Expression of Specific Class I Chitinase mRNA Levels in Different Grape Varieties and Their Antimicrobial Activity

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. RNA Extraction and cDNA Synthesis
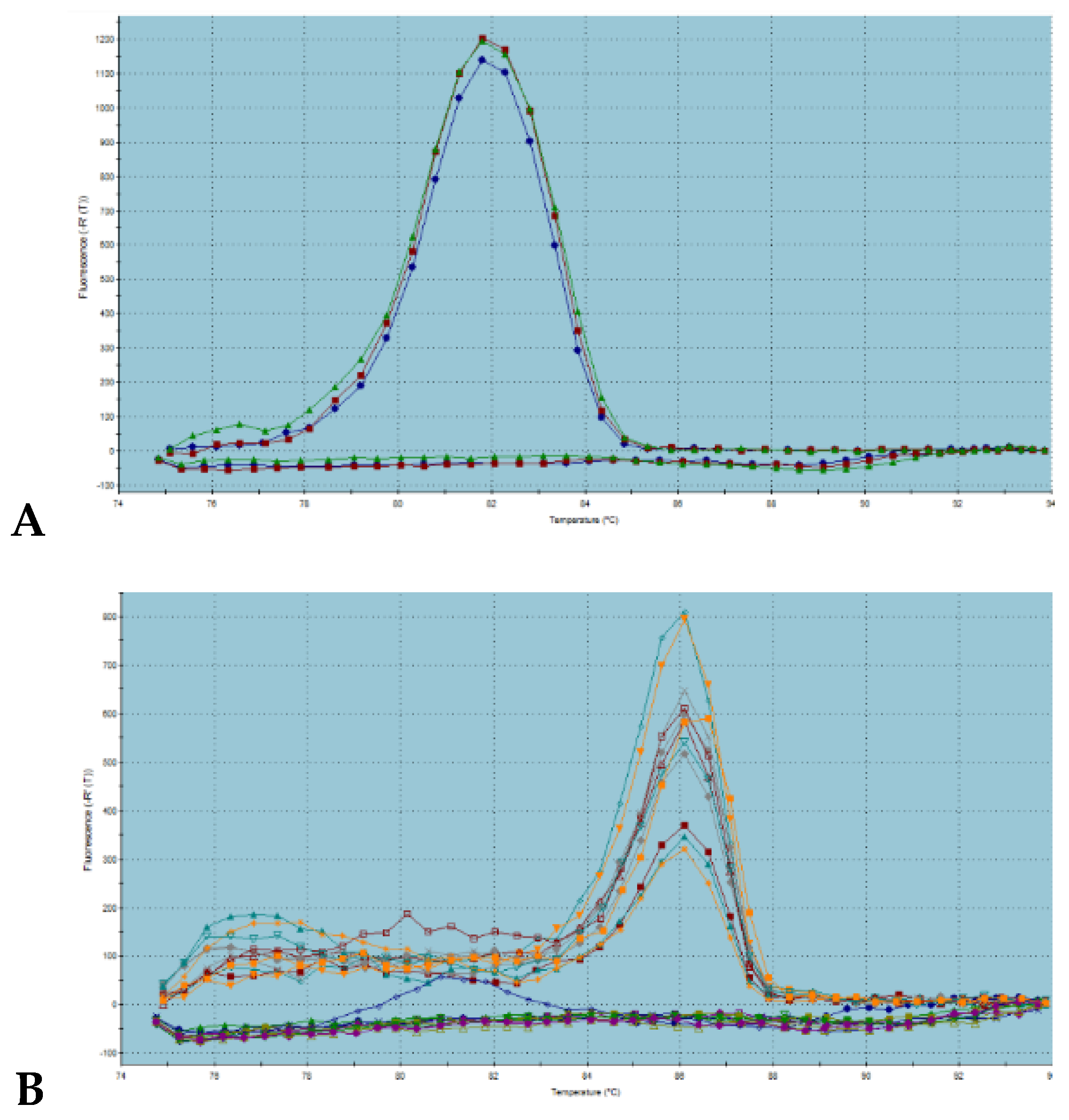
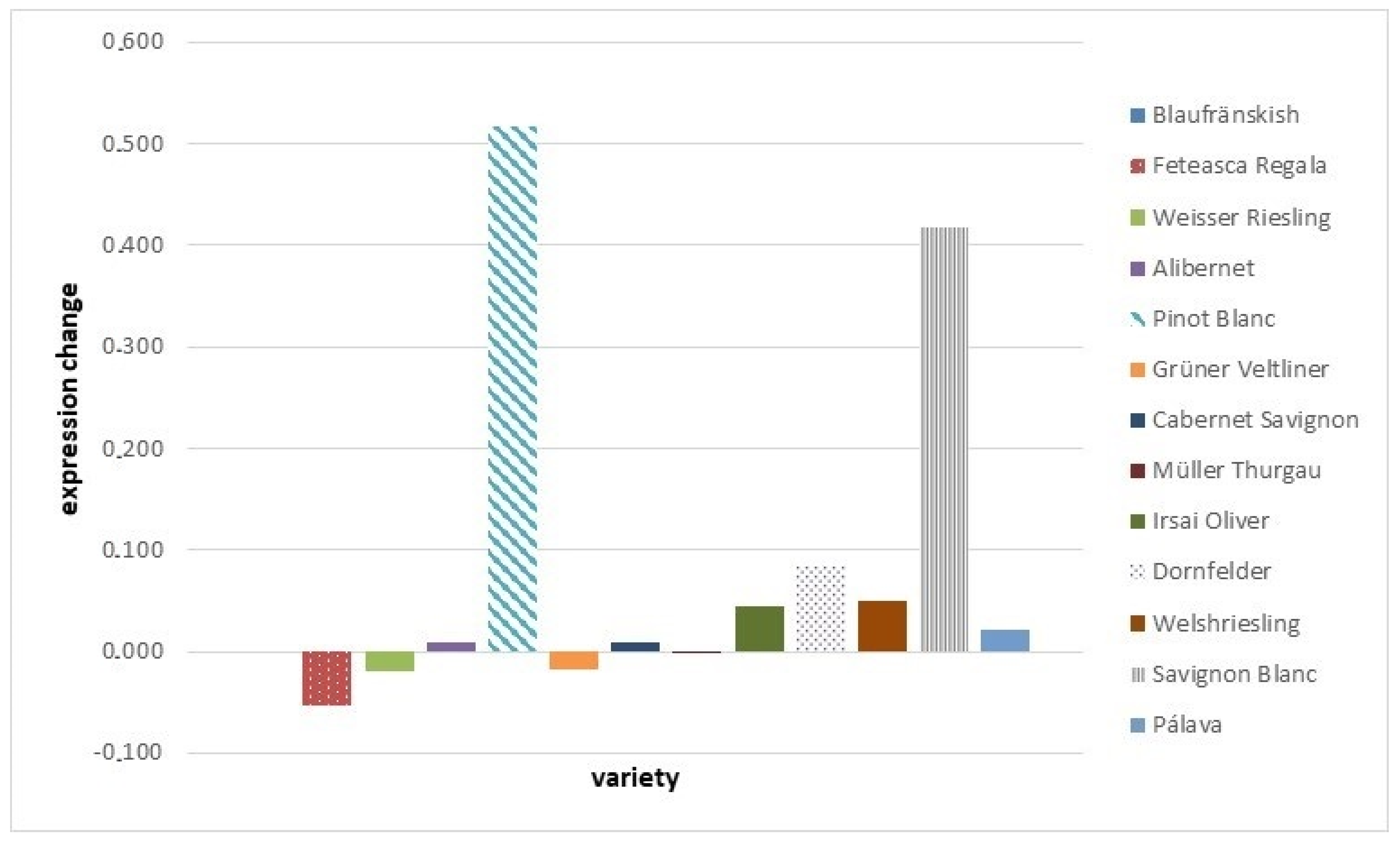
2.3. Chitinase Expression Analysis
2.4. Pomace Extract Preparation
2.5. Microorganisms
2.6. Disc Diffusion Method
2.7. Determination of Minimum Inhibitory Concentration
2.8. Data Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pastorello, E.A.; Farioli, L.; Pravettoni, V.; Ortolani, C.; Fortunato, D.; Giuffrida, M.G.; Garoffo, L.P.; Calamari, A.M.; Brenna, O.; Conti, A. Identification of grape and wine allergens as an Endochitinase 4, a lipid-transfer protein, and a Thaumatin. J. Allergy Clin. Immunol. 2003, 111, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Soural, I.; Balik, J.; Vrchotová, N.; Triska, J. Varietal Distributions of Stilbenes in Grape Cane of Vitis vinifera L. Acta Hortic. Regiotect. 2017, 20, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Arora, P.; Ansari, S.H.; Najmi, A.K.; Anjum, V.; Ahmad, S. Investigation of anthi-asthmatic potential of dried fruits of Vitis vinifera L. in animal model of bronchial asthma. Allerg. Asthma Clin. Immunol. 2016, 12, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, F.; Khan, G.M. Study of Aging and Hepatoprotective Activity of Vitis vinifera L. Seeds in Albino Rats. Asian Pac. J. Trop. Biomed. 2012, 2, 1770–1774. [Google Scholar] [CrossRef]
- Masani, Y.A.; Mathew, N.; Chakraborty, M.; Kamath, J.V. Effects of Vitis vinifera against Trition X 100 induced hyperlipidaemia in rats. Int. Res. J. of Pharm. 2012, 3, 101–103. [Google Scholar]
- Glasa, M.; Predajňa, L.; Sihelská, N.; Šoltys, K.; Ruiz-García, A.-B. Analysis of Virome by High-Throughput Sequencing Revealed Multiple Infection and Intra-Virus Diversity in a Single Grapevine Plant. Acta Hortic. Regiotect. 2020, 23, 35–39. [Google Scholar] [CrossRef]
