
Effects of Pine Pollen Polysaccharides and Sulfated Polysaccharides on Ulcerative Colitis and Gut Flora in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Polysaccharides
2.2. Monosaccharide Composition
2.3. Preparation of Sulfated Polysaccharides
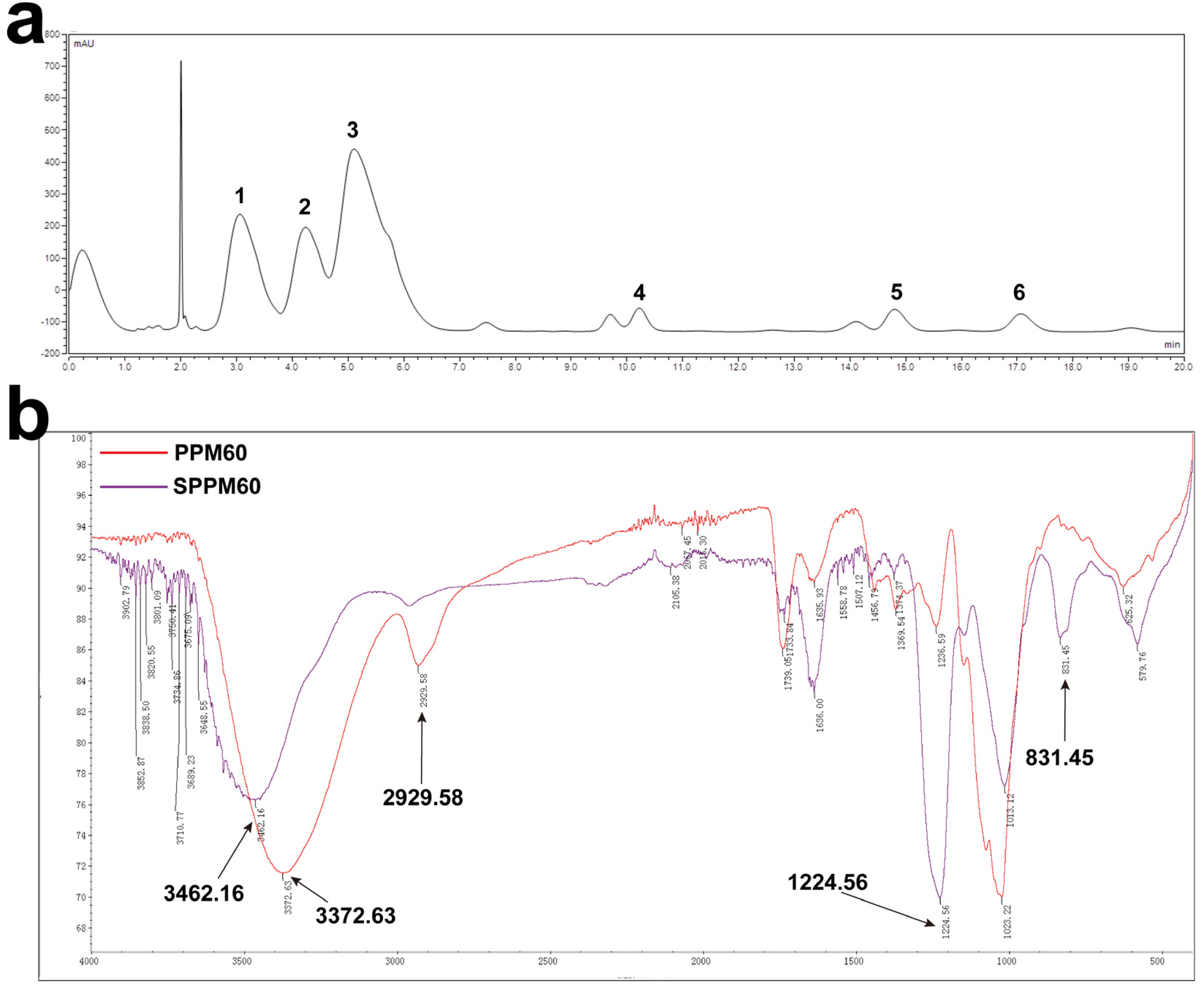
2.4. Infrared Spectroscopy Assay
2.5. Animals and DSS Colitis Model
2.6. Clinical Disease Scores
2.7. Hematoxylin and Eosin Staining
2.8. ELISA
2.9. Sample Preparation and 1H-NMR Spectroscopy
2.10. 16S rDNA Amplicon Sequencing Analysis
2.11. Statistical Analyses
3. Results
3.1. Monosaccharide Composition and Sulfating of PPM60
3.2. Effects of PPM60 and SPPM60 on Disease Progression in UC Mice
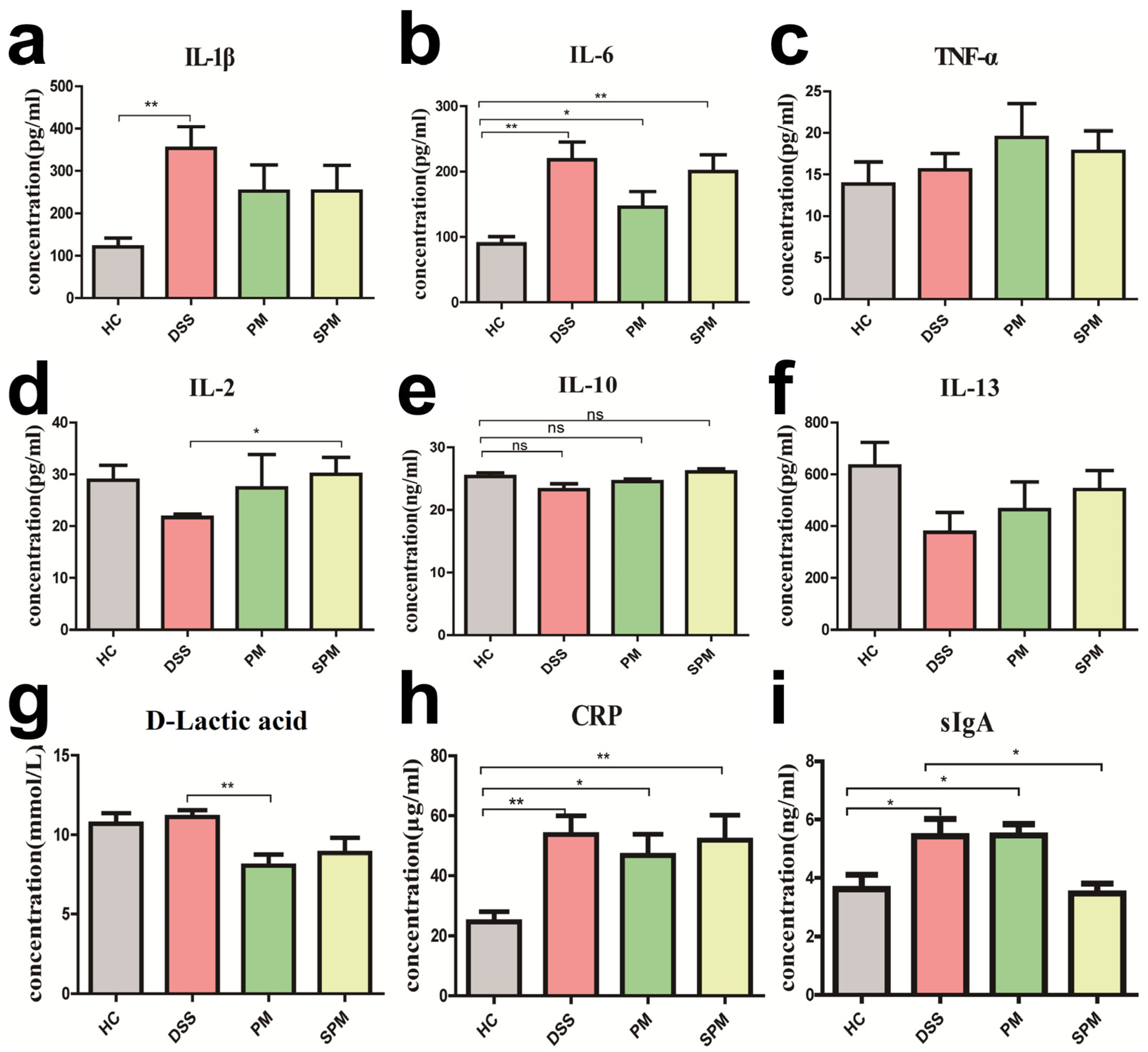
3.3. Effects of PPM60 and SPPM60 on Intestinal Immunity in UC Mice
3.4. Effects of PPM60 and SPPM60 on Serum Metabolomics in UC Mice
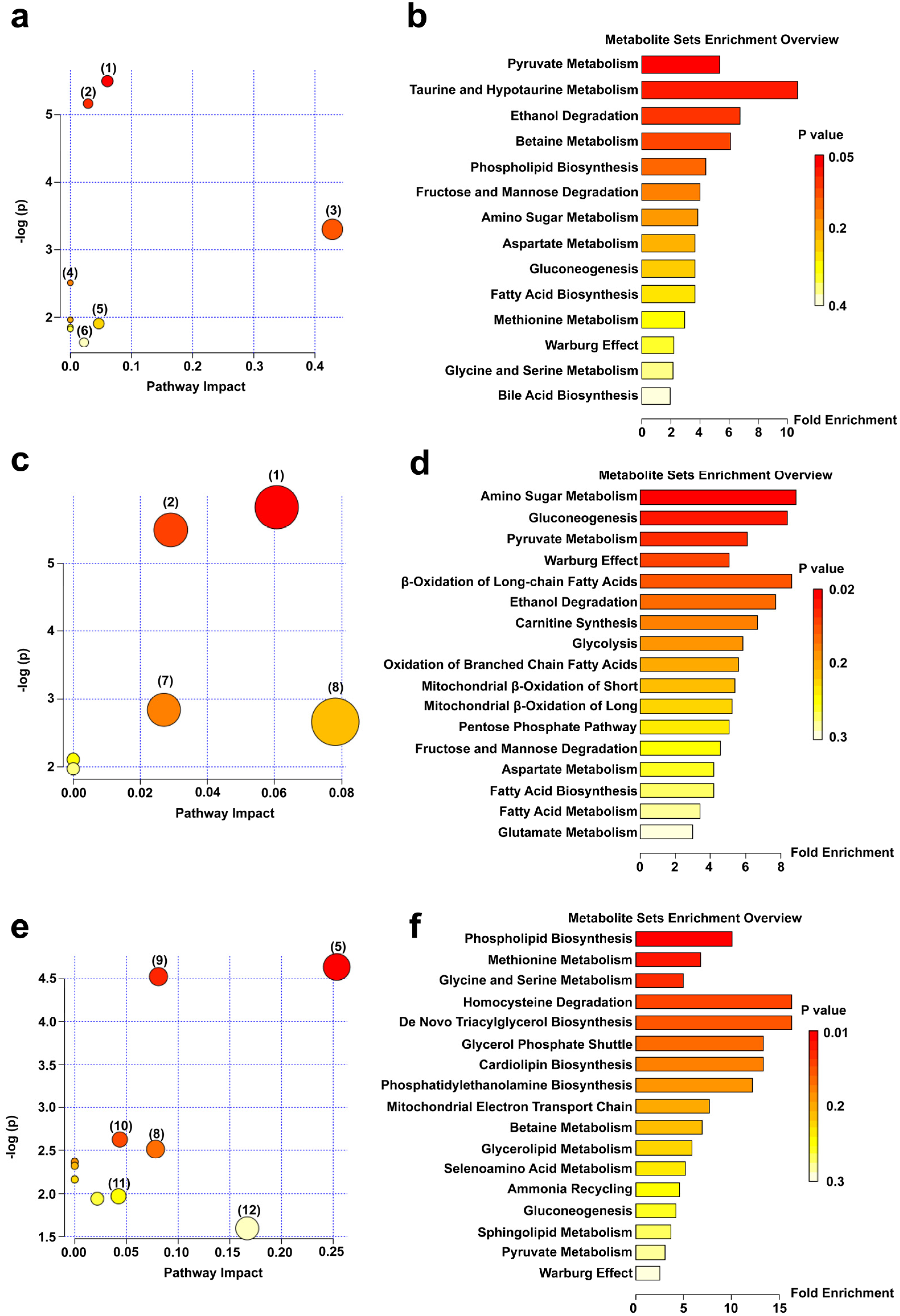
3.4.1. Multivariate Statistical Analysis
3.4.2. Qualitative Analyses of Serum Differentially Expressed Metabolites and Changes in Metabolic Pathways
3.5. Effects of PPM60 and SPPM60 on Gut Flora in UC Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, Y.R.; Rodriguez, J.R. Clinical presentation of Crohn’s, ulcerative colitis, and indeterminate colitis: Symptoms, extraintestinal manifestations, and disease phenotypes. Semin. Pediatr. Surg. 2017, 26, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Ordás, I.; Eckmann, L.; Talamini, M.; Baumgart, D.C.; Sandborn, W.J. Ulcerative colitis. Lancet 2012, 380, 1606–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Li, Y.Y. Inflammatory bowel disease: Pathogenesis. World J. Gastroenterol. 2014, 20, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Burger, D.; Travis, S. Conventional Medical Management of Inflammatory Bowel Disease. Gastroenterology 2011, 140, 1827–1837.e2. [Google Scholar] [CrossRef] [Green Version]
- Seyedian, S.S.; Nokhostin, F.; Malamir, M.D. A review of the diagnosis, prevention, and treatment methods of inflammatory bowel disease. J. Med. Life 2019, 12, 113–122. [Google Scholar] [CrossRef]
