Simple Summary
Prostate cancer is a highly heterogenous disease with respect to molecular, morphological and clinical features. Therefore, one of the major tasks in its management is to define the risk subgroups that would guide the treatment approach. Amino acid metabolism-related genes are involved in several aspects of prostate cancer progression. In this publication, we show that their expression is highly aberrant in prostate cancer, which could be potentially exploited for the establishment of disease progression parameters and therapeutic targets. We show that among the variables studied, the Gleason score was the strongest prognostic factor of progression-free survival in multivariate analysis. Additionally, the expression of SERINC3 and CSAD genes strongly differentiated between better and worse prognosis (low and high risk) for high and low Gleason scores, respectively. These results offer a suggestion for potential biomarkers of prostate cancer progression in patients that are stratified by the Gleason score.
Abstract
Prostate cancer is among the leading cancers according to both incidence and mortality. Due to the high molecular, morphological and clinical heterogeneity, the course of prostate cancer ranges from slow growth that usually does not require immediate therapeutic intervention to aggressive and fatal disease that spreads quickly. However, currently available biomarkers cannot precisely predict the course of a disease, and novel strategies are needed to guide prostate cancer management. Amino acids serve numerous roles in cancers, among which are energy production, building block reservoirs, maintenance of redox homeostasis, epigenetic regulation, immune system modulation and resistance to therapy. In this article, by using The Cancer Genome Atlas (TCGA) data, we found that the expression of amino acid metabolism-related genes is highly aberrant in prostate cancer, which holds potential to be exploited in biomarker design or in treatment strategies. This change in expression is especially evident for catabolism genes and transporters from the solute carrier family. Furthermore, by using recursive partitioning, we confirmed that the Gleason score is strongly prognostic for progression-free survival. However, the expression of the genes SERINC3 (phosphatidylserine and sphingolipids generation) and CSAD (hypotaurine generation) can refine prognosis for high and low Gleason scores, respectively. Therefore, our results hold potential for novel prostate cancer progression biomarkers.
1. Introduction
Prostate cancer is among the leading cancers according to both incidence and mortality. It is estimated that in 2020, there were 1,414,259 (7.3% of all sites) new cases diagnosed and 375,304 (3.8% of all sites) deaths from this disease [1]. Common treatment options for confined prostate cancer include surgical removal of the prostate (radical prostatectomy) and radiotherapy. However, biochemical recurrence, defined by a significant rise in blood levels of prostate-specific antigen, occurs within approximately 10 years in 20–40% of patients after radical prostatectomy and 30–50% after radiotherapy [2]. The biochemical recurrence can lead to a progressive disease, which is accompanied by symptoms or evidence of disease progression on imaging [3]. One of the major problems in prostate cancer management is to predict the course of a disease, that is, to differentiate between the tumors that will grow slowly and require minimal or no treatment and those that are more aggressive and will progress fast. Therefore, novel treatment strategies and therapeutic targets are needed, as well as better biomarkers, which would guide prostate cancer management.
Metabolic rewiring is one of the hallmarks of cancer [4], through which the cancer cell satisfies its high demands for energy and biomass building blocks to sustain its rapid proliferation. In comparison to other solid cancer types, which largely rely on aerobic glycolysis (the Warburg effect), prostate cancer cells use oxidative phosphorylation more than non-transformed prostate cells [5,6,7]. However, in advanced stages of prostate cancer, an increased glycolytic phenotype has been observed. In addition to these specificities, a hallmark of the metastatic, castration-resistant prostate cancer (mCRPC) is lipid metabolism rewiring, which manifests as increased fatty acids and cholesterol synthesis, uptake and oxidation [8].
Along with carbohydrates and fatty acids, amino acids are among the main sources of nutrients for energy homeostasis (alternative fuels) and building blocks for macromolecular biosynthesis. Additionally, amino acids help to maintain the redox balance as they are the main elements for reduced glutathione (GSH) and nicotinamide adenine dinucleotide phosphate (NADPH) generation, which are among the key molecules involved in control of the cellular redox state. Amino acid derivatives contribute to epigenetic modifications and posttranscriptional regulation. Namely, one-carbon units from the methionine and folate cycle are methyl donors for DNA and histone methylation, while acetyl-CoA derived from a group of amino acids can be used for histone acetylation. Amino acids also largely influence immune system responses in tumorigenesis and metastasis formation by creating an immunosuppressive or immunoeffective microenvironment [9]. Moreover, amino acids enable cancer cells to circumvent anticancer therapies [10]. The metabolism and uptake of amino acids, therefore, are aberrantly upregulated in many cancer types, and some of those cancer types are characterized by addiction to particular amino acids [11]. For these reasons, amino acid depletion therapies are extensively being explored in the area of cancer research [12].
The amino acid profile in prostate cancer, unlike in other solid tumors, is characterized by their anaplerotic roles more than by energy-production roles. Anaplerotic reactions are chemical reactions that form intermediates of a metabolic pathway and fuel that certain pathway. Many of the amino acids are implicated in prostate cancer, and their involvement has been recently reviewed [13,14]. For example, amino acids commonly related to prostate cancer include glutamine, leucine, serine, glycine, sarcosine, proline and arginine. In the light of the results of this paper, we describe further the roles of serine and taurine in prostate cancer.
Serine/glycine biosynthesis and one-carbon metabolism are intertwined and essential in promoting cancer cell survival and rapid proliferation. The excessive activation of serine/glycine biosynthesis pathways drives tumorigenesis and provides a single carbon unit for one-carbon metabolism. One-carbon metabolism, which is based on the chemical reactions of methionine and folate compounds, is used for the de novo synthesis of nucleotides, polyamines, amino acids, creatine and phospholipids. Serine is also a precursor for the synthesis of glycine and cysteine, both of which contribute to the production of glutathione, which is essential for redox homeostasis [15,16]. In prostate cancer, it was recently shown that increased serine and one-carbon pathway metabolism promote a neuroendocrine phenotype, which is the most lethal subtype of castration-resistant prostate cancer [17]. This characteristic represents a targetable vulnerability for prostate cancer [18,19]. In line with these findings, the role of alanine-serine-cysteine transporter 2 (ASCT2, SLC1A5) was studied. ASCT2 is a Na+-dependent transporter involved in the cellular uptake of neutral amino acids, that is, amino acids with small, hydrophilic side chains, such as serine, cysteine, asparagine and glutamine, but also alanine with the nonpolar methyl side chain [20]. The inhibition of ASCT2 suppresses prostate cancer cell growth in vitro. However, the contribution of serine to this process was not delineated, and the preferred substrate for ASCT2 is the conditionally essential amino acid glutamine [21,22]. Along with ASCT2 and several other transporters, SERINC3 (Serine Incorporator 3) protein was predicted to enable L-serine transmembrane transporter activity.
Taurine was also suggested to be potentially involved in prostate cancer progression. Namely, taurine was shown to attenuate the expression of epithelial–mesenchymal transition-related genes in human prostate cancer cells [23]. It also promoted apoptosis and inhibited proliferation of the prostate cancer cell line DU145, probably through the MST1/Hippo signaling pathway [24]. In another paper, it was shown that taurine suppressed PSA and metastasis-related genes expression in the human prostate cancer cell lines LNCaP and PC-3. In addition, taurine inhibited the migration of LNCaP and PC-3 cells [25]. Hypotaurine is a sulfinic acid that is an intermediate in the biosynthesis of taurine. An important gene in the metabolism of (hypo)taurine is CSAD (cysteine sulfinic acid decarboxylase). Its protein product catalyzes the decarboxylation of L-aspartate, 3-sulfino-L-alanine and L-cysteate to beta-alanine, hypotaurine and taurine, respectively. The preferred CSAD substrate is 3-sulfino-L-alanine.
