
RASSF6; the Putative Tumor Suppressor of the RASSF Family

Abstract
:1. Introduction
2. RASSF6 and the PDZ Domain-Containing Proteins


3. The SARAH Domain-Mediating Interaction
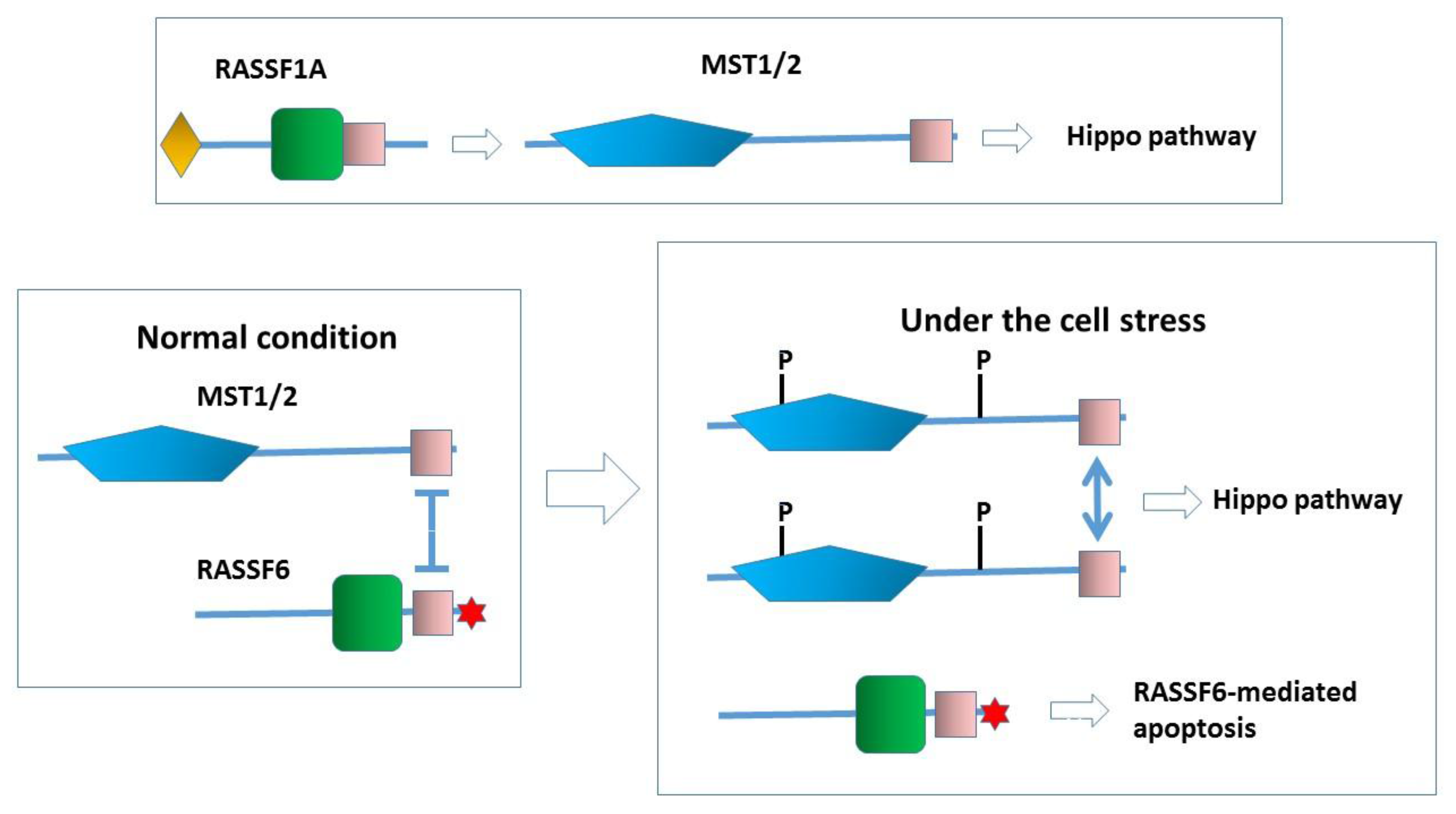
4. RASSF6 and the Hippo Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C-RASSF | Effect on MST Kinases | Ref. |
|---|---|---|
| RASSF1A |
| [20] |
| [20] | |
| [21] | |
| [8] | |
| RASSF2 |
| [22] |
| [23] | |
| [8] | |
| RASSF3 |
| [8] |
| RASSF4 |
| [8] |
| Nore1 |
| [20] |
| [20] | |
| [20] | |
| [24] | |
| RASSF6 |
| [8] |
| [8] | |
| dRASSF |
| [8] |
| [8] |

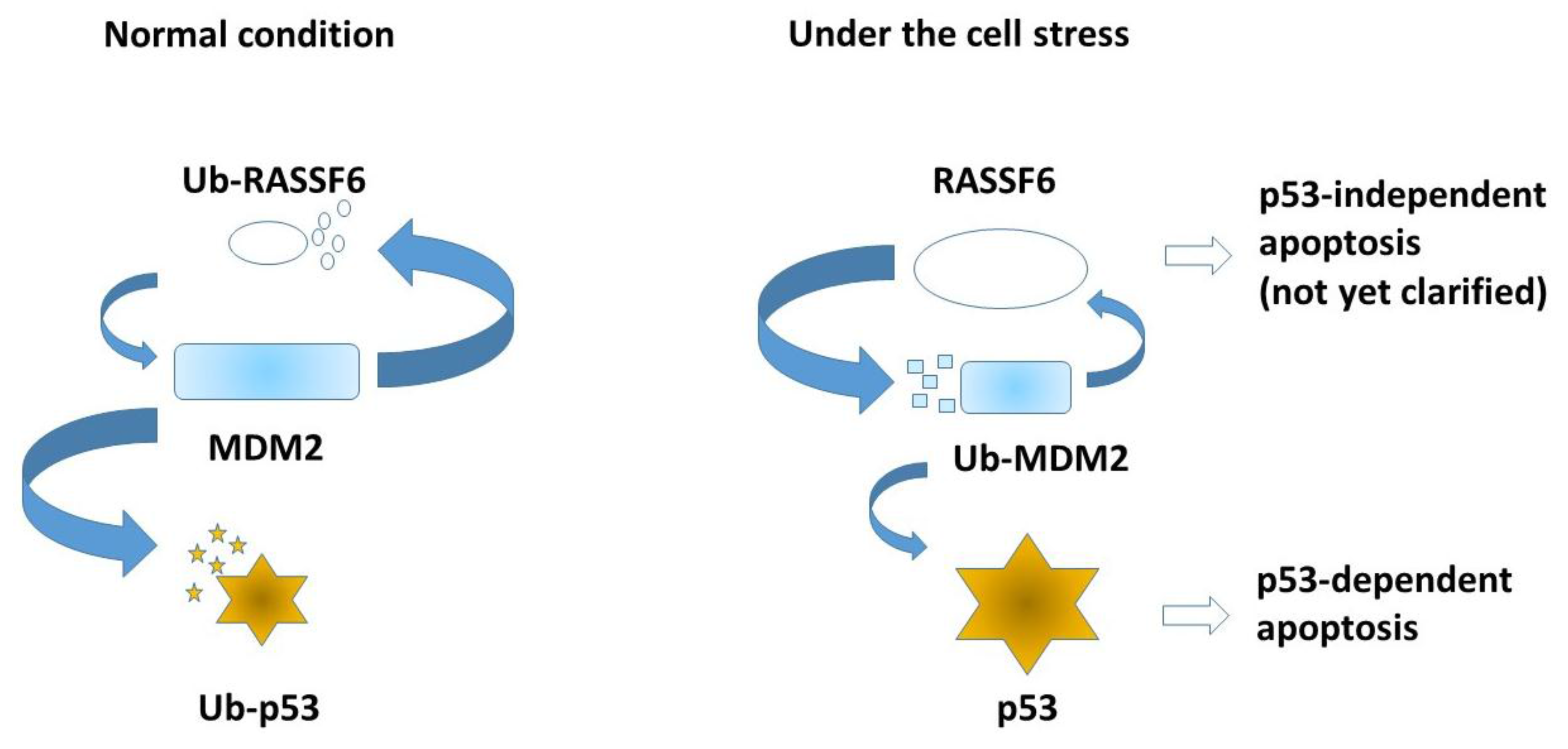
5. RASSF6, MDM2, and p53

6. RASSF6 and Other Pro-Apoptotic Molecules
7. RASSF6 and Ras
8. RASSF6 and NF-κB (Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells) Signaling
9. Conclusions
10. Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Avruch, J.; Xavier, R.; Bardeesy, N.; Zhang, X.F.; Praskova, M.; Zhou, D.; Xia, F. Rassf family of tumor suppressor polypeptides. J. Biol. Chem. 2009, 284, 11001–11005. [Google Scholar] [CrossRef] [PubMed]
- Underhill-Day, N.; Hill, V.; Latif, F. N-terminal RASSF family: RASSF7–RASSF10. Epigenetics 2011, 6, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Volodko, N.; Gordon, M.; Salla, M.; Ghazaleh, H.A.; Baksh, S. RASSF tumor suppressor gene family: Biological functions and regulation. FEBS Lett. 2014, 588, 2671–2684. [Google Scholar] [CrossRef] [PubMed]
- Pan, D. The hippo signaling pathway in development and cancer. Dev. Cell 2010, 19, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Harvey, K.; Tapon, N. The salvador-warts-hippo pathway—An emerging tumour-suppressor network. Nat. Rev. Cancer 2007, 7, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.M.; Pfeifer, G.P.; Dammann, R.H. The RASSF proteins in cancer; From epigenetic silencing to functional characterization. Biochim. Biophys. Acta 2009, 1796, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Hirabayashi, S.; Fujiwara, N.; Mori, H.; Kawata, A.; Iida, J.; Bao, Y.; Sato, Y.; Iida, T.; Sugimura, H.; et al. Ras-association domain family protein 6 induces apoptosis via both caspase-dependent and caspase-independent pathways. Exp. Cell Res. 2007, 313, 1484–1495. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Kawata, A.; Nishikawa, M.; Tateishi, Y.; Yamaguchi, M.; Nakagawa, K.; Hirabayashi, S.; Bao, Y.; Hidaka, S.; Hirata, Y.; et al. Hippo pathway-dependent and -independent roles of RASSF6. Sci. Signal. 