Broadening the Variability for Peanut Breeding with a Wild Species-Derived Induced Allotetraploid

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population Development
2.2. Morphological, Agronomic and Leaf Spots Resistance Evaluations
2.3. Linkage Map Construction
2.4. QTL Analysis
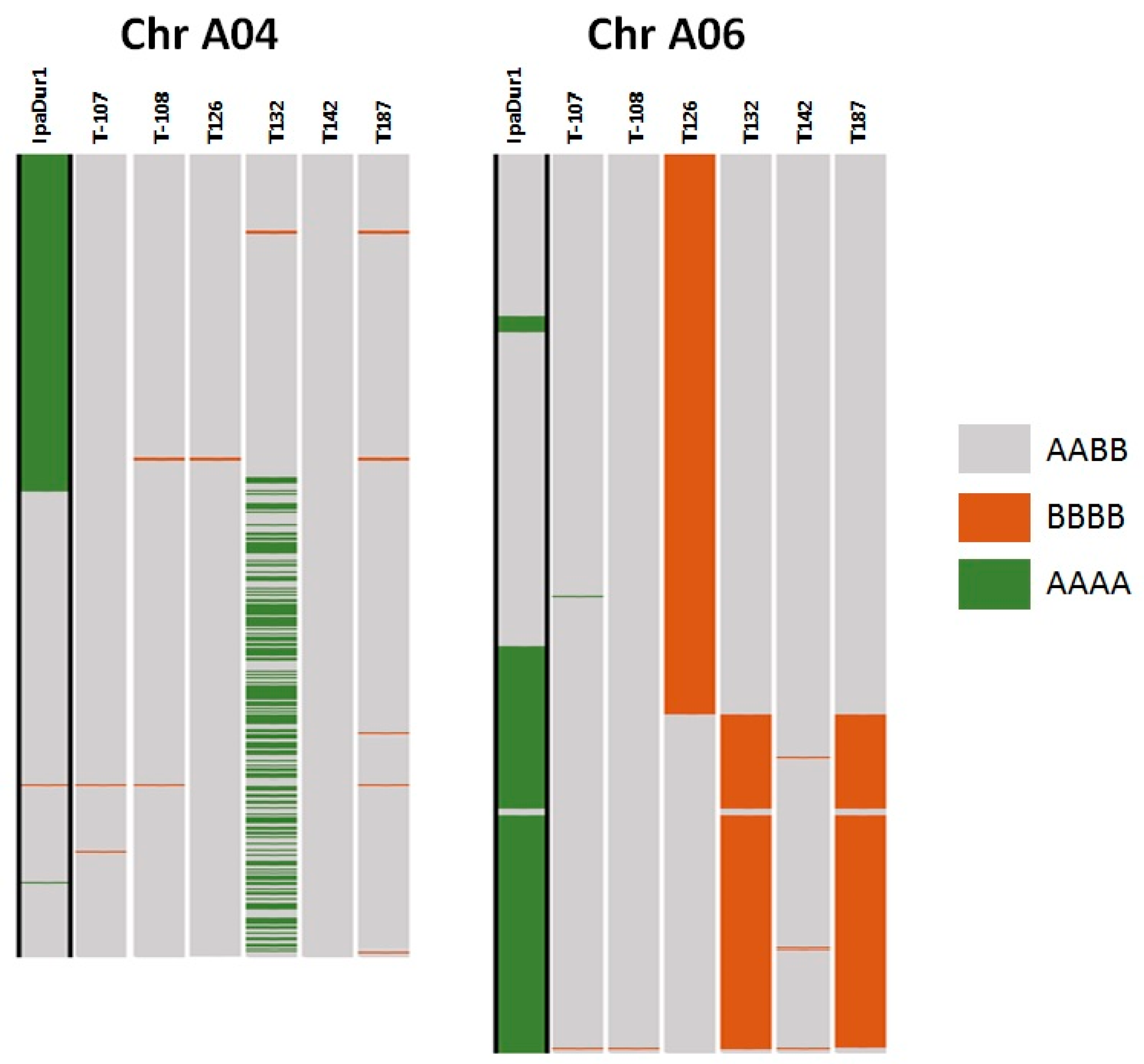
2.5. SNP Genotyping and Genetic Analysis of Selected Lines
3. Results
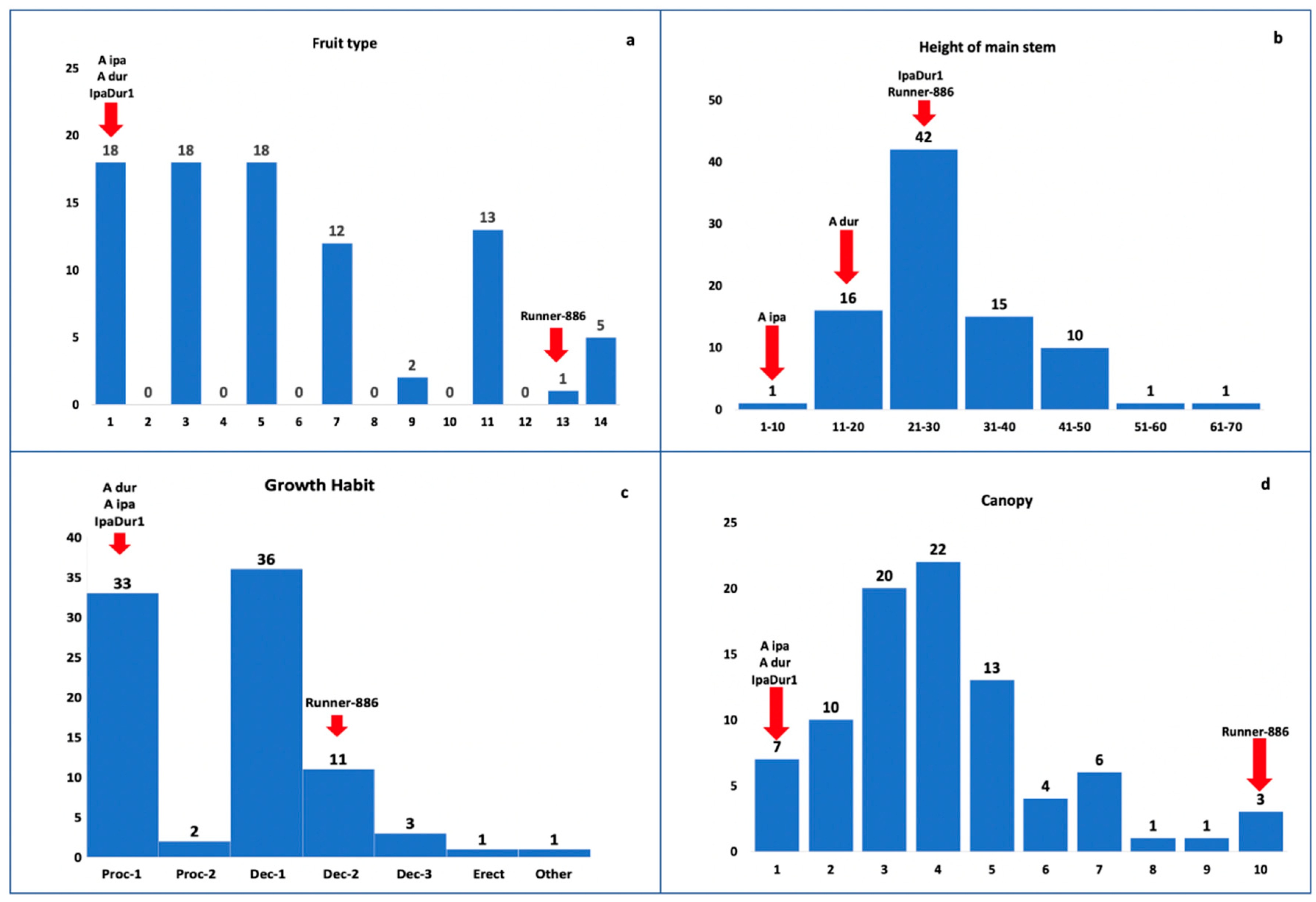
3.1. Morphological Traits
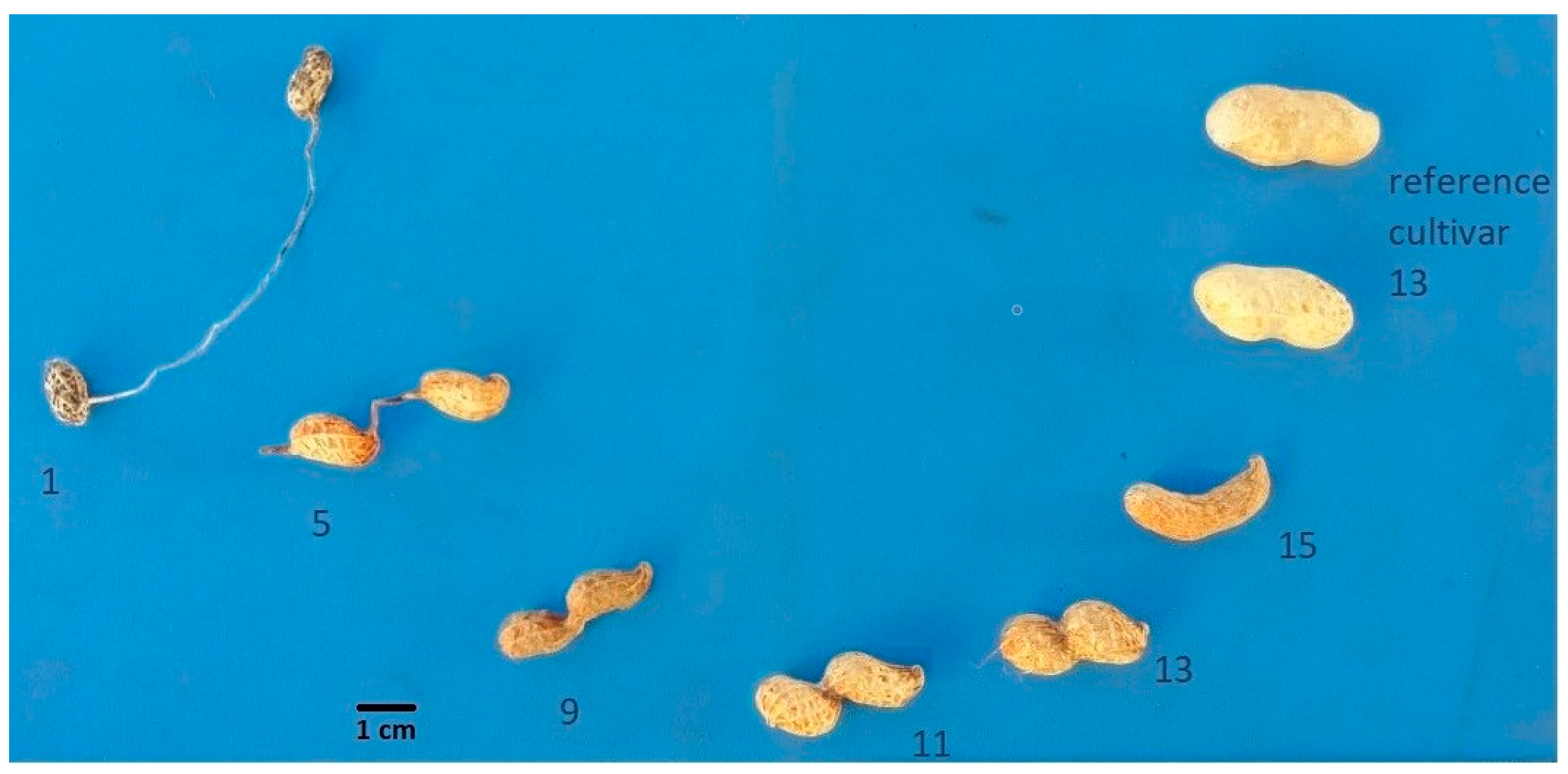
3.2. Fruit Types in an Interspecific Population
3.3. Agronomic Traits
3.4. Leaf Spot Resistance
3.5. Linkage Map
3.6. QTL Identification
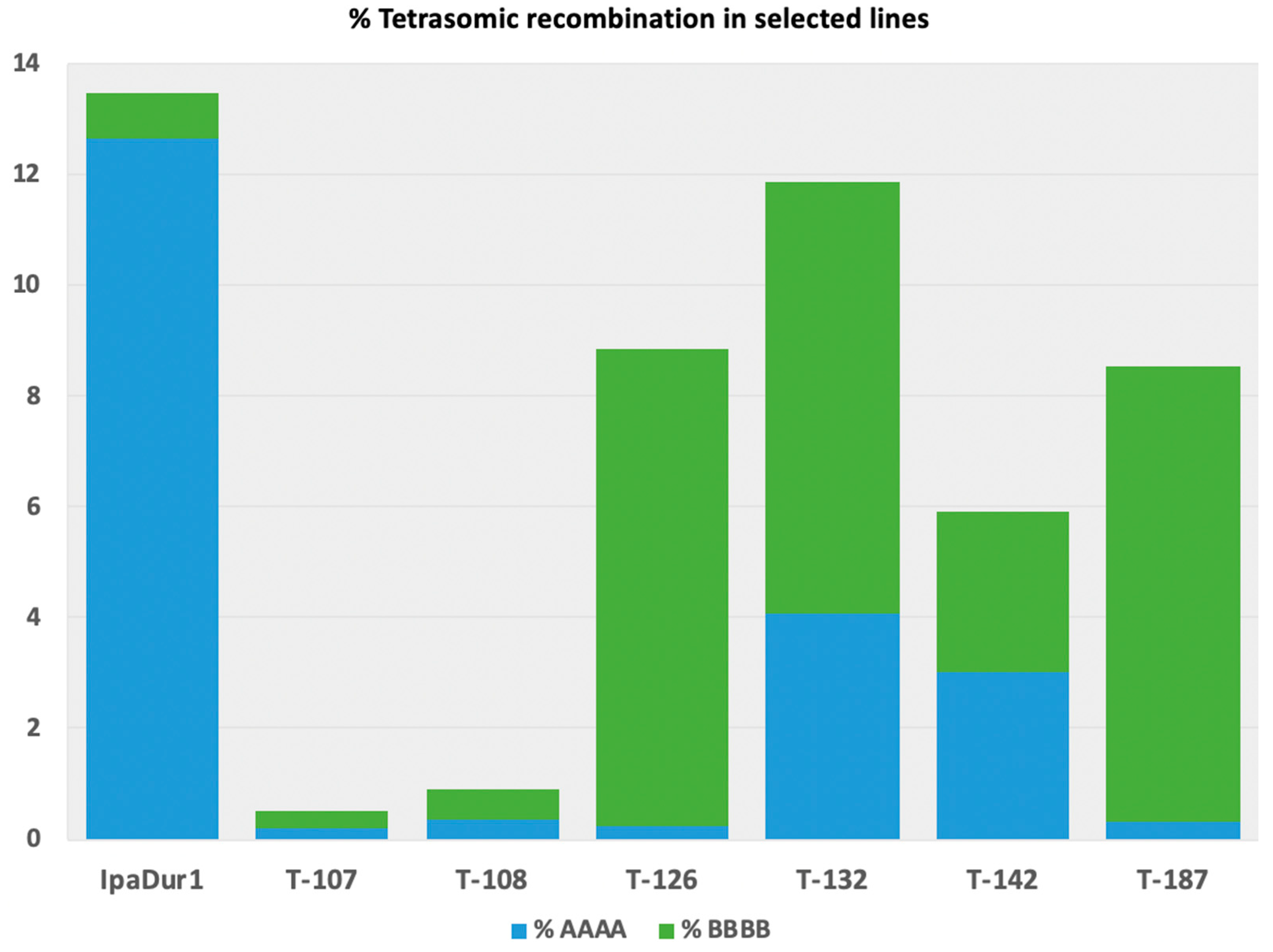
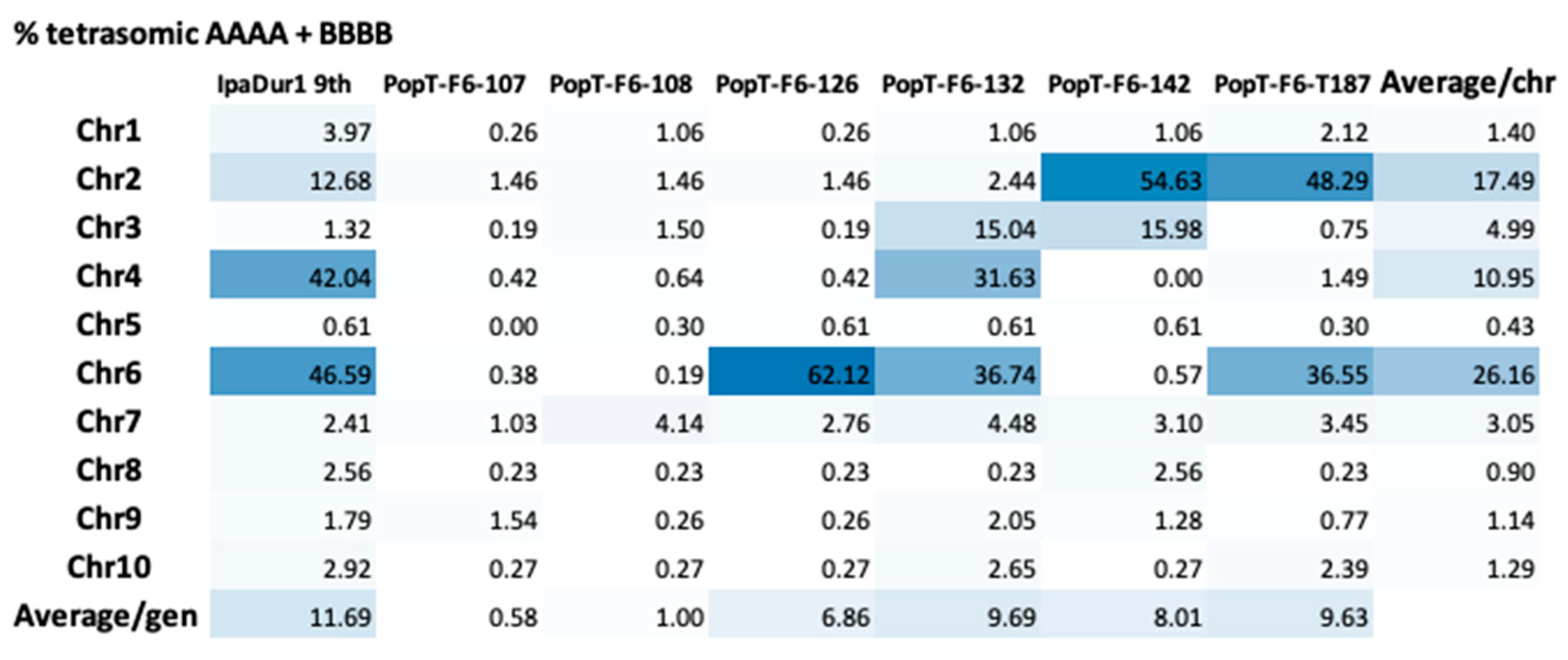
3.7. Genotyping and Genetic Analysis of Selected RILs with Contrasting Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- USDA-FAS. World agricultural production. In Circular Series WAP 9-20; Services, F.A., Ed.; USDA: Washington, DC, USA, 2020; p. 42. [Google Scholar]
- Holbrook, C.C.; Brenneman, T.B.; Stalker, H.T.; Johnson, W.C., III; Ozias-Akins, P.; Chu, Y.; Vellidis, G.; Mccluski, D. Peanut. In Yield Gains in Major US Field Crops; Smith, S., Diers, S.B., Eds.; CSSA: Madison, WI, USA, 2014; pp. 173–194. [Google Scholar] [CrossRef]
- Suassuna, T.M.F.; Suassuna, N.D.; Bogiani, J.C.; Perina, F.; Fragoso, D.B.; Sofiatti, V.; Medeiros, E.P.; Moretzsohn, M.C.; Leal-Bertioli, S.C.M.; Bertioli, D.J.; et al. BRS 425: The first runner peanut cultivar related to wild ancestral species. Crop Breed. Appl. Biotechnol. 2019, 19, 373–377. [Google Scholar] [CrossRef]
- Holbrook, C.C.; Stalker, H.T. Peanut Breeding and Genetic Resources. In Plant Breeding Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken NJ, USA, 2002; pp. 297–356. [Google Scholar] [CrossRef]
- Stalker, H.T. Utilizing Wild Species for Peanut Improvement. Crop Sci. 2017, 57, 1102–1120. [Google Scholar] [CrossRef]