- Ullah, S.; Khalil, A.A.; Shaukat, F.; Song, Y. Sources, Extraction and Biomedical Properties of Polysaccharides. Foods 2019, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Zhang, R.; Zhai, J.; Zhu, J.; Yang, F.; Yue, D.; Liu, X.; Lu, C.; Sun, X. Suppression of Th17 Cell Response in the Alleviation of Dextran Sulfate Sodium-Induced Colitis by Ganoderma lucidum Polysaccharides. J. Immunol. Res. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wang, Y.; Huang, X.; Huang, X.Y.; Huang, M.F.; Xu, R.; Yue, H.Y.; Zhou, B.G.; Huang, H.Y.; Sun, Q.M.; et al. Astragalus polysaccharide attenuates rat experimental colitis by inducing regulatory T cells in intestinal Peyer’s patches. World J. Gastroenterol. 2016, 22, 3175–3185. [Google Scholar] [CrossRef]
- Lee, M.Y.; Kim, H.; Shin, K.S. In vitro and in vivo effects of polysaccharides isolated from Korean persimmon vinegar on intestinal immunity. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 867–876. [Google Scholar] [CrossRef]
- Wu, Y.; Jiang, H.; Zhu, E.; Li, J.; Wang, Q.; Zhou, W.; Qin, T.; Wu, X.; Wu, B.; Huang, Y. Hericium erinaceus polysaccharide facilitates restoration of injured intestinal mucosal immunity in Muscovy duck reovirus-infected Muscovy ducklings. Int. J. Biol. Macromol. 2018, 107, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Neurath, M.F.; Wirtz, S. The Intestinal Microbiota in Inflammatory Bowel Disease. ILAR J. 2015, 56, 192–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Hertzberger, R.Y.; Knaus, U.G. Hydrogen peroxide production by lactobacilli promotes epithelial restitution during colitis. Redox Biol. 2018, 16, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Wang, D.; Zheng, W.; Li, X.; Zhang, H.; Zhao, D.; Wang, M. A unique polysaccharide from Hericium erinaceus mycelium ameliorates acetic acid-induced ulcerative colitis rats by modulating the composition of the gut microbiota, short chain fatty acids levels and GPR41/43 respectors. Int. Immunopharmacol. 2019, 71, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; LeLeiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef]
- Frank, D.N.; St Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, M.; Garner, A.; Vlamakis, H.; Xavier, R.J. Microbial genes and pathways in inflammatory bowel disease. Nat. Rev. Microbiol. 2019, 17, 497–511. [Google Scholar] [CrossRef]
