Impact of Winter Cover Crop Usage in Soilborne Disease Suppressiveness in Woody Ornamental Production System


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experimental Design and Layout
2.2. Cover Crop Application
2.3. Soil Moisture and Temperature Measurement
2.4. Soil Sampling
2.5. Fungal Culture and Pathogen Inoculum Preparation
2.6. Red Maple Propagation
2.7. Bioassays for Soilborne Disease Suppressiveness
2.7.1. Crop Health Assessment
2.7.2. Fluorescent Pseudomonad Population
2.8. Statistical Analysis
3. Results
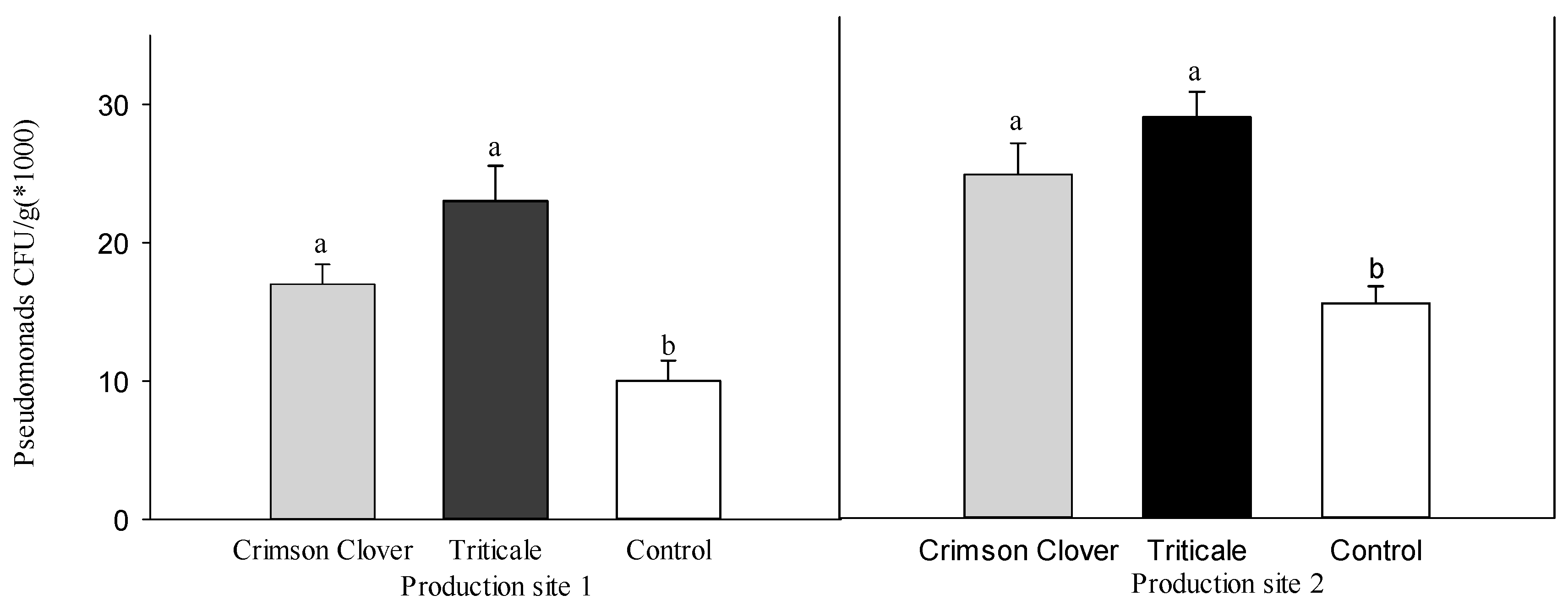
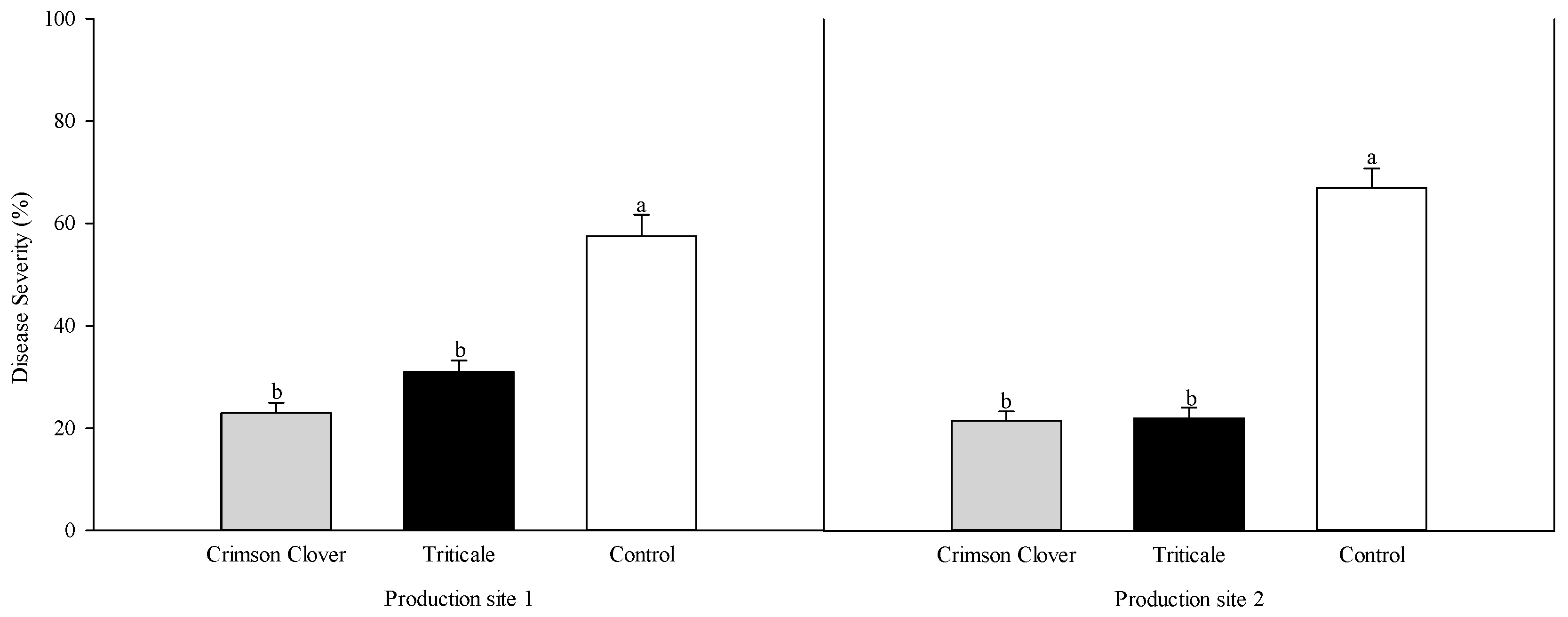
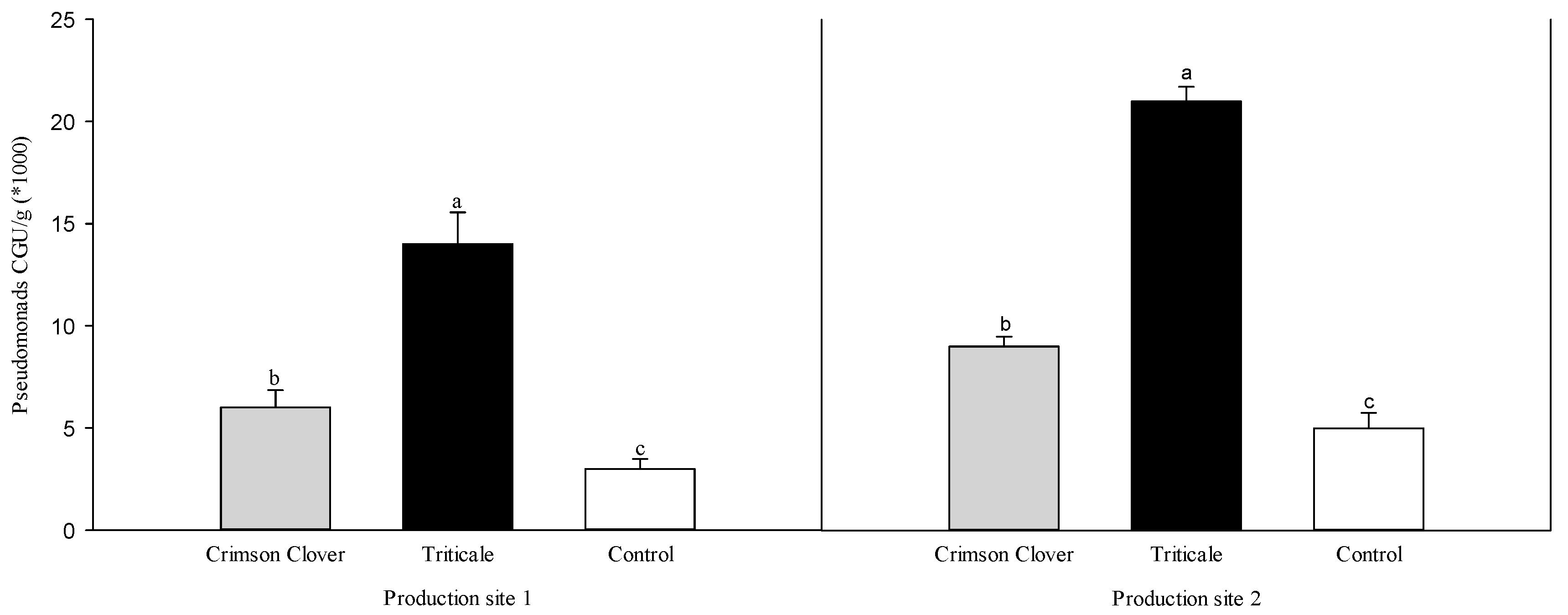
3.1. Bioassay with Natural Pathogen Pressure
3.2. Bioassay with Phytopythium vexans
3.3. Bioassay with Phytophthora nicotianae
3.4. Bioassay with Rhizoctonia solani
3.5. Soil Moisture and Temperature
3.6. Soil Chemistry
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Elumalai, L.K.; Rengasamy, R. Synergistic effect of seaweed manure and Bacillus sp. on growth and biochemical constituents of Vigna radiata L. J. Biofertil. Biopestici. 2012, 3, 121–128. [Google Scholar] [CrossRef]
- Lewis, J.A.; Papavizas, G.C. Biocontrol of cotton damping-off caused by Rhizoctonia solani in the field with formulations of Trichoderma spp. and Gliocladium virens. Crop Prot. 1991, 10, 396–402. [Google Scholar] [CrossRef]
- Mihajlovicć, M.; Rekanovicć, E.; Hrusticć, J.; Tanovicć, B. Methods for management of soilborne plant pathogens. Pestic. Fitomedicina 2017, 32, 9–24. [Google Scholar] [CrossRef]
- Baysal-Gurel, F.; Kabir, N. Comparative performance of fungicides and biocontrol products in suppression of Rhizoctonia root rot in viburnum. J. Plant Pathol. Microbiol. 2018, 9, 451. [Google Scholar]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for management of soilborne diseases in crop production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Mokhtar, M.M.; El-Mougy, N.S. Biocompost application for controlling soilborne plant pathogens—A Review. Int. J. Eng. Innov. Technol. 2014, 4, 61–68. [Google Scholar]
