Abstract
Climatic change, ecosystem imbalance, and soil salinization and desertification are serious obstacles to the restoration of degraded landscapes. Halophyte cultivation could constitute a way of mitigating these problems. Frankenia pulverulenta is used in the remediation and revegetation of areas affected by salinity and as an ornamental plant on saline soils since it can tolerate salt concentrations up to 200 mM NaCl. To increase saline tolerance, different plant growth regulators (auxins, cytokinins, gibberellins, spermidine, and salicylic acid) were tested in hydroponic conditions before the addition of NaCl (300 mM). At 52 days, growth, proline, saline excretion, free polyamines, and ethylene were determined under both saline and non-saline conditions. All growth regulators improved growth in the salt-free pretreatment; however, under conditions of salinity, pretreatment with spermidine (0.5 mM) and then salicylic acid (0.5 mM) were efficient at reversing the inhibitory effect of salt and improved saline excretion in F. pulverulenta. A strong positive correlation of polyamines and proline, and a negative correlation with ethylene, indicate that pretreatments that increase endogenous polyamine content and osmolytes are the most effective in improving salt tolerance of F. pulverulenta and could be used in the technical guidance of the cultivation of this halophyte.
Keywords:
growth; halophyte cultivation; phytohormones; polyamines; proline; salicylic acid; recretohalophyte 1. Introduction
Climate change (i.e., irregular rainfall and extreme temperatures), intensive agriculture, and the poor quality of irrigation water have provoked the degradation, salinization, and desertification of soils, and led to a loss of agriculture areas, lower crop yields and imbalances in ecosystems [1,2]. The search for alternatives that can reduce the pressure on agricultural crops to provide enough food, pharmaceutical products, energy, and other important products for the world’s population, as well as the restoration of degraded landscapes, should be a priority area of research in the twenty-first century [3]. Halophytes possess different strategies such as seed heteromorphism, dormancy, seed bank dynamics, and the ability to recover seed germination after exposure to high salinity and so improve the chances of seedling survival in diverse habitats [4,5]. These plants also have morphological (succulence, salt glands, trichomes or bladders), physiological, and bioquimic attributes (high K+/Na+ selectivity, Na+ compartmentalization that prevents ionic toxicity, osmolyte synthesis favoring osmotic adjustment, xanthophyll cycle involved in the dissipation of excess excitation energy in the PSII antenna as heat, protecting the photosynthetic machinery, water use efficiency, activation of antioxidant systems (enzymatic and non-enzymatic) to mitigate oxidative damage, and plant growth regulators that modulate transcription and translation, up-regulating antioxidants and osmolytes) that allow them to thrive under adverse conditions [6,7,8]. Thus, any increase in the use and production of halophytic crops will be an interesting line for future research.
In saline conditions, plant growth regulators (PGRs) might allow halophytes to better cope with environmental stress. Plant growth regulators are related to many aspects of plants’ life cycles. Phytohormones are structurally diverse compounds involved in many processes such as germination, growth and development, floral initiation, and fruit development and ripening. In terms of abiotic stress tolerance, they delay leaf senescence, control stomata conductance, regulate antioxidant enzymes, and act as signal molecules [9,10,11]. Auxins (IAA) and cytokinins (CKs) regulate cell expansion and cell cycles, chloroplasts, vascular tissue, apical dominance, leaf senescence, shoot differentiation, and photomorphogenic development. Gibberellic acid (GA) can lead to a significant increase in seed germination, stem elongation, leaf expansion, and flower and fruit development; however, little is known about the modulation of phytohormones in halophytes and their responses to abiotic stress [12,13]. The application of exogenous polyamines (PAs) to crops has recently been reviewed by Alcázar et al. [14], who concluded that these small amines (putrescine (Put), spermidine (Spd), spermine (Spm), and Cadaverine (Cad)) can regulate various physiological processes such as growth, biomass production, photosynthesis activity, osmolytes accumulation, redox homeostasis, and antioxidant defenses under abiotic stress. Salicylic acid (SA) is a signaling molecule involved in plant growth and development, and defense responses under stress conditions, and its exogenous application is used in many plant species to combat symptoms of toxicity induced by saline stress [15]. Nevertheless, little is known about the effect of PGR pretreatments on halophytes. Therefore, different PGRs (IAA, Kinetin, GA3, Spd, and SA) were applied during the cultivation of the halophyte F. pulverulenta to study their effect on this plant.
