
Analysis of Expression and Regulation of AKR1C2 in HPV-Positive and -Negative Oropharyngeal Squamous Cell Carcinoma

 , , ,
, , ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Materials
2.2. Ethics Statement
2.3. Immunochemistry
2.4. Cell Culture and Transfection
2.5. RT-qPCR Expression Analysis
2.6. Statistics
3. Results
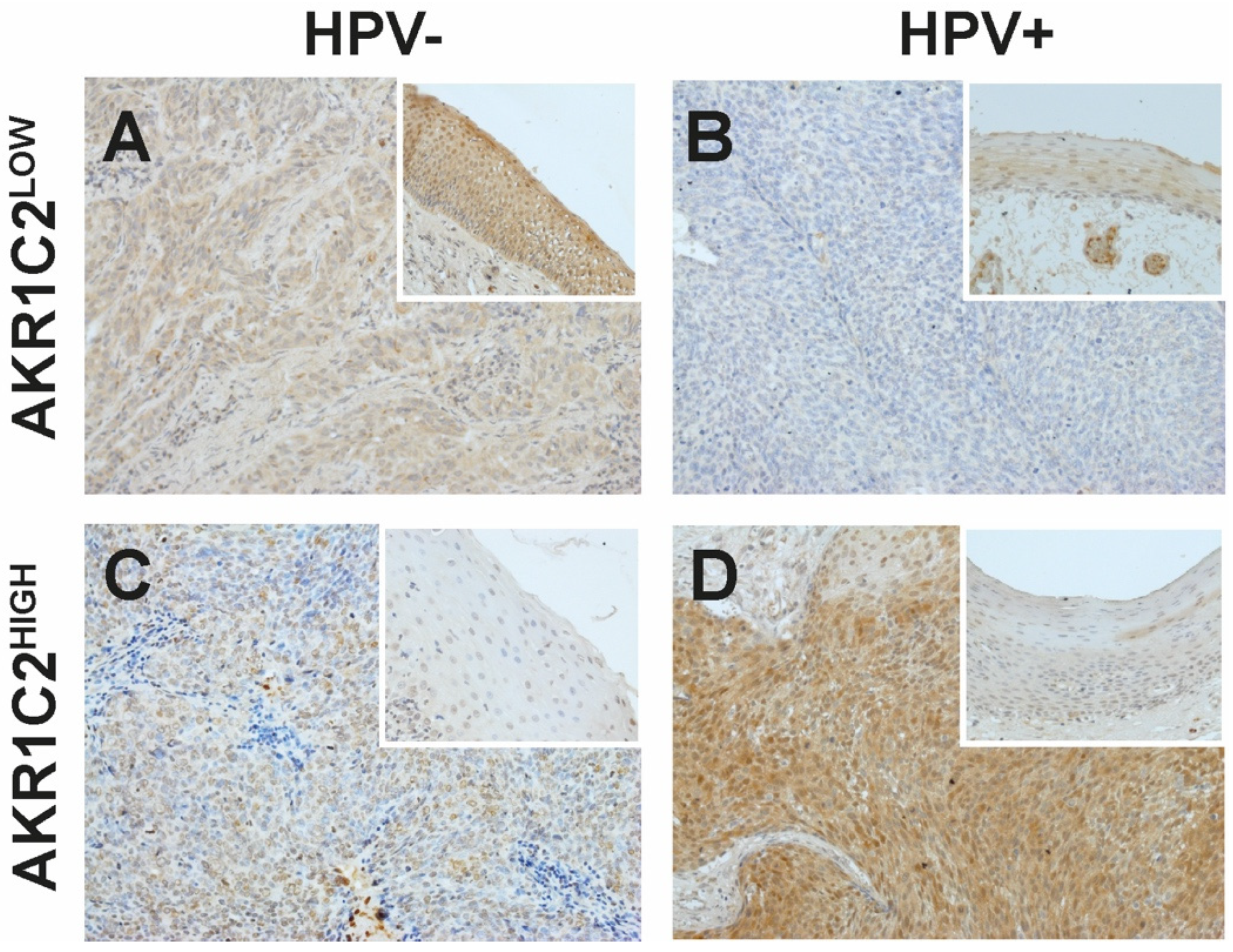
3.1. Immunohistochemical Detection of AKR1C2
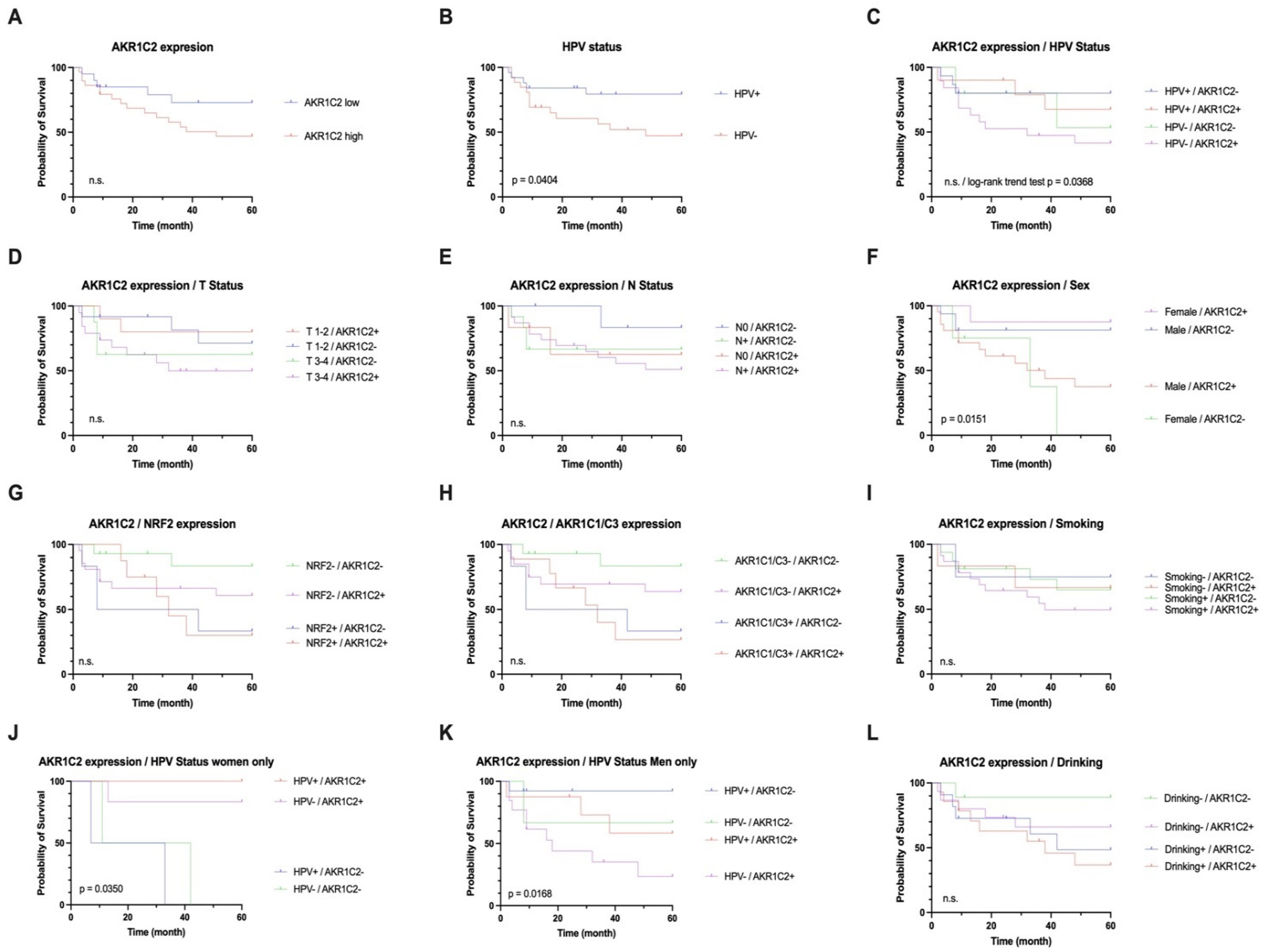
3.2. AKR1C2 Protein Expression, HPV, and Survival in OPSCC
3.3. Correlation of AKR1C2 with AKR1C1 and AKR1C3 and NRF2
3.4. Effects of HPV16-E6*I on AKR1C2 mRNA Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.B.; Gorsky, M.; Cabay, R.J.; Day, T.; Gonsalves, W. Screening for and diagnosis of oral premalignant lesions and oropharyngeal squamous cell carcinoma: Role of primary care physicians. Can. Fam. Physician 2008, 54, 870–875. [Google Scholar] [PubMed]
- De Franca, G.M.; da Silva, W.R.; Medeiros, C.K.S.; Junior, J.F.; de Moura Santos, E.; Galvao, H.C. Five-year survival and prognostic factors for oropharyngeal squamous cell carcinoma: Retrospective cohort of a cancer center. Oral Maxillofac. Surg. 2022, 26, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Pelucchi, C.; Gallus, S.; Garavello, W.; Bosetti, C.; La Vecchia, C. Alcohol and tobacco use, and cancer risk for upper aerodigestive tract and liver. Eur. J. Cancer Prev. 2008, 17, 340–344. [Google Scholar] [CrossRef]
- Olthof, N.C.; Speel, E.J.; Kolligs, J.; Haesevoets, A.; Henfling, M.; Ramaekers, F.C.; Preuss, S.F.; Drebber, U.; Wieland, U.; Silling, S.; et al. Comprehensive analysis of HPV16 integration in OSCC reveals no significant impact of physical status on viral oncogene and virally disrupted human gene expression. PLoS ONE 2014, 9, e88718. [Google Scholar] [CrossRef]
