Abstract
Seed dormancy complicates the agricultural use of many legume species. Understanding the genetic and environmental drivers of seed dormancy is necessary for advancing crop improvement for legumes, such as Vicia villosa. In this study, we quantify the magnitude of genetic and environmental effects on physical dormancy among 1488 maternal V. villosa plants from 18 diverse environments. Furthermore, we explore the relationship between physical dormancy and environmental conditions during seed development. Additive genetic variance (h2) accounted for 40% of the variance, while the growing environment explained 28% of the variance in physical dormancy. Maternal lines showed complete variance in physical dormancy, as one line was 100% dormant, and 56 lines were 0% dormant. Distributions of physical dormancy varied widely among seed production environments, with some site-years strongly skewed toward physically dormant seed, while other site-years exhibited little dormant seed. Twenty-three weather variables were associated with environmental and error effects of physical dormancy. High mean and minimum relative humidity, low mean and maximum temperature, and high precipitation weakly grouped with low physical dormancy. Weather variables calculated from fixed time windows approximating seed maturity to seed harvest at each site-year tended to be less predictive than biological seed drying windows calculated based on seed maturity of each maternal line. Overall, individual and cumulative effects of weather variables were poor predictors of physical dormancy. Moderate heritability indicates that breeding programs can select against physical dormancy and improve V. villosa for agricultural use. Marker-based approaches would maximize selection for physical dormancy by reducing the influence of unpredictable environmental effects.
1. Introduction
Legumes provide valuable services to agroecosystems, such as nitrogen supply [1,2], high-quality forage, nutrient capture [3,4,5], soil quality enhancement [5,6], and yield increases to subsequent grain crops [1,7,8]. However, seed dormancy limits agricultural use of leguminous species in agriculture. Seed dormancy reduces stand establishment when farmers plant legumes. As seeds lose dormancy over time, the legume can emerge in subsequent crop rotations and become a problematic weed [9]. Legume crops that exhibit seed dormancy, such as sweetclovers (Melilotus spp), vetches (Vicia spp), clovers (Trifolium spp), medics (Medicago spp), lupines (Lupinus spp), and trefoil (Lotus spp) are often listed as invasive species, and can cause costly ecological damage [10]. Hairy vetch (Vicia villosa Roth) is a particularly problematic species. V. villosa is a legume forage and cover crop that excels in weed suppression [11,12], erosion control [12], nitrogen supply [13,14,15,16,17], and winter survival [18,19], yet is avoided by many farmers because of seed dormancy [20,21].
V. villosa demonstrates combinational seed dormancy, with both physiological and physical dormancy mechanisms preventing immediate germination after seed maturity [22,23]. Physiological dormancy requires a post-harvest ripening process that aligns germination with conducive soil and weather conditions [24]. Physical dormancy (PY) can delay germination for more than one growing season via a water-impermeable seed coat [23,24,25]. Physical dormancy is an evolutionary adaptation that improves survival in risky natural environments (reviewed by Reference [26]). Physical dormancy breaks after accumulated exposure to heat [23] and humidity [22,27].
The dormant seed of V. villosa forms soil seedbanks that can persist and result in emergence during inopportune times of a crop rotation [24,28,29]. The emergence of V. villosa during small grain crop production can reduce crop yield and quality [30,31]. Farmers growing certified seed of small grain crops can also lose certification when seed lots become contaminated with V. villosa, as separating V. villosa seed from small grains is costly and requires specialized equipment. Previous research showed that the scarification of V. villosa seed lots can eliminate most PY; however, seed viability and seedling vigor were negatively impacted [28,29]. Understanding the genetic and environmental drivers of physical dormancy can guide the development of V. villosa varieties with low seed dormancy.
1.1. Genetic Variance in PY
No studies have evaluated the genetic control of physical dormancy in V. villosa. Among studies that evaluated diverse V. villosa germplasm, seed dormancy varied from 0 to 100% [22,27,32,33,34]. In related species, one to three large-effect loci controlled the trait. A single locus was associated with seed dormancy in Lens culinaris Medic. [35], Lupinus angustifolius L. [36], Vigna unguiculata L. [37], Vigna umbellata (Thunb.) Ohwi and Ohashi [38], and Vigna radiata L. [39]. In Pisum sativum L., two to three loci regulated seed dormancy [40]. In Glycine spp., a recessive single point mutation Gmhs1-1 created soft seed by modifying a calcium-based seed coat membrane protein [41]. However, alternative mutations at qHS1 also formed cracks in the seed coat that caused partial imbibition [41,42]. In Vicia sativa L., two genes were implicated, with homozygous recessive aa at the A locus combined with dominance at the B locus associated with soft seed [43].
The stage of maturity when seeds are harvested can impact PY. In various species of the Papilionoideae, PY was only observed in seeds that reached a mature stage of development [44,45,46]. After reaching physiological maturity, seeds must dry below a threshold of water content before an impermeable seed coat develops (reviewed by Reference [9]). Jones [22] found that PY was only observed in V. villosa seeds with a moisture content of less than 14%.
1.2. Environmental Influences on PY
No studies have evaluated how environmental conditions during seed development impact physical dormancy in V. villosa. In related species of the Papilionoideae, relative humidity (RH) and temperature influenced PY. Lower RH during seed drying increased PY in Trifolium repens L. [47,48], Trifolium subterraneum L. [46], Acacia salinga (Labill.) H.L.Wendl. [49], and Macroptilium atropurpureum (DC.) Urb. [50]. The role of the hilum in seed drying is likely responsible for the impact of RH on PY. When RH drops during seed ripening, the hilum opens and releases moisture from the seed. If RH increases, however, the hilum closes and prevents the absorption of ambient moisture [47]. Consequently, the environmental variables most likely associated with PY would be minimum RH during seed drying, and length of time that the hilum remains open (i.e., the cumulative time that RH decreased during seed drying). The minimum RH to which Acacia salinga seed were exposed explained 73% of the variance in PY [49]. Although the duration of exposure to low RH impacted PY in T. subterraneum [46], the influence was minor for A. salinga, accounting for only 4% of the variance in PY [49].
Seeds maturing at higher temperatures have shown increased PY in T. subterraneum [46] and Stylosanthes hamata [51]. High temperature and low RH, described through saturation deficit, cumulatively increased PY in T. subterraneum [46]. Bacliau-Miclau [52] found higher PY in G. max from environments with both high temperature and low RH, but the individual impact of these two variables was not distinguishable in the study.
