Abstract
Benzoxazinoids (BXs) are specialized metabolites with protective properties that are synthesized predominantly by Poaceae species, including rye (Secale cereale). Among factors known to influence BXs production, prolonged low temperature has not been studied previously. In this study, the influence of cultivation at 4 °C, which is essential for vernalization, on the concentration of BXs (HBOA, DIBOA, GDIBOA, DIMBOA, GDIMBOA, and MBOA) and the expression level of genes involved in the BX biosynthesis pathway (ScBx1–ScBx5 and ScIgl) in three rye inbred lines was investigated. After cultivation for seven weeks at 4 °C, the expression level of all analyzed genes and BX concentrations had decreased compared with those at the initiation of treatment (21 days after germination) in control and cold-treated plants. At this time point, the decrease in BX concentrations and gene expression was lower in cold-treated plants than in untreated plants. In contrast, at 77 days after germination, the gene expression levels and BX concentrations in untreated plants had generally increased. Investigation of the vernalization impact on rye BXs accumulation, as well as on Bx gene expression, may aid with determination of the most suitable winter lines and cultivars of rye for cultivation and breeding purposes.
1. Introduction
Benzoxazinoids (BXs) are specialized metabolites and an important element of the defense mechanism against biotic and abiotic stresses in plant species predominantly belonging to the Poaceae. BXs also play an important role in allelopathy [1,2,3,4]. In addition, BXs can influence other plant processes such as flowering time, auxin metabolism, iron uptake, and aluminum tolerance [5].
The first step of BX biosynthesis—conversion of indole-3-glycerol phosphate to indole—is catalyzed by indole-3-glycerol phosphate lyase. Four consecutive cytochrome P450 monooxygenases then convert indole to 2,4-dihydroxy-2H-1,4-benzoxazin-3(4H)-one (DIBOA) [6,7,8]. Subsequently, 2-oxoglutarate-dependent dioxygenase and 7-O-methyltransferase are responsible for DIBOA glucoside (DIBOA-Glc; GDIBOA) transformation to 4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one glucoside (DIMBOA-Glc; GDIMBOA) (Figure S1) [9,10]. To protect the cell from toxic aglucons, glucosyltransferases transform the aglucons into harmless, stable compounds of increased solubility, which enable their storage in vacuoles [11,12].
As a response to pathogen or herbivore attack, activity of glucosyl glucosidases leads to conversion of glucosides, ultimately resulting in release of toxic aglucons (i.e., DIMBOA, 4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one) from vacuoles into the cytoplasm [13,14,15]. DIMBOA and especially HDMBOA are unstable compounds, which are spontaneously degraded in watery solutions to MBOA [16]. An additional three O-methyltransferases catalyze the conversion of DIMBOA-Glc to 2-(2-hydroxy-4,7-dimethoxy-1,4-benzoxazin-3-one)-β-d-glucopyranose (HDMBOA-Glc) [7], whereas 2-oxoglutarate-dependent dioxygenase and O-methyltransferase, encoded by the Bx13 and Bx14 genes, catalyze the transformation of DIMBOA-Glc to 2-(2,4,7-trihydroxy-8-methoxy-1,4-benzoxazin-3-one)-β-d-glucopyranose (TRIMBOA-Glc; by Bx13) and 2-(2,4-dihydroxy-7,8-dimethoxy-1,4-benzoxazin-3-one)-β-d-glucopyranose (DIM2BOA-Glc) to 2-(2-hydroxy-4,7,8-trimethoxy-1,4-benzoxazin-3-one)-β-d-glucopyranose (HDM2BOA-Glc) and/or DIMBOA-Glc to HDMBOA-Glc (by Bx14), respectively [17]. All glucosides produced are transported and stored in the vacuole [1,2,3,7].
To date, numerous genes that control BX biosynthesis have been isolated and characterized in such species as wheat (Triticum aestivum) [15,18,19,20,21], maize (Zea mays) [7,10,11,17,22], Hordeum lechleri [23], and rye (Secale cereale) [8,15,24,25,26,27]. The ten rye genes involved in BX biosynthesis identified to date are ScBx1–ScBx7, Scglu, ScGT, and ScIgl.
A number of factors that influence BX concentration (or more rarely the expression of BX synthesis-related genes) have been studied previously, namely plant genotype [15,20,28,29,30,31,32,33], developmental stage [26,34,35], pathogen or herbivore attack [17,28,36,37,38,39], wounding [34,40,41], elevated salinity [42], allelopathy [43], fertilization [44], photoperiod [45,46], and water and/or nitrogen deficiency [47]. However, the influence of low temperature has not been considered previously. Low temperature is particularly important for winter crops, such as the majority of rye cultivars, which require a vernalization period to undergo the transition from vegetative to reproductive development.
The aim of the present study was to determine the effect of prolonged low temperature (4 °C), which is effective for vernalization of the studied rye inbred lines, on the concentration of six BXs (2-hydroxy-1,4-benzoxazin-3-one [HBOA], GDIBOA, DIBOA, GDIMBOA, DIMBOA, and 6-methoxy-2-benzoxazolinone [MBOA]) and expression of related genes ScBx1–ScBx5 and ScIgl. We hypothesized that these processes will be affected by cultivation at 4 °C for a period of at least seven weeks.
2. Materials and Methods
2.1. Plant Materials
The plant material used in the experiments consisted of three unrelated rye inbred lines—L318 (bred at the Department of Plant Genetics, Breeding and Biotechnology, Warsaw University of Life Sciences, Poland), D33, and D39 (both bred by the Polish breeding company Danko Plant Breeding Ltd., Choryń, Poland). The lines were selected based on the distinct BX concentrations measured after a natural vernalization period; line L318 was characterized by the highest, line D33 by the intermediate, and line D39 by the lowest concentration of DIBOA and the sum of BXs [48] (Supplementary Materials, Table S1). The lines chosen for experiments also differ with regard to the distribution and frequency of single-nucleotide polymorphisms detected in the ScBx1–ScBx5 genes [48].