- Ferlay, J.; Ervik, M.; Lam, F.; Laversanne, M.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Today; International Agency for Research on Cancer: Lyon, France. Available online: https://gco.iarc.fr/today/home (accessed on 4 June 2024).
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, K.; Suk, R.; Chiao, E.Y.; Chhatwal, J.; Qiu, P.; Wilkin, T.; Nyitray, A.G.; Sikora, A.G.; Deshmukh, A.A. Oral Human Papillomavirus Infection: Differences in Prevalence Between Sexes and Concordance with Genital Human Papillomavirus Infection, NHANES 2011 to 2014. Ann. Intern. Med. 2017, 167, 714–724. [Google Scholar] [CrossRef]
- CDC. CfDCaP. HPV-Associated Cancer Statistics. Available online: https://www.cdc.gov/cancer/hpv/cases.html?CDC_AAref_Val=https://www.cdc.gov/cancer/hpv/statistics/cases.htm (accessed on 4 June 2024).
- Lechner, M.; Liu, J.; Masterson, L.; Fenton, T.R. HPV-associated oropharyngeal cancer: Epidemiology, molecular biology and clinical management. Nat. Rev. Clin. Oncol. 2022, 19, 306–327. [Google Scholar] [CrossRef]
- Hoffmann, M.; Tribius, S. HPV and oropharyngeal squamous cell cancer in the 8th edition of the TNM classification. Laryngorhinootologie 2018, 97, 641–649. [Google Scholar] [CrossRef]
- Zhan, K.Y.; Puram, S.V.; Li, M.M.; Silverman, D.A.; Agrawal, A.A.; Ozer, E.; Old, M.O.; Carrau, R.L.; Rocco, J.W.; Higgins, K.M.; et al. National treatment trends in human papillomavirus-positive oropharyngeal squamous cell carcinoma. Cancer 2020, 126, 1295–1305. [Google Scholar] [CrossRef]
- Ang, K.K.; Harris, J.; Wheeler, R.; Weber, R.; Rosenthal, D.I.; Nguyen-Tan, P.F.; Westra, W.H.; Chung, C.H.; Jordan, R.C.; Lu, C.; et al. Human papillomavirus and survival of patients with oropharyngeal cancer. N. Engl. J. Med. 2010, 363, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Klussmann, J.P.; Mooren, J.J.; Lehnen, M.; Claessen, S.M.; Stenner, M.; Huebbers, C.U.; Weissenborn, S.J.; Wedemeyer, I.; Preuss, S.F.; Straetmans, J.M.; et al. Genetic signatures of HPV-related and unrelated oropharyngeal carcinoma and their prognostic implications. Clin. Cancer Res. 2009, 15, 1779–1786. [Google Scholar] [CrossRef] [PubMed]