Previous studies have included few weather variables when evaluating associations with PY. Only one [47,48,51], two [46,49], or three [50] variables were included in analyses. Without measuring or artificially controlling multiple weather variables, studies lack the ability to identify confounding weather variables on PY.
1.3. Hypotheses
We tested the hypothesis that (1) physical dormancy is genetically and environmentally controlled in V. villosa. Analyses quantified the magnitude of genetic and environmental effects on PY among 1488 maternal V. villosa plants from 18 diverse environments. Furthermore, we explored the relationship between PY and environmental conditions during seed development. We hypothesized that the environmental drivers of PY are (2) minimum RH during seed drying and (3) the cumulative time that RH decreased during seed drying.
2. Materials and Methods
2.1. Data Collection
2.1.1. Maternal Line Seed Production Environments
Material for PY analyses were derived from a V. villosa breeding program. A detailed description of nursery design, management, and selection methods are provided in Kissing Kucek et al. [53]. Site-years included a diversity of locations, soil types, planting dates, pod collection dates, and nursery selection intensity (Table 1). Between late August and October, nurseries were planted with diverse accessions, named varieties, and half-sib lines of V. villosa. Prior to flowering the following spring, the best 1.1 to 7.4% of plants in each nursery were selected based on winter survival, fall, and spring vigor. Selections were also based on flowering time for a subset of site-years (17NY, 18NYE, 17NCE, 18NCE, 19NCE). For each selected plant, collaborators recorded maturity in the field using the scale developed by Kalu and Fick [54] when 50% of plants in that nursery had flowers. Some sites collected an additional Kalu and Fick [54] maturity measurement when 50% of the lines had pods. Between early June and mid-August following planting, pods developed brown color, a signal of physiological maturity, and peak PY [55]. Collaborators at each site collected around 50 mature pods from each selected maternal plant. If pods appeared to contain few seeds per pod, more pods were collected per plant, to obtain enough seed for germination assays. The subset of half-sibling seed collected from each maternal line was considered a genetic replicate, as seed coats are constituted of maternal tissue. Seeds were collected and analyzed from a total of 1611 maternal plants.

Table 1.
Seed growing environments included in the analysis of physical dormancy. Latitude, soil types, planting, and pod collection dates include a broad gradient.
2.1.2. Physical Dormancy Assays
Pods were stored in paper bags under low humidity (<35% RH) conditions at 20 °C for five months to overcome the physiological dormancy period for V. villosa, which tends to be less than three months [24]. In some site-years, pods were frozen for three days (17MD, 17NY, 17WI, 18MN, 18NYR, 18NYE) prior to storage to kill Bruchus brachialis larvae. To accommodate a separate study of dehiscence in 2018 and 2019 [56], pods were dried to stable moisture at 30 °C for 24h to 72h. Pods were hand-threshed between rubber mats to prevent any mechanical scarification of the seed coat.
Maternal lines were evaluated for PY using germination assays [57]. Each maternal line contributed 25 seeds per replicate, for a total of four replicates in 2017 and three replicates in 2018 and 2019. Some maternal lines contributed fewer replicates, due to a lack of quality seed. Seeds that were clearly non-viable were removed prior to assays, including seeds that were eaten by B. brachialis larvae, split, shriveled, moldy, or easily crushed when squeezed by beading forceps. Green seeds were avoided in assays, as Samarah [58,59] concluded that green seeds are not physiologically mature.
Seeds were placed in Petri dishes on germination paper saturated with distilled water and placed in sealed plastic bags with a wet paper towel into a germination chamber, following best practices identified by Modisa [34]. Samples resided in a growth chamber set at 12 h at 20 °C and 12h at 15 °C, optimal temperatures for germination of V. villosa [34,60,61]. After 7d, samples were removed, and seeds in the following categories were counted: Imbibed with >1mm radicle, imbibed without radicle, and unimbibed. A noticeable increase in seed size and change in color was evident after the imbibition of water [22,23]. Seeds that did not appear to have imbibed water were squeezed with beading forceps. If the seed did not crush under pressure from the forceps, the seed coat was considered unimbibed. Unimbibed seeds were manually scarified with a razor blade to break the seed coat [22]. The hilum was avoided during cutting. Unimbibed scarified seeds reentered the germination chamber for another 7d, and were reevaluated for the three germination classes. Any seeds that remained unimbibed after 14d were determined to be inadequately scarified, and subsequently re-scarified, placed in the germination chamber for an additional 7d, and then evaluated for imbimbition with >1mm radicle or imbibition without radicle.
A seven-day germination assay was chosen as a high-throughput method for analyzing a large number of samples in the study. Calibration experiments showed that viability increased by 6.3% between 7d and 10d, and 9.0% from 7d and 14d. Consequently, the short duration of our germination assay slightly underestimated the percent viable seed and slightly overestimated the percent PY.
Final data is available [62].
2.1.3. Weather Data Aggregation
Weather data were loaded for latitude and longitude of each site-year (Table 1) on an hourly basis. Rainfall data were compiled from MRMS interpolation estimates provided by NOAA at a 0.01° accuracy, (https://www.nssl.noaa.gov/projects/mrms/). All other weather variables were compiled using NLDAS-2 interpolation estimates provided by NASA at a 0.125° accuracy (https://ldas.gsfc.nasa.gov/nldas/v2/forcing).
2.2. Data Analysis
Each seed from a maternal line received a binomial rating for PY: “0” if the seed imbibed water and produced a radicle without scarification, or “1” if the seed did not imbibe water and produced a radicle after scarification. Non-viable seed, or seed that imbibed water, but did not produce a radicle, were removed from the analysis. Maternal lines that did not have at least ten viable seeds observed were removed from the analysis.
Log-likelihood of PY from individual seed observations of each maternal plant can be described as
where Yijl(k) is the log-likelihood physical dormancy (0,1) of a seed for the maternal plant i in site-year j of technical replicate k, nested within year l; B0 is the intercept of log odds, β1 and xi1 are the partial slope and random variable associated with maternal plant i, β2 and xj2 are the partial slope and random variable associated with site-year j, β3 and xk(l)3 are the partial slope and random variable associated with technical replicate k, nested within year l (Equation (1)). The technical replicate was nested within a year because maternal lines evaluated in each year were unique.