For the first three weeks, plants were cultivated under the control condition (12 h/12 h (light/dark) photoperiod at 15 °C/12 °C (day/night)) and constant relative humidity 70% in 24-well trays filled with a mixture of peat and perlite (1:1, v/v), with six plants per single tray segment (7 cm diameter). Each inbred line was represented by 120 plants in total—six plants per one out of four biological replicates, per one out of five kinds of plant material treatment and time point of sampling: (1) untreated plants sampled at 21 days after germination (dag), (2) untreated plants sampled at 70 dag, (3) untreated plants sampled at 77 dag, (4) cold-treated plants sampled at 70 dag, and (5) cold-treated plants sampled at 77 dag (Figure S2). Three-week-old seedlings were placed either in a cold chamber maintained at 4 °C, 12 h/12 h (light/dark) photoperiod and relative humidity of 70% (cold-treated plants) or cultivated continuously under the control condition (the same as during the first three weeks) until the end of the experiment (untreated, control plants). After cultivation for seven weeks in the cold chamber at 4 °C, the cold-treated plants were transferred to the control condition for one week. Plant material (aerial parts of plants) for each biological replication and at each time point (21, 70, and 77 dag) was divided into two portions; one was assigned for gene expression analysis and the second portion for biochemical assay. The tissues were immediately frozen using liquid nitrogen and then, in the case of material destined for biochemical analysis, the samples were lyophilized.
2.2. RNA Isolation and cDNA Synthesis
Frozen tissue (100 mg) was ground in liquid N2, then total RNA was isolated using the GeneMATRIX Universal RNA Purification Kit version 1.2 (Eurx, Gdańsk, Poland) in accordance with the manufacturer’s protocol. The isolated RNA was diluted in 40 μL RNase-free water. The RNA integrity and concentration were measured using a NanoDrop 2000 spectrophotometer. To avoid genomic DNA contamination, Turbo DNase (Thermo Fisher Scientific, Waltham, MA, USA) was added. The purified RNA was then used as the template for cDNA synthesis using the High-Capacity cDNA Reverse Transcription kit (Thermo Fisher Scientific). The total volume of the reaction mixture was 20 μL, which contained 2 μg template RNA, 2 μL of 10× RT buffer, 0.8 μL of 25× dNTPs mix (100 mM), 2 μL of 10 × RT random primers, 1 μL MultiScribe reverse transcriptase (50 U/µL), and 4.2 μL nuclease-free water. The reaction was performed at 25 °C for 10 min, 37 °C for 120 min, and 85 °C for 5 min. The resulting cDNA was diluted with RNase-free water.
2.3. Quantitative Real-Time PCR Analysis
To examine the expression profiles of ScBx1–ScBx5 (GenBank, accession No. KF636825–KF636828; KF620524) and ScIgl (GenBank, accession No. MN120476), quantitative real-time PCR (qRT-PCR) analyses were conducted in a 96-well plate, with four biological and two technical replicates. A reference gene, HvAct (GenBank, accession No. AY145451), was chosen experimentally based on the results of preliminary experiments in which the utility of three genes as internal reference genes were tested: ScGADPH (GenBank, accession No. JQ659189.1), Ta54227 (GenBank, accession No. Pr0102692400), and HvAct (GenBank, accession No. AY145451). The gene HvAct was selected (using the geNorm algorithm) as the most stable gene and, therefore, the most suitable as an internal control reference gene. The same experimental approach was positively verified in our earlier works [27,43,49].
The qRT-PCR analyses were performed with the LightCycler 96 Real-Time System (Roche, Basel, Switzerland), applying the following program: 95 °C for 600 s; 32 cycles of 95 °C for 10 s, 57 °C for 10 s, and 72 °C for 15 s; and then 95 °C for 10 s, 55 °C for 60 s, and 97 °C for 1 s. The total volume of the reaction mixture was 20 µL, which contained 4 μL cDNA (5 ng/µL), 1 μL each gene-specific primer (5 mM), 4 μL RNase-free water, and 10 μL FastStart Essential DNA Green Master (Roche). The qPCR reaction efficiency for primers used in the analysis ranged from 1.94 to 2.0, and R2 value was ≥0.99. The expression level of the analyzed genes was normalized against the HvAct expression level by using the 2−ΔΔCt method [50] and the results were presented as relative gene expression. Data were presented as fold change in cold-treated rye plants relative to untreated (control) plants. The results of the gene expression analyses are summarized in Supplementary Materials, Table S2 (mean values), Table S3 (medians). The primer sequences used in the qRT-PCR reactions are listed in Table 1.

Table 1.
Primers used in quantitative real-time PCR analyses.
2.4. Biochemical Analysis
Quantitative analyses of BXs were conducted using a modified protocol published previously [48]. Briefly, samples of plant material (100 mg) were mixed with diatomaceous earth and extracted with 70% methanol (containing the internal standard: 2 µg/mL indoxyl β-d-glucoside; IbG). The extracts were evaporated to dryness under reduced pressure, reconstituted in 1 mL methanol containing 0.1% (v/v) acetic acid, and stored at −20 °C. Before the analyses, extracts were centrifuged for 20 min at 23,000× g at 4 °C and filtered using Whatman Mini-Uniprep filtering vials (GE Healthcare, Amersham, UK) with a regenerated cellulose membrane of 0.2 µm pore size.
A Waters Acquity UPLC system (Waters, Milford, MA, USA) equipped with a triple quadrupole mass spectrometer (Waters TQD) was used for the analyses. Benzoxazinoids were separated on a Waters BEH C18 column (100 × 2.1 mm, 1.7 µm) with a linear, 8.5-min-long gradient from 3% to 15% acetonitrile containing 0.1% (v/v) formic acid (solvent B) in 0.1% formic acid (solvent A). All separations were performed at 50 °C with a flow rate of 0.6 mL/min. The column effluent was diverted into the waste for the first 1.5 min of separation, to prevent contamination of the ion source with polar constituents of the sample matrix.
Subsequently, the effluent was directed into the ion source of the mass spectrometer and analyzed in the multiple reaction mode (MRM), as described previously. Briefly, the mass spectrometer operated in the negative mode with the following parameters: capillary voltage −2.8 kV, extractor 3 V, RF lens 100 mV, source temperature 130 °C, desolvation temperature 400 °C, desolvation gas flow 1000 L/h, cone gas flow 100 L/h.
Cone voltages, ranging from 15 to 35 V, were optimized individually for each analyzed compound. Collision cell entrance and exit potentials were set to −2 and 0.5 V, respectively. Parameters of quadrupoles 1 and 3 were set to achieve unit-mass resolution. Details of precursor, qualifier, and quantifier ions are shown in Supplementary Materials, Table S4. Waters MassLynx 4.1 SCN 919 software was used for data acquisition and processing.