Frankenia pulverulenta L. (Fam. Frankeniaceae) is a small annual plant found in saline endorheic coastal, and continental habitats, being a good indicator of saline soils. It forms small fragile populations of plants with many-branched spreading stems reaching 15–20 cm, and very small leaves (5–7 mm × 0.7–3.7 mm) and flowers. It blooms from April to July. Fruit forms in a capsule with two to four valves and tiny seeds [16]. The maintenance of the water content of its aerial parts is aided by the morphological characteristics of its leaves (inwardly curling towards the abaxial surface) that reduce the loss of water by transpiration. Pérez-Cuadra and Cambi [17] studied the location of salt glands at epidermal levels, which are surrounded by radiated epidermal cells. The abaxial epidermis has eglandular trichomes, a morphological adaptation that allows excess salts to be excreted. A high content of proline, phenols, and polyamines were detected in leaves [18], which provide the plant with a certain degree of tolerance to saline stress. This halophyte grows well in a concentration of 200 mM NaCl [18] and is employed in phytoremediation and in revegetation projects in gardens [19,20], being an important economic source of the production of ornamental plants [21]. This halophyte is used for medicinal purposes (antiviral effect) and as a source of neuroprotective molecules for pharmaceutical and cosmetic use due its high content of secondary metabolites (phenols, flavonoids, and antioxidants) [22,23,24].
Although the application of PGRs is used to improve crop production, little is known about their use with halophytes. Frankenia pulverulenta (a recretohalophyte) was grown as a hydroponic culture in an irrigation solution with the following PGRs pretreatments: IAA, Kinetin, GA3, Spd, and SA for 10 days, and then NaCl (300 mM NaCl). After 21 days in saline and non-saline conditions, growth parameters (dry weight and water content), osmolyte accumulation, saline excretion, endogenous-free polyamines, and ethylene production were analyzed. Our aim was to explore which PGR pretreatments produce the best results in terms of the growth and physiological characteristics of F. pulverulenta under both saline and non-saline conditions. It is hoped that the results will provide technical guidance for increasing stress tolerance of these plants and the benefits they can bring.
2. Materials and Methods
2.1. Plant Material and Growth Conditions
Seeds of Frankenia pulverulenta L. present in Brujuelo saltmarsh (GPS location: 37°52′46″ N, 3°40′11″ W) (Jaén province, South Spain) were randomly collected in September 2016. The seeds were kept dry at 4 °C until use. Then, they were washed with sterile distilled water and sown in Petri dishes at 25 ± 1 °C with a 16 h photoperiod. After 10 days, the most uniform seedlings were transferred to 1.5 L pots containing a vermiculite substrate. Four seedlings were sown per pot and cultivated hydroponically using Hoagland nutrient solution 50% pH 6.5 ± 0.1 [25]. The experiments were conducted in a growth chamber under the following environmental conditions: photosynthetic photon flux density (PPFD) 500 μmol photon m−2 s−1, 400–700 nm, provided by Sylvania Cool White (Osram Sylvania Inc., Danvers, MA, USA), and Osram Dulux Superstar (Osram Sylvania Inc., Danvers, MA, USA) lamps, photoperiod 16 h/ 8 h for day/night cycle, temperature (day) 25 °C ± 1°C and (night) 16 °C ± 1 °C, and a relative humidity of 55–75%. The pots were distributed on the growing table at random. The plants were watered every two days with Hoagland’s nutrient solution, excess solution being removed to prevent the solution from pooling around the roots.