Yijl(k) = β0 + β1xi1 + β2xj2 + β3xl(k)3
2.2.1. Estimation of Genetic and Environmental Effects on PY
A Generalized Linear Mixed animal model [63,64] evaluated log odds of physical dormancy (y) based on intercept log odds (β), random effects (u: maternal plant, site-year, and technical replicate nested within the year), and corresponding design matrices (X, Z) (Equation (2)):
y = Xβ + Zu.
As our samples were a representative subset of V. villosa genetic diversity and seed growing environments, effects were considered random. Due to the imbalance of pedigrees across environments, we were unable to include an interaction effect of genotype by environment (G*E) in the model. Consequently, the influence of G*E was absorbed into the residual error.
An inverse pedigree matrix was calculated using ASReml (v.4) [65], following Reference [66], and was included in the design matrices of Equation (2). The pedigree matrix only included maternal line information, as fathers were unknown in the polycross breeding design. Narrow-sense heritability (h2) was calculated as the variance of the maternal plant divided by the total variance.
The environmental variance was calculated as the variance of site-year divided by total variance. Analyses were completed using ASReml (v.4) [65] in R (version 3.6.0) [67].
2.2.2. Environmental Variables Influencing PY
We sought to understand the impact of weather variables on PY. Our analyses assumed that environment and G*E were the only aspects of PY influenced by weather variables. A model was generated in ASReml-R (v.4) [65] equal to Equation (2) above, but excluding the effect of site-year. Residual values extracted from this model included the effects of the environment, G*E, and residual error on PY. The residual values did not include additive genetic and technical replicate effects, which were assumed to not be influenced by weather variables. Analyses investigated relationships between the residual values and weather variables.
A priori knowledge suggested that humidity and temperature during seed drying regulate PY (see Section 1.2). Since previous studies analyzed only one to three weather variables for associations with PY, we included additional weather variables in our analysis. Eight weather metrics measured temperature, growing degree days above 4 °C (GDD), RH, precipitation, wind speed, longwave radiation, and shortwave radiation. Growing degree days above 4 °C [68] were calculated using maximum and minimum daily temperatures, according to Baskerville and Ermin [69]. Aggregate statistics summarized mean, minimum, maximum, and cumulative values of weather metrics. Time windows used to calculate aggregate statistics for weather metrics included (1) biological windows based on seed drying times of individual maternal plants in the field [70] and (2) fixed windows of seed drying for each site-year. A total of 26 aggregate weather variables were calculated for unique combinations of weather metrics, aggregate statistics, and biological or fixed time windows.
Start dates for biological seed drying windows were extrapolated from individual plant maturity ratings recorded when 50% of lines had begun to flower or when 50% of lines had pods. A linear extrapolation function determined the GDD necessary to reach physiological seed maturity from earlier maturity stages (Appendix A). Growing degree days above 4 °C were calculated starting from the planting date on October 1st, 2018. The function was based on data from 14 V. villosa lines grown in 2019 at Corvallis, OR. Data were collected in four replicate plots per line, each plot measuring 4.65 m2. Every two weeks, researchers rated the maturity of each line on a scale modified from Kalu and Fick [54] (Figure A1). A linear model explained 97.45% of the variance in GDD among V. villosa lines, where y is the GDD above 4 °C, and x is the Kalu and Fick maturity stage (Equation (3)):
y = 108.507 x + 462.167
End dates for seed drying windows were the date when pods were harvested from the field. Some site-years recorded different pod harvest timings for individual plants (Table 1), while other site-years only recorded one harvest date. Biological windows of seed drying for individual maternal plants were compared to fixed windows at each site-year. Fixed windows for each site-year included 30 days prior to the final harvest day at that site-year.
We used Least Absolute Shrinkage and Selection Operator (Lasso) to identify weather variables contributing to PY, handle multicollinearity of weather variables, and to penalize the number of variables included in the model through package ‘glmnet’ (v. 4.0-2) [71]. All weather variables were centered and scaled prior to analysis. A 100-fold training set identified a penalty term, λ, that minimized cross-validation error. When the Lasso model reduced the regression coefficient of a weather variable to zero, that variable was considered to not contribute to PY. The magnitude of coefficients for each weather variable were plotted using ‘ggplot2’ (v. 3.3.2) [72]. Relationships between residuals and each weather variable with non-zero coefficients were visualized using smoothing splines of nonparametric generalized additive models generated plotted using ‘ggplot2’ (v. 3.3.2) [72]. All analyses were conducted in R (version 3.6.0) [67].
The total impact of weather variables with non-zero coefficients on PY was analyzed through percent deviance. The deviance of the selected Lasso model (D) was calculated as two times the difference between log-likelihoods of a saturated perfect fit model and the Lasso model, including the weather variables with non-zero regression coefficients. Null deviance (D0) was calculated as two times the difference between log-likelihoods of a fully saturated model and a null model, only including an intercept. The percent of deviance evaluated the quality of the model using Equation (4):
% Deviance = 1 – D/D0
3. Results
3.1. Genetic Variance and Heritability of PY
Of 1611 maternal lines, 1488 had at least ten viable seeds for analysis. Maternal lines showed complete variance in physical dormancy among site-years included in the study (Figure 1). One maternal line exhibited 100% PY, while 56 maternal lines exhibited no physical dormancy. The additive genetic effect, h2, accounted for 40.31% of the variance of log odds of PY (Table 2) in the animal model (Equation (2)).
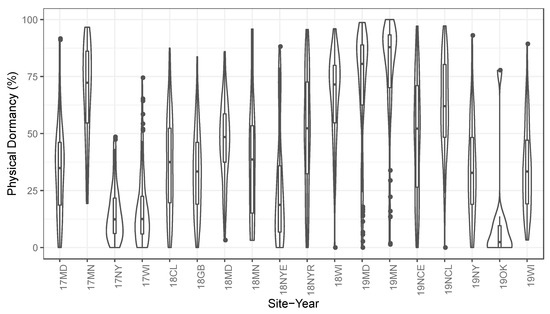
Figure 1.
Violin plots show the distribution of physical dormancy for maternal lines grown in each site-year. Boxplots embedded within the violin plots indicate means, 25th and 75th percentiles, and outliers for each site-year. The complete variance was observed among site-years and maternal lines, with extreme values, including 0 and 100% physical dormancy.
