage polarization to suppress intratumoral CD8+ T cell recruitment. J. Clin. Investig. 2019, 129, 5151–5162. [Google Scholar] [CrossRef] [PubMed]
- Valverde, L.d.F.; Pereira, T.D.A.; Dias, R.B.; Guimarães, V.S.N.; Ramos, E.A.G.; Santos, J.N.; Rocha, C.A.G. Macrophages and endothelial cells orchestrate tumor-associated angiogenesis in oral cancer via hedgehog pathway activation. Tumor Biol. 2016, 37, 9233–9241. [Google Scholar] [CrossRef]
- Guo, Y.; Xu, T.; Chai, Y.; Chen, F. TGF-β Signaling in Progression of Oral Cancer. Int. J. Mol. Sci. 2023, 24, 10263. [Google Scholar] [CrossRef]
- Sun, H.; Miao, C.; Liu, W.; Qiao, X.; Yang, W.; Li, L. Li TGF-β1/TβRII/Smad3 signaling pathway promotes VEGF expression in oral squamous cell carcinoma tumor-associated macrophages. Biochem. Biophys. Res. Commun. 2018, 497, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Mao, X.H.; Tian, T.; Wang, W.M.; Su, T.; Jiang, C.H.; Hu, C.Y. Role of PFKFB3 and CD163 in Oral Squamous Cell Carcinoma Angiogenesis. Curr. Med. Sci. 2019, 39, 410–414. [Google Scholar] [CrossRef]
- Shi, L.; Pan, H.; Liu, Z.; Xie, J.; Han, W. Roles of PFKFB3 in cancer. Signal Transduct. Target. Ther. 2017, 2, 17044. [Google Scholar] [CrossRef]
- Han, N.; Li, X.; Wang, Y.; Wang, L.; Zhang, C.; Zhang, Z.; Ruan, M.; Zhang, C. Increased tumor-infiltrating plasmacytoid dendritic cells promote cancer cell proliferation and invasion via TNF-α/NF-κB/CXCR-4 pathway in oral squamous cell carcinoma. J. Cancer 2021, 12, 3045–3056. [Google Scholar] [CrossRef]
- Rehman, A.O.; Wang, C.Y. CXCL12/SDF-1α activates NF-κB and promotes oral cancer invasion through the Carma3/Bcl10/Malt1 complex. Int. J. Oral Sci. 2009, 1, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.C.; Moreira, A.F.; de Melo-Diogo, D.; Gaspar, V.M.; Carvalho, M.P.; Correia, I.J. 3D tumor spheroids: An overview on the tools and techniques used for their analysis. Biotechnol. Adv. 2016, 34, 1427–1441. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Encarnación-Rosado, J.; Lin, E.Y.; Sohn, A.S.W.; Zhang, H.; Mancias, J.D.; Kimmelman, A.C. Autophagy supports mitochondrial metabolism through the regulation of iron homeostasis in pancreatic cancer. Sci. Adv. 2023, 9, eadf9284. [Google Scholar] [CrossRef] [PubMed]
- Dinić, M.; Pecikoza, U.; Djokić, J.; Stepanović-Petrović, R.; Milenković, M.; Stevanović, M.; Filipović, N.; Begović, J.; Golić, N.; Lukić, J. Exopolysaccharide Produced by Probiotic Strain Lactobacillus paraplantarum BGCG11 Reduces Inflammatory Hyperalgesia in Rats. Front. Pharmacol. 2018, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Pinto, B.; Henriques, A.C.; Silva, P.M.A.; Bousbaa, H. Three-Dimensional Spheroids as In Vitro Preclinical Models for Cancer Research. Pharmaceutics 2020, 12, 1186. [Google Scholar] [CrossRef]
- Pozzi, S.; Scomparin, A.; Dangoor, S.I.; Ajamil, D.R.; Ofek, P.; Neufeld, L.; Krivitsky, A.; Vaskovich-Koubi, D.; Kleiner, R.; Dey, P.; et al. Meet me halfway: Are in vitro 3D cancer models on the way to replace in vivo models for nanomedicine development? Adv. Drug Deliv. Rev. 2021, 175, 113760. [Google Scholar] [CrossRef]
- Abdolahinia, E.D.; Ahmadian, S.; Bohlouli, S.; Gharehbagh, F.J.; Jahandizi, N.G.; Vahed, S.Z.; Saadat, Y.R.; Aghbali, A.; Sharifi, S.; Dizaj, S.M.; et al. Effect of Curcumin on the Head and Neck Squamous Cell Carcinoma Cell Line HN5. Curr. Mol. Pharmacol. 2023, 16, 374–380. [Google Scholar] [PubMed]
- Memar, M.Y.; Abdolahinia, E.D.; Yekani, M.; Kouhsoltani, M.; Sharifi, S.; Dizaj, S.M. Preparation of rutin-loaded mesoporous silica nanoparticles and evaluation of its physicochemical, anticancer, and antibacterial properties. Mol. Biol. Rep. 2023, 50, 203–213. [Google Scholar] [CrossRef]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef]
- Baker, B.M.; Chen, C.S. Deconstructing the third dimension: How 3D culture microenvironments alter cellular cues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [CrossRef] [PubMed]