- Mooren, J.J.; Gultekin, S.E.; Straetmans, J.M.; Haesevoets, A.; Peutz-Kootstra, C.J.; Huebbers, C.U.; Dienes, H.P.; Wieland, U.; Ramaekers, F.C.; Kremer, B.; et al. P16(INK4A) immunostaining is a strong indicator for high-risk-HPV-associated oropharyngeal carcinomas and dysplasias, but is unreliable to predict low-risk-HPV-infection in head and neck papillomas and laryngeal dysplasias. Int. J. Cancer 2014, 134, 2108–2117. [Google Scholar] [CrossRef] [PubMed]
- Olthof, N.C.; Straetmans, J.M.; Snoeck, R.; Ramaekers, F.C.; Kremer, B.; Speel, E.J. Next-generation treatment strategies for human papillomavirus-related head and neck squamous cell carcinoma: Where do we go? Rev. Med. Virol. 2012, 22, 88–105. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- James, C.D.; Morgan, I.M.; Bristol, M.L. The Relationship between Estrogen-Related Signaling and Human Papillomavirus Positive Cancers. Pathogens 2020, 9, 403. [Google Scholar] [CrossRef]
- Penning, T.M. Aldo-Keto Reductase Regulation by the Nrf2 System: Implications for Stress Response, Chemotherapy Drug Resistance, and Carcinogenesis. Chem. Res. Toxicol. 2017, 30, 162–176. [Google Scholar] [CrossRef]
- Huebbers, C.U.; Verhees, F.; Poluschkin, L.; Olthof, N.C.; Kolligs, J.; Siefer, O.G.; Henfling, M.; Ramaekers, F.C.S.; Preuss, S.F.; Beutner, D.; et al. Upregulation of AKR1C1 and AKR1C3 expression in OPSCC with integrated HPV16 and HPV-negative tumors is an indicator of poor prognosis. Int. J. Cancer 2019, 144, 2465–2477. [Google Scholar] [CrossRef]
- Atalla, A.; Maser, E. Characterization of enzymes participating in carbonyl reduction of 4-methylnitrosamino-1-(3-pyridyl)-1-butanone (NNK) in human placenta. Chem. Biol. Interact. 2001, 130–132, 737–748. [Google Scholar] [CrossRef]