In this introductory part we aimed to briefly present the global metabolic changes in prostate cancer and to place the changes in specific amino acid metabolism-related genes into this big picture. Furthermore, in a search for biomarkers that could predict the course of prostate cancer, in this article, we analyzed The Cancer Genome Atlas (TCGA) prostate adenocarcinoma (PRAD) dataset for the expression of amino acid metabolism-related genes. We found that their expression is highly aberrant in prostate cancer. By using a machine learning approach, we found that the expression of the genes CSAD and SERINC3 discriminates between better and worse prognosis (low and high risk) for progression-free survival (PFS) of prostate cancer patients when they are stratified according to the Gleason score. In brief, this article aimed at analyzing the expression and the prognostic significance of amino acid metabolism-related genes in prostate cancer. We believe that this publication (a) adds to the big picture of potential metabolic changes in prostate cancer and (b) suggests potential biomarkers for prostate cancer prognosis. Another value of this paper, in our opinion, is methodological, and that is because (c) we used machine learning techniques (recursive partitioning and survival tree) for the definition of prognostic subgroups, unlike many of the scientific papers with a similar topic that used Cox proportional hazards regression analysis for the definition of each gene’s prognostic abilities. Considering the prostate cancer heterogeneity, we believe that our method better captures its complexity.
2. Materials and Methods
2.1. Data Preparation and Differential Gene Expression Analysis
Amino acid metabolism-related genes were retrieved from The Molecular Signatures Database (MSigDB) [26] by using Gene Ontology Biological Process (GOBP) categories. The genes that were used in our analyses are listed in the Supplementary Table S1. Briefly, the following categories were considered: amino acid activation, homeostasis, transport, salvage, biosynthesis, metabolism, catabolism, response to amino acid starvation and C- and N-terminal protein amino acid modification. The final list contained 518 genes.
The Cancer Genome Atlas [27] prostate adenocarcinoma (PRAD) dataset, containing gene expression data and clinical information for 497 prostate cancer patients and corresponding control (surrounding, non-transformed) tissues for a subset of 52 patients, was downloaded and analyzed using the TCGAbiolinks R package [28,29,30]. To obtain more thorough insight into differentially expressed amino acid metabolism-related genes and to search deeper for their transcriptional changes in prostate cancer in comparison to non-transformed prostate tissue, we chose the threshold of │log2FC│ ≥ 0.585 (│fold change│ ≥ 1.5) and p adjusted < 0.01. The data based on differentially expressed amino acid metabolism-related genes obtained in this way (N = 121) are listed in Supplementary Table S2. The expression represents the value of normalized counts.
The clinical data shown in Table 1 were obtained from cBioPortal [31] and the NCI Genomic Data Commons (GDC, TCGA) portal [32]. In total, there were 493 patients with clinical information (age, Gleason score, TNM stage, information related to residual tumor and radiation therapy) available. The event that we analyzed was progression-free survival (93 patients with this event), since, fortunately, only a smaller subset of patients experienced an event needed for the overall survival calculation. Some variables contained missing data. However, decision trees that we obtained in survival analysis by using recursive partitioning method are not as adversely affected by missing data as traditional statistical methods [33].

Table 1.
Clinical information of TCGA patients. The number (N) and the percentage (in parenthesis) of patients belonging to a certain category is shown. In some categories, there are unknowns (NAs).
2.2. Functional Enrichment Analysis
The 121 differentially expressed amino acid metabolism-related genes (DEGs) from Supplementary Table S1 were subjected to a functional enrichment analysis, which was conducted by using the Enrichr web server [34,35]. The top 10 Gene Ontology Molecular Function (MF) and Biological Process (BP) terms are shown in Table 2. Table 3 lists the functional annotation of the solute carrier family genes with differential expression in prostate cancer retrieved from www.genecards.org [36]. Additionally, Table 4 lists the functional annotation of the catabolic genes from the category Cellular amino acid catabolic process (GO:0009063) with differential expression in prostate cancer. The functional information was also retrieved from www.genecards.org [36].
Table 2.
Enrichment analysis for the differentially expressed amino acid metabolism-related genes (N = 121). The gene ontology (GO) Molecular Function (MF) and Biological Process (BP) categories are listed.
Table 3.
Functional annotation of solute carrier (SLC) family genes for which expression changes were observed in prostate cancer. The up- and down-regulated genes are listed separately. FC indicates fold change (tumor, T vs. normal, N), and FDR is the false discovery rate.
Table 4.
Functional annotation of Cellular amino acid catabolic process (GO:0009063) genes from Table 2 for which expression changes were observed in prostate cancer. The up- and down-regulated genes are listed separately. FC indicates fold change (tumor, T vs. normal, N), and FDR is the false discovery rate.
2.3. Survival Analysis
Pre-processed and normalized, but un-filtered, TCGA [27] expression data for the amino acid metabolism-related genes were obtained through the TCGAbiolinks R package [28,29,30]. The clinical data were added to expression data, organized in a data matrix and analyzed using the data analysis software R [37], version 4.2.1.
For the survival analysis, we used rpart module [38,39] in the programming language R [37]. rpart stands for Recursive PARTitioning and is the most used application for the construction of survival trees. Survival trees obtained via this method enable visual identification and comparisons of prognostic factors in a simple and straightforward manner [40,41]. The method is insensitive to missing data, in contrast to classical statistical methods, and gives reliable and robust conclusions in most clinical scenarios. The method is described in more detail in our previous publications [42]. Briefly, at the beginning of the analysis, all patients are included and in subsequent steps, they are divided into prognostic subgroups in a survival tree. At the first split (root node), a logical check is performed. If the criterion of that node is met, the left side of the tree is approached; otherwise it is the right. This is repeated at each stage (decision node) until the terminal node is reached. Therefore, a survival tree obtained in this way is composed of decision nodes and terminal nodes (leaves). Each decision node uses a provided variable to subdivide patients into two subgroups with a maximum difference in hazard ratios (HRs). The terminal nodes are reached when no further improvement in subdivision is possible. Patients in the first decision node have hazard ratio of 1. The hazard ratio for patients in each node is expressed in comparison to this value. To avoid overfitting, that is, an extensive fragmentation of the tree for which it would be hard to infer a biological meaning, we set the complexity parameter CP to 0.0373.
2.4. Kaplan–Meier Survival Estimate
The difference in survival between patients in terminal nodes was analyzed using a log-rank test and is presented as survival curves based on the Kaplan–Meier survival estimate [43]. This part of the analysis was based on the EZR package [44] in programming language R. Data were considered statistically significant if the p value of the log-rank test was ≤0.05.
3. Results
3.1. Prostate Cancer Amino Acid Metabolism-Related Gene Expression Appears to Be Highly Aberrant
As elaborated previously, amino acid metabolism-related genes play important roles in prostate cancer. To search for amino acid metabolism-related genes that are specifically changed in prostate cancer, we conducted differential gene expression analysis. The results with thresholds │log2FC│ > 0.585 and p adjusted < 0.01 revealed 4215 differentially expressed genes (DEGs) in total. Among them, there were 121 differentially expressed amino acid metabolism-related genes, which are listed in Supplementary Table S2. The enrichment analysis conducted on those 121 differentially expressed genes (Table 2) showed that the expression of genes involved in amino acid transmembrane transport (mainly of the solute carrier family) is highly perturbed. The functional annotation of the solute carrier family genes listed in Supplementary Table S2 is provided in Table 3.