- Stalker, H.T.; Wynne, J.C.; Company, M. Variation in progenies of an Arachis hypogaea x diploid wild species hybrid. Euphytica 1979, 28, 675–684. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Dudchenko, O.; Gao, D.; Seijo, G.; Leal-Bertioli, S.C.M.; Ren, L.; Farmer, A.D.; Pandey, M.K.; et al. The genome sequence of segmental allotetraploid peanut Arachis hypogaea. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Seijo, G.; Lavia, G.I.; Fernández, A.; Krapovickas, A.; Ducasse, D.A.; Bertioli, D.J.; Moscone, E.A. Genomic relationships between the cultivated peanut (Arachis hypogaea, Leguminosae) and its close relatives revealed by double GISH. Am. J. Bot. 2007, 94, 1963–1971. [Google Scholar] [CrossRef]
- Leal-Bertioli, S.C.M.; Moretzsohn, M.C.; Santos, S.P.; Brasileiro, A.C.M.; Guimarães, P.M.; Bertioli, D.J.; Araujo, A.C.G. Phenotypic effects of allotetraploidization of wild Arachis and their implications for peanut domestication. Am. J. Bot. 2017, 104, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Krapovickas, A.; Gregory, W.C. Taxonomia del genero Arachis (Leguminosae). Bonplandia 2007, 8, 1–186. [Google Scholar] [CrossRef] [Green Version]
- Fávero, A.P.; Simpson, C.E.; Valls, J.F.M.; Vello, N.A. Study of the evolution of cultivated peanut through crossability studies among Arachis ipaënsis, A. duranensis, and A. hypogaea. Crop Sci. 2006, 46, 1546–1552. [Google Scholar] [CrossRef]
- Shirasawa, K.; Bertioli, D.J.; Varshney, R.K.; Moretzsohn, M.C.; Leal-Bertioli, S.C.M.; Thudi, M.; Pandey, M.K.; Rami, J.-F.; Foncéka, D.; Gowda, M.V.C.; et al. Integrated consensus map of cultivated peanut and wild relatives reveals structures of the A and B genomes of Arachis and divergence of the legume genomes. DNA Res. 2013, 20, 173–184. [Google Scholar] [CrossRef] [Green Version]
- IBPGR; ICRISAT. Descriptors for Groundnut; Resources, I.B.F.P.G., Ed.; IBPGR/ICRISAT: Rome, Italy, 1992; p. 23. [Google Scholar]
- Leal-Bertioli, S.C.M.; Godoy, I.J.; Santos, J.F.; Doyle, J.J.; Guimarães, P.M.; Abernathy, B.L.; Jackson, S.A.; Moretzsohn, M.C.; Bertioli, D.J. Segmental allopolyploidy in action: Increasing diversity through polyploid hybridization and homoeologous recombination. Am. J. Bot. 2018, 105, 1053–1066. [Google Scholar] [CrossRef] [Green Version]