- Sun, H.; Ni, X.; Song, X.; Wen, B.; Zhou, Y.; Zou, F.; Yang, M.; Peng, Z.; Zhu, H.; Zeng, Y.; et al. Fermented Yupingfeng polysaccharides enhance immunity by improving the foregut microflora and intestinal barrier in weaning rex rabbits. Appl. Microbiol. Biotechnol. 2016, 100, 8105–8120. [Google Scholar] [CrossRef]
- Yu, Z.; Song, G.; Liu, J.; Wang, J.; Zhang, P.; Chen, K. Beneficial effects of extracellular polysaccharide from Rhizopus nigricans on the intestinal immunity of colorectal cancer mice. Int. J. Biol. Macromol. 2018, 115, 718–726. [Google Scholar] [CrossRef]
- Lu, W.; Yang, Z.; Chen, J.; Wang, D.; Zhang, Y. Recent advances in antiviral activities and potential mechanisms of sulfated polysaccharides. Carbohydr. Polym. 2021, 272, 118526. [Google Scholar] [CrossRef]
- Xiao, H.; Fu, X.; Cao, C.; Li, C.; Chen, C.; Huang, Q. Sulfated modification, characterization, antioxidant and hypoglycemic activities of polysaccharides from Sargassum pallidum. Int. J. Biol. Macromol. 2019, 121, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, Y.; Ouyang, K.; Chen, L.; Zhao, M.; Wang, W. Sulfated Cyclocarya paliurus polysaccharides improve immune function of immunosuppressed mice by modulating intestinal microbiota. Int. J. Biol. Macromol. 2022, 212, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Zheng, X.; Song, J.; Liu, J.; Ren, T.; Zhang, X.; Huang, L.; Wu, M. Radical scavenging activity of sulfated Bupleurum chinense polysaccharides and their effects against oxida-tive stress-induced senescence. Carbohydr. Polym. 2018, 192, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, D.; Hu, Y.; Lu, Y.; Guo, Z.; Kong, X.; Sun, J. Effect of sulfated astragalus polysaccharide on cellular infectivity of infectious bursal disease virus. Int. J. Biol. Macromol. 2008, 42, 166–171. [Google Scholar] [CrossRef]
- Chen, L.; Huang, G. Antioxidant activities of sulfated pumpkin polysaccharides. Int. J. Biol. Macromol. 2018, 126, 743–746. [Google Scholar] [CrossRef]
- Wang, Z.; Xie, J.; Yang, Y.; Zhang, F.; Wang, S.; Wu, T.; Shen, M.; Xie, M. Sulfated Cyclocarya paliurus polysaccharides markedly attenuates inflammation and oxidative damage in lipo-polysaccharide-treated macrophage cells and mice. Sci. Rep. 2017, 7, e40402. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, H.; Wang, Y.; Nie, S.; Li, C.; Xie, M. Sulfated modification of the polysaccharides from Ganoderma atrum and their antioxidant and immuno-modulating activities. Food Chem. 2015, 186, 231–238. [Google Scholar] [CrossRef]