- Pampaloni, F.; Reynaud, E.G.; Stelzer, E.H.K. The third dimension bridges the gap between cell culture and live tissue. Nat. Rev. Mol. Cell Biol. 2007, 8, 839–845. [Google Scholar] [CrossRef]
- Hickman, J.A.; Graeser, R.; de Hoogt, R.; Vidic, S.; Brito, C.; Gutekunst, M.; van der Kuip, H. Three-dimensional models of cancer for pharmacology and cancer cell biology: Capturing tumor complexity in vitro/ex vivo. Biotechnol. J. 2014, 9, 1115–1128. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.E.; Miller, F.R.; Heppner, G.H. Factors affecting growth and drug sensitivity of mouse mammary tumor lines in collagen gel cultures. Cancer Res. 1985, 45, 4200–4205. [Google Scholar] [PubMed]
- Miyamoto, S.; Nishikiori, N.; Sato, T.; Watanabe, M.; Umetsu, A.; Tsugeno, Y.; Hikage, F.; Sasaya, T.; Kato, H.; Ogi, K.; et al. Three-Dimensional Spheroid Configurations and Cellular Metabolic Properties of Oral Squamous Carcinomas Are Possible Pharmacological and Pathological Indicators. Cancers 2023, 15, 2793. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Sato, K.; Nakamura, T.; Yoshida, Y.; Murata, S.; Yoshida, K.; Kanemoto, H.; Umemori, K.; Kawai, H.; Obata, K.; et al. Reproduction of the Antitumor Effect of Cisplatin and Cetuximab Using a Three-dimensional Spheroid Model in Oral Cancer. Int. J. Med. Sci. 2022, 19, 1320–1333. [Google Scholar] [CrossRef]
- Vincent-Chong, V.K.; Seshadri, M. Development and Radiation Response Assessment in A Novel Syngeneic Mouse Model of Tongue Cancer: 2D Culture, 3D Organoids and Orthotopic Allografts. Cancers 2020, 12, 579. [Google Scholar] [CrossRef]
- Howard, D.; Buttery, L.D.; Shakesheff, K.M.; Roberts, S.J. Tissue engineering: Strategies, stem cells and scaffolds. J. Anat. 2008, 213, 66–72. [Google Scholar] [CrossRef]
- Ovsianikov, A.; Khademhosseini, A.; Mironov, V. The synergy of scaffold-based and scaffold-free tissue engineering strategies. Trends Biotechnol. 2018, 36, 348–357. [Google Scholar] [CrossRef]
- Lee, J.; You, J.H.; Shin, D.; Roh, J.L. Ex vivo culture of head and neck cancer explants in cell sheet for testing chemotherapeutic sensitivity. J. Cancer Res. Clin. Oncol. 2020, 146, 2497–2507. [Google Scholar] [CrossRef]
- Sekine, W.; Haraguchi, Y.; Shimizu, T.; Umezawa, A.; Okano, T. Thickness limitation and cell viability of multi-layered cell sheets and overcoming the diffusion limit by a porous-membrane culture insert. J. Biochips Tissue Chips 2011, 1, 2153–2777. [Google Scholar] [CrossRef]
- Sant, S.; Johnston, P.A. The production of 3D tumor spheroids for cancer drug discovery. Drug Discov. Today Technol. 2017, 23, 27–36. [Google Scholar] [CrossRef]
- Rodrigues, T.; Kundu, B.; Silva-Correia, J.; Kundu, S.C.; Oliveira, J.M.; Reis, R.L.; Correlo, V.M. Emerging tumor spheroids technologies for 3D in vitro cancer modeling. Pharmacol. Ther. 2018, 184, 201–211. [Google Scholar] [CrossRef]
- Friedrich, J.; Ebner, R.; Kunz-Schughart, L.A. Experimental anti-tumor therapy in 3-D: Spheroids—Old hat or new challenge? Int. J. Radiat. Biol. 2007, 83, 849–871. [Google Scholar] [CrossRef]
- Chitturi Suryaprakash, R.T.; Kujan, O.; Shearston, K.; Farah, C.S. Three-Dimensional Cell Culture Models to Investigate Oral Carcinogenesis: A Scoping Review. Int. J. Mol. Sci. 2020, 21, 9520. [Google Scholar] [CrossRef]
- Lee, C.; Lee, C.; Atakilit, A.; Siu, A.; Ramos, D.M. Differential spheroid formation by oral cancer cells. Anticancer Res. 2014, 34, 6945–6949. [Google Scholar] [PubMed]
- Singh, S.P.; Sharma, M.; Gupta, P.K. Evaluation of Phototoxic Effects of Curcumin Loaded in Organically Modified Silica Nanoparticles in Tumor Spheroids of Oral Cancer Cells. BioNanoScience 2015, 5, 10–21. [Google Scholar] [CrossRef]
- Laschke, M.W.; Menger, M.D. Life is 3D: Boosting Spheroid Function for Tissue Engineering. Trends Biotechnol. 2017, 35, 133–144. [Google Scholar] [CrossRef]