Calibration curves (seven concentration points between 0.3 and 35 ng/µL) were prepared from standard solutions of DIBOA, DIMBOA, HBOA, GDIMBOA, MBOA, and IbG (1 mg/mL each). GDIMBOA was used as a reference standard for GDIBOA quantitation. The response for each concentration point was calculated as the ratio of the analyte to the IgG peak area. The linearity of the concentration–response relationships for all investigated compounds was maintained from 0.3 to 30 ng/µL. The accuracy of the quantitation was monitored by injections of the quality-control samples, consisting of a mixture of all quantitation standards at 1, 15, and 30 ng/μL, after every ten injections of plant extract samples. Method detection limits were determined by ten repeated injections of the 0.3 ng/µL solution of the calibration standards. The results of the biochemical analysis are summarized in Supplementary Materials, Table S5 (mean values) and Table S6 (medians).
2.5. Statistical Analysis
The significance of differences (at α = 0.05 or α = 0.01) in relative gene expression or BX concentrations were determined with the Mann–Whitney U test using Statgraphics Plus ver. 3.0 software. For each inbred line the following median comparisons were performed:
- 21-day-old seedlings vs. 70-day-old untreated plants;
- 21-day-old seedlings vs. 70-day-old cold-treated plants;
- 70-day-old untreated plants vs. 70-day-old cold-treated plants;
- 77-day-old untreated plants vs. 77-day-old cold-treated plants;
- 70-day-old untreated plants vs. 77-day-old untreated plants;
- 70-day-old cold-treated plants vs. 77-day-old cold-treated plants.
3. Results
3.1. Effect of Low Temperature on Gene Expression
At the first time point (21 dag), the relative expression levels of ScBx1–ScBx5 were highest in line D33 and lowest in line D39. The opposite trend was observed for ScIgl, which was most highly expressed in line D39 and the lowest expression was observed in line D33. After seven weeks of cultivation, under both 4 °C and the control temperature, the expression level of all analyzed genes, except that of ScIgl in lines L318 and D33, decreased compared with that observed at the first time point (Figure 1, Table 2). After seven weeks, expression was no longer detectable for ScBx1 in untreated and cold-treated plants of all inbred lines, ScBx2 in cold-treated plants of lines D33 and D39, and ScBx4 in cold-treated plants of line D39. Except for a small number of cases, the expression level of the majority of genes continued to decline during the next seven days (77 dag). A noticeably different trend, i.e., an increase in expression level, was detected for ScIgl in untreated plants of line D33, and for ScBx3, ScBx4, and ScBx5 in untreated plants of line D39. Comparison of the expression levels of these genes in control and cold-treated plants at 70 dag revealed that, apart from ScBx4 in lines D33 and D39 and ScBx5 in line D33, expression was higher in cold-treated plants. The highest upregulation in cold-treated plants related to control plants was observed for ScIgl in lines L318 and D39 and for ScBx3 in all lines (Figure 2). At the third time point (77 dag), only in line L318 was the majority of genes upregulated. In the remaining two lines, D33 and D39, almost all of the analyzed genes were downregulated compared with the control plants. The exception was ScIgl in line D39, which was expressed at a higher level in cold-treated plants than in untreated plants. Similar to the second time point, at 77 dag the highest relative expression level in cold-treated plants relative to control plants was observed for ScIgl in lines L318 and D39. The lowest ratio was observed for ScBx2 in line D39.
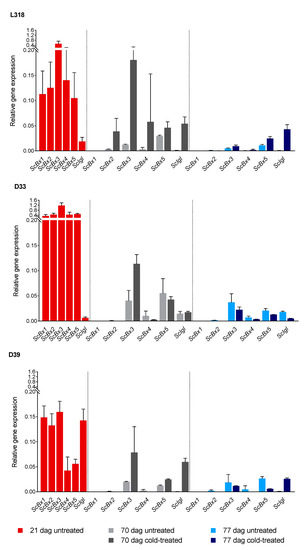
Figure 1.
Relative gene expression profiles of ScBx1–ScBx5 and ScIgl in untreated and cold-treated plants of the rye inbred lines L318, D33, and D39 measured at 21, 70, and 77 days after germination (dag). The results are presented as means ± SD of four biological replications. The significance of differences in gene expression levels between untreated and cold-treated plants (assessed using the Mann–Whitney U test) is presented in Table 2.
Table 2.
Significance of differences in ScBx1–ScBx5 and ScIgl expression levels between untreated and cold-treated plants of the rye inbred lines L318, D33, and D39 assessed using the Mann–Whitney U test.
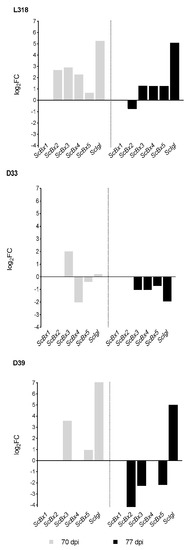
Figure 2.
ScBx1–ScBx5 and ScIgl gene expression changes represented as fold changes (FC) in cold-treated plants relative to untreated plants of rye inbred lines L318, D33, and D39 cultivated for 70 and 77 days. The results are presented as means of four biological replications.
3.2. Effect of Low Temperature on BX Concentration
At all time points, the dominant BXs detected were GDIBOA and DIBOA. Similarly, as observed for gene expression, the concentration of the majority of BXs decreased essentially at the second time point (70 dag) compared with that at the first time point (21 dag), regardless of whether the plants were cultivated at 4 °C for seven weeks or under the control condition (Figure 3, Table 3). Only in the case of HBOA in untreated plants of line L318, and MBOA in untreated plants of lines L318 and D33, the concentration of these BXs increased at 70 dag compared to the first time point. At this time point, DIMBOA in cold-treated plants of lines D33 and D39, and MBOA in untreated plants of line D39 were no longer detectable. During the next seven days, the concentration of the majority of BXs in untreated plants usually increased (except for HBOA in L318 and GDIBOA in D39), whereas for cold-treated plants the opposite trend was observed, i.e., a decrease in concentration. The only exception was DIBOA in cold-treated plants of line D33, in which the concentration increased slightly between the second and third time points. At 77 dag, the concentrations of GDIMBOA and DIMBOA of lines L318 and D39 was undetectable in cold-treated plants. At 70 dag the GDIBOA concentration increased in all inbred rye lines (Figure 4). In addition, the concentrations of HBOA in lines D33 and D39, and of DIBOA in L318 and D33 increased at this time point. The highest BX concentrations in cold-treated rye plants relative to untreated plants was observed for GDIBOA and HBOA in line D33. During the next seven days, the concentrations of the majority of analyzed BXs declined in all lines. Only the concentration of HBOA in all lines and of DIBOA in D33 increased. At the third time point, the highest ratio of BX concentrations in cold-treated plants related to control plants was observed for HBOA in line D33.