- Chu, H.-L.; Mao, H.; Feng, W.; Liu, J.-W.; Geng, Y. Effects of sulfated polysaccharide from Masson pine (Pinus massoniana) pollen on the proliferation and cell cycle of HepG2 cells. Int. J. Biol. Macromol. 2012, 55, 104–108. [Google Scholar] [CrossRef]
- Geng, Y.; Xing, L.; Sun, M.; Su, F. Immunomodulatory effects of sulfated polysaccharides of pine pollen on mouse macrophages. Int. J. Biol. Macromol. 2016, 91, 846–855. [Google Scholar] [CrossRef]
- Su, F.; Sun, M.; Geng, Y. 1H-NMR Metabolomics Analysis of the Effects of Sulfated Polysaccharides from Masson Pine Pollen in RAW264.7 Macrophage Cells. Molecules 2019, 24, 1841. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, G.; Mahajan, U.B.; Goyal, S.N.; Ojha, S.; Patil, C.R.; Subramanya, S.B. Protective effect of Lagerstroemia speciosa against dextran sulfate sodium induced ulcerative colitis in C57BL/6 mice. Am. J. Transl. Res. 2017, 9, 1792–1800. [Google Scholar] [PubMed]
- Yu, H.; Ge, S.; Huang, R.; Han-Qiao, J. A preliminary study on genetic variation and relationships of Pinus yunnanensis and its closely related species. Acta Bot. Sin. 2000, 42, 107–110. [Google Scholar]
- Mao, G.-X.; Zheng, L.-D.; Cao, Y.-B.; Chen, Z.M.; Lv, Y.-D.; Wang, Y.-Z.; Hu, X.-L.; Wang, G.-F.; Yan, J. Antiaging Effect of Pine Pollen in Human Diploid Fibroblasts and in a Mouse Model Induced by D-Galactose. Oxidative Med. Cell. Longev. 2012, 2012, 750963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Sun, X.; Yu, Z. Effective components and pharmacological function of pine pollen. J. Northeast Univ. 2007, 35, 78–80. [Google Scholar]
- Costa, L.; Fidelis, G.P.; Cordeiro, S.; Oliveira, R.; Sabry, D.; Câmara, R.; Nobre, L.; Costa, M.; Almeida-Lima, J.; Farias, E.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef]
- Tao, Y.; Zhang, L.; Cheung, P.C. Physicochemical properties and antitumor activities of water-soluble native and sulfated hyperbranched mush-room polysaccharides. Carbohydr. Res. 2006, 341, 2261–2269. [Google Scholar] [CrossRef]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Cen, M.; Chen, X.; Chen, H.; Liu, X.; Cao, Q. Correlation between Serological Biomarkers and Disease Activity in Patients with Inflammatory Bowel Disease. BioMed Res. Int. 2019, 2019, 6517549. [Google Scholar] [CrossRef] [Green Version]