- Yakavets, I.; Jenard, S.; Francois, A.; Maklygina, Y.; Loschenov, V.; Lassalle, H.-P.; Dolivet, G.; Bezdetnaya, L. Stroma-Rich Co-Culture Multicellular Tumor Spheroids as a Tool for Photoactive Drugs Screening. J. Clin. Med. 2019, 8, 1686. [Google Scholar] [CrossRef]
- Fang, Y.; Eglen, R.M. Three-Dimensional Cell Cultures in Drug Discovery and Development. SLAS Discov. 2017, 22, 456–472. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Demirci, U.; Chen, P. Emerging organoid models: Leaping forward in cancer research. J. Hematol. Oncol. 2019, 12, 142. [Google Scholar] [CrossRef]
- Driehuis, E.; Kolders, S.; Spelier, S.; Lõhmussaar, K.; Willems, S.M.; Devriese, L.A.; de Bree, R.; de Ruiter, E.J.; Korving, J.; Begthel, H. Oral mucosal organoids as a potential platform for personalized cancer therapy. Cancer Discov. 2019, 9, 852–871. [Google Scholar] [CrossRef] [PubMed]
- Saglam-Metiner, P.; Gulce-Iz, S.; Biray-Avci, C. Bioengineering-inspired three-dimensional culture systems: Organoids to create tumor microenvironment. Gene 2019, 686, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Sitarski, A.M.; Fairfield, H.; Falank, C.; Reagan, M.R. 3D Tissue Engineered In Vitro Models of Cancer in Bone. ACS Biomater. Sci. Eng. 2018, 4, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Tayebi, L.; Moharamzadeh, K. Biomaterials for Oral and Dental Tissue Engineering; Woodhead Publishing: Sawston, UK, 2017. [Google Scholar]
- Colley, H.E.; Hearnden, V.; Jones, A.V.; Weinreb, P.H.; Violette, S.M.; MacNeil, S.; Thornhill, M.H.; Murdoch, C. Development of tissue-engineered models of oral dysplasia and early invasive oral squamous cell carcinoma. Br. J. Cancer 2011, 105, 1582–1592. [Google Scholar] [CrossRef]
- Costea, D.E.; Kulasekara, K.; Neppelberg, E.; Johannessen, A.C.; Vintermyr, O.K. Species-specific fibroblasts required for triggering invasiveness of partially transformed oral keratinocytes. Am. J. Pathol. 2006, 168, 1889–1897. [Google Scholar] [CrossRef]
- Gaballah, K.; Costea, D.; Hills, A.; Gollin, S.; Harrison, P.; Partridge, M. Tissue engineering of oral dysplasia. J. Pathol. 2008, 215, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Sawant, S.; Dongre, H.; Singh, A.K.; Joshi, S.; Costea, D.E.; Mahadik, S.; Ahire, C.; Makani, V.; Dange, P.; Sharma, S.; et al. Establishment of 3D Co-Culture Models from Different Stages of Human Tongue Tumorigenesis: Utility in Understanding Neoplastic Progression. PLoS ONE 2016, 11, e0160615. [Google Scholar] [CrossRef]
- Brancato, V.; Oliveira, J.M.; Correlo, V.M.; Reis, R.L.; Kundu, S.C. Could 3D models of cancer enhance drug screening? Biomaterials 2020, 232, 119744. [Google Scholar] [CrossRef]
- Al-Samadi, A.; Poor, B.; Tuomainen, K.; Liu, V.; Hyytiäinen, A.; Suleymanova, I.; Mesimaki, K.; Wilkman, T.; Mäkitie, A.; Saavalainen, P.; et al. In vitro humanized 3D microfluidic chip for testing personalized immunotherapeutics for head and neck cancer patients. Exp. Cell Res. 2019, 383, 111508. [Google Scholar] [CrossRef]
- Bower, R.; Green, V.L.; Kuvshinova, E.; Kuvshinov, D.; Karsai, L.; Crank, S.T.; Stafford, N.D.; Greenman, J. Maintenance of head and neck tumor on-chip: Gateway to personalized treatment? Future Sci. OA 2017, 3, FSO174. [Google Scholar] [CrossRef] [PubMed]
- Duzagac, F.; Saorin, G.; Memeo, L.; Canzonieri, V.; Rizzolio, F. Microfluidic Organoids-on-a-Chip: Quantum Leap in Cancer Research. Cancers 2021, 13, 737. [Google Scholar] [CrossRef]
- Hattersley, S.M.; Sylvester, D.C.; Dyer, C.E.; Stafford, N.D.; Haswell, S.J.; Greenman, J. A microfluidic system for testing the responses of head and neck squamous cell carcinoma tissue biopsies to treatment with chemotherapy drugs. Ann. Biomed. Eng. 2012, 40, 1277–1288. [Google Scholar] [CrossRef] [PubMed]
- Lugo-Cintrón, K.M.; Ayuso, J.M.; Humayun, M.; Gong, M.M.; Kerr, S.C.; Ponik, S.M.; Harari, P.M.; Virumbrales-Muñoz, M.; Beebe, D.J. Primary head and neck tumour-derived fibroblasts promote lymphangiogenesis in a lymphatic organotypic co-culture model. EBioMedicine 2021, 73, 103634. [Google Scholar] [CrossRef]