2009, 2. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, H.; Kudo, T.; Maimaiti, S.; Ikeda, M.; Maruyama, J.; Nakagawa, K.; Hata, Y. The RASSF6 tumor suppressor protein regulates apoptosis and the cell cycle via MDM2 protein and p53 protein. J. Biol. Chem. 2013, 288, 30320–30329. [Google Scholar] [CrossRef] [PubMed]
- Justice, R.W.; Zilian, O.; Woods, D.F.; Noll, M.; Bryant, P.J. The drosophila tumor suppressor gene warts encodes a homolog of human myotonic dystrophy kinase and is required for the control of cell shape and proliferation. Genes Dev. 1995, 9, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Wang, W.; Zhang, S.; Stewart, R.A.; Yu, W. Identifying tumor suppressors in genetic mosaics: The drosophila lats gene encodes a putative protein kinase. Development 1995, 121, 1053–1063. [Google Scholar] [PubMed]
- Tapon, N.; Harvey, K.F.; Bell, D.W.; Wahrer, D.C.; Schiripo, T.A.; Haber, D.; Hariharan, I.K. Salvador promotes both cell cycle exit and apoptosis in drosophila and is mutated in human cancer cell lines. Cell 2002, 110, 467–478. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Dong, J.; Pan, D. Hippo encodes a Ste-20 family protein kinase that restricts cell proliferation and promotes apoptosis in conjunction with salvador and warts. Cell 2003, 114, 445–456. [Google Scholar] [CrossRef]
- Harvey, K.F.; Pfleger, C.M.; Hariharan, I.K. The drosophila mst ortholog, hippo, restricts growth and cell proliferation and promotes apoptosis. Cell 2003, 114, 457–467. [Google Scholar] [CrossRef]
- Pantalacci, S.; Tapon, N.; Léopold, P. The salvador partner hippo promotes apoptosis and cell-cycle exit in drosophila. Nat. Cell Biol. 2003, 5, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Udan, R.S.; Kango-Singh, M.; Nolo, R.; Tao, C.; Halder, G. Hippo promotes proliferation arrest and apoptosis in the salvador/warts pathway. Nat. Cell Biol. 2003, 5, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.C.; Wei, X.; Shimizu, T.; Ramos, E.; Rohrbaugh, M.; Nikolaidis, N.; Ho, L.L.; Li, Y. Control of cell proliferation and apoptosis by mob as tumor suppressor, mats. Cell 2005, 120, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, S.; Barrera, J.; Matthews, K.; Pan, D. The hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating yorkie, the drosophila homolog of yap. Cell 2005, 122, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Polesello, C.; Huelsmann, S.; Brown, N.H.; Tapon, N. The drosophila rassf homolog antagonizes the hippo pathway. Curr. Biol. 2006, 16, 2459–2465. [Google Scholar] [CrossRef] [PubMed]