- Iandolino, A.; Nobuta, K.; da Silva, F.G.; Cook, D.R.; Meyers, B.C. Comparative expression profiling in grape (Vitis vinifera) berries derived from frequency analysis of ESts and MPSS signatures. BMC Plant Biol. 2008, 8, 53. [Google Scholar] [CrossRef] [Green Version]
- Romero, I.; Sanchez-Ballesta, M.T.; Maldonado, R.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Expression of class I chitinase and β-1,3-glucanase genes and postharvest fungal decay control of table grapes by high CO2 pretreatment. Postharvest Boil. Technol. 2006, 41, 9–15. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.S.; Osburn, R.M. Combined Used of Lipo-Chitooligosaccharides and Chitinous Compounds for Enhanced Plant Growth and Yield. U.S. Patent 9,253,989 B2, 9 February 2016. [Google Scholar]
- Collinge, D.B.; Kragh, K.M.; Mikkelsen, J.D.; Nielsen, K.K.; Rasmussen, U.; Vad1, K. Plant chitinases. Plant J. 1993, 3, 31–40. [Google Scholar] [CrossRef]
- Kumar, M.; Brar, A.; Yadav, M.; Chawade, A.; Vivekanand, V.; Pareek, N. Chitinases—Potential Candidates for Enhanced Plant Resistance towards Fungal Pathogens. Agriculture 2018, 8, 88. [Google Scholar] [CrossRef] [Green Version]
- Malik, A. Preety Purification and properties of plant chitinases: A review. J. Food Biochem. 2019, 43, e12762. [Google Scholar] [CrossRef]
- Derckel, J.-P.; Legendre, L.; Audran, J.-C.; Haye, B.; Lambert, B. Chitinases of the grapevine (Vitis vinifera L.): Five isoforms induced in leaves by salicylic acid are constitutively expressed in other tissues. Plant Sci. 1996, 119, 31–37. [Google Scholar] [CrossRef]
- Ano, A.; Takayanagi, T.; Uchibori, T.; Okuda, T.; Yokotsuka, K. Characterization of a class III chitinase from Vitis vinifera cv. Koshu. J. Biosci. Bioeng. 2003, 95, 645–647. [Google Scholar] [CrossRef]