- Cornish, J.S.; Wirthgen, E.; Däbritz, J. Biomarkers Predictive of Response to Thiopurine Therapy in Inflammatory Bowel Disease. Front. Med. 2020, 7, 8. [Google Scholar] [CrossRef]
- Espaillat, M.P.; Kew, R.R.; Obeid, L.M. Sphingolipids in neutrophil function and inflammatory responses: Mechanisms and implications for intestinal immunity and inflammation in ulcerative colitis. Adv. Biol. Regul. 2017, 63, 140–155. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.-X.; Shao, M.-J.; Qi, Q.; Xu, Y.-S.; Yang, X.-Q.; Zhu, F.-H.; He, S.-J.; He, P.-L.; Feng, C.-L.; Wu, Y.-W.; et al. Artemisinin analogue SM934 ameliorates DSS-induced mouse ulcerative colitis via suppressing neutrophils and macrophages. Acta Pharmacol. Sin. 2018, 39, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Dai, L.-M.; Shen, H.; Gu, P.-Q.; Zheng, K.; Liu, Y.-J.; Zhang, L.; Cheng, J.-F. Qing Chang Hua Shi granule ameliorate inflammation in experimental rats and cell model of ulcerative colitis through MEK/ERK signaling pathway. Biomed. Pharmacother. 2019, 116, 108967. [Google Scholar] [CrossRef]
- Venn, E.C.; Barnes, A.J.; Hansen, R.J.; Boscan, P.L.; Twedt, D.C.; Sullivan, L.A. Serum D-lactate concentrations in dogs with parvoviral enteritis. J. Vet.-Intern. Med. 2020, 34, 691–699. [Google Scholar] [CrossRef]
- Song, W.-B.; Lv, Y.-H.; Zhang, Z.-S.; Li, Y.-N.; Xiao, L.-P.; Yu, X.-P.; Wang, Y.-Y.; Ji, H.-L.; Ma, L. Soluble intercellular adhesion molecule-1, D-lactate and diamine oxidase in patients with inflammatory bowel disease. World J. Gastroenterol. 2009, 15, 3916–3919. [Google Scholar] [CrossRef] [PubMed]
- Salerno-Goncalves, R.; Safavie, F.; Fasano, A.; Sztein, M.B. Free and complexed-secretory immunoglobulin A triggers distinct intestinal epithelial cell responses. Clin. Exp. Immunol. 2016, 185, 338–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, E.; Parkinson, E.K. Serum metabolomics in animal models and human disease. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 478–483. [Google Scholar] [CrossRef]