- Subrahmanyam, P.; Mcdonald, D.; Waliyar, F.; Reddy, L.J.; Nigam, S.N.; Gibbons, R.W.; Ramanatha, V.R.; Singh, A.K.; Pande, S.; Reddy, P.M.; et al. Screening Methods and Sources of Resistance to Rust and Late Leaf Spot of Groundnut; ICRISAT: Patancheru, India, 1992; p. 20. [Google Scholar]
- Czermainski, A.B.C. Generalização de um índice de intensidade de infecção em experimentos de doenças em plantas. Pesqui. Agropecuária Bras. 1999, 34, 1545–1555. [Google Scholar] [CrossRef] [Green Version]
- Resende, M.D.V. Software Selegen-REML/BLUP: A useful tool for plant breeding. Crop Breed. Appl. Biotechnol. 2016, 16, 330–339. [Google Scholar] [CrossRef]
- Duarte, J.B.; Vencovsky, R. Estimação e predição por modelo linear misto com ênfase na ordenação de médias de tratamentos genéticos. Sci. Agric. 2001, 58, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Resende, M.D.V.; Duarte, J.B. Precisão e controle de qualidade em experimentos de avaliação de cultivares. Pesqui. Agropecuária Trop. 2007, 37, 182–194. [Google Scholar]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.S.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The genome sequences of Arachis duranensis and Arachis ipaensis, the diploid ancestors of cultivated peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Ozias-Akins, P.; Chu, Y.; Dantas, K.M.; Santos, S.P.; Gouvea, E.; Guimarães, P.M.; Leal-Bertioli, S.C.M.; Knapp, S.J.; Moretzsohn, M.C. The Use of SNP Markers for Linkage Mapping in Diploid and Tetraploid Peanuts. G3 Genes Genomes Genet. 2014, 4, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Van Ooijen, J.W. JoinMap 4. Software for the Calculation of Genetic Linkage Maps in Experimental Populations; Kyazma B.V.: Wageningen, The Netherlands, 2006. [Google Scholar]
- Sujay, V.; Gowda, M.V.C.; Pandey, M.K.; Bhat, R.S.; Khedikar, Y.P.; Nadaf, H.L.; Gautami, B.; Sarvamangala, C.; Lingaraju, S.; Radhakrishan, T.; et al. Quantitative trait locus analysis and construction of consensus genetic map for foliar disease resistance based on two recombinant inbred line populations in cultivated groundnut (Arachis hypogaea L.). Mol. Breed. 2012, 30, 773–788. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Pandey, M.K.; Janila, P.; Nigam, S.N.; Sudini, H.; Gowda, M.V.C.; Sriswathi, M.; Radhakrishnan, T.; Manohar, S.S.; Nagesh, P. Marker-assisted introgression of a QTL region to improve rust resistance in three elite and popular varieties of peanut (Arachis hypogaea L.). Theor. Appl. Genet. 2014, 127, 1771–1781. [Google Scholar] [CrossRef] [Green Version]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newburg, L. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Lincoln, S.E.; Daly, M.; Lander, E.S. Constructiong Genetic Maps with MAPMAKER/EXP3.0; Whitehead Institute Technical Report: Cambridge, MA, USA, 1992. [Google Scholar]
- Moretzsohn, M.C.; Gouvea, E.G.; Inglis, P.W.; Leal-Bertioli, S.C.M.; Valls, J.F.M.; Bertioli, D.J. A study of the relationships of cultivated peanut (Arachis hypogaea) and its most closely related wild species using intron sequences and microsatellite markers. Ann. Bot. 2013, 111, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Basten, C.J.; Zeng, Z.-B. Windows QTL Cartographer 2.5; Department of Statistics, North Carolina State University: Raleigh, NC, USA, 2012. [Google Scholar]
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churchill, G.A.; Doerge, R.W. Empirical threshold values for quantitative trait mapping. Genetics 1994, 138, 963–971. [Google Scholar] [PubMed]
- Doerge, R.W.; Churchill, G.A. Permutation Tests for Multiple Loci Affecting a Quantitative Character. Genetics 1996, 142, 285–294. [Google Scholar] [PubMed]
- Voorrips, R.E. MapChart: Software for the Graphical Presentation of Linkage Maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clevenger, J.; Chu, Y.; Chavarro, C.; Agarwal, G.; Bertioli, D.J.; Leal-Bertioli, S.C.M.; Pandey, M.K.; Vaughn, J.; Abernathy, B.; Barkley, N.A.; et al. Genome-wide SNP Genotyping Resolves Signatures of Selection and Tetrasomic Recombination in Peanut. Mol. Plant 2017, 10, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korani, W.; Clevenger, J.P.; Chu, Y.; Ozias-Akins, P. Machine Learning as an Effective Method for Identifying True Single Nucleotide Polymorphisms in Polyploid Plants. Plant Genome 2019, 12, 180023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonceka, D.; Tossim, H.