- Praskova, M.; Khoklatchev, A.; Ortiz-Vega, S.; Avruch, J. Regulation of the MST1 kinase by autophosphorylation, by the growth inhibitory proteins, RASSF1 and NORE1, and by Ras. Biochem. J. 2004, 381, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Matallanas, D.; Romano, D.; Yee, K.; Meissl, K.; Kucerova, L.; Piazzolla, D.; Baccarini, M.; Vass, J.K.; Kolch, W.; O’neill, E. Rassf1a elicits apoptosis through an MST2 pathway directing proapoptotic transcription by the p73 tumor suppressor protein. Mol. Cell 2007, 27, 962–975. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Oh, S.; Oh, H.J.; Lim, D.S. Role of the tumor suppressor RASSF2 in regulation of MST1 kinase activity. Biochem. Biophys. Res. Commun. 2010, 391, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Cooper, W.N.; Hesson, L.B.; Matallanas, D.; Dallol, A.; von Kriegsheim, A.; Ward, R.; Kolch, W.; Latif, F. RASSF2 associates with and stabilizes the proapoptotic kinase MST2. Oncogene 2009, 28, 2988–2998. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Li, S.; Yu, J.; Min, J.; Brautigam, C.A.; Tomchick, D.R.; Pan, D.; Luo, X. Structural basis for autoactivation of human MST2 kinase and its regulation by Rassf5. Structure 2013, 21, 1757–1768. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.P.; Donninger, H.; Vos, M.D.; Eckfeld, K.; Hesson, L.; Gordon, L.; Birrer, M.J.; Latif, F.; Clark, G.J. RASSF6 is a novel member of the RASSF family of tumor suppressors. Oncogene 2007, 26, 6203–6211. [Google Scholar] [CrossRef] [PubMed]
- Song, M.S.; Song, S.J.; Kim, S.Y.; Oh, H.J.; Lim, D.S. The tumour suppressor rassf1a promotes MDM2 self-ubiquitination by disrupting the MDM2-DAXX-HAUSP complex. EMBO J. 2008, 27, 1863–1874. [Google Scholar] [CrossRef] [PubMed]
