
Define Critical Parameters of Trastuzumab-Mediated ADCC Assays via Assay Optimization Processes, Focusing on the Impact of Cryopreserved Effector Cells on Assay Performance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials & Methods
2.1. Cancer Cells
2.2. Therapeutic Monoclonal Antibody
2.3. PBMCs and NK Cells
2.4. LDH ADCC Assay
2.5. ADCC Reporter Assay
2.6. Western Blotting Analysis
2.7. Flow Cytometry
2.8. ELISA Assays
2.9. Knockdown of HER2 in SKBR-3 Cells
2.10. Statistical Analysis
3. Results
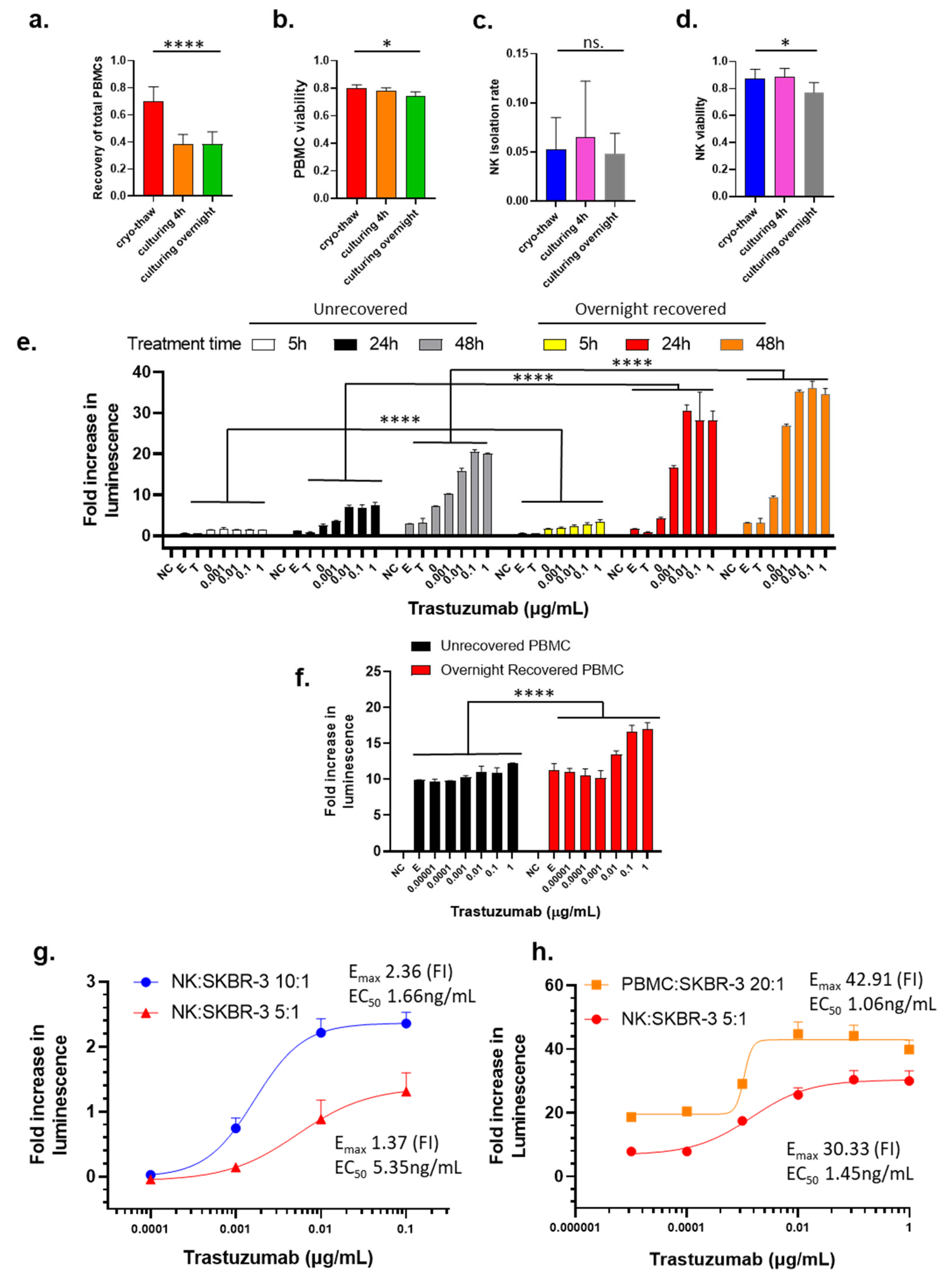
3.1. Influence of Cryopreservation and the Recovery Cultivation on PBMCs and NK Cells
3.2. Influence of Cryopreservation and Recovery Cultivation of PBMCs and NK Cells, Treatment Time, E/T Ratio, and Type of Effector Cells on ADCC Activity Measurement
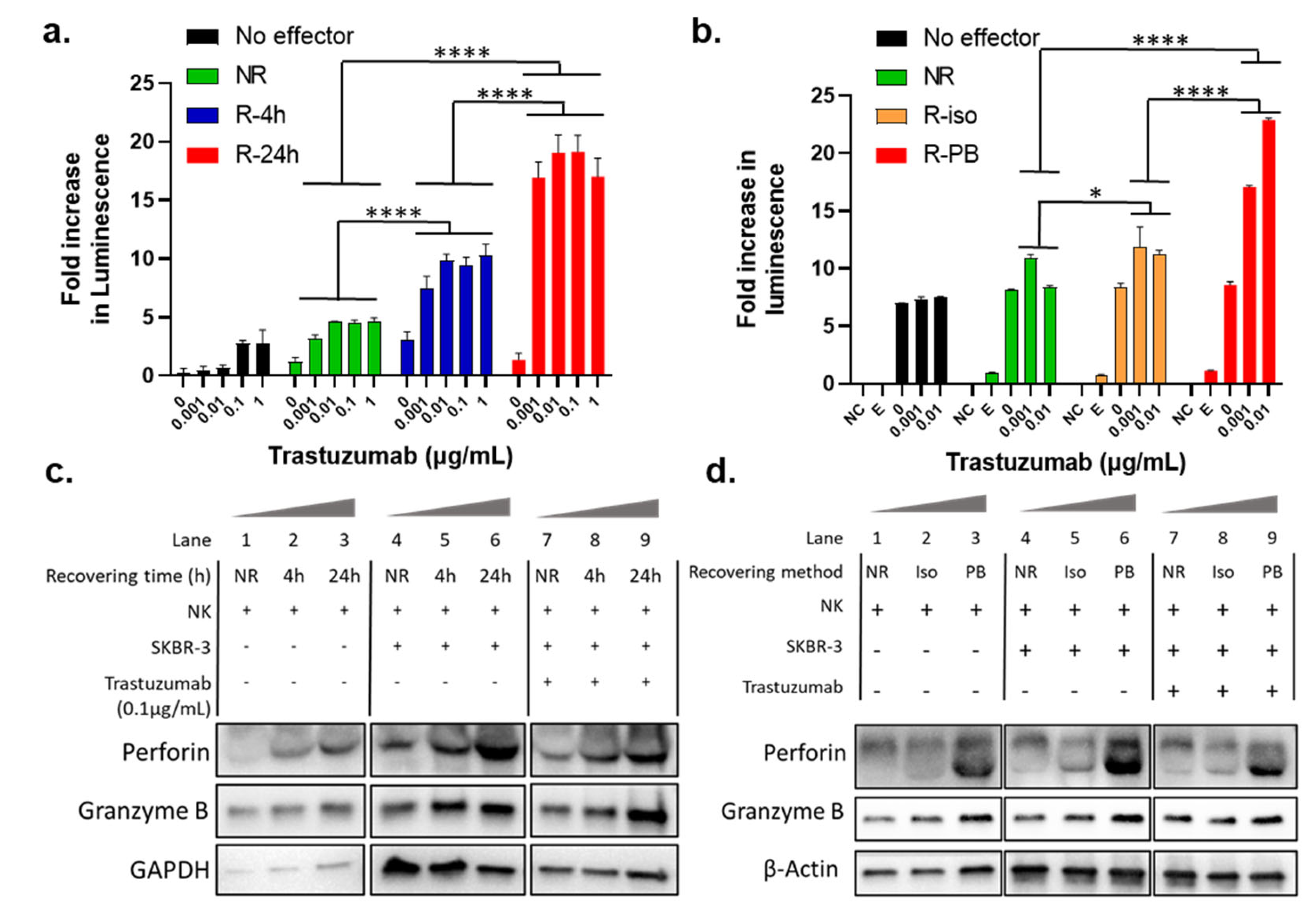
3.3. The Recovery Cultivation Condition Influences NK Cells Mediated ADCC Activity
3.4. Recovery Cultivation Influences the Expression Levels of Perforin and Granzyme B in NK Cells
3.5. Recovery Cultivation Is Essential for NK Cells to Secret Cytokines to Mediate ADCC Activity
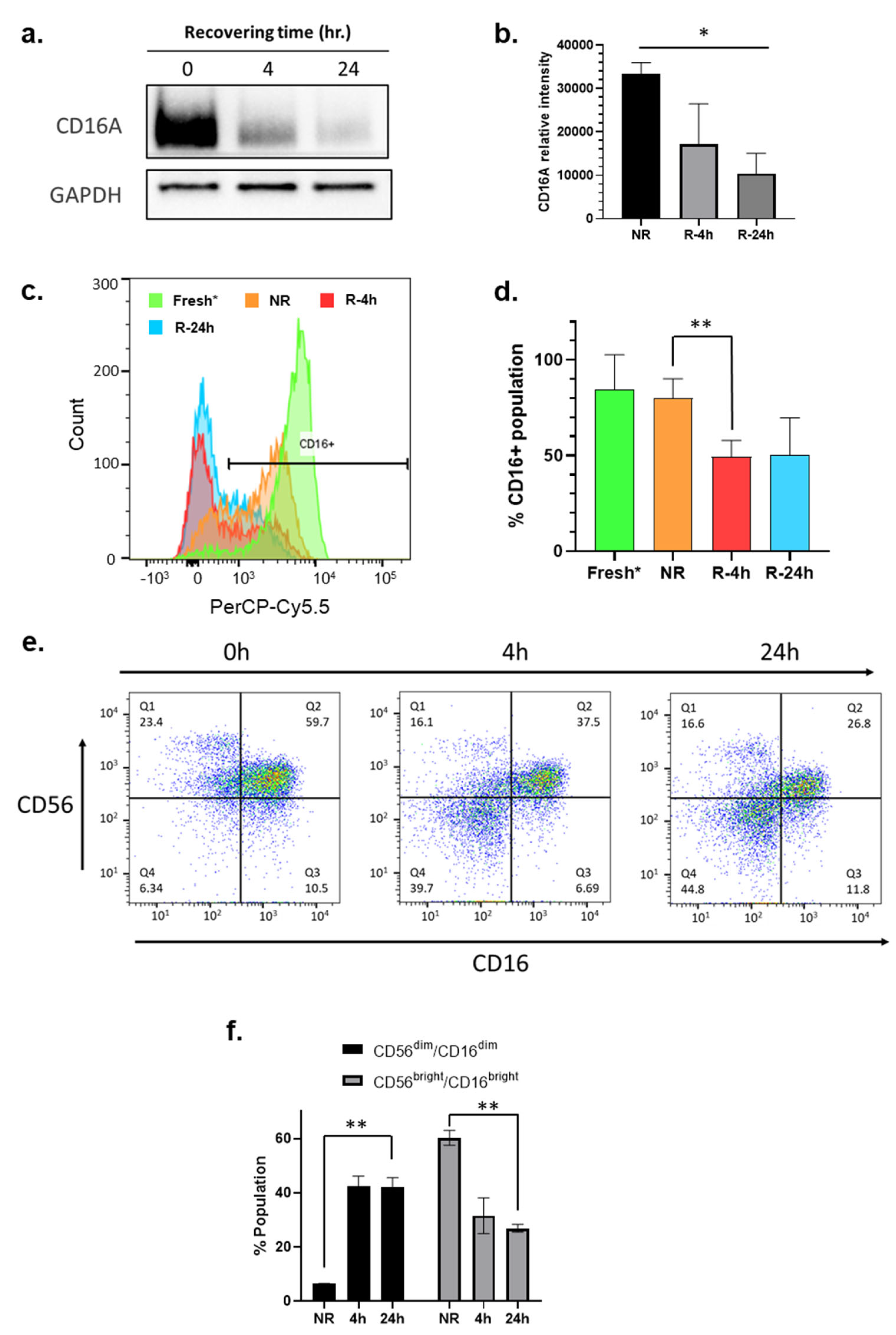
3.6. CD16/FcγRIII Expression Is Downregulated during the Recovery Culturing of NK Cells
3.7. Phenotyping Analysis Reveals the Formation of CD56dim/CD16 dim Subpopulation in NK Cells during the Recovery Cultivation Period
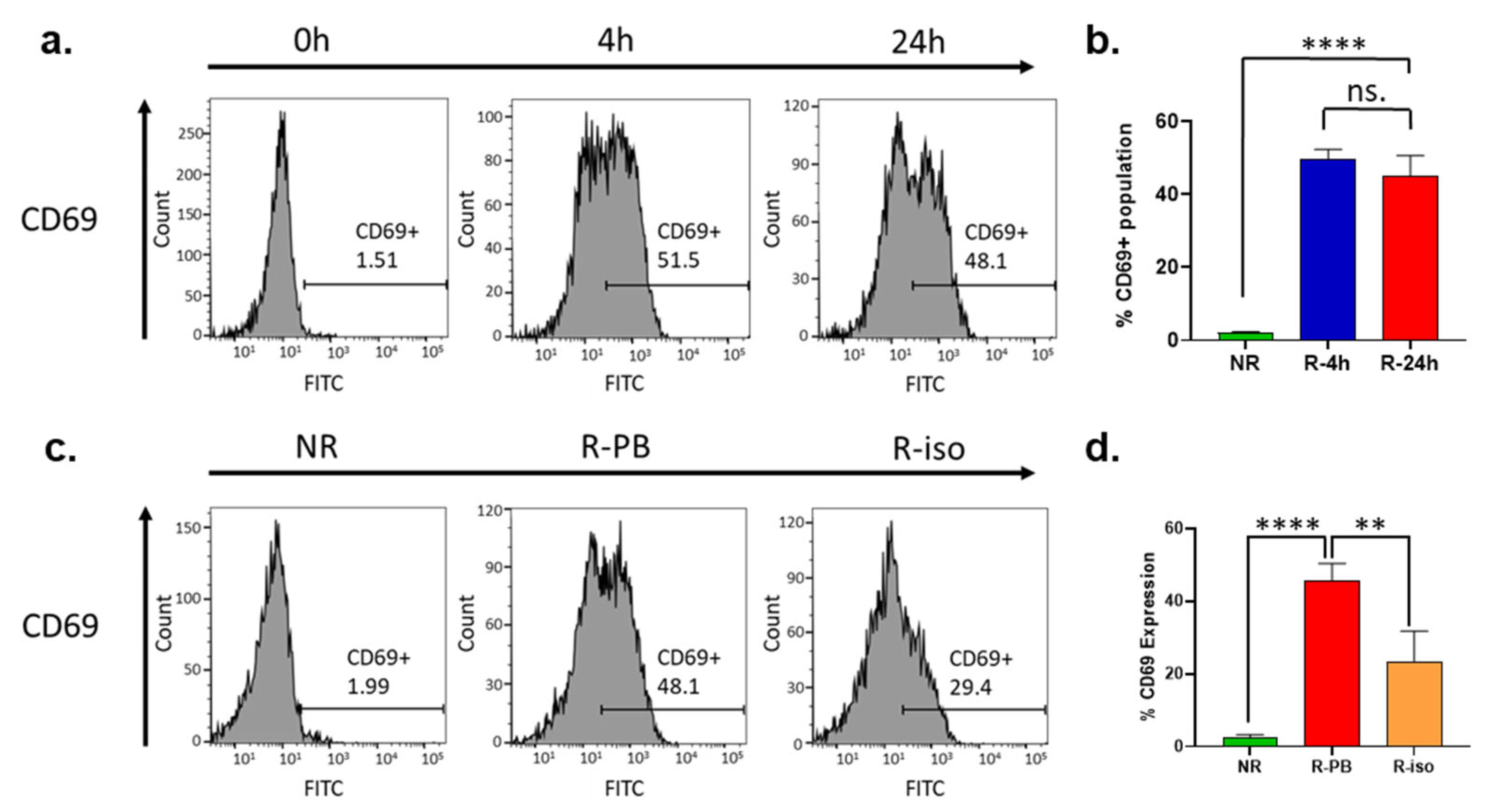
3.8. CD69 Expression Is Dramatically Upregulated on the Surface of NK Cells during the Recovery Cultivation Period
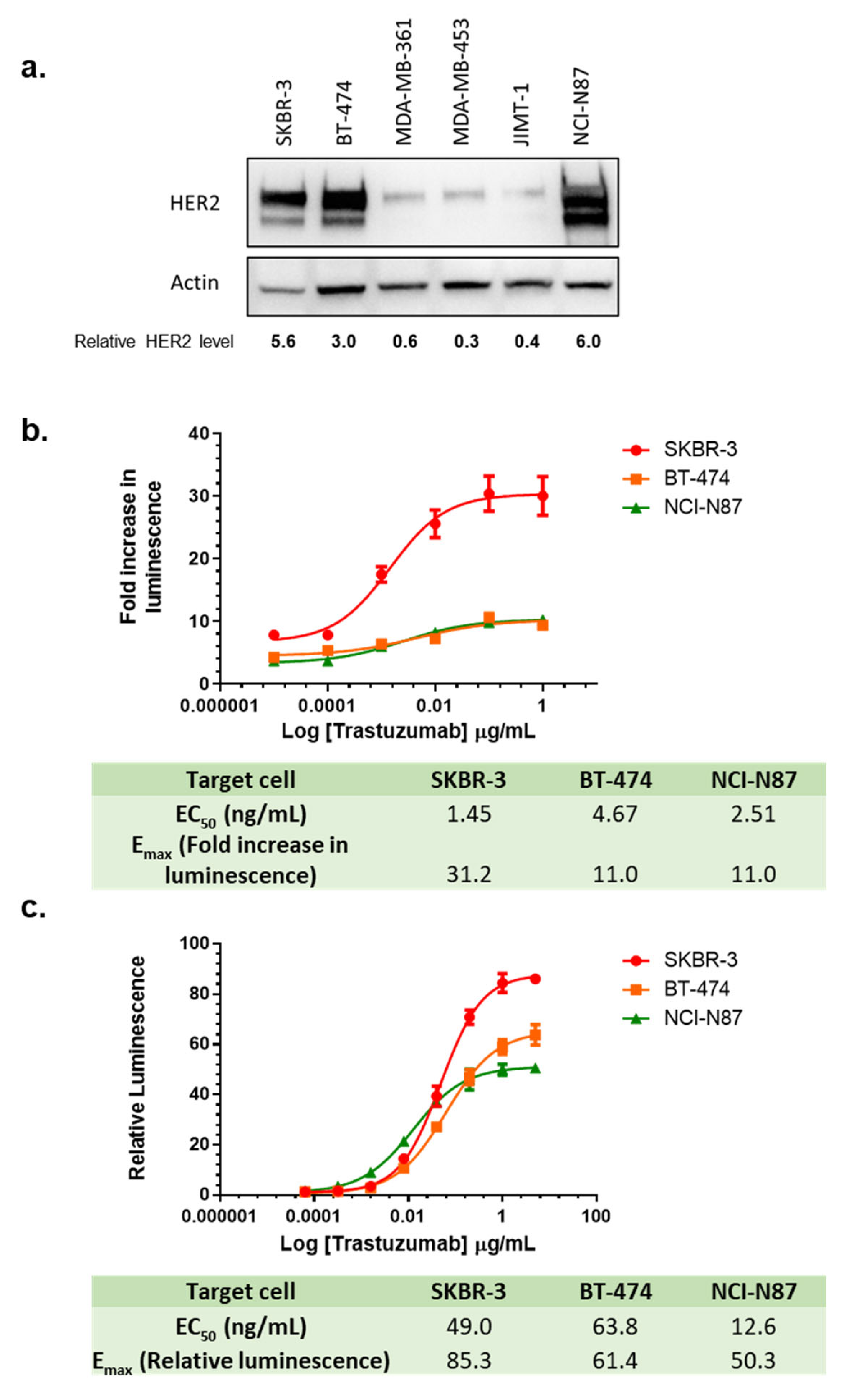
3.9. Choice of Target Cell Lines in the LDH ADCC Assay and ADCC Reporter Assay
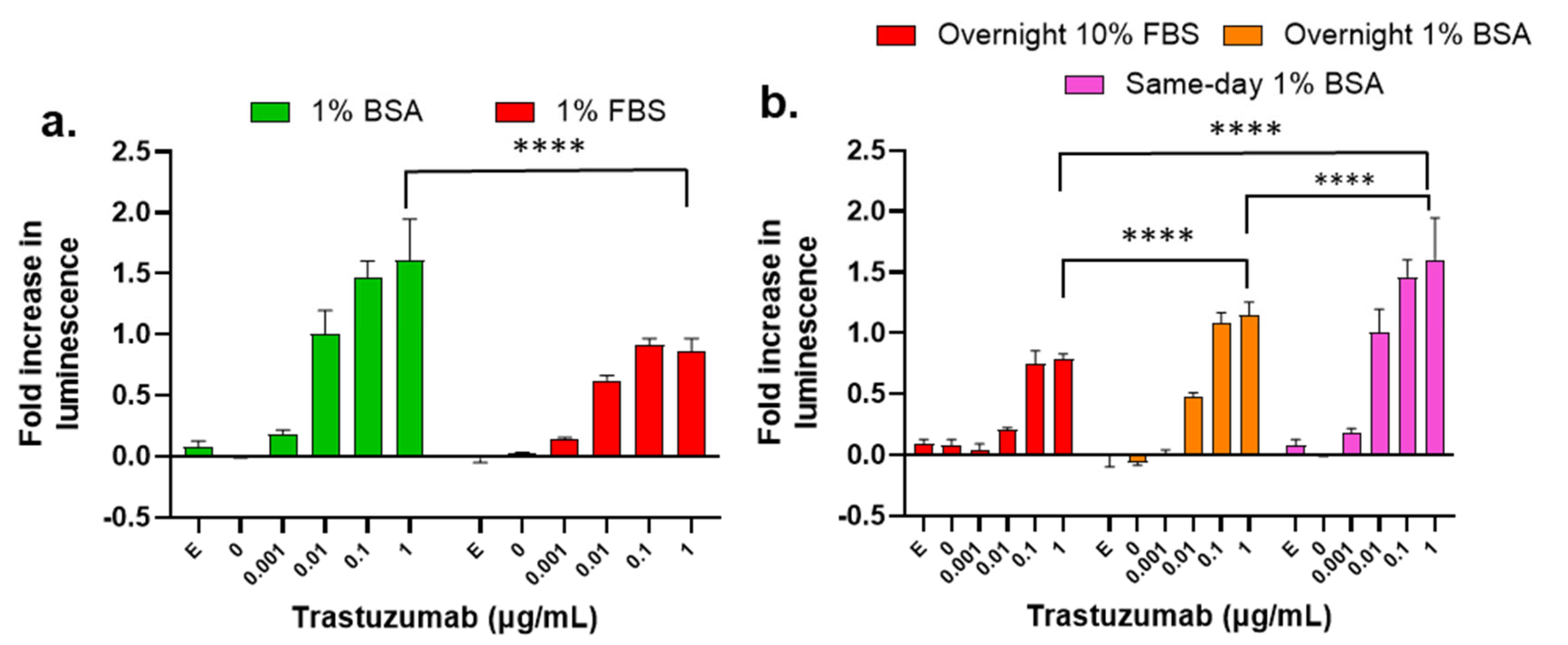
3.10. Influence of Assay Buffer, Target Cell Plating Method, and Treatment Time on the ADCC Activity of Trastuzumab
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schlam, I.; Swain, S.M. HER2-positive breast cancer and tyrosine kinase inhibitors: The time is now. NPJ Breast Cancer 2021, 7, 56. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Bang, Y.J. HER2-targeted therapies—A role beyond breast cancer. Nat. Rev. Clin. Oncol. 