-A.; Rivallan, R.; Vignes, H.; Faye, I.; Ndoye, O.; Moretzsohn, M.C.; Bertioli, D.J.; Glaszmann, J.-C.; Courtois, B.; et al. Fostered and left behind alleles in peanut: Interspecific QTL mapping reveals footprints of domestication and useful natural variation for breeding. BMC Plant Biol. 2012, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Chopra, R.; Johnson, E.B.; Emenecker, R.; Cahoon, E.B.; Lyons, J.; Kliebenstein, D.J.; Daniels, E.; Dorn, K.M.; Esfahanian, M.; Folstad, N.; et al. Identification and stacking of crucial traits required for the domestication of pennycress. Nat. Food 2020, 1, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Yang, X.; Liu, J.; Wang, B.-H.; Liu, B.-L.; Wang, Y.-Z. Pod shattering resistance associated with domestication is mediated by a NAC gene in soybean. Nat. Commun. 2014, 5, 3352. [Google Scholar] [CrossRef] [Green Version]
- Michelotto, M.D.; Barioni, W.J.; Resende, M.D.V.; Godoy, I.J.; Leonardecz, E.; Fávero, A.P. Identification of Fungus Resistant Wild Accessions and Interspecific Hybrids of the Genus Arachis. PLoS ONE 2015, 10, e0128811. [Google Scholar] [CrossRef] [Green Version]
- Suassuna, T.M.F.; Suassuna, N.D.; Moretzsohn, M.C.; Bertioli, S.C.M.L.; Bertioli, D.J.; Medeiros, E.P. Yield, market quality, and leaf spots partial resistance of interspecific peanut progenies. Crop Breed. Appl. Biotechnol. 2015, 15, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Ali, L.; Azam, S.; Rubio, J.; Kudapa, H.; Madrid, E.; Varshney, R.K.; Castro, P.; Chen, W.; Gil, J.; Millan, T. Detection of a new QTL/gene for growth habit in chickpea CaLG1 using wide and narrow crosses. Euphytica 2015, 204, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Aryamanesh, N.; Nelson, M.N.; Yan, G.; Clarke, H.J.; Siddique, K.H.M. Mapping a major gene for growth habit and QTLs for ascochyta blight resistance and flowering time in a population between chickpea and Cicer reticulatum. Euphytica 2010, 173, 307–319. [Google Scholar] [CrossRef]