- Penning, T.M.; Jonnalagadda, S.; Trippier, P.C.; Rizner, T.L. Aldo-Keto Reductases and Cancer Drug Resistance. Pharmacol. Rev. 2021, 73, 1150–1171. [Google Scholar] [CrossRef]
- Penning, T.M.; Byrns, M.C. Steroid hormone transforming aldo-keto reductases and cancer. Ann. N. Y. Acad. Sci. 2009, 1155, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Rizner, T.L. Enzymes of the AKR1B and AKR1C Subfamilies and Uterine Diseases. Front. Pharmacol. 2012, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Gang, B.P.; Bassi, C.; Wakeham, A.; Baniasadi, S.P.; Hao, Z.; Li, W.Y.; Cescon, D.W.; Li, Y.T.; Molyneux, S.; et al. Estrogen controls the survival of BRCA1-deficient cells via a PI3K-NRF2-regulated pathway. Proc. Natl. Acad. Sci. USA 2014, 111, 4472–4477. [Google Scholar] [CrossRef]
- Sannigrahi, M.K.; Rajagopalan, P.; Lai, L.; Liu, X.; Sahu, V.; Nakagawa, H.; Jalaly, J.B.; Brody, R.M.; Morgan, I.M.; Windle, B.E.; et al. HPV E6 regulates therapy responses in oropharyngeal cancer by repressing the PGC-1alpha/ERRalpha axis. JCI Insight 2022, 7, e159600. [Google Scholar] [CrossRef] [PubMed]
- Klasen, C.; Wuerdemann, N.; Rothbart, P.; Prinz, J.; Eckel, H.N.C.; Suchan, M.; Kopp, C.; Johannsen, J.; Ziogas, M.; Charpentier, A.; et al. Sex-specific aspects in patients with oropharyngeal squamous cell carcinoma: A bicentric cohort study. BMC Cancer 2023, 23, 1054. [Google Scholar] [CrossRef]
- Wanichwatanadecha, P.; Sirisrimangkorn, S.; Kaewprag, J.; Ponglikitmongkol, M. Transactivation activity of human papillomavirus type 16 E6*I on aldo-keto reductase genes enhances chemoresistance in cervical cancer cells. J. Gen. Virol. 2012, 93, 1081–1092. [Google Scholar] [CrossRef]
- AKR1C2. Available online: https://www.proteinatlas.org/ENSG00000151632-AKR1C2 (accessed on 13 April 2024).