2020, 17, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Leyland-Jones, B.; Shak, S.; Fuchs, H.; Paton, V.; Bajamonde, A.; Fleming, T.; Eiermann, W.; Wolter, J.; Pegram, M.; et al. Use of chemotherapy plus a monoclonal antibody against HER2 for metastatic breast cancer that overexpresses HER2. N. Engl. J. Med. 2001, 344, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Modi, S.; Saura, C.; Yamashita, T.; Park, Y.H.; Kim, S.B.; Tamura, K.; Andre, F.; Iwata, H.; Ito, Y.; Tsurutani, J.; et al. Trastuzumab Deruxtecan in Previously Treated HER2-Positive Breast Cancer. N. Engl. J. Med. 2020, 382, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Romond, E.H.; Perez, E.A.; Bryant, J.; Suman, V.J.; Geyer, C.E., Jr.; Davidson, N.E.; Tan-Chiu, E.; Martino, S.; Paik, S.; Kaufman, P.A.; et al. Trastuzumab plus adjuvant chemotherapy for operable HER2-positive breast cancer. N. Engl. J. Med. 2005, 353, 1673–1684. [Google Scholar] [CrossRef] [PubMed]
- Cuellar, S. Integrating trastuzumab biosimilars and HER2-directed therapies into HER2-positive breast cancer management. Am. J. Manag. Care 2020, 26, S32–S40. [Google Scholar] [CrossRef] [PubMed]
- Rugo, H.S.; Pennella, E.J.; Gopalakrishnan, U.; Hernandez-Bronchud, M.; Herson, J.; Koch, H.F.; Loganathan, S.; Deodhar, S.; Marwah, A.; Manikhas, A.; et al. Final overall survival analysis of the phase 3 HERITAGE study demonstrates equivalence of trastuzumab-dkst to trastuzumab in HER2-positive metastatic breast cancer. Breast Cancer Res. Treat. 2021, 188, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Barbier, L.; Declerck, P.; Simoens, S.; Neven, P.; Vulto, A.G.; Huys, I. The arrival of biosimilar monoclonal antibodies in oncology: Clinical studies for trastuzumab biosimilars. Br. J. Cancer 2019, 121, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Khwaja, R.; Tang, P.; Nixon, N.; King, K.; Lupichuk, S. A Review of Trastuzumab Biosimilars in Early Breast Cancer and Real World Outcomes of Neoadjuvant MYL-1401O versus Reference Trastuzumab. Curr. Oncol. 2022, 29, 4224–4234. [Google Scholar] [CrossRef] [PubMed]