- Moralejo, E.; Perez-Sierra, A.M.; Alvarez, L.A.; Belbahri, L.; Lefort, F.; Descals, E. Multiple alien Phytophthora taxa discovered on diseased ornamental plants in Spain. Plant Pathol. 2009, 58, 100–110. [Google Scholar] [CrossRef]
- Pane, A.; Martini, P.; Chimento, A.; Rapetti, S.; Savona, S.; Grasso, F.M.; Cacciola, S.O. Phytophthora species on ornamental plants in Italy. J. Plant Pathol. 2005, 87, 301. [Google Scholar]
- Hu, J.H.; Hong, C.X.; Stromberg, E.L.; Moorman, G.W. Mefenoxam sensitivity and fitness analysis of Phytophthora nicotianae isolates from nurseries in Virginia, USA. Plant Pathol. 2008, 57, 728–736. [Google Scholar] [CrossRef]
- Baysal-Gurel, F.; Liyanapathiranage, P.; Mullican, J. Biofumigation: Opportunities and challenges for control of soilborne diseases in nursery production. Plant Health Prog. 2018, 19, 332–337. [Google Scholar] [CrossRef]
- Gallup, C.A.; Sullivan, M.; Shew, H.D. Black Shank of tobacco. Plant Health Instr. 2006. [Google Scholar] [CrossRef]
- Hyakumachi, M.; Priyatmojo, A.; Kubota, M.; Fukui, H. New anastomosis groups, AG-T and AG-U, of binucleate Rhizoctonia spp. causing root and stem rot of cut-flower and miniature roses. Phytopathology 2005, 95, 784–792. [Google Scholar] [CrossRef] [Green Version]
- Rinehart, T.A.; Copes, W.E.; Toda, T.; Cubeta, M.A. Genetic characterization of binucleate Rhizoctonia species causing web blight on azalea in Mississippi and Alabama. Plant Dis. 2007, 91, 616–623. [Google Scholar] [CrossRef] [Green Version]
- Farr, D.F.; Bills, G.F.; Chamuris, G.P.; Rossman, A.Y. Fungi on Plant and Plant Products in the United States; American Phytopathological Society: St. Paul, MN, USA, 1995. [Google Scholar]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.C.; Gong, M.; Zang, H.B.; Hua, X.M.; Yao, J.; Pang, J.Y.; Yang, Y.H. Effect of methamidophos andurea application on microbial communities in soils as determined by microbial biomass and communitylevel physiological profiles. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2006, 41, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Brent, K.J.; Hollomon, D.W. Fungicide Resistance: The assessment of risk. In Monograph 1. FRAC; Global Crop Protection Federation: Brussels, Belgium, 1998; reprinted 2007; pp. 1–60. [Google Scholar]