- Akgönüllü, S.; Bakhshpour, M.; Pişkin, A.K.; Denizli, A. Microfluidic Systems for Cancer Diagnosis and Applications. Micromachines 2021, 12, 1349. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, H. Microfluidic Technologies for Head and Neck Cancer: From Single-Cell Analysis to Tumor-on-a-Chip. In Early Detection and Treatment of Head & Neck Cancers: Practical Applications and Techniques for Detection, Diagnosis, and Treatment; Springer: Berlin/Heidelberg, Germany, 2021; pp. 43–62. [Google Scholar] [CrossRef]
- Zhao, H.; Chu, M.; Huang, Z.; Yang, X.; Ran, S.; Hu, B.; Zhang, C.; Liang, J. Variations in oral microbiota associated with oral cancer. Sci. Rep. 2017, 7, 11773. [Google Scholar] [CrossRef]
- Irani, S.; Barati, I.; Badiei, M. Periodontitis and oral cancer—Current concepts of the etiopathogenesis. Oncol. Rev. 2020, 14, 465. [Google Scholar] [CrossRef]
- Isola, G.; Giudice, A.L.; Polizzi, A.; Alibrandi, A.; Murabito, P.; Indelicato, F. Identification of the different salivary Interleukin-6 profiles in patients with periodontitis: A cross-sectional study. Arch. Oral Biol. 2021, 122, 104997. [Google Scholar] [CrossRef]
- Mark Welch, J.L.; Dewhirst, F.E.; Borisy, G.G. Biogeography of the Oral Microbiome: The Site-Specialist Hypothesis. Annu. Rev. Microbiol. 2019, 73, 335–358. [Google Scholar] [CrossRef]
- Kim, J.Y.; An, C.H.; Kim, J.Y.; Jung, J.K. Experimental Animal Model Systems for Understanding Salivary Secretory Disorders. Int. J. Mol. Sci. 2020, 21, 8423. [Google Scholar] [CrossRef]
- Olek, M.; Machorowska-Pieniążek, A.; Olek, K.; Cieślar, G.; Kawczyk-Krupka, A. Photodynamic therapy in the treatment of oral squamous cell carcinoma—The state of the art in preclinical research on the animal model. Photodiagn. Photodyn. Ther. 2021, 34, 102236. [Google Scholar] [CrossRef] [PubMed]
- Toma, A.I.; Fuller, J.M.; Willett, N.J.; Goudy, S.L. Oral wound healing models and emerging regenerative therapies. Transl. Res. 2021, 236, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Johnston, W.; Delaney, C.; Rajendran, R.; Butcher, J.; Khan, S.; Bradshaw, D.; Ramage, G.; Culshaw, S. Biofilm-stimulated epithelium modulates the inflammatory responses in co-cultured immune cells. Sci. Rep. 2019, 9, 15779. [Google Scholar] [CrossRef] [PubMed]
- Dongari-Bagtzoglou, A. Pathogenesis of mucosal biofilm infections: Challenges and progress. Exp. Rev. Anti-Infect. Ther. 2008, 6, 201–208. [Google Scholar] [CrossRef]
- Valm, A.M. The Structure of Dental Plaque Microbial Communities in the Transition from Health to Dental Caries and Periodontal Disease. J. Mol. Biol. 2019, 431, 2957–2969. [Google Scholar] [CrossRef]
- Simian, M.; Bissell, M.J. Organoids: A historical perspective of thinking in three dimensions. J. Cell Biol. 2017, 216, 31–40. [Google Scholar] [CrossRef]
- Ohnemus, U.; Willers, C.; Bubenheim, M.; Horstkotte, M.A.; Houdek, P.; Fischer, F.; Schmage, P.; Moll, I.; Brandner, J.M. An ex-vivo oral mucosa infection model for the evaluation of the topical activity of antifungal agents. Mycoses 2008, 51, 21–29. [Google Scholar] [CrossRef]
- Sacks, P.G. Cell, tissue and organ culture as in vitro models to study the biology of squamous cell carcinomas of the head and neck. Cancer Metastasis Rev. 1996, 15, 27–51. [Google Scholar] [CrossRef]
- Gibbs, S.; Roffel, S.; Meyer, M.; Gasser, A. Biology of soft tissue repair: Gingival epithelium in wound healing and attachment to the tooth and abutment surface. Eur. Cells Mater. 2019, 38, 63–78. [Google Scholar] [CrossRef]
- Moutsopoulos, N.M.; Konkel, J.E. Tissue-Specific Immunity at the Oral Mucosal Barrier. Trends Immunol. 2018, 39, 276–287. [Google Scholar] [CrossRef]
- Takahashi, N.; Sulijaya, B.; Yamada-Hara, M.; Tsuzuno, T.; Tabeta, K.; Yamazaki, K. Gingival epithelial barrier: Regulation by beneficial and harmful microbes. Tissue Barriers 2019, 7, e1651158. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.J.; Wilson, M.J.; Wei, X.; Lewis, M.A.O.; Bradshaw, D.J.; Murdoch, C.; Williams, D.W. Denture-associated biofilm infection in three-dimensional oral mucosal tissue models. J. Med. Microbiol. 2018, 67, 364–375. [Google Scholar] [CrossRef]