- Pegram, M.D.; Bondarenko, I.; Zorzetto, M.M.C.; Hingmire, S.; Iwase, H.; Krivorotko, P.V.; Lee, K.S.; Li, R.K.; Pikiel, J.; Aggarwal, R.; et al. PF-05280014 (a trastuzumab biosimilar) plus paclitaxel compared with reference trastuzumab plus paclitaxel for HER2-positive metastatic breast cancer: A randomised, double-blind study. Br. J. Cancer 2019, 120, 172–182. [Google Scholar] [CrossRef]
- Pivot, X.; Bondarenko, I.; Nowecki, Z.; Dvorkin, M.; Trishkina, E.; Ahn, J.H.; Im, S.A.; Sarosiek, T.; Chatterjee, S.; Wojtukiewicz, M.Z.; et al. A phase III study comparing SB3 (a proposed trastuzumab biosimilar) and trastuzumab reference product in HER2-positive early breast cancer treated with neoadjuvant-adjuvant treatment: Final safety, immunogenicity and survival results. Eur. J. Cancer 2018, 93, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Stebbing, J.; Baranau, Y.; Baryash, V.; Manikhas, A.; Moiseyenko, V.; Dzagnidze, G.; Zhavrid, E.; Boliukh, D.; Stroyakovskii, D.; Pikiel, J.; et al. CT-P6 compared with reference trastuzumab for HER2-positive breast cancer: A randomised, double-blind, active-controlled, phase 3 equivalence trial. Lancet Oncol. 2017, 18, 917–928. [Google Scholar] [CrossRef] [PubMed]
- von Minckwitz, G.; Colleoni, M.; Kolberg, H.C.; Morales, S.; Santi, P.; Tomasevic, Z.; Zhang, N.; Hanes, V. Efficacy and safety of ABP 980 compared with reference trastuzumab in women with HER2-positive early breast cancer (LILAC study): A randomised, double-blind, phase 3 trial. Lancet Oncol. 2018, 19, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Biosimilar Development, Review, and Approval. Available online: www.fda.gov/drugs/biosimilars/biosimilar-development-review-and-approval#indication (accessed on 16 April 2024).
- Hudis, C.A. Trastuzumab—Mechanism of action and use in clinical practice. N. Engl. J. Med. 2007, 357, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Maadi, H.; Soheilifar, M.H.; Choi, W.S.; Moshtaghian, A.; Wang, Z. Trastuzumab Mechanism of Action; 20 Years of Research to Unravel a Dilemma. Cancers 2021, 13, 3540. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liu, S. Focusing on NK cells and ADCC: A promising immunotherapy approach in targeted therapy for HER2-positive breast cancer. Front. Immunol. 2022, 13, 1083462. [Google Scholar] [CrossRef] [PubMed]