- Uzun, S.; Korkmaz, Y.; Wuerdemann, N.; Arolt, C.; Puladi, B.; Siefer, O.G.; Donmez, H.G.; Hufbauer, M.; Akgul, B.; Klussmann, J.P.; et al. Comprehensive Analysis of VEGFR2 Expression in HPV-Positive and -Negative OPSCC Reveals Differing VEGFR2 Expression Patterns. Cancers 2021, 13, 5221. [Google Scholar] [CrossRef]
- Penning, T.M.; Wangtrakuldee, P.; Auchus, R.J. Structural and Functional Biology of Aldo-Keto Reductase Steroid-Transforming Enzymes. Endocr. Rev. 2019, 40, 447–475. [Google Scholar] [CrossRef]
- Idda, M.L.; Campesi, I.; Fiorito, G.; Vecchietti, A.; Urru, S.A.M.; Solinas, M.G.; Franconi, F.; Floris, M. Sex-Biased Expression of Pharmacogenes across Human Tissues. Biomolecules 2021, 11, 1206. [Google Scholar] [CrossRef]
- Ji, Q.; Aoyama, C.; Nien, Y.D.; Liu, P.I.; Chen, P.K.; Chang, L.; Stanczyk, F.Z.; Stolz, A. Selective loss of AKR1C1 and AKR1C2 in breast cancer and their potential effect on progesterone signaling. Cancer Res. 2004, 64, 7610–7617. [Google Scholar] [CrossRef]
- Detlefsen, A.J.; Paulukinas, R.D.; Penning, T.M. Germline Mutations in Steroid Metabolizing Enzymes: A Focus on Steroid Transforming Aldo-Keto Reductases. Int. J. Mol. Sci. 2023, 24, 1873. [Google Scholar] [CrossRef] [PubMed]
- Oakley, J.; Hill, M.; Giess, A.; Tanguy, M.; Elgar, G. Long read sequencing characterises a novel structural variant, revealing underactive AKR1C1 with overactive AKR1C2 as a possible cause of severe chronic fatigue. J. Transl. Med. 2023, 21, 825. [Google Scholar] [CrossRef]
- Paget-Bailly, P.; Meznad, K.; Bruyere, D.; Perrard, J.; Herfs, M.; Jung, A.C.; Mougin, C.; Pretet, J.L.; Baguet, A. Comparative RNA sequencing reveals that HPV16 E6 abrogates the effect of E6*I on ROS metabolism. Sci. Rep. 2019, 9, 5938. [Google Scholar] [CrossRef]
- Olmedo-Nieva, L.; Munoz-Bello, J.O.; Contreras-Paredes, A.; Lizano, M. The Role of E6 Spliced Isoforms (E6*) in Human Papillomavirus-Induced Carcinogenesis. Viruses 2018, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Huang, T.J.; Zhang, X.K.; Xie, Y.J.; Lin, S.T.; Luo, F.F.; Meng, D.F.; Hu, H.; Wang, J.; Peng, L.X.; et al. AKR1C2 acts as a targetable oncogene in esophageal squamous cell carcinoma via activating PI3K/AKT signaling pathway. J. Cell. Mol. Med. 2020, 24, 9999–10012. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, Q.; Fung, K.M.; Lin, H.K. AKR1C2 and AKR1C3 mediated prostaglandin D2 metabolism augments the PI3K/Akt proliferative signaling pathway in human prostate cancer cells. Mol. Cell. Endocrinol. 2008, 289, 60–66. [Google Scholar] [CrossRef]
- Lui, V.W.; Hedberg, M.L.; Li, H.; Vangara, B.S.; Pendleton, K.; Zeng, Y.; Lu, Y.; Zhang, Q.; Du, Y.; Gilbert, B.R.; et al. Frequent mutation of the PI3K pathway in head and neck cancer defines predictive biomarkers. Cancer Discov. 2013, 3, 761–769. [Google Scholar] [CrossRef]
- Yuan, T.L.; Cantley, L.C. PI3K pathway alterations in cancer: Variations on a theme. Oncogene 2008, 27, 5497–5510. [Google Scholar] [CrossRef]