Table 4 lists the roles of Cellular amino acid catabolic process (GO:0009063) genes from Table 2 for which expression changes were observed in prostate cancer. Some important genes involved, for example, in the synthesis of glycine from serine, such as SHMT2 (serine hydroxymethyltransferase 2), showed increased expression in tumor tissue. The activity of SHMT2 has been suggested to be the primary source of intracellular glycine. Genes that encode proteins involved in the catabolism of L-lysine (AADAT), valine (ACAD8), glycine (GCSH), phenylalanine/tyrosine (GSTZ1), tryptophan (IDO1, TDO2), L-phenylalanine and L-arginine (IL4I1) and serine and glycine (SDS) also showed increased expression. The genes encoding proteins involved in production of the branched-chain amino acids leucine, isoleucine and valine (BCAT2, BCAT1) were also increased in tumor vs. non-transformed tissue. On the other hand, genes with decreased expression in tumor tissue were AMT, which is involved in glycine cleavage system; GLUL, which catalyzes the synthesis of glutamine from glutamate and ammonia; NOS1, nitric oxide synthase, which synthesizes nitric oxide from L-arginine; PIPOX, which is involved in L-lysine catabolic process; and PRODH, which catalyzes the first step in proline degradation.
For the genes that we show are involved in prostate cancer prognosis (see further section), CSAD had increased expression in prostate cancer (fold-change = 1.61, FDR < 0.001, Supplementary Table S2), while the expression of SERINC3 did not change according to the criteria used.
3.2. CSAD and SERINC3 Genes Further Refine the Prognostic Value of the Gleason Score in Prostate Cancer
Prognostic values of variables listed in Table 1 (age, Gleason score, TNM staging, residual tumor information and radiation therapy) supplemented with gene expression data for amino acid metabolism-related genes were determined using recursive partitioning, the recommended method by the AJCC (American Joint Committee on Cancer) for the analysis of prognostic studies [40,41]. The importance of individual variables is shown in Figure 1. The four most informative variables were the Gleason score and the expression of CSAD, GABBR1 and SERINC3 genes. Among them, only GABBR1 did not appear in multivariate analysis. The GABBR1 gene encodes a receptor for gamma-aminobutyric acid (GABA), which is the main inhibitory neurotransmitter in the mammalian central nervous system. Its role in the progression of prostate cancer has been documented [45].
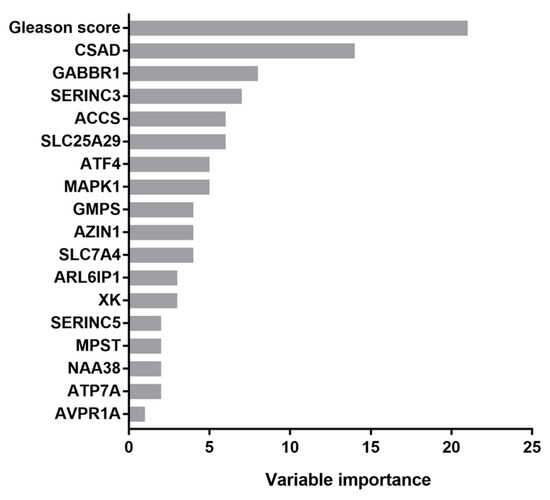
Figure 1.
Variable importance determined via the rpart method. CSAD, cysteine sulfinic acid decarboxylase; GABBR1, gamma-aminobutyric acid type B receptor subunit 1; SERINC3, serine incorporator 3; ACCS, 1-aminocyclopropane-1-carboxylate synthase homolog (inactive); SLC25A29, solute carrier family 25 member 29; ATF4, activating transcription factor 4; MAPK1, mitogen-activated protein kinase 1; GMPS, guanine monophosphate synthase; AZIN1, antizyme inhibitor 1; SLC7A4, solute carrier family 7 member 4; ARL6IP1, ADP ribosylation factor-like GTPase 6 interacting protein 1; XK, X-linked Kx blood group antigen, Kell and VPS13A-binding protein; SERINC5, serine incorporator 5; MPST, mercaptopyruvate sulfurtransferase; NAA38, N-alpha-acetyltransferase 38, NatC auxiliary subunit; ATP7A, ATPase copper transporting alpha; AVPR1A, arginine vasopressin receptor 1A.
However, AJCC criteria for prognostic studies require that a prognostic value of a variable must be always assessed in the context of other variables [40,41]. The rpart algorithm obeys this criterion since rpart uses all variables (multivariate approach) in the analysis. The rpart results are presented on a survival tree (Figure 2). Figure 2 shows that by using three variables, patients could be subdivided into three decision nodes and four terminal nodes (leaves). Variables used in the decision nodes were as follows: (1) the Gleason score, (2) CSAD gene expression (for Gleason score < 9), and (3) SERINC3 gene expression (for Gleason score ≥ 9). The importance of variables was determined by their position in the survival tree: the topmost variable (Gleason score) is the most informative, the variable below topmost is the second one by information value, and so on. The first number in a decision node rectangle denotes the hazard ratio (HR) and the numbers in the second row denote patients with the event (progression) vs. the total number of patients. The number in a third row denotes the percentage of patients in that node. Therefore, it is evident that, while the analysis starts with all patients included in the study (decision node 1; N = 493; N with progression = 93), decision node 2 is based on 71% and decision node 3 on 29% of patients. Further refinement of survival data revealed four prognostic groups: low Gleason score, low CSAD expression (28% of patients); low Gleason score, high CSAD expression (43%); high Gleason score, low SERINC3 expression (6%); and finally, high Gleason score, high SERINC3 expression (23%). The leftmost terminal node represents the group of patients at a very low risk (HR = 0.088), and the second represents patients at a medium risk (HR = 0.97). The second terminal node from the right represent patients at a low risk (HR = 0.48) and the right-most terminal node describes patients at a high risk of prostate cancer progression (HR = 2.9) (Table 5). To emphasize once again, patients in the first decision node have a hazard ratio of 1. The hazard ratio for patients in each node is expressed in comparison to this value. In conclusion, by using the information based on the Gleason score and the expression of CSAD and SERINC3 genes, a subdivision of prostate cancer patients into four prognostic groups with substantially different HRs was achieved.
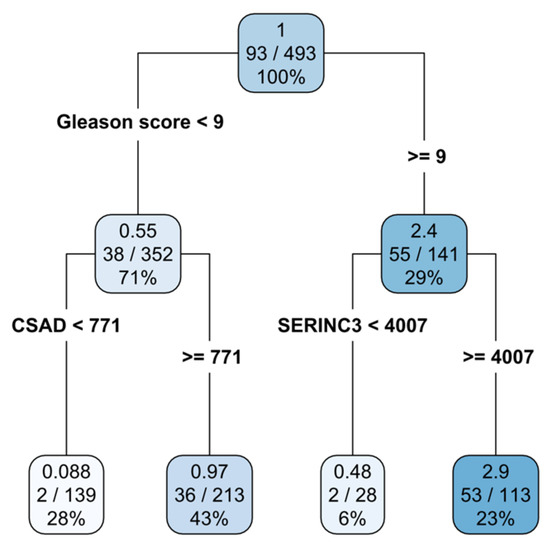
Figure 2.
Survival tree constructed using the rpart method identifies four terminal subgroups of patients. The shading of the color denotes the risk group (darker color stands for a higher risk).
Table 5.
Risk subgroups extracted via rpart analysis.
3.3. Kaplan–Meier Estimate on Prostate Cancer Patients Stratified According to Gleason Score and CSAD and SERINC3 Expression
The results of recursive partitioning (Figure 2) were further supplemented by survival curves (Kaplan–Meier method) for subgroups defined in each decision node. The difference for subgroups defined by the left and right branches of decision node 1 is shown in Figure 3, and it was statistically significant (log-rank test, p < 0.001). The subgroups defined by the left and right branches of node 2 are shown in Figure 4 (log-rank test, p < 0.001). Figure 5 shows that the difference between subgroups of node 3 was also statistically significant (log-rank test, p < 0.001).