- Lo Nigro, C.; Macagno, M.; Sangiolo, D.; Bertolaccini, L.; Aglietta, M.; Merlano, M.C. NK-mediated antibody-dependent cell-mediated cytotoxicity in solid tumors: Biological evidence and clinical perspectives. Ann. Transl. Med. 2019, 7, 105. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Garvin, D.; Paguio, A.; Moravec, R.; Engel, L.; Fan, F.; Surowy, T. Development of a robust reporter-based ADCC assay with frozen, thaw-and-use cells to measure Fc effector function of therapeutic antibodies. J. Immunol. Methods 2014, 414, 69–81. [Google Scholar] [CrossRef]
- Disis, M.L.; dela Rosa, C.; Goodell, V.; Kuan, L.Y.; Chang, J.C.; Kuus-Reichel, K.; Clay, T.M.; Kim Lyerly, H.; Bhatia, S.; Ghanekar, S.A.; et al. Maximizing the retention of antigen specific lymphocyte function after cryopreservation. J. Immunol. Methods 2006, 308, 13–18. [Google Scholar] [CrossRef]
- Mark, C.; Czerwinski, T.; Roessner, S.; Mainka, A.; Horsch, F.; Heublein, L.; Winterl, A.; Sanokowski, S.; Richter, S.; Bauer, N.; et al. Cryopreservation impairs 3-D migration and cytotoxicity of natural killer cells. Nat. Commun. 2020, 11, 5224. [Google Scholar] [CrossRef]
- Ramachandran, H.; Laux, J.; Moldovan, I.; Caspell, R.; Lehmann, P.V.; Subbramanian, R.A. Optimal thawing of cryopreserved peripheral blood mononuclear cells for use in high-throughput human immune monitoring studies. Cells 2012, 1, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Tollerud, D.J.; Brown, L.M.; Clark, J.W.; Neuland, C.Y.; Mann, D.L.; Pankiw-Trost, L.K.; Blattner, W.A. Cryopreservation and long-term liquid nitrogen storage of peripheral blood mononuclear cells for flow cytometry analysis: Effects on cell subset proportions and fluorescence intensity. J. Clin. Lab. Anal. 1991, 5, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Joo, Y.; Lee, E.J.; Byeon, Y.; Kim, J.H.; Pyo, K.H.; Kim, Y.S.; Lim, S.M.; Kilbride, P.; Iyer, R.K.; et al. Successful expansion and cryopreservation of human natural killer cell line NK-92 for clinical manufacturing. PLoS ONE 2024, 19, e0294857. [Google Scholar] [CrossRef] [PubMed]