- Souza, J.G.S.; Bertolini, M.; Thompson, A.; Barão, V.A.R.; Dongari-Bagtzoglou, A. Biofilm Interactions of Candida albicans and Mitis Group Streptococci in a Titanium-Mucosal Interface Model. Appl. Environ. Microbiol. 2020, 86, e02950-19. [Google Scholar] [CrossRef]
- Beklen, A.; Torittu, A.; Ihalin, R.; Pöllänen, M. Aggregatibacter actinomycetemcomitans Biofilm Reduces Gingival Epithelial Cell Keratin Expression in an Organotypic Gingival Tissue Culture Model. Pathogens 2019, 8, 278. [Google Scholar] [CrossRef]
- Ingendoh-Tsakmakidis, A.; Mikolai, C.; Winkel, A.; Szafrański, S.P.; Falk, C.S.; Rossi, A.; Walles, H.; Stiesch, M. Commensal and pathogenic biofilms differently modulate peri-implant oral mucosa in an organotypic model. Cell. Microbiol. 2019, 21, e13078. [Google Scholar] [CrossRef] [PubMed]
- Bugueno, I.M.; Batool, F.; Keller, L.; Kuchler-Bopp, S.; Benkirane-Jessel, N.; Huck, O. Porphyromonas gingivalis bypasses epithelial barrier and modulates fibroblastic inflammatory response in an in vitro 3D spheroid model. Sci. Rep. 2018, 8, 14914. [Google Scholar] [CrossRef]
- Dongari-Bagtzoglou, A.; Kashleva, H. Development of a highly reproducible three-dimensional organotypic model of the oral mucosa. Nat. Protoc. 2006, 1, 2012–2018. [Google Scholar] [CrossRef]
- Pinnock, A.; Murdoch, C.; Moharamzadeh, K.; Whawell, S.; Douglas, C.W. Characterisation and optimisation of organotypic oral mucosal models to study Porphyromonas gingivalis invasion. Microbes Infect. 2014, 16, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Iannelli, F.; Zotti, A.I.; Roca, M.S.; Grumetti, L.; Lombardi, R.; Moccia, T.; Vitagliano, C.; Milone, M.R.; Ciardiello, C.; Bruzzese, F.; et al. Valproic Acid Synergizes With Cisplatin and Cetuximab in vitro and in vivo in Head and Neck Cancer by Targeting the Mechanisms of Resistance. Front. Cell Dev. Biol. 2020, 8, 732. [Google Scholar] [CrossRef]
- Melissaridou, S.; Wiechec, E.; Magan, M.; Jain, M.V.; Chung, M.K.; Farnebo, L.; Roberg, K. The effect of 2D and 3D cell cultures on treatment response, EMT profile and stem cell features in head and neck cancer. Cancer Cell Int. 2019, 19, 16. [Google Scholar] [CrossRef]
- Van Vuuren, R.J.; Botes, M.; Jurgens, T.; Joubert, A.M.; van den Bout, I. Novel sulphamoylated 2-methoxy estradiol derivatives inhibit breast cancer migration by disrupting microtubule turnover and organization. Cancer Cell Int. 2019, 19, 1. [Google Scholar] [CrossRef] [PubMed]
- Azharuddin, M.; Roberg, K.; Dhara, A.K.; Jain, M.V.; Darcy, P.; Hinkula, J.; Slater, N.K.H.; Patra, H.K. Dissecting multi drug resistance in head and neck cancer cells using multicellular tumor spheroids. Sci. Rep. 2019, 9, 20066. [Google Scholar] [CrossRef] [PubMed]
- Perche, F.; Torchilin, V.P. Cancer cell spheroids as a model to evaluate chemotherapy protocols. Cancer Biol. Ther. 2012, 13, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.G.; da Rocha, G.O.; de Andrade, J.B. Occurrence of the potent mutagens 2-nitrobenzanthrone and 3-nitrobenzanthrone in fine airborne particles. Sci. Rep. 2019, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, J.; Jacobi, C.; Gstoettner, S.; Welz, C.; Schwenk-Zieger, S.; Stauber, R.; Strieth, S.; Kuenzel, J.; Baumeister, P.; Becker, S. Therapy testing in a spheroid-based 3D cell culture model for head and neck squamous cell carcinoma. J. Vis. Exp 2018, 134, e57012. [Google Scholar] [CrossRef]
- Hagemann, J.; Jacobi, C.; Hahn, M.; Schmid, V.; Welz, C.; Schwenk-Zieger, S.; Stauber, R.; Baumeister, P.; Becker, S. Spheroid-based 3D Cell Cultures Enable Personalized Therapy Testing and Drug Discovery in Head and Neck Cancer. Anticancer Res. 2017, 37, 2201–2210. [Google Scholar] [CrossRef]