- Klittich, C.J. Milestones in fungicide discovery: Chemistry that changed agriculture. Plant Health Prog. 2008. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, M.; Freilich, S. Prospects for biological soilborne disease control: Application of indigenous versus synthetic microbiomes. Phytopathology 2017, 107, 256–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.C.; Watkins, K.B.; Teasdale, J.R.; Abdul-Baki, A.A. Cover crops in sustainable food production. Food Rev. Int. 2000, 16, 121–157. [Google Scholar] [CrossRef]
- Halcomb, M. Summer Cover Crops for Nursery Row Middles. In Field Nursery Weed Control; Univ. Tenn. Coop. Ext. Serv. Handout, University of Tennessee: Knoxville, TN, USA, 2012. [Google Scholar]
- Yeager, T.H.; Bilderback, T.E.; Fare, D.; Gilliam, C.; Lea-Cox, J.D.; Niemiera, A.X.; Ruter, J.M.; Tilt, K.; Warren, S.; Whitwell, T.; et al. Best Management Practices: Guide for Producing Nursery Crops, 3rd ed.; Southern Nursery Association: Atlanta, GA, USA, 2013. [Google Scholar]
- Finney, D.M.; Buyer, J.S.; Kaye, J.P. Living cover crops have immediate impacts on soil microbial community structure and function. J. Soil Water Conserv. 2017, 72, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Couillerot, O.; Prigent-Combaret, C.; Caballero-Mellado, J.; Moënne-Loccoz, Y. Pseudomonas fluorescens and closely related fluorescent pseudomonads as biocontrol agents of soil-borne phytopathogens. Lett. Appl. Microbiol. 2009, 48, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Dawadi, S.; Baysal-Gurel, F.; Addesso, K.M.; Oliver, J.B.; Simmons, T. Impact of cover crop usage on soilborne diseases in field nursery production. Agronomy 2019, 9, 753. [Google Scholar] [CrossRef] [Green Version]
- Holmes, K.A.; Benson, D.M. Evaluation of Phytophthora parasitica var. nicotianae for biocontrol of Phytophthora parasitica on Catharanthus roseus. Plant Dis. 1994, 78, 193–199. [Google Scholar]