- Emwas, A.-H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR Spectroscopy for Metabolomics Research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Xue, J.; Liu, C.; Gao, B.; Chi, L.; Tu, P.; Lu, K.; Ru, H. Serum Metabolomics Identifies Altered Bioenergetics, Signaling Cascades in Parallel with Exposome Markers in Crohn’s Disease. Molecules 2019, 24, 449. [Google Scholar] [CrossRef] [Green Version]
- Dawiskiba, T.; Deja, S.; Mulak, A.; Ząbek, A.; Jawień, E.; Pawełka, D.; Banasik, M.; Mastalerz-Migas, A.; Balcerzak, W.; Kaliszewski, K.; et al. Serum and urine metabolomic fingerprinting in diagnostics of inflammatory bowel diseases. World J. Gastroenterol. 2014, 20, 163–174. [Google Scholar] [CrossRef]
- Kolho, K.-L.; Pessia, A.; Jaakkola, T.; de Vos, W.M.; Velagapudi, V. Faecal and serum metabolomics in paediatric inflammatory bowel disease. J. Crohn’s Colitis 2016, 11, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Weingarden, A.R.; Vaughn, B.P. Intestinal microbiota, fecal microbiota transplantation, and inflammatory bowel disease. Gut Microbes 2017, 8, 238–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Zhu, C.; Quan, Y.; Yang, Z.Y.; Wu, S.; Luo, W.W.; Tan, B.; Wang, X.Y. Relationship between intestinal microbiota and ulcerative colitis: Mechanisms and clinical application of pro-biotics and fecal microbiota transplantation. World J. Gastroenterol. 2018, 24, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Narayanasamy, S.; King, K.; Dennison, A.; Spelman, D.W.; Aung, A.K. Clinical Characteristics and Laboratory Identification of Aerococcus Infections: An Australian Tertiary Centre Perspective. Int. J. Microbiol. 2017, 2017, 5684614. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-J.; Wu, H.; Wu, S.-D.; Lu, N.; Wang, Y.-T.; Liu, H.-N.; Dong, L.; Liu, T.-T.; Shen, X.-Z. Parasutterella, in association with irritable bowel syndrome and intestinal chronic inflammation. J. Gastroenterol. Hepatol. 2018, 33, 1844–1852. [Google Scholar] [CrossRef] [PubMed]