- Preissner, S.H.; Nahles, S.; Preissner, S.; Heiland, M.; Koerdt, S. Influence of sex on survival rates of HPV-positive oropharyngeal cancers. Front. Oncol. 2022, 12, 917890. [Google Scholar] [CrossRef]
- Hashibe, M.; Brennan, P.; Chuang, S.C.; Boccia, S.; Castellsague, X.; Chen, C.; Curado, M.P.; Dal Maso, L.; Daudt, A.W.; Fabianova, E.; et al. Interaction between tobacco and alcohol use and the risk of head and neck cancer: Pooled analysis in the International Head and Neck Cancer Epidemiology Consortium. Cancer Epidemiol. Biomark. Prev. 2009, 18, 541–550. [Google Scholar] [CrossRef]
- Byrns, M.C.; Duan, L.; Lee, S.H.; Blair, I.A.; Penning, T.M. Aldo-keto reductase 1C3 expression in MCF-7 cells reveals roles in steroid hormone and prostaglandin metabolism that may explain its over-expression in breast cancer. J. Steroid Biochem. Mol. Biol. 2010, 118, 177–187. [Google Scholar] [CrossRef]
- Badmann, S.; Mayr, D.; Schmoeckel, E.; Hester, A.; Buschmann, C.; Beyer, S.; Kolben, T.; Kraus, F.; Chelariu-Raicu, A.; Burges, A.; et al. AKR1C1/2 inhibition by MPA sensitizes platinum resistant ovarian cancer towards carboplatin. Sci. Rep. 2022, 12, 1862. [Google Scholar] [CrossRef] [PubMed]
- Shiiba, M.; Yamagami, H.; Yamamoto, A.; Minakawa, Y.; Okamoto, A.; Kasamatsu, A.; Sakamoto, Y.; Uzawa, K.; Takiguchi, Y.; Tanzawa, H. Mefenamic acid enhances anticancer drug sensitivity via inhibition of aldo-keto reductase 1C enzyme activity. Oncol. Rep. 2017, 37, 2025–2032. [Google Scholar] [CrossRef]
- Byrns, M.C.; Steckelbroeck, S.; Penning, T.M. An indomethacin analogue, N-(4-chlorobenzoyl)-melatonin, is a selective inhibitor of aldo-keto reductase 1C3 (type 2 3alpha-HSD, type 5 17beta-HSD, and prostaglandin F synthase), a potential target for the treatment of hormone dependent and hormone independent malignancies. Biochem. Pharmacol. 2008, 75, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Hagedorn, C.H.; Cullen, B.R. Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA 2004, 10, 1957–1966. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Yousef, A.; Grandis, J.R.; Johnson, D.E. NSAID therapy for PIK3CA-Altered colorectal, breast, and head and neck cancer. Adv. Biol. Regul. 2020, 75, 100653. [Google Scholar] [CrossRef]
- Trabert, B.; Ness, R.B.; Lo-Ciganic, W.H.; Murphy, M.A.; Goode, E.L.; Poole, E.M.; Brinton, L.A.; Webb, P.M.; Nagle, C.M.; Jordan, S.J.; et al. Aspirin, nonaspirin nonsteroidal anti-inflammatory drug, and acetaminophen use and risk of invasive epithelial ovarian cancer: A pooled analysis in the Ovarian Cancer Association Consortium. J. Natl. Cancer Inst. 2014, 106, djt431. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| AKR1C2 Tumor Staining | ||||||||
|---|---|---|---|---|---|---|---|---|
| Total (1) | AKR1C2HIGH (2) | AKR1C2LOW (2) | ||||||
| Clinicopathological Feature | n | % | n | % | n | % | χ2 | |
| Mean age (years) | 51 | 55.125 | 60.162 | |||||
| Sex | ||||||||
| Male | 39 | 76.5 | 21 | 42.9 | 16 | 32.7 | ||
| Female | 12 | 23.5 | 8 | 16.3 | 4 | 8.2 | 0.738 | |
| T classification | ||||||||
| pT1 and pT2 | 23 | 45.1 | 10 | 20.4 | 12 | 24.5 | ||