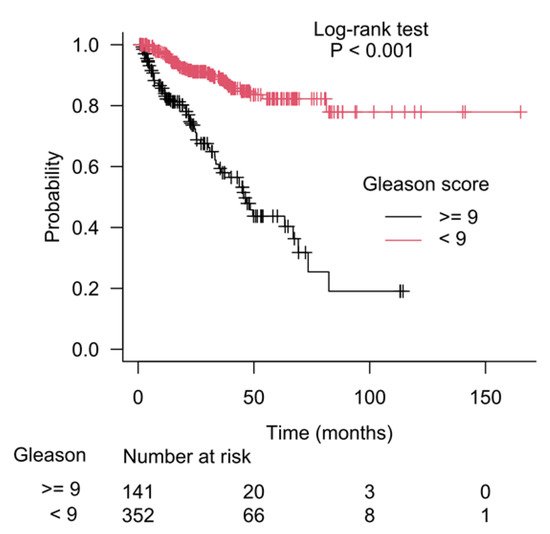
Figure 3.
Difference in patients’ survival for the left and the right branches of the starting decision node (node 1), which used the Gleason score as a separation criterion.
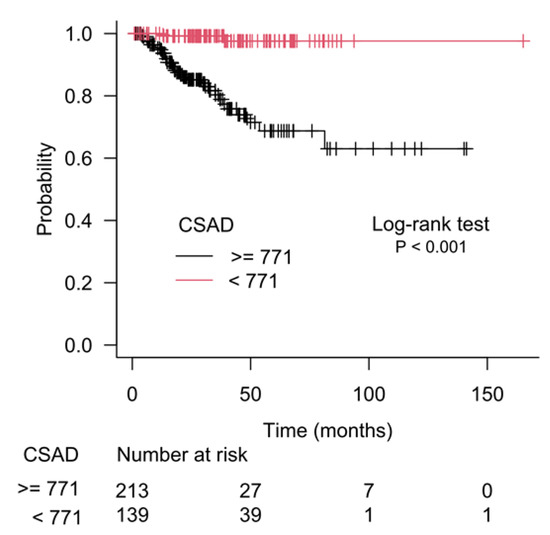
Figure 4.
Difference in patients’ survival for the left and the right branches of the second decision node (node 2), which used CSAD gene expression as a separation criterion.
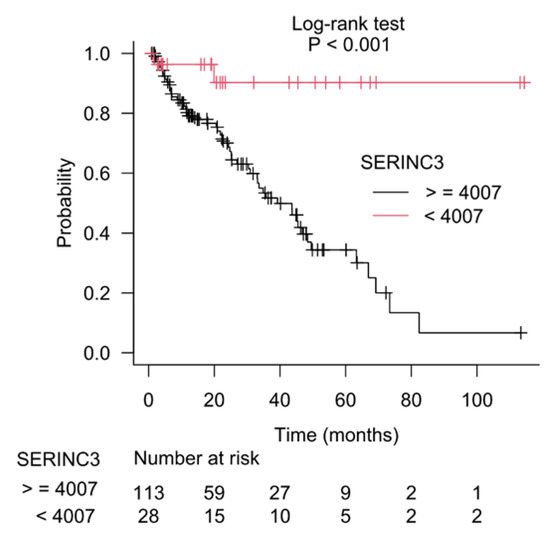
Figure 5.
Difference in patients’ survival for the left and the right branches of the third decision node (node 3), which used SERINC3 gene expression as a separation criterion.
4. Discussion
4.1. Metabolites and Metabolism-Related Genes in the Prognosis of Prostate Cancer
The driving events in prostate cancer progression include entangled actions of several signaling pathways that are potentiated by changes in gene expression, genetic and epigenetic alterations [46] and post-transcriptional and post-translational modifications [47]. However, although a substantial amount of information is gathered in regard to the mentioned processes, one of the major obstacles in prostate cancer management is still the inability to predict the course of a disease, that is, to differentiate between slowly growing cancers that do not require immediate treatment and those that are more aggressive and will progress fast.
The metabolic landscape in cancers is highly perturbed in comparison to that in healthy tissue and metabolic genes and molecules, therefore, hold potential to be exploited in a search for disease biomarkers and novel therapeutic targets. This is especially the case since, not only primary tumors, but also metastases from certain tissues (e.g., liver and some other sites [48,49]), acquire changes in metabolism-related gene expression profiles. Metabolic profiles in prostate cancer have been thoroughly studied and reviewed by Kelly et al. [50] who analyzed the articles reporting metabolites in prostate tissue, blood, urine and prostatic secretions. They showed that amino acids are among the most promising metabolic diagnostic biomarkers and biomarkers of tumor aggressiveness. Some amino acids (e.g., glutamine) were also used in terms of predicting disease recurrence [5]. In addition to metabolites themselves, the repertoire of metabolic genes as a source of prostate cancer biomarkers has already been studied. Namely, Zhang et al. identified three metabolism-associated prostate cancer clusters that were characterized by significantly different outcomes in disease-free survival (DFS), clinical stage, stemness index, tumor microenvironment (including stromal and immune cells), presence of DNA mutation (TP53 and SPOP), copy number variation and microsatellite instability [51]. In a further paper, they established metabolism-scores of tumors to predict the prognosis of prostate cancer. This metabolic score was closely related to the tumor microenvironment, presence of DNA mutations and drug sensitivity [52]. Feng et al. studied energy metabolism-related genes in prostate cancer and defined an energy metabolism-related gene prognostic index, which proved to predict biochemical recurrence for patients with prostate cancer that were undergoing radical prostatectomy [53]. Finally, Zhao et al. were able to predict biochemical-recurrence-free survival (BRFS) using a three-metabolic-gene risk score model in prostate cancer patients [2].
4.2. Differentially Expressed Amino Acid Metabolism-Related Genes in Prostate Cancer
Although, as elaborated, several papers already dealt with the potential of metabolic genes in predicting the outcome of prostate cancer patients, none of them, to the best of our knowledge, analyzed the amino-acid metabolism-related genes separately. Since amino acids themselves, as already mentioned [50], are involved in the prognosis for prostate cancer patients, it is to be expected that the genes encoding proteins that participate in their metabolism would also show prognostic capabilities. In very recent papers, the amino acid metabolism genes already showed good performance in the prognosis of e.g., colorectal cancer [54], hepatocellular carcinoma [55], clear cell renal cell carcinoma [56], glioma [57] and head and neck squamous cell carcinomas [58]. In this research, we studied the potential of amino acid metabolism-related genes to predict progression-free survival (PFS) using The Cancer Genome Atlas prostate adenocarcinoma (PRAD) dataset.
The first relevant finding of this paper is that the expression of the genes encoding proteins that are involved in amino acid transport across both the cellular (majority) and the mitochondrial (to a lesser extent) membrane show changed expression. Namely, the solute carrier (SLC) family genes were among the top terms in functional enrichment analysis of both Gene Ontology (GO) Molecular Function and GO Biological Process categories of differentially expressed genes (DEGs) (Table 2 and Table 3). The SLC group of membrane transport proteins include over 400 members organized into 66 families. Solutes that are transported by the various SLC proteins are extremely diverse and include charged and uncharged organic molecules, inorganic ions and the gas ammonia. However, most of the SLC group members listed in Table 3 are involved in amino acid transport as they were selected because of their connection with amino acid metabolism. Although more of the SLCs are up-regulated (15) than down-regulated (10) in prostate cancer, it is hard to speculate about the ‘big picture’, that is, to establish which of the amino acids are largely influenced by these changes in the expression of SLCs. What is known is that some of these gene products were shown to be implicated in prostate cancer progression, such as, for example, SLC7A5 [59], SLC7A11 [60], SLC11A1 [61], SLC43A1 [62] and SLC1A3 [63]. Although not listed in the Table 3, a recent paper documented metabolic reprogramming and the predominance of several solute carrier genes (SLC12A5, SLC25A17 and SLC27A6) during acquired enzalutamide resistance in prostate cancer [64], emphasizing the importance of the SLC family members in prostate cancer.