- Horie, M.; Miura, T.; Hirakata, S.; Hosoyama, A.; Sugino, S.; Umeno, A.; Murotomi, K.; Yoshida, Y.; Koike, T. Comparative analysis of the intestinal flora in type 2 diabetes and nondiabetic mice. Exp. Anim. 2017, 66, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Talib, N.; Mohamad, N.E.; Yeap, S.K.; Hussin, Y.; Aziz, M.N.M.; Masarudin, M.J.; Sharifuddin, S.A.; Hui, Y.W.; Ho, C.L.; Alitheen, N.B. Isolation and Characterization of Lactobacillus spp. from Kefir Samples in Malaysia. Molecules 2019, 24, 2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liévin-Le Moal, V.; Servin, A.L. Anti-infective activities of Lactobacillus strains in the human intestinal microbiota: From probiotics to gastrointes-tinal anti-infectious biotherapeutic agents. Clin. Microbiol. Rev. 2014, 27, 167–199. [Google Scholar] [CrossRef] [Green Version]
- Slattery, C.; Cotter, P.D.; O’Toole, P.W. Analysis of Health Benefits Conferred by Lactobacillus Species from Kefir. Nutrients 2019, 11, 1252. [Google Scholar] [CrossRef] [Green Version]
- Geerlings, S.Y.; Kostopoulos, I.; De Vos, W.M.; Belzer, C. Akkermansia muciniphila in the Human Gastrointestinal Tract: When, Where, and How? Microorganisms 2018, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Reunanen, J.; Kainulainen, V.; Huuskonen, L.; Ottman, N.; Belzer, C.; Huhtinen, H.; de Vos, W.M.; Satokari, R. Akkermansia muciniphila Adheres to Enterocytes and Strengthens the Integrity of the Epithelial Cell Layer. Appl. Environ. Microbiol. 2015, 81, 3655–3662. [Google Scholar] [CrossRef] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Håkansson, Å.; Tormobadia, N.; Baridi, A.; Xu, J.; Molin, G.; Hagslätt, M.-L.; Karlsson, C.; Jeppsson, B.; Cilio, C.M.; Ahrné, S. Immunological alteration and changes of gut microbiota after dextran sulfate sodium (DSS) administration in mice. Clin. Exp. Med. 2015, 15, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagalingam, N.A.; Kao, J.Y.; Young, V.B. Microbial ecology of the murine gut associated with the development of dextran sodium sulfate-induced colitis. Inflamm. Bowel Dis. 2011, 17, 917–926. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 C57BL/6 male, 6 weeks C57BL/6 male, 6 weeks | |||
| Day 1 | Day 5 | Day 7 (euthanasia) | |
| HC group | water (free drinking) + water (i.g) | water (free drinking) + water (i.g) | n = 10 |
| DSS group | 3%DSS (free drinking) + water (i.g) | water (free drinking) + water (i.g) | n = 10 |
| PM group | 3%DSS (free drinking) +200 mg/kg PPM60 (i.g) | water (free drinking) + 200 mg/kg PPM60 (i.g) | n = 10 |