- Baysal−Gurel, F.; Simmons, T.; Liyanapathiranage, P.; Kabir, M.N. Evaluation of biorational products and fungicides for the control of rhizoctonia root rot of viburnum. In Plant Disease Management Report No. 11: OT003; American Phytopathological Society: St. Paul, MN, USA, 2017. [Google Scholar]
- Jeffers, S.N.; Martin, S.B. Comparison of two media selective for Phytophthora and Pythium species. Plant Dis. 1986, 70, 1038–1043. [Google Scholar] [CrossRef]
- Ferguson, A.J.; Jeffers, S.N. Detecting multiple species of Phytophthora in container mixes from ornamental crop nurseries. Plant Dis. 1999, 83, 1129–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, W.A.; Shew, H.D.; Melton, T.A. A semi-selective medium to isolate Rhizoctonia solani from soil and tissue. Plant Path. Ext., NC State Ext. 2001, 1–2. [Google Scholar]
- Das, A.K.; Nerkar, S.; Kumar, A.; Bawage, S. Detection, identification, and characterization of Phytophthora spp. infecting citrus in India. J. Plant Pathol. 2016, 98, 55–69. [Google Scholar]
- de Jesus, A.L.; Gonçalves, D.R.; Rocha, S.C.O.; Marano, A.V.; Jeronimo, G.H.; De Souza, J.I.; Boro, M.C.; Pires-Zottarelli, C.L.A. Morphological and phylogenetic analyses of three Phytopythium species (Peronosporales, Oomycota) from Brazil. Cryptogam. Mycol. 2016, 37, 117–128. [Google Scholar] [CrossRef]
- Gould, W.D.; Hagedorn, C.; Bardinelli, T.R.; Zablotowicz, R.M. New selective media for enumeration and recovery of fluorescent Pseudomonad from various habitats. Appl. Environ. Microbiol. 1985, 49, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Lazzeri, L.; Baruzzi, G.; Malaguti, L.; Antoniacci, L. Replacing methyl bromide in annual strawberry production with glucosinolate-containing green manure crops. Pest. Manage. Sci. 2003, 59, 983–990. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, T.S. Control of soilborne potato diseases using brassica green manures. Crop Prot. 2006, 26, 1067–1077. [Google Scholar] [CrossRef]
- Mutch, D.R.; Snapp, S. Cover crop choices for Michigan. Michigan State University Extension Bulletin E 2884. 2003. Available online: http://www.covercrops.msu.edu/pdf_files/ext_bul_E2884.pdf (accessed on 13 March 2020).