- Kudo, T.; Ikeda, M.; Nishikawa, M.; Yang, Z.; Ohno, K.; Nakagawa, K.; Hata, Y. The RASSF3 candidate tumor suppressor induces apoptosis and G1-S cell-cycle arrest via p53. Cancer Res. 2012, 72, 2901–2911. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Park, S.J.; Sung, K.S.; Park, J.; Lee, S.B.; Park, S.Y.; Lee, H.J.; Ahn, J.W.; Choi, S.J.; Lee, S.G.; et al. MDM2 associates with Ras effector NORE1 to induce the degradation of oncoprotein HIPK1. EMBO Rep. 2012, 13, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Vos, M.D.; Dallol, A.; Eckfeld, K.; Allen, N.P.; Donninger, H.; Hesson, L.B.; Calvisi, D.; Latif, F.; Clark, G.J. The RASSF1A tumor suppressor activates bax via MOAP-1. J. Biol. Chem. 2006, 281, 4557–4563. [Google Scholar] [CrossRef] [PubMed]
- Foley, C.J.; Freedman, H.; Choo, S.L.; Onyskiw, C.; Fu, N.Y.; Yu, V.C.; Tuszynski, J.; Pratt, J.C.; Baksh, S. Dynamics of RASSF1A/MOAP-1 association with death receptors. Mol. Cell. Biol. 2008, 28, 4520–4535. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.Y.; Zheng, L.S.; Wu, Y.Z.; Peng, L.X.; Cao, Y.; Cao, X.; Xie, P.; Huang, B.J.; Qian, C.N. RASSF6 promotes p21(cip1/waf1)-dependent cell cycle arrest and apoptosis through activation of the jnk/sapk pathway in clear cell renal cell carcinoma. Cell Cycle 2014, 13, 1440–1449. [Google Scholar] [CrossRef] [PubMed]
- Vavvas, D.; Li, X.; Avruch, J.; Zhang, X.F. Identification of NORE1 as a potential Ras effector. J. Biol. Chem. 1998, 273, 5439–5442. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.J.; Flatters, D.; Rodrigues-Lima, F.; Yan, J.; Thalassinos, K.; Katan, M. Comparative analysis of interactions of RASSF1–10. Adv. Biol. Regul. 2013, 53, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Khokhlatchev, A.; Rabizadeh, S.; Xavier, R.; Nedwidek, M.; Chen, T.; Zhang, X.F.; Seed, B.; Avruch, J. Identification of a novel Ras-regulated proapoptotic pathway. Curr. Biol. 2002, 12, 253–265. [Google Scholar] [CrossRef]
- Donninger, H.; Calvisi, D.F.; Barnoud, T.; Clark, J.; Schmidt, M.L.; Vos, M.D.; Clark, G.J. NORE1A is a Ras senescence effector that controls the apoptotic/senescent balance of p53 via HIPK2. J. Cell Biol. 2015, 208, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, H.; Kuroyanagi, H.; Maimaiti, S.; Ikeda, M.; Nakagawa, K.; Hata, Y. Characterization of rsf-1, the caenorhabditis elegans homolog of the Ras-association domain family protein 1. Exp. Cell Res. 2013, 319, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, Y.; Avruch, J.; Zhang, X.F. NORE1 inhibits tumor cell growth independent of Ras or the MST1/2 kinases. Oncogene 2004, 23, 3426–3433. [Google Scholar] [CrossRef] [PubMed]