- Huyghe, C. Genetics and genetic modifications of plant architetcture in grain legumes: A review. Agronomie 1998, 18, 383–411. [Google Scholar] [CrossRef]
- Saha, G.C.; Sarker, A.; Chen, W.; Vandemark, G.J.; Muehlbauer, F.J. Inheritance and Linkage Map Positions of Genes Conferring Agromorphological Traits in Lens culinaris Medik. Int. J. Agron. 2013, 2013, 618926. [Google Scholar] [CrossRef] [Green Version]
- Leal-Bertioli, S.C.M.; Shirasawa, K.; Abernathy, B.; Moretzsohn, M.C.; Chavarro, C.; Clevenger, J.; Ozias-Akins, P.; Jackson, S.A.; Bertioli, D.J. Tetrasomic Recombination Is Surprisingly Frequent in Allotetraploid. Arachis. Genetics 2015, 199, 1093–1105. [Google Scholar] [CrossRef] [Green Version]
- Nguepjop, J.R.; Tossim, H.-A.; Bell, J.M.; Rami, J.-F.; Sharma, S.; Courtois, B.; Mallikarjuna, N.; Sane, D.; Fonceka, D. Evidence of Genomic Exchanges between Homeologous Chromosomes in a Cross of Peanut with Newly Synthetized Allotetraploid Hybrids. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Gowda, M.V.C.; Motagi, B.N.; Naidu, G.K.; Diddimani, S.N.; Sheshagiri, R. GPBD-4: A Spanish bunch groundnut genotype resistant to rust and late leaf spot. Int. Arachis Newsl. 2002, 22, 29–32. [Google Scholar]
- Lamon, S.; Chu, Y.; Guimaraes, L.A.; Bertioli, D.J.; Leal-Bertioli, S.C.M.; Santos, J.F.; Godoy, I.J.; Culbreath, A.K.; Holbrook, C.C.; Ozias-Akins, P. Characterization of peanut lines with interspecific introgressions conferring Late Leaf Spot resistance. Crop Sci. 2020, in press. [Google Scholar] [CrossRef]
- Simpson, C.E.; Starr, J.L.; Baring, M.R.; Burow, M.D.; Cason, J.M.; Wilson, J.N. Registration of ‘Webb’ Peanut. J. Plant Regist. 2013, 7, 265–268. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fruit Type | Description | Rate |
|---|---|---|
| Wild | Biarticulate fruit, articles separated by a long isthmus | 1 |
| Mixed wild | Biarticulate fruit, articles separated by isthmus with long and intermediate length. | 3 |
| Wild intermediate | Biarticulate fruit, articles separated by isthmus of intermediate length | 5 |
| Mixed wild and cultivated | Biarticulated and not articulated fruits; varying isthmus lengths or pod constriction | 7 |