- Welke, S.E. The effect of compost extract on the yield of strawberries and the severity of Botrytis cinerea. J. Sustain. Agric. 2005, 25, 57–68. [Google Scholar] [CrossRef]
- Bhadoria, B.D.S. Allelopathy: A natural way towards weed management. Am. J. Exp. Agric. 2011, 1, 7–20. [Google Scholar] [CrossRef]
- Hausbeck, M.K.; Lamour, K.H. Phytophthora capsica on vegetable crops: Research progress and management challenges. Plant Dis. 2004, 88, 1292–1303. [Google Scholar] [CrossRef] [Green Version]
- Turrini, A.; Sbrana, C.; Avio, L.; Njeru, E.M.; Bocci, G.; Bàrberi, P.; Giovannetti, M. Changes in the composition of native root arbuscular mycorrhizal fungal communities during a short-term cover crop-maize succession. Biol. Fertil. Soils 2016, 53, 643–653. [Google Scholar] [CrossRef]
- Lehman, R.M.; Taheri, W.I.; Osborne, S.L.; Buyer, J.S.; Douds, D.D.J. Fall cover cropping can increase arbuscular mycorrhizae in soils supporting intensive agricultural production. Agric. Ecosyst. Environ. Appl. Soil Ecol. 2012, 61, 300–304. [Google Scholar] [CrossRef]
- Pfleger, F.L.; Linderman, R.G. Mycorrhiza and Plant Growth; The American Phytopathological Society Press: St. Paul, MN, USA, 2000; p. 360. [Google Scholar]
- Baysal-Gurel, F.; Liyanapathiranage, P. Pathogenecity of Rhizoctonia solani and Phytophthora nicotianae to Brassicaceae cover crops. Arch. Phytopathol. Plant Prot. 2019, 52, 288–302. [Google Scholar] [CrossRef]
- Grazieli, A.D.S. Studies on the Impact of cover Crops on Soybean Productivity and Root Rot Disease. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2019. [Google Scholar]
- Bailey, K.L.; Lazarovits, G. Suppressing soilborne diseases with residue management and organic amendments. Soil Tillage Res. 2003, 72, 169–180. [Google Scholar] [CrossRef]
- Blok, W.J.; Lamers, J.G.; Termorshuizen, A.J.; Bollen, G.J. Control of soilborne plant pathogens by incorporating fresh organic amendments followed by tarping. Phytopathology 2000, 90, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Bonanomi, G.; Antignani, V.; Capodilupo, M.; Scala, F. Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem. 2010, 42, 136–144. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Graber, E.R.; Elad, Y.; Frenkel, O. Biochar as a management tool for soilborne diseases affecting early stage nursery seedling production. Crop Prot. 2019, 120, 34–42. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Johnston, S.A. Ecologically based approaches to management of Phytophthora blight on bell pepper. Plant Dis. 1999, 83, 1080–1089. [Google Scholar] [CrossRef] [Green Version]
- Civitello, D.J.; Cohen, J.; Fatima, H.; Halstead, N.T.; Liriano, J.; McMahon, T.A.; Ortega, C.N.; Sauer, E.L.; Sehgal, T.; Young, S.; et al. Biodiversity inhibits parasites: Broad evidence for the dilution effect. Proc. Natl. Acad. Sci. USA 2015, 112, 8667–8671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keesing, F.; Belden, L.K.; Daszak, P.; Dobson, A.; Harvell, C.D.; Holt, R.D.; Hudson, P.; Jolles, A.; Jones, K.E.; Mitchell, C.E.; et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature 2010, 468, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Vukicevich, E.; Lowery, T.; Bowen, P.; Urbez-Torres, J.R.; Hart, M. Cover crops to increase soil microbial diversity and mitigate decline in perennial agriculture. A review. Agron. Sustain. Dev. 2016, 36, 48. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Lee-Marzano, S.; Ortiz-Ribbing, L.M.; Gruver, J.; Hartman, G.L.; Eastburn, D.M. Suppression of soilborne diseases of soybean with cover crops. Plant Dis. 2017, 101, 1918–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, R.P.; Griffin, T.S.; Honeycutt, C.W. Rotation and cover crop effects on soilborne potato diseases, tuber yield, and soil microbial communities. Plant Dis. 2010, 94, 1491–1502. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.F.; Yamasaki, H.; Mazzola, M. Brassica napus seed meal soil amendment modifies microbial community structure, nitric oxide production and incidence of Rhizoctonia root rot. Soil Biol. Biochem. 2005, 37, 1215–1227. [Google Scholar] [CrossRef]
- Motisi, N.; Montfort, F.; Faloya, V.; Lucas, P.; Dore, T. Growing Brassica juncea as a cover crop, then incorporating its residues provides complementary control of Rhizoctonia root rot of sugar beet. Field Crop Res. 2009, 113, 238–245. [Google Scholar] [CrossRef]
- Baysal-Gurel, F.; Liyanapathiranage, P.; Addesso, M. Effect of Brassica crop-based biofumigation on soilborne disease suppression in woody ornamentals. Can. J. Plant Pathol. 2019, 8, 138. [Google Scholar] [CrossRef]
- Huber, D.M.; Graham, R.D. The role of nutrition in crop resistance and tolerance to diseases. In Mineral Nutrition of Crops: Fundamental Mechanisms and Implications; Rengel, Z., Ed.; Food Products Press: New York, NY, USA, 1999; pp. 169–206. [Google Scholar]
- Sanford, G.B. Studies on Rhizoctonia solani kühn.: IV. Effect of soil temperature and moisture on virulence. Can. J. Res. 1938, 16, 203–213. [Google Scholar] [CrossRef]
- Schlatter, D.; Kinkel, L.L.; Thomashow, L.S.; Weller, D.M.; Paulitz, T.C. Disease suppressive soils: New insights from the soil microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, D.M.; Raaijmakers, J.M.; McSpadden Gardener, B.B.; Thomashow, L.S. Microbial populations responsible for specific suppressiveness to plant pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, D.M.; Landa, B.B.; Mavrodi, O.V.; Schroeder, K.L.; De La Fuente, L.; Blouin Bankhead, S.; Allende Molar, R.; Bonsall, R.F.; Mavrodi, D.V.; Thomashow, L.S. Role of 2,4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. in the defense of plant roots. Plant Biol. 2007, 9, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramamoorthy, V.; Samiyappan, R. Induction of defense-related genes in Pseudomonas fluorescens treated chilli plants in response to infection by Colletotrichum capsici. J. Mycol. Plant Pathol. 2001, 31, 146–155. [Google Scholar]
- Mathivanan, N.; Prabavathy, V.R.; Vijayanandraj, V.R. Application of talc formulations of Pseudomonas fluorescens Migula and Trichoderma viridae Pers. ex SF Gray decrease the sheath blight disease and enhance the plant growth and yield in rice. J. Phytopathol. 2005, 153, 697–701. [Google Scholar] [CrossRef]
- Marimuthu, S.; Ramamoorthy, V.; Samiyappan, R.; Subbian, P. Intercropping system with combined application of Azospirillum and Pseudomonas fluorescens reduces root rot incidence caused by Rhizoctonia bataticola and increases seed cotton yield. J. Phytopathol. 2013, 161, 405–411. [Google Scholar] [CrossRef]