4.3. Prognostic Value of Amino Acid Metabolism-Related Genes in Prostate Cancer
To get back to the primary question of this publication, which would be the prediction of prostate cancer outcomes, several publications already used gene expression profiles to foresee the prostate cancer prognosis (e.g., [65,66,67,68,69,70,71,72]). However, as already mentioned, those still did not make it to the clinics; that is, the course of prostate cancer remains mainly unpredictable. Therefore, in this paper, we extended the knowledge on potential prostate cancer progression-free survival biomarkers to amino acid metabolism-related genes. The changes in expression of those genes are extensive in prostate cancer and therefore hold potential for biomarkers and therapeutic targets. We found that the Gleason score is the strongest variable influencing prostate cancer progression-free survival in a multivariate analysis. This is to be expected, since the Gleason score is highly informative of the characteristics of tumor cells that constitute the tumor tissue. However, when the patients were stratified according to a low/high Gleason score, the genes CSAD (for the low Gleason score) and SERINC3 (for the high Gleason score) differentiated the risk of progression. That is, patients with higher CSAD and higher SERINC3 expression are at a higher risk of progression (Figure 2).
CSAD protein is involved in the generation of beta-alanine, hypotaurine and taurine. Although papers suggest that taurine has a beneficial role in prostate cancer (see Introduction), it needs to be emphasized that hypotaurine is the preferential product of the biochemical reaction involving CSAD. It was shown that hypotaurine potentiates a malignant phenotype in glioma through aberrant hypoxic signaling. The authors show that taurine, the oxidation metabolite of hypotaurine, decreased intracellular hypotaurine and resulted in glioma cell growth arrest [73]. Therefore, the ratio of hypotaurine/taurine could play a role in prostate cancer as well. Additionally, long non-coding RNA TUG1 (taurine up-regulated 1) was originally identified in a genomic screen of taurine-treated mouse retinal cells [74]. TUG1 accelerates prostate cancer progression [75,76]. Its knockdown inhibits the tumorigenesis and progression of prostate cancer in vitro and in vivo [77] and enhances radiosensitivity [78]. Finally, high expression of TUG1 correlates with progression of the disease and less favorable survival profiles in prostate cancer patients [79]. To emphasize that CSAD plays versatile roles in different cancer types, data from The Human Protein Atlas [80,81] state that CSAD is an unfavorable prognostic marker in renal and colorectal cancer, which would agree with our study. However, it is favorable in urothelial, liver, pancreatic and head and neck cancer. To add more complexity to the potential mechanisms of action involving hypotaurine/taurine, CSAD also catalyzes the generation of beta-alanine. It would be interesting to further detangle these complex relationships (hypotaurine–taurine–beta–alanine), of which taurine is the most studied, and define their impact on prostate cancer.
As elaborated in an introductory part, serine metabolism potentiates the malignancy of prostate cancer. The serine incorporator (SERINC) proteins are a family of multipass transmembrane proteins associated with the biosynthesis of serine-containing phospholipids and sphingolipids [82]. More precisely, SERINC2–4 are carrier proteins that incorporate the polar amino acid serine into membranes to facilitate the synthesis of phosphatidylserine and sphingolipids [83]. SERINC proteins were most studied in the context of viral infections during which they are constitutive host resistance factors, which suppress viral infection by incorporating into virus particles [83]. Phosphatidylserine (PS) is a serine-containing phospholipid and a component of the cell membrane. It plays a key role in cell cycle signaling, specifically in relation to apoptosis. Studies using pre-clinical models of prostate cancer showed that antibody-mediated PS blockade reprograms the innate immune system to promote anti-tumor responses. Therefore, bavituximab, a PS-targeting antibody, is being assessed in multiple clinical trials, including those for prostate cancer [84]. Sphingolipids are synthesized from serine and palmitoyl-CoA. Inhibitors of sphingolipid metabolism were shown to antagonize pro-survival responses. Moreover, cancer cells use sphingolipid-driven escape mechanisms to evade therapies. Sphingolipids have also been implicated in prostate cancer, as recently reviewed [85]. This brief overview of the promoting roles of phosphatidylserine and sphingolipids in prostate tumorigenesis agrees with our findings that the increased expression of SERINC3, which potentiates their biosynthesis, represents a higher risk of disease progression for prostate cancer patients that are stratified according to the Gleason score. In addition to SERINC3, SERINC5, for which the gene product has a similar function to SERINC3, is present on the list of genes implicated by our univariate analysis (Figure 1). This further indicates that the processes conducted by proteins encoded by these genes are potentially critically involved in prostate tumorigenesis.
4.4. Methodological Considerations
Besides dealing with biological processes involved in prostate cancer progression, our paper differs from those with a similar topic in that we used machine learning to define prognostic subgroups instead of using Cox proportional hazards regression analysis to define gene-based prognosis. From the technical point of view, the recursive partitioning method used has the advantage in that it establishes the hierarchy of the variables studied; that is, this method lists the variables by their importance for prognosis. In this way, subgroups of patients are defined, and the knowledge on their specificities is refined. Given the heterogeneity of prostate cancer, we believe that this method is more suitable to define gene expression-specific prostate cancer characteristics. Additionally, the survival tree, generated through recursive partitioning, is easier to interpret than the Cox regression results.
5. Conclusions
In our study, we analyzed differentially expressed genes between prostate cancer and surrounding non-transformed prostate tissue by using TCGA data. We found that the expression of amino acid metabolism-related genes is highly aberrant in prostate cancer. The groups of genes that are the most affected include solute carrier family of amino acid transporters and the genes involved in the catabolism of amino acids, which are mainly up-regulated. Furthermore, we found that the Gleason score is the strongest prognostic factor for progression-free survival in prostate cancer patients, which is expected given the amount of information provided by this parameter. However, when the patients are stratified according to the Gleason score, the genes CSAD (low Gleason score) and SERINC3 (high Gleason score) further refine the prognosis. The high expression of both CSAD and SERINC3 is correlated with worse outcomes. The CSAD gene product is involved in hypotaurine generation, and the SERINC3 gene product is involved in the generation of phosphatidylserine and sphingolipids. There are indications that hypotaurine, phosphatidylserine and sphingolipids promote prostate cancer progression. We believe that our results hold potential for the future design of prognostic biomarkers in prostate cancer, which is an intensive field of research, considering that the progression of prostate cancer is currently hard to predict. Functional studies on CSAD and SERINC3 genes and their regulators are needed to further delineate their roles in prostate cancer, which would reveal their potential for further interventions.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cancers15041309/s1, Table S1: Human amino-acid metabolism-related genes categorized through Gene Ontology Biological Processes (GOBPs; retrieved from https://www.gsea-msigdb.org/gsea/msigdb/index.jsp (accessed on 1 November 2022)); Table S2: Differentially expressed amino-acid metabolism-related genes between prostate cancer tumor and non-tumor tissue with a threshold |log2FC| ≥ 0.585 and FDR < 0.01.
Author Contributions
Conceptualization, I.S. and P.K.; methodology, I.S. and P.K.; formal analysis, I.S. and P.K.; investigation, I.S., P.K. and K.G.T.; data curation, I.S. and P.K.; writing—original draft preparation, I.S.; writing—review and editing, I.S., P.K. and K.G.T.; supervision, P.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a MY ZABA START 2019 donation from Zagrebačka banka (I.S.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable (this study uses publicly available data).