2.2. Experimental Design and Treatments
After the acclimation period (3 weeks in hydroponic conditions), plants were pretreated for 10 days with different growth regulators (PGRs) applied to the nutrient solution before irrigation. These PGRs were:
- -
- Indole-Acetic acid (1 μM)
- -
- Kinetin (1 μM)
- -
- Gibberellic acid (1 μM)
- -
- Spermidine (0.5 mM)
- -
- Salicylic acid (0.5 mM)
The PGR concentrations used were based on previous experimentations and information in the literature [15,26,27]. An experimental design of six treatments was established: (1) control (-PGRs); (2) indole-acetic acid (IAA); (3) kinetin (K); (4) gibberellic acid (GA3); (5) spermidine (Spd); and (6) salicylic acid (SA) with 12 pots per treatment. Subsequently, these pots were divided into two groups of six pots, to which two levels of NaCl were applied: 0 and 300 mM. The NaCl levels were selected according to the germination and growth trials conducted prior to this experiment. Salt concentrations were increased progressively to avoid osmotic shock until the required concentrations were reached [18]. Six pots with 24 plants were grown in free saline conditions, and six other pots with 24 plants were used in saline treatments. After 21 days of treatment, the plant materials were collected. The F. pulverulenta plants were 52 days old (10 days of PGR pretreatment after 21 days in saline or non-saline conditions).
2.3. Growth Parameters
All plants were used to calculate the fresh weight (FW) (roots, stems and leaves). Part of the plant material was placed in a forced-air oven at 70 °C for 96 h to determine the dry weight (DW), water content, osmolytes, and free PAs. The water percentage was calculated using the following formula: WC (%) = (FW − DW/FW) × 100, in SL (stem and leaves) and R (roots) [18]. Part of the fresh leaves were washed with distilled water (any soluble salt crystals were removed from the surface of the leaves and collected for ionic analysis), and afterwards dried with towels before the fresh weight was determined. Fresh leaves were also used to determine the ethylene production.
2.4. Determination of Proline
The method described by Bates et al. [28] was used to extract and quantify the proline (Pro): dry leaves (at 52 days of culture) were homogenized with 3% sulfosalicylic acid and centrifuged at 1000× g, the supernatant being used for the quantification of Pro. The endogenous Pro content was determined at 520 nm in a VARIAN spectrophotometer (Cary 4000 UV-VIS, Santa Clara, CA, USA). A standard curve with Pro was used for the final calculations.
2.5. Ion Content Measurement of Salt Crystals in Surface Leaves of F. pulverulenta
The aerial parts of the plants treated with salt (300 mM NaCl) were carefully and rapidly washed in distilled water to remove the soluble salt crystals and to estimate the excreted soluble salt. Na+ concentrations were measured using a flame photometer (Ciba Corning, Essex, UK), while Cl− was determined with a chloridometer analyzer (MKII Choride 926, Sherwood, UK) following the same method as used for other recretohalophytes [29].
2.6. Analysis of Free Polyamines
The method used by Bueno et al. [18] was followed. Briefly, dry material was homogenized in 4% (v/v) cold perchloric acid and centrifuged at 14,000× g for 35 min at 4 °C. Aliquots of the supernatant, and standard pattern (1–6-diaminohexane, cadaverine and putrescine (dihydrochloride), spermidine (trihydrochloride), and spermine (tetrahydrochloride)), were derivatized with dansyl chloride, extracted with toluene and analyzed by HPLC (high-performance liquid chromatography). Aliquots (20 μL) of the samples were injected into a reverse phase Spheri-5 C18 ODS (80 Å, 4.6 × 220 mm) column and eluted with a water–acetonitrile variable solvent gradient at a flow rate of 1.5 mL min−1. A Shimadzu (Kyoto, Japan) LC-10A HPLC equipped with a fluorescence spectrophotometer (excitation and emission wavelengths were 252 nm and 500 nm band pass) was used to quantify the dansyl derivatives. A relative calibration procedure was used to determine free polyamines in samples using 1–6-diaminohexane as an internal standard, and standard curves of putrescine, spermidine, spermine and cadaverine from Sigma.
2.7. Ethylene Production
The method used by Bueno et al. [30] was followed with minor modifications: fresh leaves (200 mg) were aseptically transferred to a 5 mL flask containing 50 μL of distilled water. The flask was sealed with a silicone-rubber stopper and incubated in darkness at 30 °C. After 1 h, a 1 mL gas sample was injected into a HP5890 (series II, Hewlett Packard, Palo Alto, CA, USA) gas chromatograph fitted with a flame ionization detector and a 2 m × 4 mm stainless-steel column packed with 50–80 mesh Poropack-R. The N2, H2, and synthetic air flow rates were 50, 86, and 400 mL min−1, respectively. Ethylene identification was based on the retention time compared with an ethylene (C2H4) standard (purity 99.9%).