- Magan, M.; Wiechec, E.; Roberg, K. CAFs affect the proliferation and treatment response of head and neck cancer spheroids during co-culturing in a unique in vitro model. Cancer Cell Int. 2020, 20, 599. [Google Scholar] [CrossRef]
- Lin, K.-C.; Ting, L.-L.; Chang, C.-L.; Lu, L.-S.; Lee, H.-L.; Hsu, F.-C.; Chiou, J.-F.; Wang, P.-Y.; Burnouf, T.; Ho, D.C.-Y.; et al. Ex Vivo Expanded Circulating Tumor Cells for Clinical Anti-Cancer Drug Prediction in Patients with Head and Neck Cancer. Cancers 2021, 13, 6076. [Google Scholar] [CrossRef]
- Tanaka, N.; Osman, A.A.; Takahashi, Y.; Lindemann, A.; Patel, A.A.; Zhao, M.; Takahashi, H.; Myers, J.N. Head and neck cancer organoids established by modification of the CTOS method can be used to predict in vivo drug sensitivity. Oral Oncol. 2018, 87, 49–57. [Google Scholar] [CrossRef]
- Zhao, L.; Huang, L.; Yu, S.; Zheng, J.; Wang, H.; Zhang, Y. Decellularized tongue tissue as an in vitro model for studying tongue cancer and tongue regeneration. Acta Biomater. 2017, 58, 122–135. [Google Scholar] [CrossRef]
- Ayuso, J.M.; Vitek, R.; Swick, A.D.; Skala, M.C.; Wisinski, K.B.; Kimple, R.J.; Lambert, P.F.; Beebe, D.J. Effects of culture method on response to EGFR therapy in head and neck squamous cell carcinoma cells. Sci. Rep. 2019, 9, 12480. [Google Scholar] [CrossRef]
- Tuomainen, K.; Al-Samadi, A.; Potdar, S.; Turunen, L.; Turunen, M.; Karhemo, P.-R.; Bergman, P.; Risteli, M.; Åström, P.; Tiikkaja, R.; et al. Human Tumor–Derived Matrix Improves the Predictability of Head and Neck Cancer Drug Testing. Cancers 2020, 12, 92. [Google Scholar] [CrossRef]
- Jin, D.; Ma, X.; Luo, Y.; Fang, S.; Xie, Z.; Li, X.; Qi, D.; Zhang, F.; Kong, J.; Li, J. Application of a microfluidic-based perivascular tumor model for testing drug sensitivity in head and neck cancers and toxicity in endothelium. RSC Adv. 2016, 6, 29598–29607. [Google Scholar] [CrossRef]
- Wahbi, W.; Korelin, K.; Sieviläinen, M.; Karihtala, P.; Wilkman, T.; Tarkkanen, J.; Salo, T.; Al-Samadi, A. Evaluation of in vitro and in vivo personalized cancer treatment assays for oral squamous cell carcinoma. Transl. Oncol. 2023, 33, 101677. [Google Scholar] [CrossRef]
- Sieviläinen, M.; Saavalainen, J.; Adnan-Awad, S.; Salo, T.; Al-Samadi, A. IDO1 Inhibition Reduces Immune Cell Exclusion Through Inducing Cell Migration While PD-1 Blockage Increases IL-6 and -8 Secretion From T Cells in Head and Neck Cancer. Front. Immunol. 2022, 13, 812822. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Yu, D. Tumor microenvironment as a therapeutic target in cancer. Pharmacol. Ther. 2021, 221, 107753. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 2012, 196, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun. Signal. 2020, 18, 59. [Google Scholar] [CrossRef] [PubMed]
- Micalet, A.; Moeendarbary, E.; Cheema, U. 3D In Vitro Models for Investigating the Role of Stiffness in Cancer Invasion. ACS Biomater. Sci. Eng. 2021, 9, 3729–3741. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Coussens, L.M. Accessories to the crime: Functions of cells recruited to the tumor microenvironment. Cancer Cell 2012, 21, 309–322. [Google Scholar] [CrossRef]
- Heimdal, J.; Aarstad, H.J.; Olofsson, J. Monocytes secrete interleukin-6 when co-cultured in vitro with benign or malignant autologous fragment spheroids from squamous cell carcinoma patients. Scand. J. Immunol. 2000, 51, 271–278. [Google Scholar] [CrossRef]
- Heimdal, J.H.; Olsnes, C.; Olofsson, J.; Aarstad, H.J. Monocyte and monocyte-derived macrophage secretion of MCP-1 in co-culture with autologous malignant and benign control fragment spheroids. Cancer Immunol. Immunother. 2001, 50, 300–306. [Google Scholar] [CrossRef]