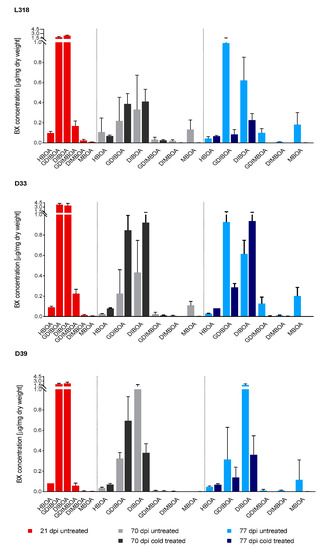
Figure 3.
Benzoxazinoid (BX) concentrations in untreated and cold-treated plants of the rye inbred lines L318, D33, and D39 measured at 21, 70, and 77 days after germination. The results are presented as means ± SD of four biological replications. The significance of differences in BX concentrations between untreated and cold-treated plants (assessed using the Mann–Whitney U test) are presented in Table 3.
Table 3.
Significance of differences in benzoxazinoid (BX) concentration between untreated and cold- treated plants of the rye inbred lines L318, D33, and D39 assessed using the Mann–Whitney U test.
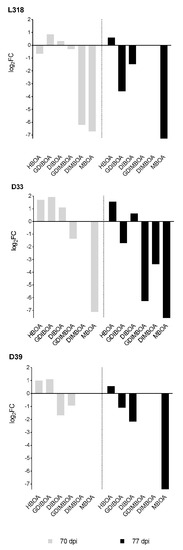
Figure 4.
Benzoxazinoid (BX) concentrations in cold-treated plants relative to untreated plants of the rye inbred lines L318, D33, and D39 cultivated for 70 and 77 days. The results are presented as means of four biological replications.
4. Discussion
The properties and biosynthesis regulatory pathway for BXs have been intensively studied for more than 60 years. Although a variety of factors that affect BX biosynthesis have been thoroughly investigated, the effect of prolonged low temperature has been overlooked. Understanding the impact of low temperature is important for at least two reasons: first, winter cereal species must undergo a vernalization period under a temperature ranging from 0.2 °C to 10 °C [51], which is essential for the transition from vegetative to reproductive development, and vernalization requirements for rye can vary between 1.1 °C to 6 °C for 15 to 63 days [52,53,54]; and second, no standardized protocols have been established for plant material sampling, neither for biochemical nor for molecular analyses. Therefore, in the present study we examined the impact of prolonged low temperature on the concentration of BXs (HBOA, GDIBOA, DIBOA, GDIMBOA, DIMBOA, and MBOA) and the expression level of related genes (ScBx1–ScBx5 and ScIgl).
As might be expected, the concentration of analyzed BXs and the expression level of the BX synthesis-related genes decreased substantially over time. This phenomenon is a natural trend reported by many authors, and documented previously for rye: e.g., Sue et al. [55], La Hovary [34], Groszyk et al. [35], and Tanwir et al. [26].
With regard to genes, the most distinct pattern observed was a significant increase in expression level at 70 dag followed by a decrease during the next seven days for ScIgl in cold-treated plants of lines L318 and D33 after their transfer to the control condition. In line D33 cultivated under the control condition for the entire experiment, the expression level of ScIgl increased continuously. These observations essentially agree with the results published previously by Wlazło et al. [27], who observed that ScIgl assumes the role of ScBx1 at advanced developmental stages and its expression level may even increase between the tenth and eleventh week of cultivation as observed in line D33. With regard to BXs, the late change in the concentration of two metabolites (DIBOA and GDIBOA) in plants not exposed to low temperature, i.e., the concentration increased significantly at 77 dag, was contrary to expectations. The same phenomenon was described by Wlazło et al. [27], which the authors attributed to ScIgl gene activity.
The gene expression level in plants exposed to low temperature (4 °C) for seven weeks generally was initially higher than that detected in plants cultivated under the control condition except for ScBx4 and ScBx5 in line D33—the expression level of these two genes was higher in plants that were not subjected to cold treatment. After cultivation for seven days under the control condition after cold treatment, the same responses (except for ScBx2) were observed in line L318, whereas those in line D33 changed completely, i.e., the expression level of the majority of genes was higher in the control plants. The line D39 was characterized by the opposite relationship compared with expression levels at 70 dag in the case of four genes (ScBx2, ScBx3, and ScBx5). Therefore, it may be concluded that cultivation for seven weeks at a low temperature, by slowing plant metabolism, results in a lower rate of reduction in gene expression level. Cultivation under the control temperature for seven days after cold treatment could be perceived by cold-treated plants as a new stress, which explains why in these plants the gene expression level decreased generally to a greater extent than that of untreated plants. However, the specificity of the reaction to temperature and age-related changes clearly depended on the plant genotype. The most visible difference was that in line L318, with the exception of ScBx2, a higher decrease in gene expression level was observed for plants subjected to low temperature, whereas in lines D33 and D39 reduced gene expression was observed at 70 dag; at 77 dag higher expression levels were observed for control plants.
Similarly, as for gene expression, the BX concentrations generally declined with plant age except for the aforementioned instances. The gradual decline in BX concentrations is typical for rye and was described previously, inter alia, by Rice et al. [56], Sue et al. [15], La Hovary [34], La Hovary et al. [57], and Groszyk et al. [35].
In all lines, regardless of the time point and treatment, GDIBOA and DIBOA were consistently the dominant BXs detected, which was reported previously as characteristic for rye [26,27,34,58,59]. The impact of low temperature on BX synthesis was not as distinct as it was on gene expression, and depended on plant genotype and time point to a greater extent, especially at 70 dag. After cultivation for seven days after cold treatment, a predominantly negative influence of cold stress was observed. Nevertheless, a number of consistent responses to low temperature and subsequent transfer to the control condition were observed, i.e., the concentrations of MBOA and GDIMBOA were consistently (at 70 and 77 dag) negatively affected, GDIBOA concentration was positively affected at 70 dag and negatively at 77 dag, and HBOA concentration was usually positively influenced (except for line L318 at 70 dag).