- Robinson, S.P.; Jacobs, A.K.; Dry, I.B. A class IV chitinase is highly expressed in grape berries during ripening. Plant. Physiol. 1997, 114, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Vincenzi, S.; Bierma, J.; Wickramasekara, S.I.; Curioni, A.; Gazzola, D.; Bakalinsky, A. Characterization of a Grape Class IV Chitinase. J. Agric. Food Chem. 2014, 62, 5660–5668. [Google Scholar] [CrossRef]
- Farshidi, N.; Moghaddam, M.; Yaghoubi, V.; Ayati, S.H.; Varasteh, A.R.; Sankian, M. Type IV chitinase quantification in four different grape cultivars (Vitis vinifera) in northeast of Iran by an indirect sandwich enzyme-linked immunosorbent assay. J. Immunoass. Immunochem. 2018, 40, 139–148. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Couderchet, M.; Vernet, G.; Aziz, A. Chitosan Stimulates Defense Reactions in Grapevine Leaves and Inhibits Development of Botrytis Cinerea. Eur. J. Plant Pathol. 2006, 114, 405–413. [Google Scholar] [CrossRef]
- Saito, S.; Odagiri, M.; Furuya, S.; Suzuki, S.; Takayanagi, T. Inhibitory effect of chitinases isolated from Semillon grapes (Vitis vinifera) on growth of grapevine pathogens. J. Plant Biochem. Biotechnol. 2011, 20, 47–54. [Google Scholar] [CrossRef]
- Punja, Z.K.; Zhang, Y.-Y. Plant Chitinases and Their Roles in Resistance to Fungal Diseases. J. Nematol. 1993, 25, 526–540. [Google Scholar]
- Falconer, R.; Marangon, M.; Van Sluyter, S.C.; Neilson, K.A.; Chan, C.; Waters, E.J. Thermal Stability of Thaumatin-Like Protein, Chitinase, and Invertase Isolated from Sauvignon blanc and Semillon Juice and Their Role in Haze Formation in Wine. J. Agric. Food Chem. 2010, 58, 975–980. [Google Scholar] [CrossRef]
- Marangon, M.; Van Sluyter, S.C.; Neilson, K.A.; Chan, C.; Haynes, P.A.; Waters, E.J.; Falconer, R. Roles of Grape Thaumatin-like Protein and Chitinase in White Wine Haze Formation. J. Agric. Food Chem. 2011, 59, 733–740. [Google Scholar] [CrossRef]