- Withanage, K.; Nakagawa, K.; Ikeda, M.; Kurihara, H.; Kudo, T.; Yang, Z.; Sakane, A.; Sasaki, T.; Hata, Y. Expression of RASSF6 in kidney and the implication of RASSF6 and the hippo pathway in the sorbitol-induced apoptosis in renal proximal tubular epithelial cells. J. Biochem. 2012, 152, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Sanada, Y.; Kumoto, T.; Suehiro, H.; Nishimura, F.; Kato, N.; Hata, Y.; Sorisky, A.; Yanaka, N. RASSF6 expression in adipocytes is down-regulated by interaction with macrophages. PLoS ONE 2013, 8, e61931. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Tommasi, S.; Lee, D.H.; Dammann, R.; Pfeifer, G.P. Control of microtubule stability by the RASSF1A tumor suppressor. Oncogene 2003, 22, 8125–8136. [Google Scholar] [CrossRef] [PubMed]
- Dallol, A.; Agathanggelou, A.; Fenton, S.L.; Ahmed-Choudhury, J.; Hesson, L.; Vos, M.D.; Clark, G.J.; Downward, J.; Maher, E.R.; Latif, F. RASSF1A interacts with microtubule-associated proteins and modulates microtubule dynamics. Cancer Res. 2004, 64, 4112–4116. [Google Scholar] [CrossRef] [PubMed]
- Vos, M.D.; Martinez, A.; Elam, C.; Dallol, A.; Taylor, B.J.; Latif, F.; Clark, G.J. A role for the RASSF1A tumor suppressor in the regulation of tubulin polymerization and genomic stability. Cancer Res. 2004, 64, 4244–4250. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.; Cheong, H.K.; ul Mushtaq, A.; Kim, H.Y.; Yeo, K.J.; Kim, E.; Lee, W.C.; Hwang, K.Y.; Cheong, C.; Jeon, Y.H. Structural basis of the heterodimerization of the MST and RASSF sarah domains in the hippo signalling pathway. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1944–1953. [Google Scholar] [CrossRef] [PubMed]
- Crose, L.E.; Galindo, K.A.; Kephart, J.G.; Chen, C.; Fitamant, J.; Bardeesy, N.; Bentley, R.C.; Galindo, R.L.; Chi, J.T.; Linardic, C.M. Alveolar rhabdomyosarcoma-associated pax3-foxo1 promotes tumorigenesis via hippo pathway suppression. J. Clin. Investig. 2014, 124, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Hesson, L.B.; Dunwell, T.L.; Cooper, W.N.; Catchpoole, D.; Brini, A.T.; Chiaramonte, R.; Griffiths, M.; Chalmers, A.D.; Maher, E.R.; Latif, F. The novel RASSF6 and RASSF10 candidate tumour suppressor genes are frequently epigenetically inactivated in childhood leukaemias. Mol. Cancer 2009, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezzanotte, J.J.; Hill, V.; Schmidt, M.L.; Shinawi, T.; Tommasi, S.; Krex, D.; Schackert, G.; Pfeifer, G.P.; Latif, F.; Clark, G.J. RASSF6 exhibits promoter hypermethylation in metastatic melanoma and inhibits invasion in melanoma cells. Epigenetics 2014, 9, 1496–1503. [Google Scholar] [CrossRef] [PubMed]