For the most important BX, DIBOA, we did not observe a common response to low temperature; on the contrary, each line showed a different response to cold treatment. The only line in which treatment with low temperature positively influenced DIBOA concentration during the entire experiment was D33. With regard to line L318, a positive effect of low temperature was noted at 70 dag, immediately after transfer to the control condition, and in line D39 the DIBOA concentration was higher in the control plants. The results of the present experiments in relation to DIBOA concentration are not completely consistent with those of Reberg-Horton et al. [33], who reported that DIBOA concentration declined more slowly in winter rye genotypes compared with that of facultative types of rye, which suggested that vernalization had a promotive impact. In addition, La Hovary et al. [57] observed that rye plants in the vegetative phase accumulate BXs at higher concentrations than plants in the reproductive phase. Nevertheless, the authors pointed to the dependence of this trend on the genotype.
Do the Changes in Gene Expression Correspond to Changes in BX Synthesis?
In the present study, the gene expression levels did not entirely correspond with the BX concentrations. A relatively strong correspondence was observed at the first time point (three-week-old seedlings), at 70 dag in untreated plants, and at 77 dag in cold-treated plants. The most distinct and surprising finding was a significant increase in the concentrations of GDIBOA and DIBOA in untreated plants of lines L318 and D33 and a simultaneous decrease in expression level of the majority of genes except for ScIgl. It can be concluded that the high level of transcripts of this particular gene is responsible for this, which agrees with the results published recently by Wlazło et al. [27]. In turn, a relatively high accordance between gene expression and BX concentration was observed in untreated plants of line D39 at 77 dag. With regard to cold-treated plants, the most distinct discordance was noted for lines L318 and D33 at 70 dag, when a significant increase in ScIgl transcript abundance was not accompanied by higher BX concentrations. The present observations concerning cold-treated rye plants correspond only partially with the results of La Hovary et al. [56], who suggested that lower BX concentrations in rye plants appear to be due to the progression of flowering despite the higher abundance of Bx gene transcripts.
The tissues used for BX biochemical analyses have been collected from plants at diverse developmental stages, generally, but not always, before the vernalization period. Groszyk et al. [35] and Wlazło et al. [27] analyzed nonvernalized plants at 1 to 90 and 14 to 77 days old, respectively. Reberg-Horton et al. [33], Rice et al. [56], and La Hovary [34] sampled leaves from several-month-old, field-grown plants that had experienced vernalization. Thus, it is extremely difficult to compare and discuss the data from these different experiments.
In the present study, we showed that exposure to low temperature (4 °C) for seven weeks, which is necessary for winter crops to undergo the transition from vegetative to reproductive growth, affects BX synthesis and expression of BX synthesis-related genes. In general, a positive effect of cold temperature was observed immediately after the change in temperature from 4 °C to the control condition, and cultivation for seven days under the control temperature after cold treatment had a negative impact. This observation suggests that BXs production can be modified by manipulation of the duration of the vernalization period, which, of course, is fully achievable under greenhouse cultivation. However, to some extent, we can also influence the potential of winter arable crops cultivated in the field by choosing the length of the vernalization period. This is particularly important in the era of ongoing climate change. The changing global climate system is evident throughout the world [60,61]. Moreover, winter rye belongs to the crops most influenced by climate factors [62]. Based on the results of Reberg-Horton et al. [33] and La Hovary et al. [57], the naturally prolonged vernalization (through earlier sowing) of field-cultivated rye is expected to stimulate BXs production, at least in some genotypes. The same can be achieved by spring sowing of winter rye cultivars that may prevent or delay generative growth and thus, as proposed by La Hovary [34], stimulate BXs production. Such an effect was observed in the present study for GDIBOA and MBOA in all three inbred lines cultivated under the control condition throughout the experimental period. Besides the increase in BX production, it can be presumed that the prolonged vernalization should positively affect the general performance of overwintering cereals, particularly grain yield, and better winter tolerance [63,64] and disease resistance to several fungal pathogens by enhanced production of pathogenesis-related (PR) proteins [65,66,67]. However, this assumption must be verified experimentally in field trials lasting many years, in different locations, and by comprising sufficiently large plant material.
A specialized, but important conclusion that can be drawn from the present experiments is that researchers must be more careful with the interpretation of their results in comparison with those of other research teams.
5. Conclusions
- The cultivation for seven weeks at 4 °C causes a decrease of BX concentration and expression level of related genes compared with those on the 21st day after seed germination.
- The decrease over time in BX concentration and in expression level of related genes is greater in cold-treated than in untreated plants.
- Manipulating the duration of the vernalization period can significantly affect the production of BXs in rye.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/10/9/1260/s1, Table S1: Benzoxazinoid (BX) concentrations of the rye inbred lines L318, D33, and D39 chosen for the experiments, Table S2: Mean relative expression levels of ScBx1–ScBx5 and ScIgl genes in the rye inbred lines L318, D33, and D39 cultivated under low and control temperatures, Table S3: Medians for gene expression values measured at 3 time points in untreated and cold-treated plants, Table S4. Details of multiple-reaction monitoring analyses, Table S5. Mean benzoxazinoid (BX) concentration in the rye inbred lines L318, D33, and D39 cultivated under low and control temperatures, Table S6. Medians for BX concentration values measured at 3 time points in untreated and cold-treated plants, Figure S1. Biosynthesis pathway of benzoxazinoids in maize (Zea mays), Figure S2. Schematic illustration of the experimental design.
Author Contributions
Conceptualization, M.R.-T.; methodology, B.B., M.Ś., A.S., and M.K.; statistical analysis, L.B.; data analysis, B.B. and M.Ś.; writing—original draft preparation, review and editing, M.R.-T., B.B., and M.Ś. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by National Science Centre, grant No. 2015/19/B/NZ9/00921.