- Ferreira, R.B.; A Piçarra-Pereira, M.; Monteiro, S.A.; Loureiro, V.B.; Teixeira, A.R. The wine proteins. Trends Food Sci. Technol. 2001, 12, 230–239. [Google Scholar] [CrossRef]
- Van Sluyter, S.C.; McRae, J.; Falconer, R.; Smith, P.; Bacic, A.; Waters, E.J.; Marangon, M. Wine Protein Haze: Mechanisms of Formation and Advances in Prevention. J. Agric. Food Chem. 2015, 63, 4020–4030. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.; Alexander, G.; Muhlack, R.; Pocock, K.; Colby, C.; O’Neill, B.; Høj, P.; Jones, P. Preventing protein haze in bottled white wine. Aust. J. Grape Wine Res. 2005, 11, 215–225. [Google Scholar] [CrossRef]
- Kunová, S.; Felšöciová, S.; Tvrda, E.; Ivanišová, E.; Kántor, A.; Žiarovská, J.; Terentjeva, M.; Kačániová, M. Antimicrobial activity of resveratrol and grape pomace extract. Potravinarstvo Slovak J. Food Sci. 2019, 13, 363–368. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Smyth, G.K. Linear Models and Empirical Bayes Methods for Assessing Differential Expression in Microarray Experiments. Stat. Appl. Genet. Mol. Boil. 2004, 3, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Social Science Statistics. T-Test Calculator for 2 Independent Means. 2019. Available online: www.socscistatistics.com/tests/studentttest (accessed on 10 June 2020).
- Rhodes, P.; Mitchell, J.; Wilson, M.; Melton, L. Antilisterial activity of grape juice and grape extracts derived from Vitis vinifera variety Ribier. Int. J. Food Microbiol. 2006, 107, 281–286. [Google Scholar] [CrossRef] [PubMed]
- De Medeiros, S.C.; Júnior, J.E.M.; Sales, G.W.P.; Grangeiro, T.B.; Nogueira, N.A.P. Chitinases as Antibacterial Proteins: A Systematic Review. J. Young- Pharm. 2018, 10, 144–148. [Google Scholar] [CrossRef]
- Chen, Y.-T.; Hsu, L.-H.; Huang, I.-P.; Tsai, T.-C.; Lee, G.-C.; Shaw, J.-F. Gene Cloning and Characterization of a Novel Recombinant Antifungal Chitinase from Papaya (Carica papaya). J. Agric. Food Chem. 2007, 55, 714–722. [Google Scholar] [CrossRef]