- Oyer, J.L.; Croom-Perez, T.J.; Dieffenthaller, T.A.; Robles-Carillo, L.D.; Gitto, S.B.; Altomare, D.A.; Copik, A.J. Cryopreserved PM21-Particle-Expanded Natural Killer Cells Maintain Cytotoxicity and Effector Functions. Front. Immunol. 2022, 13, 861681. [Google Scholar] [CrossRef] [PubMed]
- Voshol, H.; Dullens, H.F.; Den Otter, W.; Vliegenthart, J.F. Human natural killer cells: A convenient purification procedure and the influence of cryopreservation on cytotoxic activity. J. Immunol. Methods 1993, 165, 21–30. [Google Scholar] [CrossRef]
- Mata, M.M.; Mahmood, F.; Sowell, R.T.; Baum, L.L. Effects of cryopreservation on effector cells for antibody dependent cell-mediated cytotoxicity (ADCC) and natural killer (NK) cell activity in Cr-51-release and CD107a assays. J. Immunol. Methods 2014, 406, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pross, H.F.; Maroun, J.A. The standardization of NK cell assays for use in studies of biological response modifiers. J. Immunol. Methods 1984, 68, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Avila, I.V.; Marmol, P.; Kiessling, R.; de Coaña, Y.P. Evaluating Antibody-Dependent Cell-Mediated Cytotoxicity by Chromium Release Assay. Immune Checkp. Blockade: Methods Protoc. 2019, 1913, 167–179. [Google Scholar] [CrossRef]
- Buller, C.W.; Mathew, S.O. NK Cell Isolation and Cytotoxicity by Radioactive Chromium Release Assay and DELFIA-EuTDA Cytotoxicity Assay. In Natural Killer (NK) Cells: Methods and, Protocols; Shimasaki, N., Ed.; Springer: New York, NY, USA, 2022; pp. 221–233. [Google Scholar]
- Wieckowski, S.; Avenal, C.; Orjalo, A.V., Jr.; Gygax, D.; Cymer, F. Toward a Better Understanding of Bioassays for the Development of Biopharmaceuticals by Exploring the Structure-Antibody-Dependent Cellular Cytotoxicity Relationship in Human Primary Cells. Front. Immunol. 2020, 11, 552596. [Google Scholar] [CrossRef]
- Broussas, M.; Broyer, L.; Goetsch, L. Evaluation of antibody-dependent cell cytotoxicity using lactate dehydrogenase (LDH) measurement. Methods Mol. Biol. 2013, 988, 305–317. [Google Scholar] [CrossRef]