- Kross, K.W.; Heimdal, J.H.; Aarstad, H.J. Mononuclear phagocytes in head and neck squamous cell carcinoma. Eur. Arch. Otorhinolaryngol. 2010, 267, 335–344. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kross, K.W.; Heimdal, J.H.; Olsnes, C.; Olofsson, J.; Aarstad, H.J. Co-culture of head and neck squamous cell carcinoma spheroids with autologous monocytes predicts prognosis. Scand. J. Immunol. 2008, 67, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.K.; Schirlau, K.; Sonkoly, E.; Brandau, S.; Lang, S.; Pivarcsi, A.; Balz, V.; Müller, A.; Homey, B.; Boelke, E.; et al. A novel mechanism for anti-EGFR antibody action involves chemokine-mediated leukocyte infiltration. Int. J. Cancer 2009, 124, 2589–2596. [Google Scholar] [CrossRef]
- Hirschhaeuser, F.; Leidig, T.; Rodday, B.; Lindemann, C.; Mueller-Klieser, W. Test System for Trifunctional Antibodies in 3D MCTS Culture. J. Biomol. Screen. 2009, 14, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Q.; Kuo, J.-C.; Wei, M.-T.; Wu, M.-C.; Yang, M.-H.; Chiou, A. Fibroblast promotes head and neck squamous cell carcinoma cell invasion through mechanical barriers in 3D collagen microenvironments. ACS Appl. Bio Mater. 2020, 3, 6419–6429. [Google Scholar] [CrossRef]
- Gomez, K.E.; Wu, F.; Keysar, S.B.; Morton, J.J.; Miller, B.; Chimed, T.S.; Le, P.N.; Nieto, C.; Chowdhury, F.N.; Tyagi, A.; et al. Cancer Cell CD44 Mediates Macrophage/Monocyte-Driven Regulation of Head and Neck Cancer Stem Cells. Cancer Res. 2020, 80, 4185–4198. [Google Scholar] [CrossRef]
- Huang, C.; Yoon, C.; Zhou, X.H.; Zhou, Y.C.; Zhou, W.W.; Liu, H.; Yang, X.; Lu, J.; Lee, S.Y.; Huang, K. ERK1/2-Nanog signaling pathway enhances CD44+ cancer stem-like cell phenotypes and epithelial-to-mesenchymal transition in head and neck squamous cell carcinomas. Cell Death Dis. 2020, 11, 266. [Google Scholar] [CrossRef]
- Engelmann, L.; Thierauf, J.; Laureano, N.K.; Stark, H.-J.; Prigge, E.-S.; Horn, D.; Freier, K.; Grabe, N.; Rong, C.; Federspil, P. Organotypic co-cultures as a novel 3D model for head and neck squamous cell carcinoma. Cancers 2020, 12, 2330. [Google Scholar] [CrossRef]
- Kochanek, S.J.; Close, D.A.; Johnston, P.A. High Content Screening Characterization of Head and Neck Squamous Cell Carcinoma Multicellular Tumor Spheroid Cultures Generated in 384-Well Ultra-Low Attachment Plates to Screen for Better Cancer Drug Leads. Assay Drug Dev. Technol. 2019, 17, 17–36. [Google Scholar] [CrossRef]
- Braunholz, D.; Saki, M.; Niehr, F.; Öztürk, M.; Puértolas, B.B.; Konschak, R.; Budach, V.; Tinhofer, I. Spheroid Culture of Head and Neck Cancer Cells Reveals an Important Role of EGFR Signalling in Anchorage Independent Survival. PLoS ONE 2016, 11, e0163149. [Google Scholar] [CrossRef]
- He, C.K.; Chen, Y.W.; Wang, S.H.; Hsu, C.H. Hydrodynamic shuttling for deterministic high-efficiency multiple single-cell capture in a microfluidic chip. Lab Chip 2019, 19, 1370–1377. [Google Scholar] [CrossRef]
- Byun, J.-Y.; Huang, K.; Lee, J.S.; Huang, W.; Hu, L.; Zhen, X.; Tang, X.; Li, F.; Jo, D.-G.; Song, X.; et al. Targeting HIF-1α/NOTCH1 pathway eliminates CD44+ cancer stem-like cell phenotypes, malignancy, and resistance to therapy in head and neck squamous cell carcinoma. Oncogene 2022, 41, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, R.; Chen, Y.; Yang, X.; Shang, Z. Stromal nicotinamide N-methyltransferase orchestrates the crosstalk between fibroblasts and tumour cells in oral squamous cell carcinoma: Evidence from patient-derived assembled organoids. Oncogene 2023, 42, 1166–1180. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-R.; Brown, J.; Yu, Y.; Lee, D.; Zhou, K.; Dunn, Z.S.; Hon, R.; Wilson, M.; Kramer, A.; Zhu, Y.; et al. Targeting Immunosuppressive Tumor-Associated Macrophages Using Innate T Cells for Enhanced Antitumor Reactivity. Cancers 2022, 14, 2749. [Google Scholar] [CrossRef] [PubMed]
- Roma-Rodrigues, C.; Mendes, R.; Baptista, P.V.; Fernandes, A.R. Targeting Tumor Microenvironment for Cancer Therapy. Int. J. Mol. Sci. 2019, 20, 840. [Google Scholar] [CrossRef] [PubMed]