Table 2.
Additive genetic variance, environmental variance, technical replicate variance, and residual variance of physical dormancy using an animal model.
3.2. Environmental Effect
3.2.1. Environmental Variance in PY
Distributions of physical dormancy varied strongly among seed production environments, with some site-years (19MD, 19MN) strongly skewed toward physically dormant seed, and other site-years (17NY, 17WI, 19OK) exhibiting little dormant seed (Figure 1). In the animal model (Equation (2)), the growing environment explained 27.95% of the variance in PY (Table 2). Growing environment explained less variance in PY than the genetic effect, but more variance than technical replicate or residual error (including G*E). Locations did not show consistent PY among years. Each environment that was measured in multiple years had vastly different PY means and distributions (Figure 1)
Residual values measured the influence of the environment, G*E, and error on PY (Figure 2). For each observation, residual values explained deviance in log-likelihood of physical dormancy after removing the pedigree matrix and technical replicate. Negative residuals indicated observations with high PY values that deviated from genetic or technical replicate effects (Figure 2). Conversely, positive residuals indicated observations with low PY values that deviated from genetic or technical replicate effects (Figure 2).
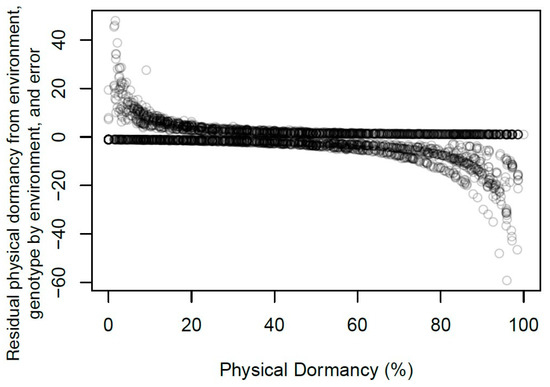
Figure 2.
Relationship between percent physical dormancy and residuals, including environment, genotype by environment, and error. Residuals indicate the deviance in log-likelihood of physical dormancy from genetic and technical replicate effects. Negative residuals indicate observations with high physical dormancy relative to genetic and technical replicate effects. Positive residuals indicate observations with low physical dormancy relative to genetic and technical replicate effects.
3.2.2. Weather Variables Associated with PY
Twenty-three weather variables showed association with PY environment, G*E, and error residuals (Figure 3). However, all 23 weather variables with non-zero coefficients together explained only 12.17% of the deviance in residuals. Weather variables that were most to least associated with PY were mean precipitation, mean RH, mean temperature, minimum RH, cumulative precipitation, maximum temperature, mean wind speed, cumulative hours below 50% RH, mean shortwave radiation, cumulative GDD above 4 °C, cumulative longwave radiation, and mean longwave radiation (Figure 3). Weather variables calculated from fixed time windows prior to seed harvest tended to be less predictive than the biological seed drying windows calculated for each maternal line, especially for measures of RH. Regression coefficients for biological seed drying windows were further from zero, and therefore, more important to model fit, compared with fixed time windows for mean RH, mean temperature, cumulative precipitation, minimum RH, mean shortwave radiation, and mean longwave radiation (Figure 3).
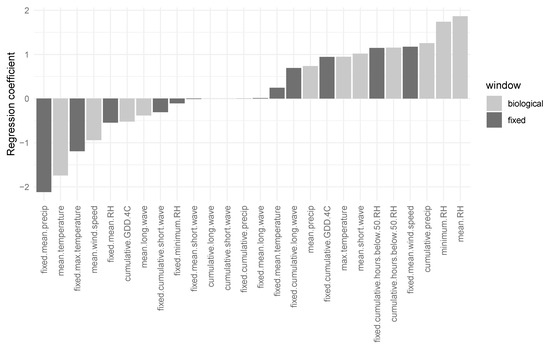
Figure 3.
Relationships of weather variables with the environment, genotype by environment interaction, and error residuals on physical dormancy. A Least Absolute Shrinkage and Selection Operator fitted 26 weather variables to residuals. A 100-fold training set identified a prior distribution for the Lasso, λ, that minimized cross-validation error. Aggregate statistics from hourly weather data were calculated as fixed time windows at 30 days prior to the final pod harvest at a site-year (‘fixed’) and biological time windows extrapolated as the date of seed physiological maturity to the date of pod harvest for individual maternal plants (‘biological’). Bars show the magnitude of the contribution of weather variable to environment, genotype by environment, and error residuals.
Smoothing splines did not show strong relationships between weather variables and PY residuals. The most important six weather variables in the Lasso model (Figure 3; fixed mean precipitation, mean RH, mean temperature, minimum RH, cumulative precipitation, and fixed maximum temperature) showed weak visual relationships with PY (Figure 4). Seeds that dried in conditions with high mean and minimum RH, low mean and maximum temperature, and high precipitation weakly grouped with lower PY (Figure 4).
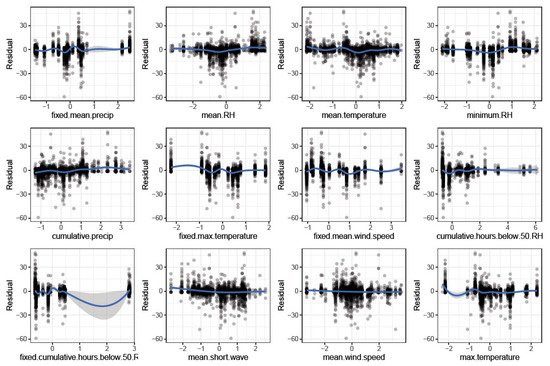
Figure 4.
Relationships between weather variables and percent physical dormancy residuals of the environment, genotype by environment, and error. Negative residuals indicate observations with high physical dormancy, while positive residuals indicate observations with low physical dormancy relative to genetic and technical replicate effects. Weather variables on the x-axes were centered and scaled prior to analysis. Blue lines show nonparametric smoothing splines created using generalized additive models. Shaded blue areas around blue lines show the standard error of the spline fit.
4. Discussion
V. villosa lines often contain greater diversity within lines than among lines [53]. Large datasets are, therefore, necessary to accurately estimate genetic and environmental effects. The high number and broad diversity of genotypes (1488) and environments (18) in this study provide a robust estimate of genotype and environmental effects on PY.