| pT3 and pT4 | 28 | 54.9 | 19 | 38.8 | 8 | 16.3 | 0.090 | |
| N classification | ||||||||
| pN0 | 13 | 25.5 | 6 | 12.2 | 7 | 14.3 | ||
| pN1–2 (3) | 39 | 74.5 | 23 | 46.9 | 13 | 26.5 | 0.331 | |
| M classification | ||||||||
| pM0 | 49 | 96.1 | 28 | 57.1 | 19 | 38.8 | ||
| pM1 | 2 | 3.9 | 1 | 2.0 | 1 | 2.0 | 1.000 | |
| AJCC classification | ||||||||
| I | 14 | 27.5 | 5 | 10.2 | 8 | 16.3 | ||
| II | 12 | 23.5 | 4 | 8.2 | 8 | 16.3 | ||
| III | 10 | 19.6 | 8 | 16.3 | 2 | 4.1 | ||
| IV | 15 | 29.4 | 12 | 24.5 | 2 | 4.1 | 0.009 | |
| Relapse | ||||||||
| Yes | 23 | 45.1 | 13 | 26.5 | 9 | 18.4 | ||
| No | 28 | 54.9 | 16 | 32.7 | 11 | 22.4 | 1.000 | |
| Death | ||||||||
| Yes | 23 | 47.0 | 13 | 26.5 | 6 | 12.2 | ||
| No | 26 | 53.3 | 16 | 32.7 | 14 | 28.6 | 0.377 | |
| HPV-status | ||||||||
| Negative | 25 | 49.0 | 19 | 38.8 | 5 | 10.2 | ||
| Positive | 26 | 51.0 | 10 | 20.4 | 15 | 30.6 | 0.008 | |
| Smoking | ||||||||
| Yes | 40 | 78.4 | 22 | 45.8 | 16 | 33.3 | ||
| No | 11 | 21.6 | 6 | 12.5 | 4 | 8.3 | 1.000 | |
| Alcohol | ||||||||
| Yes | 25 | 49 | 14 | 28.6 | 11 | 22.4 | ||
| No | 26 | 47 | 15 | 30.6 | 9 | 18.4 | 0.773 | |
| Localization | ||||||||
| Tonsil | 31 | 60.8 | 16 | 32.7 | 15 | 30.6 | ||
| Tongue base | 15 | 29.4 | 8 | 16.3 | 5 | 10.2 | ||
| Soft palate | 5 | 9.8 | 5 | 10.2 | 0 | 0 | 0.122 | |
| NRF2 expression | ||||||||
| Nuclear | 14 | 27.5 | 8 | 16.3 | 6 | 12.2 | ||
| Cytoplasmic | 37 | 72.5 | 21 | 42.9 | 14 | 28.6 | 1.000 | |
| AKR1C1 expression | ||||||||
| AKR1C1 (+) | 15 | 29.4 | 20 | 40.8 | 6 | 12.2 | ||
| AKR1C1 (−) | 36 | 70.6 | 9 | 18.4 | 14 | 28.6 | 1.000 | |
| AKR1C3 expression | ||||||||
| AKR1C3 (+) | 15 | 29.4 | 20 | 40.8 | 6 | 12.2 | ||
| AKR1C3 (−) | 36 | 70.6 | 9 | 18.4 | 14 | 28.6 | 1.000 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziogas, M.; Siefer, O.; Wuerdemann, N.; Balaji, H.; Gross, E.; Drebber, U.; Klussmann, J.P.; Huebbers, C.U. Analysis of Expression and Regulation of AKR1C2 in HPV-Positive and -Negative Oropharyngeal Squamous Cell Carcinoma. Cancers 2024, 16, 2976. https://doi.org/10.3390/cancers16172976
Ziogas M, Siefer O, Wuerdemann N, Balaji H, Gross E, Drebber U, Klussmann JP, Huebbers CU. Analysis of Expression and Regulation of AKR1C2 in HPV-Positive and -Negative Oropharyngeal Squamous Cell Carcinoma. Cancers. 2024; 16(17):2976. https://doi.org/10.3390/cancers16172976
Chicago/Turabian StyleZiogas, Maria, Oliver Siefer, Nora Wuerdemann, Harini Balaji, Elena Gross, Uta Drebber, Jens Peter Klussmann, and Christian U. Huebbers. 2024. "Analysis of Expression and Regulation of AKR1C2 in HPV-Positive and -Negative Oropharyngeal Squamous Cell Carcinoma" Cancers 16, no. 17: 2976. https://doi.org/10.3390/cancers16172976
APA StyleZiogas, M., Siefer, O., Wuerdemann, N., Balaji, H., Gross, E., Drebber, U., Klussmann, J. P., & Huebbers, C. U. (2024). Analysis of Expression and Regulation of AKR1C2 in HPV-Positive and -Negative Oropharyngeal Squamous Cell Carcinoma. Cancers, 16(17), 2976. https://doi.org/10.3390/cancers16172976