Acknowledgments
We thank Robert McKenzie, from Edanz Group (https://en-author-services.edanzgroup.com/), for editing a draft of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Frey, M.; Schullehner, K.; Dick, R.; Fiesselmann, A.; Gierl, A. Benzoxazinoid biosynthesis, a model for evolution of specialized metabolic pathways in plants. Phytochemistry 2009, 70, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, H.M. Hydroxamic acids derived from 2-hydroxy-2H-1,4-benzoxazin-3(4H)-one: Key defense chemicals of cereals. J. Agric. Food Chem. 2009, 57, 1677–1696. [Google Scholar] [CrossRef] [PubMed]
- Makowska, B.; Bakera, B.; Rakoczy-Trojanowska, M. The genetic background of benzoxazinoid biosynthesis in cereals. Acta Physiol. Plant 2015, 37, 176. [Google Scholar] [CrossRef]
- Wouters, F.C.; Blanchett, B.; Gershenzo, J.; Vassão, D.G. Plant defense and herbivore counter-defense: Benzoxazinoids and insect herbivores. Phytochem. Rev. 2016, 15, 1127–1151. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Richter, A.; Jander, G. Beyond Defense: Multiple Functions of Benzoxazinoids in Maize Metabolism. Plant Cell Physiol. 2018, 59, 1528–1537. [Google Scholar] [CrossRef]
- Chu, H.Y.; Wegel, E.; Osbourn, A. The plant genome: An evolutionary view on structure and function, from hormones to specialized metabolism: The emergence of metabolic gene clusters in plants. Plant J. 2011, 66, 66–79. [Google Scholar] [CrossRef]
- Meihls, L.N.; Handrick, V.; Glauser, G.; Barbier, H.; Kaur, H.; Haribal, M.M.; Lipka, A.E.; Gershenzon, J.; Buckler, E.S.; Erb, M.; et al. Natural variation in maize aphid resistance is associated with 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one glucoside methyltransferase activity. Plant Cell 2013, 25, 2341–2355. [Google Scholar] [CrossRef]
- Bakera, B.; Rakoczy-Trojanowska, M. Isolation and structural analysis of the Bx6 and Bx7 genes controlling the biosynthesis of benzoxazinoids in rye (Secale cereale L.). Acta Physiol. Plant 2020, 42, 56. [Google Scholar] [CrossRef]
- Frey, M.; Huber, K.; Park, W.J.; Sicker, D.; Lindberg, P.; Meeley, R.B.; Simmons, C.R.; Yalpani, N.; Gierl, A. A 2-oxoglutarate-dependent Dioxygenase Is Integrated in DIMBOA-biosynthesis. Phytochemistry 2003, 62, 371–376. [Google Scholar] [CrossRef]
- Jonczyk, R.; Schmidt, H.; Osterrieder, A.; Fiesselmann, A.; Schullehner, K.; Haslbeck, M.; Sicker, D.; Hofmann, D.; Yalpani, N.; Simmons, C.; et al. Elucidation of the final reactions of DIMBOA-glucoside biosynthesis in maize: Characterization of Bx6 and Bx7. Plant Physiol. 2008, 146, 1053–1063. [Google Scholar] [CrossRef]
- Rad Von, U.; Huttl, R.; Lottspeich, F.; Gierl, A.; Frey, M. Two glucosyltransferases are involved in detoxification of benzoxazinoids in maize. Plant J. 2001, 28, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Dick, R.; Rattei, T.; Haslbeck, M.; Schwab, W.; Gierl, A.; Frey, M. Comparative analysis of benzoxazinoid biosynthesis in monocots and dicots: Independent recruitment of stabilization and activation functions. Plant Cell 2012, 24, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Babcock, G.D.; Esen, A. Substrate specificity of maize b-glucosidase. Plant Sci. 1994, 101, 31–39. [Google Scholar] [CrossRef]
- Oikawa, A.; Ishihara, A.; Iwamura, H. Induction of HDMBOA-Glc accumulation and DIMBOA-Glc4-O-methyltransferase by jasmonic acid in poaceous plants. Phytochemistry 2002, 61, 331–337. [Google Scholar] [CrossRef]
- Sue, M.; Nakamura, C.; Nomura, T. Dispersed benzoxazinone gene cluster: Molecular characterization and chromosomal localization of glucosyltransferase and glucosidase genes in wheat and rye. Plant Physiol. 2011, 157, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Niculaes, C.; Abramov, A.; Hannemann, L.; Frey, M. Plant protection by benzoxazinoids–recent insights into biosynthesis and function. Agronomy 2018, 8, 143. [Google Scholar] [CrossRef]
- Handrick, V.; Robert, C.A.M.; Ahern, K.R.; Zhou, S.; Machado, R.A.R.; Maag, D.; Glauser, G.; Fernandez-Penny, F.E.; Chandran, J.N.; Rodgers-Melnik, E.; et al. Biosynthesis of 8-O-methylated benzoxazinoid defence compounds in maize. Plant Cell Online 2016, 28, 1682–1700. [Google Scholar] [CrossRef]
- Nomura, T.; Ishihara, A.; Imaishi, H.; Endo, T.R.; Ohkawa, H.; Iwamura, H. Molecular characterization and chromosomal localization of cytochrome P450 genes involved in the biosynthesis of cyclic hydroxamic acids in hexaploid wheat. Mol. Genet. Genom. 2002, 267, 210–217. [Google Scholar] [CrossRef]
- Nomura, T.; Ishihara, A.; Imaishi, H.; Ohkawa, H.; Endo, T.R.; Iwamura, H. Rearrangement of the genes for the biosynthesis of benzoxazinones in the evolution of Triticeae species. Planta 2003, 217, 776–782. [Google Scholar] [CrossRef]
- Nomura, T.; Ishihara, A.; Yanagita, R.C.; Endo, T.R.; Iwamura, H. Three genomes differentially contribute to the biosynthesis of benzoxazinones in hexaploid wheat. Proc. Natl. Acad. Sci. USA 2005, 102, 16490–16495. [Google Scholar] [CrossRef]