- Goswami, D.; Vaghela, H.; Parmar, S.; Dhandhukia, P.C.; Thakker, J.N. Plant growth promoting potentials of Pseudomonas spp. strain OG isolated from marine water. J. Plant Interact. 2013, 8, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Barahona, E.; Navazo, A.; Martinez-Granero, F.; Zea-Bonilla, T.; Perez-Jimenez, R.M.; Martin, M.; Rivilla, R. Pseudomonas fluorescens F113 mutant with enhanced competitive colonization ability and improved biocontrol activity against fungal root pathogens. Appl. Environ. Microbiol. 2011, 77, 5412–5419. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, J. Use of cover crops and green manures to attract beneficial insects. In Integrated Pest Management Program; University of Connecticut: Storrs, CT, USA, 1999; Retrieved: 20 May 2020. [Google Scholar]
- Bakker, M.G.; Acharya, J.; Moorman, T.B.; Robertson, A.E.; Kaspar, T.C. The potential for cereal rye cover crops to host corn seedling pathogens. Phytopathology 2016, 106, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Robertson, A.; Kaspar, T.; Leandro, L.; Mueller, D.; Acharya, J. Disease risks associated with cover crops and soybean production. In Proceedings of the Integrated Crop Management Conference, Ames, IA, USA, 29–30 November 2017; 21, pp. 125–128. Available online: https://lib.dr.iastate.edu/icm/2017/proceedings/21 (accessed on 20 April 2020).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Production Site 1 | Production Site 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Rhizoctonia solani Recovery (%) z | Oomycete Recovery (%) z | Total Height Increment (cm) y | Total Plant Fresh wt. (g) | Total Root Fresh wt. (g) | Rhizoctonia solani Recovery (%) z | Oomycete Recovery (%) z | Total Height Increment (cm) y | Total Plant Fresh wt. (g) | Total Root Fresh wt. (g) | |
| Crimson Clover | 12.0 ± 3.3 b x | 24.0 ± 2.7 ab | 9.3 ± 1.8 a | 47.3 ± 4.3 a | 28.6 ± 2.7 a | 16.0 ± 2.7 ab | 16.0 ± 2.7 b | 9.6 ± 2.9 a | 48.9 ± 3.9 a | 33.2 ± 2.7 a |
| Triticale | 10.0 ± 4.5 b | 18.0 ± 3.6 b | 14.3 ± 3.0 a | 51.6 ± 6.4 a | 34.2 ± 5.1 a | 14.0 ± 4.3 b | 16.0 ± 4.0 b | 4.0 ± 1.3 a | 58.7 ± 5.3 a | 41.3 ± 3.7 a |
| Control | 26.0 ± 3.1 a | 34.0 ± 3.1 a | 8.1 ± 2.0 a | 64.5 ± 9.5 a | 43.5 ± 7.8 a | 28.0 ± 3.3 a | 36.0 ± 2.7 a | 10.8 ± 2.6 a | 46.7 ± 4.8 a | 32.9 ± 3.9 a |
| F value | 5.70 | 6.68 | 1.97 | 1.62 | 1.79 | 4.78 | 13.24 | 2.37 | 1.49 | 1.83 |
| df | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| P value | 0.0086 | 0.0044 | 0.1585 | 0.2169 | 0.1870 | 0.0167 | <0.001 | 0.1128 | 0.2441 | 0.1796 |
| Treatments | Production Site 1 | Production Site 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| Pathogen Recovery of P. vexans z | Total Height Increment (cm) y | Total Plant Fresh wt. (g) | Total Root Fresh wt. (g) | Pathogen Recovery of P. vexans z | Total Height Increment (cm) y | Total Plant Fresh wt. (g) | Total Root Fresh wt. (g) | |
| Crimson Clover | 38.0 ± 4.7 b x | 10.5 ± 1.0 a | 45.5 ± 4.1 a | 29.4 ± 2.7 a | 28.0 ± 3.3 b | 8.1 ± 1.6 a | 59.5 ± 3.6 a | 36.4 ± 3.3 a |
| Triticale | 32.0 ± 4.4 b | 6.8 ± 1.4 ab | 48.6 ± 4.9 a | 31.4 ± 3.1 a | 30.0 ± 4.5 b | 4.7 ± 2.3 a | 69.5 ± 4.0 a | 49.6 ± 3.7 a |
| Control | 64.0 ± 5.8 a | 5.3 ± 1.0 b | 51.7 ± 3.7 a | 33.6 ± 2.7 a | 74.0 ± 4.3 a | 5.6 ± 2.0 a | 57.2 ± 5.3 a | 39.5 ± 4.3 a |
| F value | 11.56 | 5.55 | 0.52 | 0.55 | 41.48 | 0.78 | 2.25 | 3.34 |
| df | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| P value | 0.0002 | 0.0095 | 0.5992 | 0.5824 | <0.001 | 0.4688 | 0.1243 | 0.0507 |