| Cultivated 1 | Fruit not articulated; very deep constriction | 9 |
| Cultivated 2 | Fruit not articulated; deep constriction | 11 |
| Cultivated 3 | Fruit not articulated; moderate constriction | 13 |
| Mixed cultivated | Fruit not articulated; different degrees of constriction | 14 |
| Cultivated 4 | No isthmus or constriction | 15 |
| Effects | MSEV | ISEV | AUDPC-MSEV | AUDPC-ISEV |
|---|---|---|---|---|
| Treatment | −29.46 | 449.82 | 665.14 | 1148.65 |
| Complete | −38.67 | 440.64 | 653.88 | 1137.90 |
| LRT (chi-square 1) | 9.21 ** | 9.18 ** | 11.26 ** | 10.75 ** |
| Trait | Chrom. | Position (cM) 2 | Nearest Marker | LOD | Additive Effect 3 | R2 (%) 4 |
|---|---|---|---|---|---|---|
| MSEV 1 | A08 | 64.1 | AHGS1385_A | 4.0 | 0.3523 | 11.0 |
| B02 | 13.5 | AhTE0791 | 6.3 | −0.6309 | 20.1 | |
| ISEV 1 | A08 | 64.1 | AHGS1385_A | 3.6 | 2.8201 | 10.0 |
| B02 | 13.5 | AhTE0791 | 5.9 | −5.3011 | 19.7 | |
| B02 | 17.3 | Ah_FI501986 | 2.7 | −2.5501 | 8.4 | |
| AUDPC-MSEV | A02 | 73.2 | RM2H10 | 5.2 | 16.5797 | 12.8 |
| A08 | 85.0 | RM17H09 | 5.9 | 14.7175 | 14.9 | |
| B05 | 4.2 | AHGS2285_2 | 3.8 | 11.5223 | 9.0 | |
| B05 | 45.9 | DS_c6795_188 | 3.7 | −11.3779 | 8.7 | |
| AUDPC-ISEV | A02 | 74.5 | Seq15D2 | 4.6 | 127.3087 | 12.6 |
| A06 | 84.3 | AHGS1478 | 2.5 | −84.2252 | 6.4 | |
| A08 | 85.0 | RM17H09 | 4.7 | 123.2982 | 13.1 | |
| B05 | 4.2 | AHGS2285_2 | 3.3 | 99.8630 | 8.6 | |
| B05 | 45.9 | DS_c6795_188 | 6.7 | −147.5304 | 19.2 | |
| HMS | A04 | 58.5 | AHGS2785 | 7.4 | −5.7618 | 21.6 |
| A04 | 88.6 | AHGS2390_A | 7.9 | 5.7943 | 20.0 | |
| A08 | 46.4 | AHGS3698 | 5.4 | 3.6912 | 12.3 | |
| B02 | 24.9 | Seq18E7 | 6.2 | 4.1874 | 15.6 | |
| B05 | 10.4 | AHGS1203 | 5.0 | 3.8839 | 11.0 | |
| B05 | 45.9 | DS_c6795_188 | 5.3 | −3.7708 | 11.9 | |
| GH | A04 | 106.0 | DS_c3103_115 # | 4.2 | n.d 5 | n.d 5 |
| Canopy | A02 | 87.4 | AHS1008 | 7.3 | −1.6459 | 21.3 |
| Fruit type | A04 | 0.0 | TC21A09 | 6.9 | 0.1706 | 16.7 |
| A08 | 37.2 | DS_c582_344 | 7.1 | 0.1649 | 17.3 | |
| A08 | 42.3 | RN22A12 | 8.5 | 0.1823 | 19.8 | |
| B03 | 43.4 | AHGS1661 | 5.1 | −0.1676 | 12.9 |