Data Availability Statement
In this article, we used The Cancer Genome Atlas (TCGA) prostate adenocarcinoma (PRAD) dataset available at https://gdc.cancer.gov/ (accessed on 1 November 2022).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Zhao, Y.; Tao, Z.; Li, L.; Zheng, J.; Chen, X. Predicting Biochemical-Recurrence-Free Survival Using a Three-Metabolic-Gene Risk Score Model in Prostate Cancer Patients. BMC Cancer 2022, 22, 239. [Google Scholar] [CrossRef]
- van den Broeck, T.; van den Bergh, R.C.N.; Briers, E.; Cornford, P.; Cumberbatch, M.; Tilki, D.; de Santis, M.; Fanti, S.; Fossati, N.; Gillessen, S.; et al. Biochemical Recurrence in Prostate Cancer: The European Association of Urology Prostate Cancer Guidelines Panel Recommendations. Eur. Urol. Focus 2020, 6, 231–234. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Zadra, G.; Loda, M. Metabolic Vulnerabilities of Prostate Cancer: Diagnostic and Therapeutic Opportunities. Cold Spring Harb Perspect Med 2018, 8, a030569. [Google Scholar] [CrossRef]
- Eidelman, E.; Twum-Ampofo, J.; Ansari, J.; Siddiqui, M.M. The Metabolic Phenotype of Prostate Cancer. Front. Oncol. 2017, 7, 131. [Google Scholar] [CrossRef]
- Ahmad, F.; Cherukuri, M.K.; Choyke, P.L. Metabolic Reprogramming in Prostate Cancer. Br. J. Cancer 2021, 125, 1185–1196. [Google Scholar] [CrossRef]
- Chetta, P.; Zadra, G. Metabolic Reprogramming as an Emerging Mechanism of Resistance to Endocrine Therapies in Prostate Cancer. Cancer Drug Resist. 2021, 4, 143–162. [Google Scholar] [CrossRef]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino Acids in Cancer. Exp. Mol. Med. 2020, 52, 15–30. [Google Scholar] [CrossRef]
- Yoo, H.C.; Han, J.M. Amino Acid Metabolism in Cancer Drug Resistance. Cells 2022, 11, 140. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, X.; Cheng, C.; Yu, W.; Yi, P. Metabolism of Amino Acids in Cancer. Front. Cell Dev. Biol. 2021, 8, xxx. [Google Scholar] [CrossRef]
- Butler, M.; van der Meer, L.T.; van Leeuwen, F.N. Amino Acid Depletion Therapies: Starving Cancer Cells to Death. Trends Endocrinol. Metab. 2021, 32, 367–381. [Google Scholar] [CrossRef]
- Strmiska, V.; Michalek, P.; Eckschlager, T.; Stiborova, M.; Adam, V.; Krizkova, S.; Heger, Z. Prostate Cancer-Specific Hallmarks of Amino Acids Metabolism: Towards a Paradigm of Precision Medicine. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 248–258. [Google Scholar] [CrossRef]
- Schcolnik-Cabrera, A.; Juárez-López, D. Dual Contribution of the MTOR Pathway and of the Metabolism of Amino Acids in Prostate Cancer. Cell Oncol. 2022, 45, 831–859. [Google Scholar] [CrossRef]
- Yang, M.; Vousden, K.H. Serine and One-Carbon Metabolism in Cancer. Nat. Rev. Cancer 2016, 16, 650–662. [Google Scholar] [CrossRef]
- Pan, S.; Fan, M.; Liu, Z.; Li, X.; Wang, H. Serine, Glycine and One-Carbon Metabolism in Cancer (Review). Int. J. Oncol. 2021, 58, 158–170. [Google Scholar] [CrossRef]
- Reina-Campos, M.; Linares, J.F.; Duran, A.; Cordes, T.; L’Hermitte, A.; Badur, M.G.; Bhangoo, M.S.; Thorson, P.K.; Richards, A.; Rooslid, T.; et al. Increased Serine and One-Carbon Pathway Metabolism by PKCλ/ι Deficiency Promotes Neuroendocrine Prostate Cancer. Cancer Cell 2019, 35, 385–400.e9. [Google Scholar] [CrossRef]
- Gao, X.; Locasale, J.W.; Reid, M.A. Serine and Methionine Metabolism: Vulnerabilities in Lethal Prostate Cancer. Cancer Cell 2019, 35, 339–341. [Google Scholar] [CrossRef]
- Ganini, C.; Amelio, I.; Bertolo, R.; Candi, E.; Cappello, A.; Cipriani, C.; Mauriello, A.; Marani, C.; Melino, G.; Montanaro, M.; et al. Serine and One-Carbon Metabolisms Bring New Therapeutic Venues in Prostate Cancer. Discov. Oncol. 2021, 12, 45. [Google Scholar] [CrossRef]
- Ndaru, E.; Garibsingh, R.A.A.; Shi, Y.Y.; Wallace, E.; Zakrepine, P.; Wang, J.; Schlessinger, A.; Grewer, C. Novel Alanine Serine Cysteine Transporter 2 (ASCT2) Inhibitors Based on Sulfonamide and Sulfonic Acid Ester Scaffolds. J. Gen. Physiol. 2019, 151, 357–368. [Google Scholar] [CrossRef]
- Saruta, M.; Takahara, K.; Yoshizawa, A.; Niimi, A.; Takeuchi, T.; Nukaya, T.; Takenaka, M.; Zennami, K.; Ichino, M.; Sasaki, H.; et al. Alanine-Serine-Cysteine Transporter 2 Inhibition Suppresses Prostate Cancer Cell Growth In Vitro. J. Clin. Med. 2022, 11, 5466. [Google Scholar] [CrossRef]
- Scalise, M.; Pochini, L.; Console, L.; Losso, M.A.; Indiveri, C. The Human SLC1A5 (ASCT2) Amino Acid Transporter: From Function to Structure and Role in Cell Biology. Front. Cell Dev. Biol. 2018, 6, 96. [Google Scholar] [CrossRef]
- Tang, Y.; Kim, Y.S.; Choi, E.J.; Hwang, Y.J.; Yun, Y.S.; Bae, S.M.; Park, P.J.; Kim, E.K. Taurine Attenuates Epithelial-Mesenchymal Transition-Related Genes in Human Prostate Cancer Cells. Adv. Exp. Med. Biol. 2017, 975, 1203–1212. [Google Scholar] [CrossRef]
- Song, X.; Yuan, B.; Zhao, S.; Zhao, D. Effect of Taurine on the Proliferation, Apoptosis and MST1/Hippo Signaling in Prostate Cancer Cells. Transl. Cancer Res. 2022, 11, 1705–1712. [Google Scholar] [CrossRef]
- Tang, Y.; Choi, E.J.; Cheong, S.H.; Hwang, Y.J.; Arokiyaraj, S.; Park, P.J.; Moon, S.H.; Kim, E.K. Effect of Taurine on Prostate-Specific Antigen Level and Migration in Human Prostate Cancer Cells. Adv. Exp. Med. Biol. 2015, 803, 203–214. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database Hallmark Gene Set Collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Sander, C.; Stuart, J.M.; Chang, K.; Creighton, C.J.; et al. The Cancer Genome Atlas Pan-Cancer Analysis Project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Silva, T.C.; Colaprico, A.; Olsen, C.; D’Angelo, F.; Bontempi, G.; Ceccarelli, M.; Noushmehr, H. TCGA Workflow: Analyze Cancer Genomics and Epigenomics Data Using Bioconductor Packages. F1000 Res. 2016, 5, 1542. [Google Scholar] [CrossRef]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor Package for Integrative Analysis of TCGA Data. Nucleic Acids Res. 2016, 44, e17. [Google Scholar] [CrossRef]
- Mounir, M.; Lucchetta, M.; Silva, T.C.; Olsen, C.; Bontempi, G.; Chen, X.; Noushmehr, H.; Colaprico, A.; Papaleo, E. New Functionalities in the TCGAbiolinks Package for the Study and Integration of Cancer Data from GDC and GTEX. PLoS Comput. Biol. 2019, 15. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the CBioPortal. Sci. Signal 2013, 6, pl1. [Google Scholar] [CrossRef]
- Heath, A.P.; Ferretti, V.; Agrawal, S.; An, M.; Angelakos, J.C.; Arya, R.; Bajari, R.; Baqar, B.; Barnowski, J.H.B.; Burt, J.; et al. The NCI Genomic Data Commons. Nat. Genet. 2021, 53, 257–262. [Google Scholar] [CrossRef]
- Narayanachar Tattar, P.; Vaman, H.J. Survival Analysis; CRC Press: Boca Raton, FL, USA, 2023. [Google Scholar]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Iny Stein, T.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 2016. [Google Scholar] [CrossRef]
- R Core Team. R Core Team; R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 1 November 2022).