- Nomura, T.; Nasuda, S.; Kawaura, K.; Ogihara, Y.; Kato, N.; Sato, F.; Kojima, T.; Toyoda, A.; Iwamura, H.; Endo, T.R. Structures of the three homoeologous loci of wheat benzoxazinone biosynthetic genes TaBx3 and TaBx4 and characterization of their promoter sequences. Theor. Appl. Genet. 2008, 116, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Frey, M.; Chomet, P.; Glawischnig, E.; Stettner, C.; Grün, S.; Winklmair, A.; Eisenreich, W.; Bacher, A.; Meeley, R.B.; Briggs, S.P.; et al. Analysis of a chemical plant defence mechanism in grasses. Science 1997, 277, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Grün, S.; Frey, M.; Gierl, A. Evolution of the Indole Alkaloid Biosynthesis in the Genus Hordeum: Distribution of Gramine and DIBOA and Isolation of the Benzoxazinoid Biosynthesis Genes FromHordeum Lechleri. Phytochemistry 2005, 66, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Nikus, J.; Esen, A.; Jonsson, L.M.V. Cloning of a plastidic rye (Secale cereale) b-glucosidase cDNA and its expression in Escherichia coli. Physiol. Plant 2003, 118, 337–345. [Google Scholar] [CrossRef]
- Bakera, B.; Makowska, B.; Groszyk, J.; Niziołek, M.; Orczyk, W.; Bolibok-Brągoszewska, H.; Hromada-Judycka, A.; Rakoczy-Trojanowska, M. Structural characteristics of ScBx genes controlling the biosynthesis of hydroxamic acids in rye (Secale cereale L.). J. Appl. Genet. 2015, 56, 287–298. [Google Scholar] [CrossRef]
- Tanwir, F.; Dionisio, G.; Adhikari, K.B.; Fomsgaard, I.S.; Gregersen, P.L. Biosynthesis and chemical transformation of benzoxazinoids in rye during seed germination and the identification of a rye Bx6-like gene. Phytochemistry 2017, 140, 95–107. [Google Scholar] [CrossRef]
- Wlazło, A.; Święcicka, M.; Koter, M.D.; Krępski, T.; Bolibok, L.; Stochmal, A.; Kowalczyk, M.; Rakoczy-Trojanowska, M. Genes ScBx1 and ScIgl–Competitors or Cooperators? Genes 2020, 20, 223. [Google Scholar] [CrossRef]
- Gianoli, E.; Papp, M.; Niemeyer, H.M. Costs and benefits of hydroxamic acids-related resistance in winter wheat against the bird cherry-oat aphid, Rhopalosiphum padi L. Ann. Appl. Biol. 1996, 129, 83–90. [Google Scholar] [CrossRef]
- Copaja, S.V.; Villarroel, E.; Bravo, H.R.; Pizarro, L.; Argandoña, V.H. Hydroxamic acids in Secale cereale L., and the relationship with their antifeedant and allelopathic properties. Z. Naturforsch. C. 2006, 61, 670–676. [Google Scholar] [CrossRef]
- Barrίa, B.N.; Copaja, S.V.; Niemeyer, H.M. Occurrence of DIBOA in wild Hordeum species and its relation to aphid resistance. Phytochemistry 1992, 31, 89–91. [Google Scholar] [CrossRef]
- Niemeyer, H.M.; Copaja, S.V.; Barrıa, B.N. The Triticeae as sources of hydroxamic acids, specialized metabolites in wheat conferring resistance against aphids. Hereditas 1992, 116, 295–299. [Google Scholar] [CrossRef]
- Frey, M.; Kliem, R.; Saedler, H.; Gierl, A. Expression of a cytochrome P450 gene family in maize. Mol. Gen. Genet. 1995, 246, 100–109. [Google Scholar] [CrossRef]
- Reberg-Horton, S.C.; Burton, J.D.; Danehower, D.A.; Ma, G.Y.; Monks, D.W.; Murphy, J.P.; Ranells, N.N.; Williamson, J.D.; Creamer, N.G. Changes over time in the allelochemical concentration of ten cultivars of rye (Secale cereale L.). J. Chem. Ecol. 2005, 31, 179–193. [Google Scholar] [CrossRef]
- La Hovary, C. Allelochemicals in Secale cereale: Biosynthesis and Molecular Biology of Benzoxazinones. 2011. Available online: https://repository.lib.ncsu.edu/handle/1840.16/6844 (accessed on 5 May 2020).
- Groszyk, J.; Kowalczyk, M.; Yanushevska, Y.; Stochmal, A.; Rakoczy-Trojanowska, M.; Orczyk, W. Identification and VIGS-based characterization of Bx1 ortholog in rye (Secale cereale L.). PLoS ONE 2017, 12, e0171506. [Google Scholar] [CrossRef]
- Zheng, Y.Q.; Zhao, Y.; Dong, F.S.; Yao, J.R.; Hurle, K. Relationship of DIMBOA concentration in wheat seedlings and its resistance to plant pathogens. Allelopath. J. 2005, 15, 137–143. [Google Scholar]
- Ahmad, S.; Veyrat, N.; Gordon-Weeks, R. Benzoxazinoid metabolites regulate innate immunity against aphids and fungi in maize. Plant Physiol. 2011, 157, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Praz, C.; Li, B.; Singla, J.; Robert, C.A.; Kessel, B.; Scheuermann, D.; Lüthi, L.; Ouzunova, M.; Erb, M.; et al. Fungal resistance mediated by maize wall-associated kinase ZmWAK-RLK1 correlates with reduced benzoxazinoid concentration. New Phytol. 2019, 221, 976–987. [Google Scholar] [CrossRef]
- Ding, X.; Yang, M.; Huang, H.; Chuan, Y.; He, X.; Li, C.; Zhu, Y.; Zhu, S. Priming maize resistance by its neighbors: Activating 1,4-benzoxazine-3-ones synthesis and defense gene expression to alleviate leaf disease. Front. Plant Sci. 2015, 6, 830. [Google Scholar] [CrossRef]
- Morse, S.; Wratten, S.D.; Edwards, P.J.; Niemeyer, H.M. Changes in the hydroxamic acid concentration of maize leaves with time and after artificial damage; implications for insect attack. Ann. Appl. Biol. 1991, 119, 239–249. [Google Scholar] [CrossRef]
- Kruidhof, H.M.; Van Dam, N.M.; Ritz, C.; Lotz, L.A.P.; Kropff, M.J.; Bastiaans, L. Mechanical wounding under field conditions: A potential tool to increase the allelopathic inhibitory effect of cover crops on weeds? Europ. J. Agron. 2014, 52, 229–236. [Google Scholar] [CrossRef]
- Makleit, P. Changes in cyclic hydroxamic acid concentration of various rye varieties for the effect of abiotic stress. Acta Biol. Szeged. 2005, 49, 103–104. [Google Scholar]
- Rakoczy-Trojanowska, M.; Święcicka, M.; Bakera, B.; Kowalczyk, M.; Stochmal, A.; Bolibok, L. Co-cultivating rye with berseem clover affects benzoxazinoid production and expression of related genes. Crop Sci. 2020. [CrossRef]