- Wang, S.; Shao, B.; Rao, P.; Lee, Y.; Ye, X. Hypotin, a Novel Antipathogenic and Antiproliferative Protein from Peanuts with a Sequence Similar to Those of Chitinase Precursors. J. Agric. Food Chem. 2007, 55, 9792–9799. [Google Scholar] [CrossRef] [PubMed]
- Van Scheltinga, A.C.T.; Hennig, M.; Dijkstra, B.W. The 1.8 Å Resolution Structure of Hevamine, a Plant Chitinase/Lysozyme, and Analysis of the Conserved Sequence and Structure Motifs of Glycosyl Hydrolase Family 18. J. Mol. Biol. 1996, 262, 243–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippi, A.; Petrussa, E.; Boscutti, F.; Vuerich, M.; Vrhovsek, U.; Rabiei, Z.; Braidot, E. Bioactive Polyphenols Modulate Enzymes Involved in Grapevine Pathogenesis and Chitinase Activity at Increasing Complexity Levels. Int. J. Mol. Sci. 2019, 20, 6357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, B.J.; Kwak, J.-H.; Son, J.-H.; Park, J.-M.; Lee, J.-Y.; Jo, C.; Byun, M.-W. Biological and anti-microbial activity of irradiated green tea polyphenols. Food Chem. 2004, 88, 549–555. [Google Scholar] [CrossRef]
- Grover, A.; Yashpal; Sen, A.; Arora, N.; Kirti, P.B.; Sharma, R.P. Removal of Vacuolar Targeting Signal from Class I Vacuolar Chitinase leads to its Extracellular Secretion in Transgenic Tobacco. J. Plant Biochem. Biotechnol. 2001, 10, 139–142. [Google Scholar] [CrossRef]
- Ruggiero, P.; Rossi, G.; Tombola, F.; Pancotto, L.; Lauretti, L.; Del Giudice, G.; Zoratti, M. Red wine and green tea reduce H pylori- or VacA-induced gastritis in a mouse model. World J. Gastroenterol. 2007, 13, 349. [Google Scholar] [CrossRef] [Green Version]
- Ja Jayaprakasha, G.; Selvi, T.; Sakariah, K. Antibacterial and antioxidant activities of grape (Vitis vinifera) seed extracts. Food Res. Int. 2003, 36, 117–122. [Google Scholar] [CrossRef]