- Atkinson, E.J.; Therneau, T.M. An Introduction to Recursive Partitioning Using the RPART Routines. Mayo Clin. 2000, 61, xxx. [Google Scholar]
- Therneau, T.; Atkinson, B. Rpart: Recursive Partitioning and Regression Trees. R Package Version 4.1-15. 2019. Available online: https://CRAN.R-project.org/package=rpart (accessed on 1 November 2022).
- Ulm, K.; Kriner, M.; Eberle, S.; Reck, M.; Hessler, S. Statistical Methods to Identify Predictive Factors. In Handbook of Statistics in Clinical Oncology; Crowley, J., Ankerst, D., Eds.; Chapman & Hall/CRC: Boca Raton, FL, USA, 2006; pp. 335–345. [Google Scholar]
- Schumacher, M.; Hollander, N.; Schwarzer, G.; Sauerbrei, W. Prognostic Factor Studies. In Handbook of Statistics in Clinical Oncology; Crowley, J., Ankerst, D., Eds.; Chapman & Hall/CRC: Boca Raton, FL, USA, 2006; pp. 289–333. [Google Scholar]
- Aralica, G.; Šarec Ivelj, M.; Pačić, A.; Baković, J.; Milković Periša, M.; Krištić, A.; Konjevoda, P. Prognostic Significance of Lacunarity in Preoperative Biopsy of Colorectal Cancer. Pathol. Oncol. Res. 2020, 26, 2567–2576. [Google Scholar] [CrossRef]
- Rowe, P. Essential Statistics for the Pharmaceutical Sciences; Wiley Online Library: Hoboken, NJ, USA, 2015. [Google Scholar]
- Kanda, Y. Investigation of the Freely Available Easy-to-Use Software “EZR” for Medical Statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef]
- Solorzano, S.R.; Imaz-Rosshandler, I.; Camacho-Arroyo, I.; García-Tobilla, P.; Morales-Montor, G.; Salazar, P.; Arena-Ortiz, M.L.; Rodríguez-Dorantes, M. GABA Promotes Gastrin-Releasing Peptide Secretion in NE/NE-like Cells: Contribution to Prostate Cancer Progression. Sci. Rep. 2018, 8, 10272. [Google Scholar] [CrossRef]
- Testa, U.; Castelli, G.; Pelosi, E. Cellular and Molecular Mechanisms Underlying Prostate Cancer Development: Therapeutic Implications. Medicines 2019, 6, 82. [Google Scholar] [CrossRef]
- Samaržija, I. Post-Translational Modifications That Drive Prostate Cancer Progression. Biomolecules 2021, 11, 247. [Google Scholar] [CrossRef]
- Samaržija, I. Site-Specific and Common Prostate Cancer Metastasis Genes as Suggested by Meta-Analysis of Gene Expression Data. Life 2021, 11, 636. [Google Scholar] [CrossRef]
- Samaržija, I. A Need for Stratification of Metastasis Samples According to Secondary Site in Gene Expression Studies. Biocell 2022, 46, 1747–1750. [Google Scholar] [CrossRef]
- Kelly, R.S.; Heiden, M.G.V.; Giovannucci, E.; Mucci, L.A. Metabolomic Biomarkers of Prostate Cancer: Prediction, Diagnosis, Progression, Prognosis, and Recurrence. Cancer Epidemiol. Biomark. Prev. 2016, 25, 887–906. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, R.; Liang, F.; Zhang, L.; Liang, X. Identification of Metabolism-Associated Prostate Cancer Subtypes and Construction of a Prognostic Risk Model. Front Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, X.; Zhang, L.; Wang, D. Metabolic Characterization and Metabolism-Score of Tumor to Predict the Prognosis in Prostate Cancer. Sci. Rep. 2021, 11, 22486. [Google Scholar] [CrossRef]
- Feng, D.; Shi, X.; Zhang, F.; Xiong, Q.; Wei, Q.; Yang, L. Energy Metabolism-Related Gene Prognostic Index Predicts Biochemical Recurrence for Patients With Prostate Cancer Undergoing Radical Prostatectomy. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef]
- Peng, X.; Zheng, T.; Guo, Y.; Zhu, Y. Amino Acid Metabolism Genes Associated with Immunotherapy Responses and Clinical Prognosis of Colorectal Cancer. Front. Mol. Biosci. 2022, 9. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, J.; Wang, S.; Jiang, Q.; Xu, K. Identification and Validation of a Nine-Gene Amino Acid Metabolism-Related Risk Signature in HCC. Front Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Zhang, F.; Lin, J.; Zhu, D.; Tang, Y.; Lu, Y.; Liu, Z.; Wang, X. Identification of an Amino Acid Metabolism-Associated Gene Signature Predicting the Prognosis and Immune Therapy Response of Clear Cell Renal Cell Carcinoma. Front. Oncol. 2022, 12. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Chai, R.C.; Wang, Y.Z.; Wang, Z.; Liu, X.; Wu, F.; Jiang, T. Amino Acid Metabolism-Related Gene Expression-Based Risk Signature Can Better Predict Overall Survival for Glioma. Cancer Sci. 2019, 110, 321–333. [Google Scholar] [CrossRef]
- Li, W.; Zou, Z.; An, N.; Wang, M.; Liu, X.; Mei, Z. A Multifaceted and Feasible Prognostic Model of Amino Acid Metabolism-Related Genes in the Immune Response and Tumor Microenvironment of Head and Neck Squamous Cell Carcinomas. Front. Oncol. 2022, 12. [Google Scholar] [CrossRef]
- Xu, M.; Sakamoto, S.; Matsushima, J.; Kimura, T.; Ueda, T.; Mizokami, A.; Kanai, Y.; Ichikawa, T. Up-Regulation of LAT1 during Antiandrogen Therapy Contributes to Progression in Prostate Cancer Cells. J. Urol. 2016, 195, 1588–1597. [Google Scholar] [CrossRef]
- Cordova, R.A.; Misra, J.; Amin, P.H.; Klunk, A.J.; Damayanti, N.P.; Carlson, K.R.; Elmendorf, A.J.; Kim, H.-G.; Mirek, E.T.; Elzey, B.D.; et al. GCN2 EIF2 Kinase Promotes Prostate Cancer by Maintaining Amino Acid Homeostasis. Elife 2022, 11, e81083. [Google Scholar] [CrossRef]
- Zhu, Q.; Meng, Y.; Li, S.; Xin, J.; Du, M.; Wang, M.; Cheng, G. Association of Genetic Variants in Autophagy-Lysosome Pathway Genes with Susceptibility and Survival to Prostate Cancer. Gene 2022, 808, 145953. [Google Scholar] [CrossRef]
- Rii, J.; Sakamoto, S.; Sugiura, M.; Kanesaka, M.; Fujimoto, A.; Yamada, Y.; Maimaiti, M.; Ando, K.; Wakai, K.; Xu, M.; et al. Functional Analysis of LAT3 in Prostate Cancer: Its Downstream Target and Relationship with Androgen Receptor. Cancer Sci. 2021, 112, 3871–3883. [Google Scholar] [CrossRef]