- Doedens, A.L.; Stockmann, C.; Rubinstein, M.P.; Liao, D.; Zhang, N.; DeNardo, D.G.; Coussens, L.M.; Karin, M.; Goldrath, A.W.; Johnson, R.S. Macrophage expression of hypoxia-inducible factor-1α suppresses T-cell function and promotes tumor progression. Cancer Res. 2010, 70, 7465–7475. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-J.; Song, B.; Kim, Y.-S.; Kim, E.-K.; Lee, J.-M.; Lee, G.-E.; Kim, J.-O.; Kim, Y.-J.; Chang, W.-S.; Kang, C.-Y. Tumor Microenvironmental Conversion of Natural Killer Cells into Myeloid-Derived Suppressor CellsNK Cell Conversion into MDSCs. Cancer Res. 2013, 73, 5669–5681. [Google Scholar] [CrossRef]
- Noy, R.; Pollard, J.W. Tumor-associated macrophages: From mechanisms to therapy. Immunity 2014, 41, 49–61. [Google Scholar] [CrossRef]
- Gutschalk, C.M.; Herold-Mende, C.C.; Fusenig, N.E.; Mueller, M.M. Granulocyte Colony-Stimulating Factor and Granulocyte-Macrophage Colony-Stimulating Factor Promote Malignant Growth of Cells from Head and Neck Squamous Cell Carcinomas In vivo. Cancer Res. 2006, 66, 8026–8036. [Google Scholar] [CrossRef]
- Linde, N.; Gutschalk, C.M.; Hoffmann, C.; Yilmaz, D.; Mueller, M.M. Integrating macrophages into organotypic co-cultures: A 3D in vitro model to study tumor-associated macrophages. PLoS ONE 2012, 7, e40058. [Google Scholar] [CrossRef]
- Lu, Y.; Li, S.; Ma, L.; Li, Y.; Zhang, X.; Peng, Q.; Mo, C.; Huang, L.; Qin, X.; Liu, Y. Type conversion of secretomes in a 3D TAM2 and HCC cell co-culture system and functional importance of CXCL2 in HCC. Sci. Rep. 2016, 6, 24558. [Google Scholar] [CrossRef]
- Chávez-Galán, L.; Olleros, M.L.; Vesin, D.; Garcia, I. Much More than M1 and M2 Macrophages, There are also CD169+ and TCR+ Macrophages. Front. Immunol. 2015, 6, 263. [Google Scholar] [CrossRef]
- Laoui, D.; Van Overmeire, E.; Movahedi, K.; Van den Bossche, J.; Schouppe, E.; Mommer, C.; Nikolaou, A.; Morias, Y.; De Baetselier, P.; Van Ginderachter, J.A. Mononuclear phagocyte heterogeneity in cancer: Different subsets and activation states reaching out at the tumor site. Immunobiology 2011, 216, 1192–1202. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.; Kulawiec, D.; Jazwinska, D.; Zervantonakis, I. Paracrine CSF1 signaling regulates macrophage migration dynamics towards ovarian cancer cells in a 3D microfluidic model that recapitulates in vivo infiltration patterns in patient-derived xenograft models. bioRxiv 2022. [Google Scholar] [CrossRef]
- Lewis, C.E.; Pollard, J.W. Distinct role of macrophages in different tumor microenvironments. Cancer Res. 2006, 66, 605–612. [Google Scholar] [CrossRef]
- Wyckoff, J.B.; Wang, Y.; Lin, E.Y.; Li, J.-F.; Goswami, S.; Stanley, E.R.; Segall, J.E.; Pollard, J.W.; Condeelis, J. Direct visualization of macrophage-assisted tumor cell intravasation in mammary tumors. Cancer Res. 2007, 67, 2649–2656. [Google Scholar] [CrossRef]
- Heldin, C.H.; Rubin, K.; Pietras, K.; Ostman, A. High interstitial fluid pressure—An obstacle in cancer therapy. Nat. Rev. Cancer 2004, 4, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Hompland, T.; Ellingsen, C.; Øvrebø, K.M.; Rofstad, E.K. Interstitial fluid pressure and associated lymph node metastasis revealed in tumors by dynamic contrast-enhanced MRI. Cancer Res. 2012, 72, 4899–4908. [Google Scholar] [CrossRef] [PubMed]
- Penny, H.L.; Sieow, J.L.; Adriani, G.; Yeap, W.H.; Ee, P.S.C.; Luis, B.S.; Lee, B.; Lee, T.; Mak, S.Y.; Ho, Y.S. Warburg metabolism in tumor-conditioned macrophages promotes metastasis in human pancreatic ductal adenocarcinoma. Oncoimmunology 2016, 5, e1191731. [Google Scholar] [CrossRef] [PubMed]
- Battat, S.; Weitz, D.A.; Whitesides, G.M. An outlook on microfluidics: The promise and the challenge. Lab Chip 2022, 22, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.L.; Seager, R.J.; Litvak, F.; Spill, F.; Sieow, J.L.; Leong, P.H.; Kumar, D.; Tan, A.S.M.; Wong, S.C.; Adriani, G.; et al. Integrated in silico and 3D in vitro model of macrophage migration in response to physical and chemical factors in the tumor microenvironment. Integr. Biol. 2020, 12, 90–108. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).