| SPM group | 3%DSS (free drinking) +200 mg/kg SPPM60 (i.g) | water (free drinking) + 200 mg/kg SPPM60 (i.g) | n = 10 |
| Name | Shifts | VIP | Trend |
|---|---|---|---|
| N-Acetyl-L-alanine | 1.32 (d), 4.11 (t) | 3.29 | ↑ |
| Acetic acid | 1.25 (t), 3.28 (s), 4.17 (q) | 3.09 | ↑ |
| L-Fucose | 3.88 (dd), 3.79 (m), 3.64 (dd), 3.45 (dd), 1.25 (d) | 3.22 | ↑ |
| Lactic acid | 4.1 (q), 1.32 (d) | 2.42 | ↑ |
| Taurine | 3.25 (t), 3.42 (t) | 2.76 | ↑ |
| Betaine | 3.89 (s), 3.25 (s) | 3.10 | ↑ |
| Acetylcholine | 3.75 (t), 3.23 (s) | 2.38 | ↑ |
| Allose | 3.41 (dd), 3.69 (dd), 3.78 (m), 3.87 (dd) | 2.92 | ↓ |
| Name | Shifts | VIP | Trend |
|---|---|---|---|
| N-Acetyl-L-alanine | 1.32 (d), 4.11 (t) | 2.73 | ↓ |
| Acetic acid | 1.25 (t), 4.17 (q) | 2.43 | ↓ |
| Lactic acid | 4.1 (q), 1.32 (d) | 2.73 | ↓ |
| Fructose-6-phosphate | 3.92 (m), 3.65 (m), 3.55 (m) | 3.26 | ↑ |
| Allose | 3.41 (dd), 3.63 (dd), 3.69 (dd), 3.87 (dd) | 2.92 | ↑ |
| D-xylose | 3.21 (dd), 3.31 (dd), 3.42 (t), 3.51 (dd), 3.63 (m) | 2.38 | ↑ |
| L-carnitine | 3.419 (s), 3.215 (s), 2.425 (s) | 2.38 | ↓ |
| Name | Shifts | VIP | Trend |
|---|---|---|---|
| Betaine | 3.89 (s), 3.25 (s) | 2.99 | ↑ |
| Glyceryl phosphate | 3.67 (dd), 3.82 (m) | 2.24 | ↑ |
| L-serine | 3.832 (dd), 3.958 (m) | 2.09 | ↑ |
| D-xylose | 3.21 (dd), 3.31 (dd), 3.42 (t), 3.51 (dd), 3.63 (m) | 2.34 | ↑ |
| Acetylcholine | 3.75 (t), 3.23 (s), 2.15 (s) | 2.43 | ↑ |
| Lactic acid | 4.1 (q), 1.32 (d) | 3.00 | ↓ |
| Allose | 3.41 (dd), 3.63 (dd), 3.69 (dd), 3.78 (m) | 2.33 | ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Song, X.; Wang, Z.; Li, Z.; Geng, Y. Effects of Pine Pollen Polysaccharides and Sulfated Polysaccharides on Ulcerative Colitis and Gut Flora in Mice. Polymers 2023, 15, 1414. https://doi.org/10.3390/polym15061414
Wang Y, Song X, Wang Z, Li Z, Geng Y. Effects of Pine Pollen Polysaccharides and Sulfated Polysaccharides on Ulcerative Colitis and Gut Flora in Mice. Polymers. 2023; 15(6):1414. https://doi.org/10.3390/polym15061414
Chicago/Turabian StyleWang, Yali, Xiao Song, Zhanjiang Wang, Zhenxiang Li, and Yue Geng. 2023. "Effects of Pine Pollen Polysaccharides and Sulfated Polysaccharides on Ulcerative Colitis and Gut Flora in Mice" Polymers 15, no. 6: 1414. https://doi.org/10.3390/polym15061414