The complete variance and moderate heritability of PY indicate the potential to select against PY in V. villosa [20]. Moderate heritability indicates that the trait is not likely to be highly quantitative, similar to related species [35,36,37,38,39,40,41,42,43]. If PY is controlled by a small number of genes, marker-based selection should be an effective method to select against PY. The broad genetic diversity of pedigrees included in this study improves the accuracy of our heritability estimate. However, there could be a potential bias of heritability, due to indirect selection in the breeding program. Although we did not select for PY directly, plants were only sampled if they germinated the first year after harvest in 2018 and 2019. Mechanical threshing of material would have broken some physically dormant seed coats, but lines with PY may have been less likely to germinate in 2018 and 2019. Since many lines demonstrated high PY in all years of the breeding program, there is little evidence for this bias in the dataset.
PY means and distributions were highly variable among locations and within locations in different years (Figure 1). The narrow window of seed drying likely increases the impact of year effects on PY. Each location included in the study did not have consistent relative humidity, precipitation, temperatures, wind speed, nor shortwave radiation during seed drying windows among years. Within a single site-year, genotypes with divergent maturity timings also experienced different environmental conditions during seed drying windows. To obtain variance in PY, accurately identify molecular markers associated with PY, and make meaningful genetic improvements, breeding programs should include a diversity of environments.
Weather variables were weakly related to environmental effects of PY. Lasso models indicated the influence of weather variables previously associated with PY, including RH and temperature. Validating the first part of our hypothesis, minimum RH during seed drying was associated with PY. However, the second part of our hypothesis was less supported. Similar to [49], cumulative time that seeds were exposed to low RH conditions impacted PY to a lesser degree than other weather variables (Figure 3), and the relationship with PY was not clear (Figure 4). In this data set, high mean and minimum RH, low mean and maximum temperature, and high precipitation are very weakly grouped with low PY. Weather variables calculated from specific seed drying windows for individual maternal lines tended to be more related to PY than fixed time windows, especially for the important metrics of temperature and RH. Future studies should identify biologically specific seed drying windows prior to assessing correlations with PY. Norman [73] found no correlation between mean annual rainfall of a collection site and PY in various species of Trifolium, likely because the annual time window over which the weather variable was calculated was too broad to represent the specific environment of seed drying.
Weather variables individually and cumulatively explained a small portion of the environmental effects on PY. Several sources of variability that were not included in our models could have reduced the influence of weather variables on PY. First, the indeterminate nature of vetch altered seed drying windows experienced by different seeds of a maternal line. The time window of seed drying was a mean estimate for each plant. This study did not track the developmental timeline of each individual seed analyzed for PY. Some seeds collected on a plant could have dried for longer or shorter time periods than the plant mean estimate, due to flowering indeterminacy. Second, microclimatic differences among seeds on a single plant could have impacted PY. Microclimatic weather variables experienced by a seed depending on the location of a pod on a plant. Accurately measuring RH can be especially challenging, as microclimates can be different in the upper and lower parts of the plant [48]. Consequently, accurate measurements of RH would require a demanding sampling at the site of each individual ripening pod. Third, variations in soil could have influenced hard seed. Fourth, post-harvest conditions could have impacted PY. Seeds experienced similar storage conditions once shipped to a common storage location, typically within a month after harvest. Prior to shipping, some site-years froze seed for three days (see Section 2.1.2). A model incorporating an effect for seed freezing did not show improved fit over Equation (2). Beyond the recorded information of freezing, the site-year variability in seed storage conditions prior to shipping is unknown and could have added unaccounted variability to our response of PY. Fifth, the record-keeping detail of pod harvest dates was not consistent among studied site-years. The accuracy of the weather variable analysis depended on known seed drying time in the field. Twelve site-years recorded variable harvest timings according to maturity timing of maternal lines. However, three site-years did not vary pod harvest based on maternal lines maturity timings, and three site-years did not record pod harvest timing (Table 1). Some errors were introduced into the analyses due to la lack of detailed information on pod harvest for three of the 18 site-years (likely due to reduced accuracy of weather variables calculated for biological windows). Potential sources of unaccounted variability in our dataset are common for research and breeding programs. Consequently, weather variables are likely not a reliable way to measure non-genetic effects on PY.
5. Conclusions
Physical dormancy in V. villosa shows moderate heritability and potential for alteration by breeding programs. Environmental effects also impact physical dormancy, with wide variance among locations and within locations in different years. Weather variables poorly predict environment, genotype by environment, and residual effects of PY. Due to the low predictability of environmental effects, and large contributions of genetic effects, breeding programs would make most progress using marker-based approaches to select for or against physical dormancy.
Author Contributions
Conceptualization, L.K.K. and H.R.; methodology, L.K.K., H.R., N.P.W., M.D.A., R.J.H.; software, L.K.K.; validation, L.K.K., H.R.; formal analysis, L.K.K.; investigation, L.K.K., H.R., N.P.W., N.J.E., M.D.A., S.S.E., S.W.; resources, H.R., S.B.M., C.R.-H., N.J.E., M.R.R., M.D.A., R.J.H.; data curation, L.K.K., S.W., N.P.W.; writing—original draft preparation, L.K.K.; writing—review and editing, L.K.K., H.R., M.D.A., S.S.E., R.J.H., M.R.R., S.W., N.J.E., N.P.W., S.B.M.; visualization, L.K.K., H.R., M.D.A.; supervision, L.K.K., H.R., S.S.E.; project administration, L.K.K., S.S.E., H.R., C.R.-H., S.B.M.; funding acquisition, L.K.K., H.R., C.R.-H., S.B.M., M.R.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Institute for Food and Agriculture grant numbers 2015-51300-24192, 2018-51300-28424, and 2018-67013-27570.
Acknowledgments
We would like to thank Rick Hitchcock and Aurélie Poncet for compilation of NOAA and NASA weather data. Field support was provided by Rebecca Heidelberger, Megan Poskaitis, and Allen Burke. We thank David Podoll, Steve Groff, Jeoffry Drillaud, and the Institute for Field and Vegetable Crops Maksima Gorkog for donation of germplasm.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A

Figure A1.