- Simonetti, G.; Santamaria, A.R.; D’Auria, F.; Mulinacci, N.; Innocenti, M.; Cecchini, F.; Pericolini, E.; Gabrielli, E.; Panella, S.; Antonacci, D.; et al. Evaluation of Anti-Candida Activity of Vitis vinifera L. Seed Extracts Obtained from Wine and Table Cultivars. BioMed Res. Int. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pomace Sample | EC | PA | SE | EF | LM | SA | CA | CG | CT |
|---|---|---|---|---|---|---|---|---|---|
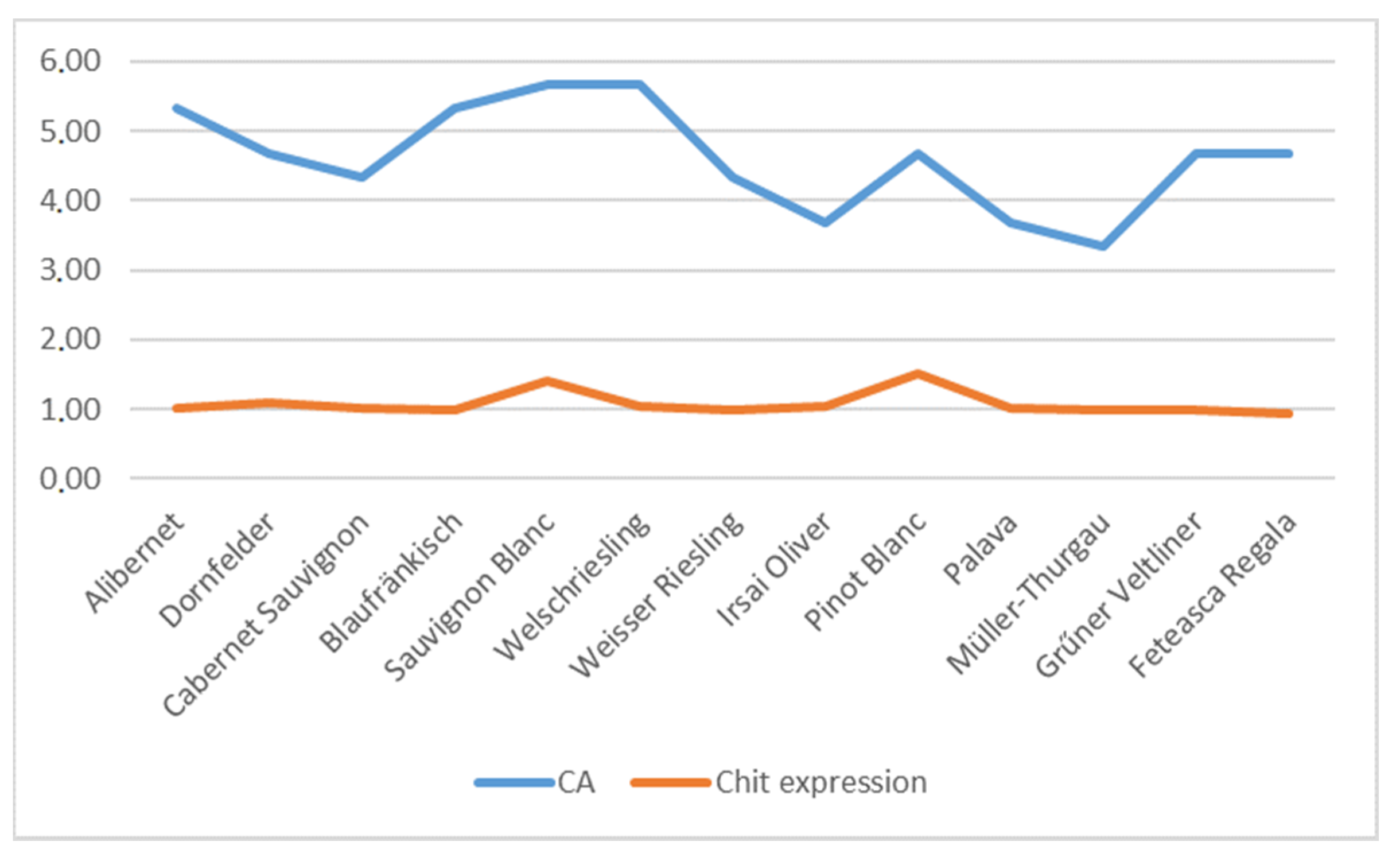
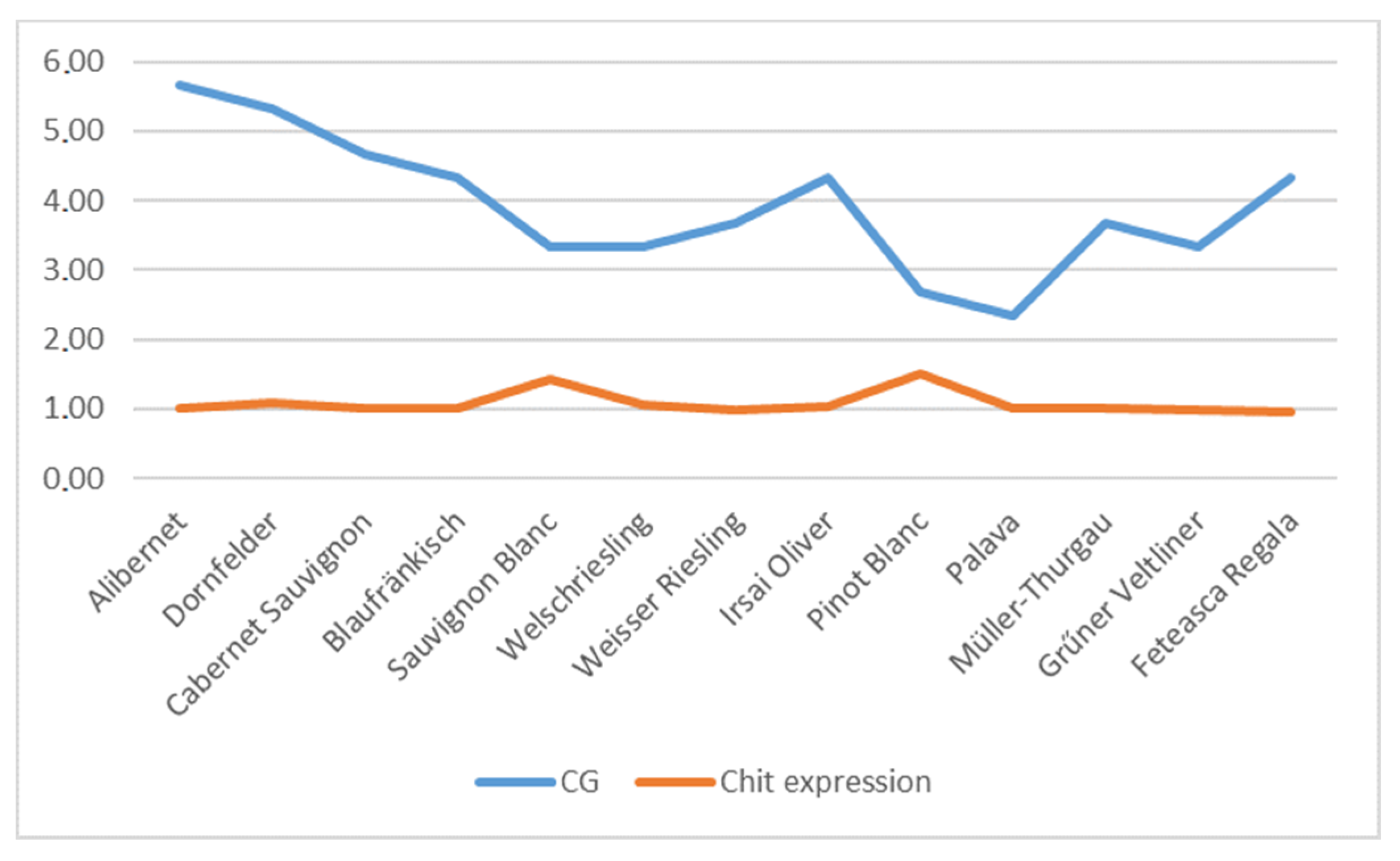
| Alibernet | 8.33 ± 0.58 a | 8.33 ± 1.15 b | 6.33 ± 0.58 c | 8.33 ± 1.15 d | 10.33 ± 0.58 c,e | 7.67 ± 0.58 e | 5.33 ± 0.58 a,b,d,e | 5.66 ± 0.58 a,b,d,e | 4.33 ± 0.58 a,b,d,e |
| Dornfelder | 9.33 ± 0.58 a | 8.00 ± 1.73 b | 6.33 ± 1.15 a | 8.67 ± 1.15 d | 8.67 ± 0.58 e | 7.33 ± 0.58 f | 4.67 ± 0.58 a,b,d,e,f | 5.33 ± 0.58 a,b,d,e | 6.67 ± 0.58 a |
| Cabernet Sauvignon | 7.33 ± 0.58 a | 6.33 ± 0.58 b | 5.67 ± 0.58 a,c | 8.67 ± 0.58 b,c,d | 7.67 ± 0.58 c,e | 6.67 ± 0.58 d,f | 4.33 ± 0.58 a,b,d,e,f | 4.67 ± 0.58 a,b,d,e,f | 3.67 ± 0.58 a,b,c,d,e,f |
| Blaufränkisch | 8.33 ± 1.15 a | 6.67 ± 1.15 b | 5.33 ± 0.58 a,c | 8.67 ± 1.15 c, d | 9.33 ± 0.58 b, c, e | 7.33 ± 0.58 f | 5.33 ± 0.58 a,d,e | 4.33 ± 0.58 a,d,e,f | 4.67 ± 1.15 a,d,e,f |
| Sauvignon Blanc | 6.33 ± 0.58 a | 7.67 ± 1.53 b | 6.33 ± 1.15 c | 8.67 ± 1.15 d | 8.67 ± 0.58 e | 6.67 ± 0.58 f | 5.67 ± 1.15 d,e | 3.33 ± 0.58 a,b,c,d,e,f | 4.33 ± 0.58 b,d,e |
| Welschriesling | 5.67 ± 0.58 a | 6.00 ± 1.00 b | 5.67 ± 1.15 c | 8.67 ± 0.58 a,b,c,d | 7.33 ± 0.58 e | 6.33 ± 1.15 d,f | 5.67 ± 0.58 d,g | 3.33 ± 0.58 a,b,c,d,e,f,g | 2.67 ± 0.58 a,b,c,d,e,f,g |
| Weisser Riesling | 4.67 ± 0.58 a | 4.67 ± 0.58 a,b | 5.33 ± 0.58 c | 9.67 ± 0.58 a,b,c | 6.67 ± 0.58 a,e | 7.67 ± 0.58 a,c,e,f | 4.33 ± 0.58 b,e,f | 3.67 ± 0.58 b,c,e,f | 3.33 ± 0.58 b,c,e,f |
| Irsai Oliver | 6.33 ± 0.58 a | 5.67 ± 0.58 b | 6.00 ± 1.00 c | 8.67 ± 0.58 a,b,c,d | 7.67 ± 0.58 e | 7.33 ± 1.15 e | 3.67 ± 0.58 a,c,d,e,f | 4.33 ± 0.58 d,e,f | 3.33 ± 0.58 a,b,c,d,e,f |