- Djos, A.; Martinsson, T.; Kogner, P.; Carén, H. The RASSF gene family members RASSF5, RASSF6 and RASSF7 show frequent dna methylation in neuroblastoma. Mol. Cancer 2012, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.L.; Li, D.D.; Lin, Q.; Zhou, Y.; Zhou, Q.B.; Zeng, B.; Fu, Z.Q.; Gao, W.C.; Liu, Y.M.; Chen, R.W.; et al. Low RASSF6 expression in pancreatic ductal adenocarcinoma is associated with poor survival. World J. Gastroenterol. 2015, 21, 6621–6630. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Dong, Z.; Guo, Y.; Shen, S.; Guo, X.; Kuang, G.; Yang, Z. Decreased expression and frequent promoter hypermethylation of RASSF2 and RASSF6 correlate with malignant progression and poor prognosis of gastric cardia adenocarcinoma. Mol. Carcinog. 2015. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.Y.; Chen, M.Y.; Hua, Y.J.; Chen, S.; Zheng, L.S.; Cao, X.; Peng, L.X.; Xie, P.; Huang, B.J.; Sun, R.; et al. Downregulation of Ras association domain family member 6 (RASSF6) underlies the treatment resistance of highly metastatic nasopharyngeal carcinoma cells. PLoS ONE 2014, 9, e100843. [Google Scholar] [CrossRef] [PubMed]
- Gnyszka, A.; Jastrzebski, Z.; Flis, S. DNA methyltransferase inhibitors and their emerging role in epigenetic therapy of cancer. Anticancer Res. 2013, 33, 2989–2996. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwasa, H.; Jiang, X.; Hata, Y. RASSF6; the Putative Tumor Suppressor of the RASSF Family. Cancers 2015, 7, 2415-2426. https://doi.org/10.3390/cancers7040899
Iwasa H, Jiang X, Hata Y. RASSF6; the Putative Tumor Suppressor of the RASSF Family. Cancers. 2015; 7(4):2415-2426. https://doi.org/10.3390/cancers7040899
Chicago/Turabian StyleIwasa, Hiroaki, Xinliang Jiang, and Yutaka Hata. 2015. "RASSF6; the Putative Tumor Suppressor of the RASSF Family" Cancers 7, no. 4: 2415-2426. https://doi.org/10.3390/cancers7040899
APA StyleIwasa, H., Jiang, X., & Hata, Y. (2015). RASSF6; the Putative Tumor Suppressor of the RASSF Family. Cancers, 7(4), 2415-2426. https://doi.org/10.3390/cancers7040899