Maturity development time series for fourteen V. villosa lines grown at the USDA-ARS Forage Seed and Cereal Research Unit in Corvallis, Oregon in 2019. Data were collected in four replicate plots per line, each plot measuring 4.65 m2. Every two weeks, researchers rated the maturity of each line on a scale modified from Kalu and Fick [54]. A linear extrapolation function of the data determined the growing degree days above 4 °C (GDD) necessary to reach physiological seed maturity. In a linear model (Equation (3)), maturity stage explained 97.45% of the variance in GDD among V. villosa lines.
Figure A1.
Maturity development time series for fourteen V. villosa lines grown at the USDA-ARS Forage Seed and Cereal Research Unit in Corvallis, Oregon in 2019. Data were collected in four replicate plots per line, each plot measuring 4.65 m2. Every two weeks, researchers rated the maturity of each line on a scale modified from Kalu and Fick [54]. A linear extrapolation function of the data determined the growing degree days above 4 °C (GDD) necessary to reach physiological seed maturity. In a linear model (Equation (3)), maturity stage explained 97.45% of the variance in GDD among V. villosa lines.

References
- Frye, W.W.; Blevins, R.L.; Smith, M.S.; Corak, S.J.; Varco, J.J. Role of Annual Legume Cover Crops in Efficient Use of Water and Nitrogen. Crop. Strat. Effic. Use Water Nitrogen 1988, 51, 129–154. [Google Scholar] [CrossRef]
- Dabney, S.M.; Delgado, J.A.; Reeves, D.W. Using winter cover crops to improve soil and water quality. Commun. Soil Sci. Plant Anal. 2001, 32, 1221–1250. [Google Scholar] [CrossRef]
- Drinkwater, L.E.; Wagoner, P.; Sarrantonio, M. Legume-based cropping systems have reduced carbon and nitrogen losses. Nature 1998, 396, 262–265. [Google Scholar] [CrossRef]
- Langdale, G.W.; Blevins, R.L.; Karlen, D.L.; McCool, D.K.; Nearing, M.A.; Skidmore, E.L.; Thomas, A.W.; Tyler, D.D.; Williams, J.R. Cover crop effects on soil erosion by wind and water. In Cover Crops for Clean Water; Hargrove, W.L., Ed.; Soil & Water Conservation Society: Ankeny, IA, USA, 1991; pp. 15–22. [Google Scholar]
- Kaspar, T.C.; Singer, J.W. The Use of Cover Crops to Manage Soil. Soil Manag. Build. A Stable Base Agric. 2015, 321–337. [Google Scholar] [CrossRef]
- Tiemann, L.K.; Grandy, A.S.; Atkinson, E.E.; Marin-Spiotta, E.; McDaniel, M.D. Crop rotational diversity enhances belowground communities and functions in an agroecosystem. Ecol. Lett. 2015, 18, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Torbert, H.A.; Reeves, D.W.; Mulvaney, R.L. Winter legume cover crop benefits to corn: Rotation vs. fixed-nitrogen effects. Agron. J. 1996, 88, 527–535. [Google Scholar] [CrossRef]
- Miguez, F.E.; Bollero, G.A. Winter cover crops in Illinois: Evaluation of ecophysiological characteristics of corn. Crop Sci. 2006, 46, 1536–1545. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Germination Ecology of Seeds with Physical Dormancy; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780124166776. [Google Scholar]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Daughtry, C.S. Weed Suppression by Live and Desiccated Hairy Vetch (Vicia villosa). Weed Sci. 1993, 41, 207–212. [Google Scholar] [CrossRef]
- Mutchler, C.K.; McDowell, L.L. Soil loss from cotton with winter cover crops. Trans. Am. Soc. Agric. Eng. 1990, 33, 432–436. [Google Scholar] [CrossRef]
- Sainju, U.M.; Singh, B.P.; Yaffa, S. Soil organic matter and tomato yield following tillage, cover cropping, and nitrogen fertilization. Agron. J. 2002, 94, 594–602. [Google Scholar] [CrossRef]
- Yaffa, S.; Sainju, U.M.; Singh, B.P. Fresh market tomato yield and soil nitrogen as affected by tillage, cover cropping, and nitrogen fertilization. HortScience 2000, 35, 1258–1262. [Google Scholar] [CrossRef]
- Parr, M.; Grossman, J.M.; Reberg-Horton, S.C.; Brinton, C.; Crozier, C. Nitrogen Delivery from Legume Cover Crops in No-Till Organic Corn Production. Agron. J. 2011, 103, 1578–1590. [Google Scholar] [CrossRef]
- Power, J.F.; Zachariassen, J.A. Relative nitrogen utilization by legume cover crop species at three soil temperatures. Agron. J. 1993, 85, 134–140. [Google Scholar] [CrossRef]
- Decker, A.M.; Clark, A.J.; Meisinger, J.J.; Mulford, F.R.; McIntosh, M.S. Legume cover crop contributions to no-tillage corn production. Agron. J. 1994, 86, 126–135. [Google Scholar] [CrossRef]
- Brandsæter, L.O.; Olsmo, A.; Tronsmo, A.M.; Fykse, H. Freezing Resistance of Winter Annual and Biennial Legumes at Different Developmental Stages. Crop Sci. 2002, 42, 437–443. [Google Scholar] [CrossRef]
- Stute, J.K.; Posner, J.L. Legume cover crop options for grain rotations in Wisconsin. Agron. J. 1993, 85, 1128–1132. [Google Scholar] [CrossRef]
- Wayman, S.; Kissing Kucek, L.; Mirsky, S.B.; Ackroyd, V.; Cordeau, S.; Ryan, M.R. Organic and conventional farmers differ in their perspectives on cover crop use and breeding. Renew. Agric. Food Syst. 2016, 1–10. [Google Scholar] [CrossRef]
- CTICS. Annual Report 2016–2017 Cover Crop Survey, Joint publication of the Conservation Technology Information Center, the North Central Region Sustainable Agriculture Research and Education Program, and the American Seed Trade Association; CTICS: Wahington, DC, USA, 2017; pp. 1–46. [Google Scholar] [CrossRef]