| Treatments | Production Site 1 | Production Site 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| Pathogen Recovery of P. nicotianae z | Total Height Increment (cm) y | Total Plant Fresh wt. (g) | Total Root Fresh wt. (g) | Pathogen Recovery of P. nicotianae z | Total Height Increment (cm) y | Total Plant Fresh wt. (g) | Total Root Fresh wt. (g) | |
| Crimson Clover | 48.0 ± 4.4 b x | 12.0 ± 1.5 a | 51.1 ± 5.0 a | 35.2 ± 3.7 a | 24.0 ± 4.0 b | 5.7 ± 1.8 a | 56.6 ± 4.0 a | 40.6 ± 3.1 a |
| Triticale | 36.0 ± 2.7 b | 8.1 ± 2.4 ab | 59.7 ± 3.3 a | 38.5 ± 2.3 a | 30.0 ± 4.5 b | 3.8 ± 1.9 a | 46.4 ± 5.2 a | 32.4 ± 3.8 a |
| Control | 66.0 ± 5.2 a | 4.9 ± 0.9 b | 62.2 ± 5.1 a | 41.4 ± 3.4 a | 70.0 ± 5.4 a | 5.9 ± 1.9 a | 55.3 ± 5.4 a | 38.5 ± 4.1 a |
| F value | 12.72 | 4.18 | 1.65 | 0.96 | 28.91 | 0.39 | 1.30 | 1.32 |
| df | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| P value | 0.0001 | 0.0261 | 0.2106 | 0.3944 | <0.001 | 0.681 | 0.289 | 0.2851 |
| Treatments | Production Site 1 | Production Site 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| Pathogen Recovery of R. solani z | Total Height Increment (cm) y | Total Plant Fresh wt. (g) | Total Root Fresh wt. (g) | Pathogen Recovery of R. solani z | Total Height Increment (cm) y | Total Plant Fresh wt. (g) | Total Root Fresh wt. (g) | |
| Crimson Clover | 52.0 ± 4.4 bx | 11.4 ± 2.1 a | 41.9 ± 4.6 a | 29.7 ± 1.9 a | 32.0 ± 3.3 b | 5.1 ± 1.4 a | 62.a ± 6.2 a | 44.7 ± 4.6 a |
| Triticale | 28.0 ± 3.3 c | 6.6 ± 1.6 ab | 51.1 ± 3.5 a | 32.6 ± 2.3 a | 36.0 ± 4.0 b | 2.3 ± 0.5 a | 57.8 ± 2.9 a | 41.0 ± 2.3 a |
| Control | 76.0 ± 5.8 a | 4.0 ± 1.1 b | 51.9 ± 3.7 a | 34.3 ± 2.8 a | 70.0 ± 3.3 a | 6.7 ± 2.5 a | 59.0 ± 6.3 a | 40.9 ± 5.2 a |
| F value | 27.00 | 5.35 | 1.98 | 2.73 | 34.62 | 1.73 | 0.17 | 0.26 |
| df | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| P value | <.0001 | 0.0110 | 0.1575 | 0.0834 | <0.001 | 0.1967 | 0.8476 | 0.7705 |
| Treatments | Production Site 1 | Production Site 2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OM y | CEC y (meq/100 g) | C:N Ratio y | pH | P (kg/ha) | K (kg/ha) | OM y | CEC y (meq/100g) | C:N Ratio y | pH | P (kg/ha) | K (kg/ha) | |
| Crimson Clover | 2.9 a z | 5.9 a | 8.5 a | 5.9 a | 41.4 a | 411.6 a | 2.6 a | 5.8 a | 9.3 ab | 6.0 a | 27.4 a | 242.5 a |
| Triticale | 2.9 a | 6.6 a | 11.5 a | 5.9 a | 31.4 a | 320.9 a | 2.5 a | 6.0 a | 11.9 a | 5.9 a | 29.7 a | 241.4 a |
| Control | 2.9 a | 6.3 a | 9.1 a | 5.9 a | 36.9 a | 371.8 a | 2.8 a | 5.3 a | 8.2 b | 5.9 a | 22.4 a | 201.6 a |
| P value | 1.0000 | 0.7103 | 0.2391 | 0.9866 | 0.5928 | 0.7132 | 0.2933 | 0.3939 | 0.0401 | 0.5413 | 0.5368 | 0.7016 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panth, M.; Baysal-Gurel, F.; Simmons, T.; Addesso, K.M.; Witcher, A. Impact of Winter Cover Crop Usage in Soilborne Disease Suppressiveness in Woody Ornamental Production System. Agronomy 2020, 10, 995. https://doi.org/10.3390/agronomy10070995
Panth M, Baysal-Gurel F, Simmons T, Addesso KM, Witcher A. Impact of Winter Cover Crop Usage in Soilborne Disease Suppressiveness in Woody Ornamental Production System. Agronomy. 2020; 10(7):995. https://doi.org/10.3390/agronomy10070995
Chicago/Turabian StylePanth, Milan, Fulya Baysal-Gurel, Terri Simmons, Karla M. Addesso, and Anthony Witcher. 2020. "Impact of Winter Cover Crop Usage in Soilborne Disease Suppressiveness in Woody Ornamental Production System" Agronomy 10, no. 7: 995. https://doi.org/10.3390/agronomy10070995