| Pinot Blanc | 5.33 ± 0.58 a | 4.33 ± 0.58 b | 4.67 ± 0.58 c | 7.67 ± 0.58 a,b,c,d | 6.33 ± 0.58 b,e | 8.33 ± 0.58 a,b,c,e,f | 4.67 ± 0.58 d,e,f,g | 2.67 ± 1.15 a,c,d,e,f,g | 3.33 ± 0.58 a,d,e,f |
| Pálava | 8.67 ± 0.58 a | 8.33 ± 1.15 b | 5.33 ± 1.53 a,b,c | 7.67 ± 0.58 d | 6.67 ± 0.58 e | 7.33 ± 0.58 f | 3.67 ± 0.58 a,b,d,e,f | 2.33 ± 0.58 a,b,c,d,e,f | 3.33 ± 0.58 a,b,d,e,f |
| Müller-Thurgau | 7.67 ± 0.58 a | 6.33 ± 0.58 b | 4.33 ± 0.58 a,c | 7.67 ± 0.58 c,d | 5.67 ± 1.15 e | 6.33 ± 0.58 f | 3.33 ± 1.15 a,b,d,f | 3.67 ± 1.15 a,b,d,f | 2.33 ± 0.58 a,b,d,e,f |
| Grűner Veltliner | 5.67 ± 0.58 a | 6.33 ± 0.58 b | 4.33 ± 0.58 b,c | 8.00 ± 1.00 a,c,d | 6.33 ± 0.58 c,e | 7.33 ± 0.58 c,f | 4.67 ± 0.58 d,f,g | 3.33 ± 0.58 a,b,d,e,f | 2.67 ± 0.58 a,b,d,e,f,g |
| Feteasca Regala | 6.33 ± 1.15 a | 5.67 ± 0.58 b | 4.33 ± 0.58 c | 8.67 ± 0.58 b,c,d | 7.67 ± 1.15 c,e | 7.00 ± 1.00 c,f | 4.67 ± 1.15 d,e | 4.33 ± 0.58 d,e,f | 3.67 ± 0.58 a,d,e,f |
| Source of Variation | SS | df | MS | F | p-Value | F Crit |
|---|---|---|---|---|---|---|
| Sample | 138.4729 | 12 | 11.53941 | 19.19589 | 9.78321E-29 | 1.793719 |
| Columns | 896.7407 | 8 | 112.0926 | 186.4668 | 5.40308E-97 | 1.978112 |
| Interaction | 196.2963 | 96 | 2.044753 | 3.401461 | 1.70267E-14 | 1.314387 |
| Within | 140.6667 | 234 | 0.60114 | |||
| Total | 1372.177 | 350 |
| Grape Sample | EC | PA | SE | EF | LM | SA | CA | CG | CT |
|---|---|---|---|---|---|---|---|---|---|
| Alibernet | 500 | 1000 | 1000 | 500 | 250 | 125 | 1000 | 1000 | 1000 |
| Dornfelder | 500 | 500 | 1000 | 500 | 250 | 250 | 1000 | 2000 | 500 |
| Cabernet Sauvignon | 250 | 500 | 500 | 500 | 500 | 250 | 1000 | 2000 | 500 |
| Blaufränkisch | 500 | 1000 | 500 | 500 | 1000 | 125 | 2000 | 1000 | 500 |
| Sauvignon Blanc | 500 | 1000 | 1000 | 1000 | 500 | 500 | 2000 | 2000 | 2000 |
| Welschriesling | 1000 | 1000 | 1000 | 1000 | 250 | 500 | 1000 | 2000 | 1000 |