- Jones, J.P. A physiological study of dormancy in vetch seed. Cornell Univ. Agric. Exp. Stn. 1928, 120, 1–50. [Google Scholar]
- Renzi, J.P.; Chantre, G.R.; Cantamutto, M.A. Development of a thermal-time model for combinational dormancy release of hairy vetch (Vicia villosa ssp. villosa). Crop Pasture Sci. 2014, 65, 470–478. [Google Scholar] [CrossRef]
- Renzi, J.P.; Chantre, G.R.; Cantamutto, M.A. Vicia villosa ssp. villosa Roth field emergence model in a semiarid agroecosystem. Grass Forage Sci. 2018, 73, 146–158. [Google Scholar] [CrossRef]
- Rolston, P.M. Water Impermeable Dormancy. Bot. Rev. 1978, 44, 365–396. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Biogeographical and Evolutionary Aspects of Seed Dormancy; Academic Press: London, UK, 2014; ISBN 9780124166776. [Google Scholar]
- Renzi, J.P.; Chantre, G.R.; Cantamutto, M.A. Effect of water availability and seed source on physical dormancy break of Vicia villosa ssp. villosa. Seed Sci. Res. 2016, 26, 1–10. [Google Scholar] [CrossRef]
- Mirsky, S.B.; Wallace, J.M.; Curran, W.S.; Crockett, B.C. Hairy vetch seedbank persistence and implications for cover crop management. Agron. J. 2015, 107, 2391–2400. [Google Scholar] [CrossRef]
- Jacobsen, K.L.; Gallagher, R.S.; Burnham, M.; Bradley, B.B.; Larson, Z.M.; Walker, C.W.; Watson, J.E. Mitigation of seed germination impediments in hairy vetch. Agron. J. 2010, 102, 1346–1351. [Google Scholar] [CrossRef]
- Curran, W.S.; Wallace, J.M.; Mirsky, S.; Crockett, B. Effectiveness of Herbicides for Control of Hairy Vetch (Vicia villosa) in Winter Wheat. Weed Technol. 2015, 29, 509–518. [Google Scholar] [CrossRef]
- Keene, C.L.; Curran, W.S.; Wallace, J.M.; Ryan, M.R.; Mirsky, S.B.; Vangessel, M.J.; Barbercheck, M.E. Cover crop termination timing is critical in organic rotational no-till systems. Agron. J. 2017, 109, 272–282. [Google Scholar] [CrossRef]
- Elkins, D.M.; Hoveland, C.S.; Donnelly, E.D. Germination of Vicia Species and Interspecific Lines as Affected by Temperature Cycles. Crop Sci. 1966, 6, 45–48. [Google Scholar] [CrossRef]
- Jannink, J.; Merrick, L.C.; Liebman, M.; Dyck, E.A.; Corson, S. Management and winter hardiness of hairy vetch in Maine. Maine Agric. Exp. Stn. Tech. Bull. 1997, 167, 35. [Google Scholar]
- Modisa, O. Seed Dormancy in Vetch; University of Melbourne: Melbourne, Australia, 1999. [Google Scholar]
- Ladizinsky, G. The genetics of hard seed coat in the genus Lens. Euphytica 1985, 34, 539–543. [Google Scholar] [CrossRef]
- Forbes, I.; Wells, H. Hard and soft seededness in Blue Lupine, Lupinus angustifolius L.: Inheritance and pheno-type classification. Crop Sci. 1968, 8, 195–197. [Google Scholar] [CrossRef]
- Kongjaimun, A.; Kaga, A.; Tomooka, N.; Somta, P.; Vaughan, D.A.; Srinives, P. The genetics of domestication of yardlong bean, Vigna unguiculata (L.) Walp. ssp. unguiculata cv.-gr. sesquipedalis. Ann. Bot. 2012, 109, 1185–1200. [Google Scholar] [CrossRef] [PubMed]
- Isemura, T.; Kaga, A.; Tomooka, N.; Shimizu, T.; Vaughan, D.A. The genetics of domestication of rice bean, Vigna umbellata. Ann. Bot. 2010, 106, 927–944. [Google Scholar] [CrossRef] [PubMed]
- Isemura, T.; Kaga, A.; Tabata, S.; Somta, P.; Srinives, P.; Shimizu, T.; Jo, U.; Vaughan, D.A.; Tomooka, N. Construction of a genetic linkage map and genetic analysis of domestication related traits in Mungbean (Vigna radiata). PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Weeden, N.F. Genetic changes accompanying the domestication of Pisum sativum: Is there a common genetic basis to the “domestication syndrome” for legumes? Ann. Bot. 2007, 100, 1017–1025. [Google Scholar] [CrossRef]
- Sun, L.; Miao, Z.; Cai, C.; Zhang, D.; Zhao, M.; Wu, Y.; Zhang, X.; Swarm, S.A.; Zhou, L.; Zhang, Z.J.; et al. GmHs1-1, encoding a calcineurin-like protein, controls hard-seededness in soybean. Nat. Genet. 2015, 47, 939–943. [Google Scholar] [CrossRef]
- Jang, S.; Sato, M.; Sato, K.; Jitsuyama, Y.; Fujino, K.; Mori, H.; Takahashi, R.; Benitez, E.R.; Liu, B.; Yamada, T. A Single-Nucleotide Polymorphism in an Endo-1, 4-β-Glucanase Gene Controls Seed Coat Permeability in Soybean. PLoS ONE 2015, 1–19. [Google Scholar] [CrossRef]
- Donnelly, E.D.; Watson, J.E.; John, A. Inheritance of Hard Seed in Vicia. J. Hered. 1972, 7, 361–365. [Google Scholar] [CrossRef]
- Chinnasamy, G.; Bal, A.K. The pattern of seed development and maturation in beach pea (Lathyrus maritimus). Can. J. Bot. 2003, 81, 531–540. [Google Scholar] [CrossRef]
- Gresta, F.; Avola, G.; Onofri, A.; Anastasi, U.; Cristaudo, A. When does hard coat impose dormancy in legume seeds? Lotus and scorpiurus case study. Crop Sci. 2011, 51, 1739–1747. [Google Scholar] [CrossRef]
- Aitken, Y. The problem of hard seeds in subterranean clover. Proc. R. Soc. Vic. 1939, 51, 187–213. [Google Scholar]
- Hyde, E.O. The Function of the Hilum in Some Papilionaceae in Relation to the Ripening of the Seed and the Permeability of the Testa. Ann. Bot. 1954, 18, 241–256. [Google Scholar] [CrossRef]