- Mwaja, V.N.; Masiunas, J.B.; Weston, L.A. Effects of fertility on biomass, phytotoxicity, and allelochemical concentration of cereal rye. J. Chem. Ecol. 1995, 21, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Epstein, W.W.; Rowsemitt, C.N.; Berger, P.J.; Negus, N.C. Dynamics of 6 methoxybenzoxazolinone in winter wheat, Effects of photoperiod and temperature. J. Chem. Ecol. 1986, 12, 2011–2021. [Google Scholar] [CrossRef] [PubMed]
- Gianoli, E.; Niemeyer, H.M. Environmental effects on the accumulation of hydroxamic acids in wheat seedlings: The importance of plant growth rate. J. Chem. Ecol. 1997, 23, 543–551. [Google Scholar] [CrossRef]
- Nie, C.R.; Luo, S.M.; Lin, C.X.; Zeng, R.S.; Huang, J.H.; Wang, J.W. Status of DIMBOA and phenolic acids in transgenic Bt corn. Aust. J. Agri. Res. 2005, 56, 833–837. [Google Scholar] [CrossRef]
- Rakoczy-Trojanowska, M.; Orczyk, W.; Krajewski, P.; Bocianowski, J.; Stochmal, A.; Kowalczyk, M. ScBx gene based association analysis of hydroxamate concentration in rye (Secale cereale L.). J. Appl. Genet. 2017, 58, 1–9. [Google Scholar] [CrossRef]
- Świecicka, M.; Dmochowska-Boguta, M.; Orczyk, W.; Grądzielewska, A.; Stochmal, A.; Kowalczyk, M.; Bolibok, L.; Rakoczy-Trojanowska, M. Changes in benzoxazinoid contents and the expression of the associated genes in rye (Secale cereal L.) due to brown rust and the inoculation procedure. PLoS ONE 2020, 15, e0233807. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Stoskopf, N.C. Cereal Grain Crops; Reston Publishing: Reston VA, USA, 1985; pp. 403–414. [Google Scholar]
- Nuttonson, M.Y. Rye–Climate Relationships and the Use of Phenology in Ascertaining the Thermal and Photo-Thermal Requirements of Rye–Based on Data of North America and of Some Thermally Analogous Areas of North America in the Soviet Union, Finland, Poland, and Czechoslovakia; American Institute of Crop Ecology: Washington, DC, USA, 1957; p. 219. [Google Scholar]
- Hartman, T.A. Influence of daylength on vernalization of winter rye. NJAS Wagening. J. Life Sci. 1964, 12, 132–155. [Google Scholar] [CrossRef]
- Zieliński, K.; Krzewska, M.; Żur, I.; Juzoń, K.; Kopeć, P.; Nowicka, A.; Moravčiková, J.; Skrzypek, E.; Dubas, E. The effect of glutathione and mannitol on androgenesis in anther and isolated microspore cultures of rye (Secale cereale L.). Plant Cell Tiss. Organ Cult. 2020, 140, 577–592. [Google Scholar] [CrossRef]
- Sue, M.; Yamazaki, K.; Yajima, S.; Nomura, T.; Matsukawa, T.; Iwamura, H.; Miyamoto, T. Molecular and structural characterization of hexameric β-d-glucosidases in wheat and rye. Plant Physiol. 2006, 141, 1237–1247. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rice, C.P.; Park, Y.B.; Adam, F.; Abdul-Baki, A.A.; Teasdale, J.R. Hydroxamic acid concentration and toxicity of rye at selected growth stages. J. Chem. Ecol. 2005, 31, 1887–1905. [Google Scholar] [CrossRef]
- La Hovary, C.; Danehower, D.A.; Ma, G.; Reberg-Horton, C.; Williamson, J.D.; Baerson, S.R.; Burton, J.D. Phytotoxicity and benzoxazinone concentration in field grown cereal rye (Secale cereale L.). Int. J. Agron. 2016. [Google Scholar] [CrossRef]
- Gierl, A.; Frey, M. Evolution of benzoxazinone biosynthesis and indole production in maize. Planta 2001, 213, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Zasada, I.A.; Meyer, S.L.F.; Halbrendt, J.M.; Rice, C. Activity of hydroxamic acids from Secale cereale against the plant-parasitic nematodes Meloidogyne incognita and Xiphinema americanum. Phytopathology 2005, 95, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.; Lathrop, R.G.; Obropta, C.C. Climate change vulnerability assessment and adaptation strategies through best management practices. J. Hydrol. 2020, 580, 124311. [Google Scholar] [CrossRef]
- Kitetu, G.M.; Ko, J.H. Climate Change on Agriculture in 2050: A CGE Approach. 2020. In Proceedings of the 23rd GTAP Annual Conference on Global Economic Analysis “Global Economic Analysis Beyond 2020”, Division of International & Area Studies, Busan, Korea, 17–19 June 2020. [Google Scholar]
- Wittchen, U.; Chmielewski, F.M. Phytoclimate of winter rye stands. Meteorol. Z. 2005, 14, 183–189. [Google Scholar] [CrossRef]
- Pomortsev, A.V.; Dorofeev, N.V.; Zorina, S.Y.; Katysheva, N.B.; Sokolova, L.G. The effect of planting date on Winter rye and triticale overwinter survival and yield in Eastern Siberia. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Siberian Institute of Plant Physiology and Biochemistry, Krasnoyarsk, Russia, 22 June 2019. [Google Scholar]
- Xie, Y.; Wang, C.; Yang, W.; Yang, W.; Feng, M.; Qiao, X.; Song, J. Canopy hyperspectral characteristics and yield estimation of winter wheat (Triticum aestivum) under low temperature injury. Sci. Rep. 2020, 10, 244. [Google Scholar] [CrossRef]
- Rodriguez-Algaba, J.; Sørensen, C.K.; Labouriau, R.; Justesen, A.F.; Hovmøller, M.S. Susceptibility of winter wheat and triticale to yellow rust influenced by complex interactions between vernalisation, temperature, plant growth stage and pathogen race. Agronomy 2020, 10, 13. [Google Scholar] [CrossRef]
- Kuwabara, C.; Imai, R. Molecular basis of disease resistance acquired through cold acclimation in overwintering plants. J. Plant Biol. 2009, 52, 19–26. [Google Scholar] [CrossRef]
- Gaudet, D.A.; Laroche, A.; Frick, M.; Davoren, J.; Puchalski, B.; Ergon, A. Expression of plant defence-related (PR-protein) transcripts during hardening and dehardening of winter wheat. Physiol. Mol. Plant Pathol. 2000, 57, 15–24. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).