- Sun, J.; Nagel, R.; Zaal, E.A.; Ugalde, A.P.; Han, R.; Proost, N.; Song, J.; Pataskar, A.; Burylo, A.; Fu, H.; et al. SLC 1A3 Contributes to L-asparaginase Resistance in Solid Tumors. EMBO J. 2019, 38, e102147. [Google Scholar] [CrossRef]
- Verma, S.; Shankar, E.; Chan, E.R.; Gupta, S. Metabolic Reprogramming and Predominance of Solute Carrier Genes during Acquired Enzalutamide Resistance in Prostate Cancer. Cells 2020, 9, 2535. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Y.; Wang, X.; Ping, Y.; Wang, D.; Cao, Y.; Dai, Y.; Liu, W.; Tao, Z. Five-Gene Signature Associating with Gleason Score Serve as Novel Biomarkers for Identifying Early Recurring Events and Contributing to Early Diagnosis for Prostate Adenocarcinoma. J. Cancer 2021, 12, 3626–3647. [Google Scholar] [CrossRef]
- Meng, J.; Guan, Y.; Wang, B.; Chen, L.; Chen, J.; Zhang, M.; Liang, C. Risk Subtyping and Prognostic Assessment of Prostate Cancer Based on Consensus Genes. Commun. Biol. 2022, 5, 233. [Google Scholar] [CrossRef]
- Wu, X.; Lv, D.; Lei, M.; Cai, C.; Zhao, Z.; Eftekhar, M.; Gu, D.; Liu, Y. A 10-Gene Signature as a Predictor of Biochemical Recurrence after Radical Prostatectomy in Patients with Prostate Cancer and a Gleason Score ≥ 7. Oncol. Lett. 2020, 20, 2906–2918. [Google Scholar] [CrossRef]
- Glinsky, G.V.; Glinskii, A.B.; Stephenson, A.J.; Hoffman, R.M.; Gerald, W.L. Gene Expression Profiling Predicts Clinical Outcome of Prostate Cancer. J. Clin. Investig. 2004, 113, 913–923. [Google Scholar] [CrossRef]
- Mou, Z.; Spencer, J.; Knight, B.; John, J.; McCullagh, P.; McGrath, J.S.; Harries, L.W. Gene Expression Analysis Reveals a 5-Gene Signature for Progression-Free Survival in Prostate Cancer. Front. Oncol. 2022, 12. [Google Scholar] [CrossRef]
- Zhou, R.; Feng, Y.; Ye, J.; Han, Z.; Liang, Y.; Chen, Q.; Xu, X.; Huang, Y.; Jia, Z.; Zhong, W. Prediction of Biochemical Recurrence-Free Survival of Prostate Cancer Patients Leveraging Multiple Gene Expression Profiles in Tumor Microenvironment. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef]
- Lu, G.; Cai, W.; Wang, X.; Huang, B.; Zhao, Y.; Shao, Y.; Wang, D. Identifying Prognostic Signatures in the Microenvironment of Prostate Cancer. Transl. Androl. Urol. 2021, 10. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Z. A Gleason Score-Related Outcome Model for Human Prostate Cancer: A Comprehensive Study Based on Weighted Gene Co-Expression Network Analysis. Cancer Cell Int. 2020, 20, 159. [Google Scholar] [CrossRef]
- Gao, P.; Yang, C.; Nesvick, C.L.; Feldman, M.J.; Sizdahkhani, S.; Liu, H.; Chu, H.; Yang, F.; Tang, L.; Tian, J.; et al. Hypotaurine Evokes a Malignant Phenotype in Glioma through Aberrant Hypoxic Signaling. Oncotarget 2016, 7, 15200–15214. [Google Scholar] [CrossRef]
- Zhou, H.; Sun, L.; Wan, F. Molecular Mechanisms of TUG1 in the Proliferation, Apoptosis, Migration and Invasion of Cancer Cells (Review). Oncol. Lett. 2019, 18, 4393–4402. [Google Scholar] [CrossRef]
- Hao, S.D.; Ma, J.X.; Liu, Y.; Liu, P.J.; Qin, Y. Long Non-Coding TUG1 Accelerates Prostate Cancer Progression through Regulating MiR-128-3p/YES1 Axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 619–632. [Google Scholar] [CrossRef]
- Yang, B.; Tang, X.; Wang, Z.; Sun, D.; Wei, X.; Ding, Y. TUG1 Promotes Prostate Cancer Progression by Acting as a CeRNA of MiR-26a. Biosci. Rep. 2018, 38, BSR20180677. [Google Scholar] [CrossRef]
- Li, G.; Yang, J.; Chong, T.; Huang, Y.; Liu, Y.; Li, H. TUG1 Knockdown Inhibits the Tumorigenesis and Progression of Prostate Cancer by Regulating MicroRNA-496/Wnt/β-Catenin Pathway. Anticancer Drugs 2020, 31, 592–600. [Google Scholar] [CrossRef]
- Xiu, D.; Liu, L.; Cheng, M.; Sun, X.; Ma, X. Knockdown of LncRNA TUG1 Enhances Radiosensitivity of Prostate Cancer via the TUG1/MiR-139-5p/SMC1A Axis. Onco. Targets Ther. 2020, 13, 2319–2331. [Google Scholar] [CrossRef]
- Yu, J.; Wang, Y.; Peng, S. The High Expression of LncRNA TUG1 Correlates with Progressive Tumor Condition and Less Satisfying Survival Profiles in Prostate Cancer Patients. Transl. Cancer Res. 2019, 8. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science 2015, 347. [Google Scholar] [CrossRef]
- Uhlén, M.; Björling, E.; Agaton, C.; Szigyarto, C.A.K.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A Human Protein Atlas for Normal and Cancer Tissues Based on Antibody Proteomics. Mol. Cell. Proteom. 2005, 4, 1920–1932. [Google Scholar] [CrossRef]
- Tu, M.; Saputo, S. From Beginning to End: Expanding the SERINC3 Interactome Through an in Silico Analysis. Bioinform. Biol. Insights 2022, 16. [Google Scholar] [CrossRef]
- Xu, S.; Zheng, Z.; Pathak, J.L.; Cheng, H.; Zhou, Z.; Chen, Y.; Wu, Q.; Wang, L.; Zeng, M.; Wu, L. The Emerging Role of the Serine Incorporator Protein Family in Regulating Viral Infection. Front. Cell Dev. Biol. 2022, 10, 856468. [Google Scholar] [CrossRef]
- Birge, R.B.; Boeltz, S.; Kumar, S.; Carlson, J.; Wanderley, J.; Calianese, D.; Barcinski, M.; Brekken, R.A.; Huang, X.; Hutchins, J.T.; et al. Phosphatidylserine Is a Global Immunosuppressive Signal in Efferocytosis, Infectious Disease, and Cancer. Cell Death Differ. 2016, 23, 962–978. [Google Scholar] [CrossRef]
- Voelkel-Johnson, C.; Norris, J.S.; White-Gilbertson, S. Interdiction of Sphingolipid Metabolism Revisited: Focus on Prostate Cancer. Adv. Cancer Res. 2018, 140, 265–293. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).