| Weisser Riesling | 1000 | 500 | 1000 | 1000 | 500 | 500 | 2000 | 2000 | 2000 |
| Irsai Oliver | 250 | 500 | 1000 | 1000 | 250 | 500 | 2000 | 1000 | 2000 |
| Pinot Blanc | 500 | 500 | 1000 | 500 | 250 | 500 | 2000 | 2000 | 1000 |
| Pálava | 500 | 1000 | 1000 | 500 | 250 | 500 | 1000 | 2000 | 2000 |
| Müller-Thurgau | 1000 | 1000 | 1000 | 500 | 250 | 250 | 2000 | 2000 | 2000 |
| Grűner Veltliner | 1000 | 500 | 500 | 1000 | 500 | 250 | 1000 | 2000 | 2000 |
| Feteasca Regala | 1000 | 1000 | 500 | 500 | 500 | 500 | 1000 | 2000 | 1000 |
| Antimicrobial Activity | R Correlation to the Chitinase mRNA Expression |
|---|---|
| Escherichia coli | −0.264 |
| Pseudomonas aeruginosa | −0.154 |
| Salmonella enteritidis subsp. enteritidis | 0.145 |
| Enterococcus faecalis | −0.251 |
| Listeria monocytogenes | −0.036 |
| Staphylococcus aureus subsp. aureus | 0.300 |
| Candida albicans | 0.296 |
| Candida glabrata | −0.393 |
| Candida tropicalis | 0.111 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Žiarovská, J.; Zamiešková, L.; Bilčíková, J.; Fialková, V.; Sabo, J.; Kunová, S.; Kačániová, M. Expression of Specific Class I Chitinase mRNA Levels in Different Grape Varieties and Their Antimicrobial Activity. Agronomy 2020, 10, 1176. https://doi.org/10.3390/agronomy10081176
Žiarovská J, Zamiešková L, Bilčíková J, Fialková V, Sabo J, Kunová S, Kačániová M. Expression of Specific Class I Chitinase mRNA Levels in Different Grape Varieties and Their Antimicrobial Activity. Agronomy. 2020; 10(8):1176. https://doi.org/10.3390/agronomy10081176
Chicago/Turabian StyleŽiarovská, Jana, Lucia Zamiešková, Jana Bilčíková, Veronika Fialková, Jozef Sabo, Simona Kunová, and Miroslava Kačániová. 2020. "Expression of Specific Class I Chitinase mRNA Levels in Different Grape Varieties and Their Antimicrobial Activity" Agronomy 10, no. 8: 1176. https://doi.org/10.3390/agronomy10081176