| RIL/Accession | Standard Petal Color | Height of Main Stem (cm) | Growth Habit | Canopy | Fruit Type | GV MSEV * |
|---|---|---|---|---|---|---|
| T107 | Yellow | 25.3 | 1 | 5 | 11 | 5.9 |
| T108 | Yellow | 38.5 | 3 | 7 | 14 | 5.6 |
| T126 | Yellow | 41.0 | 3 | 2 | 7 | 5.4 |
| T132 | Yellow | 38.5 | 1 | 5 | 11 | 5.8 |
| T142 | Orange | 23.8 | 3 | 7 | 5 | 5.5 |
| T187 | Yellow | 17.0 | 6 | 10 | 11 | 5.6 |
| ‘Runner IAC 886′ | Orange | 27.3 | 4 | 10 | 13 | 6.1 |
| K 30076 | Orange | 7.5 | 1 | 1 | 1 | 5.3 |
| V 14167 | Yellow | 14.8 | 1 | 1 | 1 | 5.8 |
| IpaDur1 | Yellow | 27.8 | 1 | 1 | 1 | 5.3 |
| IpaDur1 | T107 | T108 | T126 | T132 | T142 | T187 | |
|---|---|---|---|---|---|---|---|
| 2x A. duranensis loci | 236 | 145 | 162 | 86 | 168 | 130 | 100 |
| 2x A. ipaënsis loci | 208 | 107 | 127 | 148 | 79 | 114 | 156 |
| Ratio wild/cultivated | * | 1.01 | 1.16 | 0.86 | 1.03 | 1.12 | 0.92 |
| A. duranensis/A. ipaënsis | 1.50 | 1.09 | 1.10 | 0.59 | 1.43 | 0.82 | 0.60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suassuna, T.; Suassuna, N.; Martins, K.; Matos, R.; Heuert, J.; Bertioli, D.; Leal-Bertioli, S.; Moretzsohn, M. Broadening the Variability for Peanut Breeding with a Wild Species-Derived Induced Allotetraploid. Agronomy 2020, 10, 1917. https://doi.org/10.3390/agronomy10121917
Suassuna T, Suassuna N, Martins K, Matos R, Heuert J, Bertioli D, Leal-Bertioli S, Moretzsohn M. Broadening the Variability for Peanut Breeding with a Wild Species-Derived Induced Allotetraploid. Agronomy. 2020; 10(12):1917. https://doi.org/10.3390/agronomy10121917
Chicago/Turabian StyleSuassuna, Taís, Nelson Suassuna, Kennedy Martins, Ramon Matos, Jair Heuert, David Bertioli, Soraya Leal-Bertioli, and Márcio Moretzsohn. 2020. "Broadening the Variability for Peanut Breeding with a Wild Species-Derived Induced Allotetraploid" Agronomy 10, no. 12: 1917. https://doi.org/10.3390/agronomy10121917
APA StyleSuassuna, T., Suassuna, N., Martins, K., Matos, R., Heuert, J., Bertioli, D., Leal-Bertioli, S., & Moretzsohn, M. (2020). Broadening the Variability for Peanut Breeding with a Wild Species-Derived Induced Allotetraploid. Agronomy, 10(12), 1917. https://doi.org/10.3390/agronomy10121917