- Vincken, R.; Ruiz-Saenz, A. A co-culture model system to quantify antibody-dependent cellular cytotoxicity in human breast cancer cells using an engineered natural killer cell line. STAR Protoc. 2023, 4, 102224. [Google Scholar] [CrossRef] [PubMed]
- Petricevic, B.; Laengle, J.; Singer, J.; Sachet, M.; Fazekas, J.; Steger, G.; Bartsch, R.; Jensen-Jarolim, E.; Bergmann, M. Trastuzumab mediates antibody-dependent cell-mediated cytotoxicity and phagocytosis to the same extent in both adjuvant and metastatic HER2/neu breast cancer patients. J. Transl. Med. 2013, 11, 307. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Kitano, S.; Aikawa, H.; Kuchiba, A.; Hayashi, M.; Yamamoto, N.; Tamura, K.; Hamada, A. A novel method for evaluating antibody-dependent cell-mediated cytotoxicity by flowcytometry using cryopreserved human peripheral blood mononuclear cells. Sci. Rep. 2016, 6, 19772. [Google Scholar] [CrossRef] [PubMed]
- Garvin, D.; Stecha, P.; Gilden, J.; Wang, J.; Grailer, J.; Hartnett, J.; Fan, F.; Cong, M.; Cheng, Z.J. Determining ADCC Activity of Antibody-Based Therapeutic Molecules using Two Bioluminescent Reporter-Based Bioassays. Curr. Protoc. 2021, 1, e296. [Google Scholar] [CrossRef] [PubMed]
- Kute, T.; Stehle, J.R.; Ornelles, D.; Walker, N.; Delbono, O.; Vaughn, J.P. Understanding key assay parameters that affect measurements of trastuzumab-mediated ADCC against Her2 positive breast cancer cells. Oncoimmunology 2012, 1, 810–821. [Google Scholar] [CrossRef]
- Tang, Y.; Lou, J.; Alpaugh, R.K.; Robinson, M.K.; Marks, J.D.; Weiner, L.M. Regulation of antibody-dependent cellular cytotoxicity by IgG intrinsic and apparent affinity for target antigen. J. Immunol. 2007, 179, 2815–2823. [Google Scholar] [CrossRef]
- Klingemann, H. Challenges of cancer therapy with natural killer cells. Cytotherapy 2015, 17, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.P.; Jaw, J.J.; Stutzman, N.C.; Zou, Z.C.; Sun, P.D. Natural killer cell-produced IFN-γ and TNF-α induce target cell cytolysis through up-regulation of ICAM-1. J. Leukoc. Biol. 2012, 91, 299–309. [Google Scholar] [CrossRef]
- Ackerman, M.E.; Nimmerjahn, F. Antibody Fc: Linking Adaptive and Innate Immunity; Academic Press: Cambridge, MA, USA, 2014; pp. 1–363. [Google Scholar]