2.8. Data Analysis
Analyses of variance (ANOVA) of the data were computed using a Statgraphics Centurion v. 17, provided by the University of Jaén. Significant differences between means were determined using Tukey’s multiple range test (p ≤ 0.05 and p ≤ 0.01). Pearson’s correlation coefficients were calculated using the parameters analyzed under saline and non-saline conditions.
3. Results
3.1. Effect of Pretreatment PGRs under Saline and Non-Saline Conditions
Growth in F. pulverulenta is stimulated at concentration of up to 200 mM NaCl. However, fresh, and dry weights decrease at 300 mM NaCl compared to the controls grown under non-saline conditions [18]. Thus, concentrations of 0 and 300 mM NaCl were chosen for growing this plant. Growth parameters after 52 days of culture are shown in Table 1. In pretreatments without salt, all PGRs increased the dry weight of the stem + leaves (SLDW) and roots (RDW). Significant differences (p ≤ 0.05) were found only in aerial parts (except for 1 μM IAA) (Table 1A), which indicates that there was a better plant growth. In terms of water content, all pretreatments exerted a positive effect, above all on roots (RWC), with Kinetin, Spd, and SA having the highest values (Table 1A). In pretreatments PGRs + NaCl (300 mM) (Table 1B), control, IAA, Kinetin, and GA3 all had values that were lower than those obtained for growth without salt (Table 1A). Nevertheless, pretreatments with Spd and SA significantly improved all studied parameters and even gave rise to dry weight values that were similar to those obtained in pretreatments without salt (Table 1A). The highest values were found in the pretreatment Spd (0.5 mM) + NaCl (300 mM), which increased SLDW by 56%, RDW by 44%, SLWC by 4%, and RWC by 17%, compared to plants treated only with salt (-PGRs + NaCl 300 mM). There was even an increase of 4% in RWC relative to untreated plants (-PGRs). In Figure 1, it can be observed that the best growth of F. pulverulenta was cultivated with Spd + salt and SA + salt.

Table 1.
(A) Effect of pretreatment PGRs (plant growth regulators) under non-saline conditions, and 1 (B) pretreatment PGRs with salt (300 mM NaCl) in Frankenia pulverulenta, at 52 days of culture on SLDW (stem + leaves dry weight), RDW (root dry weight), SLWC (stem + leaves water content), and roots water content (RWC). The values represent means ± SE (n = 16). Different letters next to column represent significant differences between treatments, according to Tukey’s test (p ≤ 0.05).

Figure 1.
Frankenia pulverulenta cultivated with Spd + salt compared to control (A), and F. pulverulenta cultivated with SA + salt compared to control (B).
3.2. Effect of Pretreatment PGRs under Saline and Non-Saline Conditions on Proline Accumulation
The osmolyte Pro increased under high salt concentrations [18]. Figure 2A shows the accumulation of Pro in F. pulverulenta leaves grown with pretreatment PGRs. The maximum values were observed with all PGRs, whose values increased 2.6-fold when compared to non-treated plants (no PGRS). The increase in Pro at 300 mM NaCl was 4.2-fold when compared to the values of Pro (no PGRs) (Figure 2A,B). Pretreatment (Spd + salt) and (SA + salt) resulted in higher values of Pro content (Figure 2B), and a 1.4-fold increase compared to plants grown only with salt.
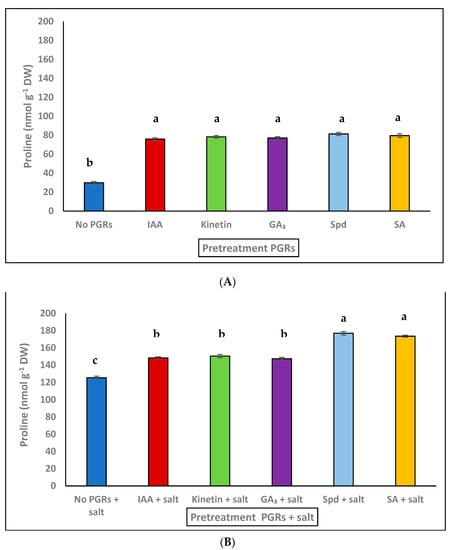
Figure 2.