- D’Hondt, B.; Brys, R.; Hoffmann, M. The incidence, field performance and heritability of non-dormant seeds in white clover (Trifolium repens L.). Seed Sci. Res. 2010, 20, 169–177. [Google Scholar] [CrossRef]
- Tozer, M.G.; Ooi, M.K.J. Humidity-regulated dormancy onset in the Fabaceae: A conceptual model and its ecological implications for the Australian wattle Acacia saligna. Ann. Bot. 2014, 114, 579–590. [Google Scholar] [CrossRef]
- Jones, R.M.; Bunch, G.A. The effect of stocking rate on the population dynamics of siratro in siratro (Macroptilium atropurpureum)-setaria (Setaria sphacelata) pastures in south-east Queensland. II* Seed set, soil seed reserves, seedling recruitment and seedling survival. Aust. J. Agric. Res. 1987, 39, 221–234. [Google Scholar] [CrossRef]
- Argel, P.; Humphreys, L. Environmental effects on seed development and hardseededness in Stylosanthes hamata cv. Verano. I. Temperature. Aust. J. Agric. 1983, 34, 261–270. [Google Scholar] [CrossRef]
- Baciu-Miclau, D. Contribution to the study of hard seed and coat structure properties of soybean. Proc. Int. Seed Test. Assoc. 1970, 35, 599–617. [Google Scholar]
- Kissing Kucek, L.; Riday, H.; Ehlke, N.; Reberg-Horton, C.; Maul, J.; Mirsky, S.B.; Pelzer, C.J.; Poskaitis, M.; Ryan, M.R.; Seehaver, S.; et al. Environmental Influences on the Relationship between Fall and Spring Vigor in Hairy Vetch. Crop Sci. 2019, 59, 2443–2454. [Google Scholar] [CrossRef]
- Kalu, B.A.; Fick, G.W. Quantifying morphological development of alfalfa for studies of herbage quality. Crop Sci. 1981, 21, 267–271. [Google Scholar] [CrossRef]
- Renzi, J.P. Efecto de la Estructura del Cultivo y el Grado de Madurez a Cosecha Sobre el Rendimie to y la Calidad de Semillas de Vicia Sativa L., Bajo Riego. Ph.D. Thesis, La Universidad Nacional del Sur, Bahía Blanca, Argentina, 2009; p. 143. [Google Scholar]
- Kissing Kucek, L.; Riday, H.; Rufener, B.P.; Burke, A.N.; Eagen, S.S.; Ehlke, N.; Krogman, S.; Mirsky, S.B.; Reberg-Horton, C.; Ryan, M.R.; et al. Pod Dehiscence in Hairy Vetch (Vicia villosa Roth). Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Elias, S.G.; Copeland, L.O.; Mcdonald, M.B.; Baalbaki, R.Z. Germination and Viability Testing. In Seed Testing: Principles and Practices; Michigan State University Press: East Lansing, MI, USA, 2012; pp. 59–81. [Google Scholar]
- Samarah, N.H.; Allataifeh, N.; Turk, M.A.; Tawaha, A.M. Seed germination and dormancy of fresh and air-dried seeds of common vetch (Vicia sativa L.) harvested at different stages of maturity. Seed Sci. Technol. 2004, 32, 11–19. [Google Scholar] [CrossRef]
- Samarah, N.H. Effect of air-drying immature seeds in harvested pods on seed quality of common vetch (Vicia sativa L.). New Zeal. J. Agric. Res. 2006, 49, 331–339. [Google Scholar] [CrossRef]
- Brar, G.S.; Gomez, J.F.; McMichael, B.L.; Matches, A.G.; Taylor, H.M. Germination of Twenty Forage Legumes as Influenced by Temperature. Agron. J. 1991, 83, 173–175. [Google Scholar] [CrossRef]
- Renzi, J.; Cantamutto, M. Dormancia y germinacion en semillas de Vicia villosa Roth. Análisis Semillas 2009, 3, 84–89. [Google Scholar]
- Kissing, K.L.; Riday, H.; Wiering, N.P.; Azevedo, M.; Eagen, S.S.; Ehlke, N.; Hayes, R.J.; Krogman, S.; Mirsky, S.B.; Reberg-Horton, C.; et al. Vicia villosa seed physical dormancy dataset from 2017–2019. Ag Data Commons 2020. [Google Scholar] [CrossRef]
- Lynch, M.; Walsh, B. Genetics and Analysis of Quantitative Traits; Sinauer Assoc Inc.: Sunderland, MA, USA, 1998. [Google Scholar]
- Vazquez, A.I.; Bates, D.M.; Rosa, G.J.M.; Gianola, D.; Weigel, K.A. Technical note: An R package for fitting generalized linear mixed models in animal breeding. J. Anim. Sci. 2010, 88, 497–504. [Google Scholar] [CrossRef]
- Butler, D.G.; Cullis, B.R.; Gilmour, A.R.; Gogel, B.J.; Thompson, R. ASReml-R Reference Manual Version 4. 2017. p. 176. Available online: https://asreml.kb.vsni.co.uk/wp-content/uploads/sites/3/2018/02/ASReml-R-Reference-Manual-4.pdf (accessed on 1 September 2020).
- Meuwissen, T.H.E.; Luo, Z. Computing inbreeding coefficients in large populations. Genet. Sel. Evol. 1992, 24, 305–313. [Google Scholar] [CrossRef]
- R Core Team, R. A Language and Environment for Statistical Computing Organization; R Core Team R: Vienna, Autria, 2019. [Google Scholar]
- Teasdale, J.R.; Devine, T.E.; Mosjidis, J.A.; Bellinder, R.R.; Beste, C.E.; June, M. Growth and Development of Hairy Vetch Cultivars in the Northeastern United States as Influenced by Planting and Harvesting Date. Agron. J. 2004, 96, 1266–1271. [Google Scholar] [CrossRef]
- Baskerville, A.G.L.; Emin, P. Rapid Estimation of Heat Accumulation from Maximum and Minimum Temperatures. Ecology 1969, 50, 514–517. [Google Scholar] [CrossRef]
- van de Pol, M.; Cockburn, A. Identifying the critical climatic time window that affects trait expression. Am. Nat. 2011, 177, 698–707. [Google Scholar] [CrossRef]
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar]
- Norman, H.C.; Cocks, P.S.; Galwey, N.W. Hardseededness in annual clovers: Variation between populations from wet and dry environments. Aust. J. Agric. Res. 2002, 53, 821–829. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).