- Borrego, F.; Robertson, M.J.; Ritz, J.; Pena, J.; Solana, R. CD69 is a stimulatory receptor for natural killer cell and its cytotoxic effect is blocked by CD94 inhibitory receptor. Immunology 1999, 97, 159–165. [Google Scholar] [CrossRef]
- Hui, X.W.; Yuan, C.; Cao, W.R.; Ge, W.L.; Zhang, D.; Dan, M.; Zhao, Q.; Liu, B.N.; Yao, B. An Innovative Site-Specific Anti-HER2 Antibody-Drug Conjugate with High Homogeneity and Improved Therapeutic Index. Oncotargets Ther. 2022, 15, 331–343. [Google Scholar] [CrossRef]
- Leibson, P.J. Signal transduction during natural killer cell activation: Inside the mind of a killer. Immunity 1997, 6, 655–661. [Google Scholar] [CrossRef]
- Osinska, I.; Popko, K.; Demkow, U. Perforin: An important player in immune response. Cent. Eur. J. Immunol. 2014, 39, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.; Jason, J.; Wallace, K.; Slaughter, J.; Whatley, V.; Han, A.; Nwanyanwu, O.C.; Kazembe, P.N.; Dobbie, H.; Archibald, L.; et al. Spontaneous cytokine production and its effect on induced production. Clin. Vaccine Immunol. 2002, 9, 1049–1056. [Google Scholar] [CrossRef]
- Rabinowich, H.; Sedlmayr, P.; Herberman, R.B.; Whiteside, T.L. Response of human NK cells to IL-6 alterations of the cell surface phenotype, adhesion to fibronectin and laminin, and tumor necrosis factor-alpha/beta secretion. J. Immunol. 1993, 150, 4844–4855. [Google Scholar] [CrossRef]
- Cibrian, D.; Sanchez-Madrid, F. CD69: From activation marker to metabolic gatekeeper. Eur. J. Immunol. 2017, 47, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Cabrera, M.; Santis, A.G.; Fernandez-Ruiz, E.; Blacher, R.; Esch, F.; Sanchez-Mateos, P.; Sanchez-Madrid, F. Molecular cloning, expression, and chromosomal localization of the human earliest lymphocyte activation antigen AIM/CD69, a new member of the C-type animal lectin superfamily of signal-transmitting receptors. J. Exp. Med. 1993, 178, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, S.F.; Ramsdell, F.; Hjerrild, K.A.; Armitage, R.J.; Grabstein, K.H.; Hennen, K.B.; Farrah, T.; Fanslow, W.C.; Shevach, E.M.; Alderson, M.R. Molecular characterization of the early activation antigen CD69: A type II membrane glycoprotein related to a family of natural killer cell activation antigens. Eur. J. Immunol. 1993, 23, 1643–1648. [Google Scholar] [CrossRef]
- Marden, C.M.; North, J.; Anderson, R.; Bakhsh, I.A.; Addison, E.; Pittman, H.; Mackinnon, S.; Lowdell, M.W. CD69 is required for activated NK cell-mediated killing of resistant targets. Blood 2005, 106, 928a–929a. [Google Scholar] [CrossRef]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Romee, R.; Foley, B.; Lenvik, T.; Wang, Y.; Zhang, B.; Ankarlo, D.; Luo, X.; Cooley, S.; Verneris, M.; Walcheck, B.; et al. NK cell CD16 surface expression and function is regulated by a disintegrin and metalloprotease-17 (ADAM17). Blood 2013, 121, 3599–3608. [Google Scholar] [CrossRef]
- Srpan, K.; Ambrose, A.; Karampatzakis, A.; Saeed, M.; Cartwright, A.N.R.; Guldevall, K.; De Matos, G.; Onfelt, B.; Davis, D.M. Shedding of CD16 disassembles the NK cell immune synapse and boosts serial engagement of target cells. J. Cell Biol. 2018, 217, 3267–3283. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, S.; Weiss, E.; Schmitt, A.L.; Schlegel, J.; Burgert, A.; Terpitz, U.; Sauer, M.; Moretta, L.; Sivori, S.; Leonhardt, I.; et al. CD56 Is a Pathogen Recognition Receptor on Human Natural Killer Cells. Sci. Rep. UK 2017, 7. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, H.; Endo, Y.; Wu, W.J. Define Critical Parameters of Trastuzumab-Mediated ADCC Assays via Assay Optimization Processes, Focusing on the Impact of Cryopreserved Effector Cells on Assay Performance. Cancers 2024, 16, 2367. https://doi.org/10.3390/cancers16132367
Peng H, Endo Y, Wu WJ. Define Critical Parameters of Trastuzumab-Mediated ADCC Assays via Assay Optimization Processes, Focusing on the Impact of Cryopreserved Effector Cells on Assay Performance. Cancers. 2024; 16(13):2367. https://doi.org/10.3390/cancers16132367
Chicago/Turabian StylePeng, Hanjing, Yukinori Endo, and Wen Jin Wu. 2024. "Define Critical Parameters of Trastuzumab-Mediated ADCC Assays via Assay Optimization Processes, Focusing on the Impact of Cryopreserved Effector Cells on Assay Performance" Cancers 16, no. 13: 2367. https://doi.org/10.3390/cancers16132367