(A) Proline content in F. pulverulenta leaves cultivated with pretreatment PGRs: IAA, Kinetin, GA3, Spd, and SA respect to control (no PGRs). (B) Proline content cultivated with PGRs + salt: IAA + NaCl, Kinetin + NaCl, GA3 + NaCl, Spd + NaCl and SA + NaCl, respect to control only with salt (NaCl 300 mM). Values represent means ± SE (n = 4). Different letters above bars represent significant differences between treatments (p ≤ 0.01).
3.3. Effects of Pretreatment PGRs + Salt on Ionic Content in the Leaf-Washing Solution in F. pulverulenta
Frankenia pulverulenta is a recretohalophyte that can be used in phytoremediation due to its saline glands that facilitate the excretion of salts from the plant. We aimed to discover whether any of the pretreatments improved saline excretion. Salt crystals were visible to the naked eye on the surface of leaves of plants grown with salt (300 mM) (Figure 3).

Figure 3.
Salt crystals on surface leaves of F. pulverulenta.
The mineral analyses of the washing solution showed that similar amounts of sodium and chloride were excreted to the amounts excreted in the respective controls for Na+ and Cl−. The exception was the pretreatment Spd (0.5 mM) and NaCl (300 mM), after which the increase in Na+ was 17% compared to the control (-PGRs and NaCl 300 mM), indicating that significant saline excretion occurred in this pretreatment (Table 2).
Table 2.
Sodium and chloride determination from the leaf washing solution expressed per gram of freshly rinsed leaves of F. pulverulenta in response to pretreatment PGRs with salt (300 mM NaCl), after 52 days of culture. The values represent means ± SE (n = 4). Different letters represent significant differences among pretreatments, according to Tukey’s test (p ≤ 0.01).
3.4. Effect of Pretreatment PGRs on Free Polyamine Content
In general, pretreatments Spd (0.5 mM) − salt and Spd (0.5 mM) + salt in F. pulverulenta gave the best growth results and physiological characteristics. Therefore, we decided to measure the endogenous content of free PAs cultivated with different PGRs under saline and non-saline conditions. The results are shown in Table 3. Putrescine, Spd, Spm, and Cad were detected, Spd being the triamine with the highest values under non-saline conditions, while Spd and Spm had the highest values under saline conditions. In all salt-free PGR pretreatments, Put, Spd, and Cad decreased compared to the control pretreatments (-PGRs) (Table 3A). Nevertheless, an increase in endogenous Spm (0.64-fold) was observed, especially in the Spd and SA (0.5 mM) pretreatments. Table 3B shows the results for the pretreatment PGRs + NaCl (300 mM), in which a significant increase (p ≤ 0.01) in Spd, Spm, and Cad was observed in all PGRs. Indeed, exogenous Spd and SA increased 0.38- (endogenous Spd) and 0.31- (endogenous Spm) fold compared to their respective controls. On the other hand, endogenous Spm and Cad increased one- and 12-fold, respectively (Table 3B) compared to results obtained from Spd (0.5 mM) without salt (Table 3A) pretreatments. Therefore, the saline treatments (300 mM NaCl) and exogenous application of PGRs and, above all, Spd increase the PA pool favor an increase in Spm that promotes tolerance to abiotic stress. Cadaverine levels also increased under saline conditions in the Spd and SA pretreatments, which indicated that a modulation of these amines occurred during the growth of F. pulverulenta.
Table 3.
(A). Effect of pretreatment PGRs − salt, and (B) PGRs + salt (300 mM NaCl) in F. pulverulenta leaves, at 52 days of culture, on free PAs content: putrescine (Put), spermidine (Spd), spermine (Spm) and cadaverine (Cad). The values represent means ± SE (n = 4). Different letters next to columns represent significant differences among treatments, according to Tukey’s test (p ≤ 0.01).
Total PAs (Put + Spd + Spm + Cad) and ethylene production (C2H4) on leaves of F. pulverulenta are shown in Table 4. The results indicate a marked decrease in ethylene but an overall increase in PA levels under saline conditions, which may indicate that this decrease in ethylene will contribute to an increase in endogenous PA content.
Table 4.
(A). Effect of pretreatment PGRs − salt, and (B) PGRs + salt (300 mM NaCl) in F. pulverulenta leaves on total PAs and ethylene production. The values represent means ± SE (n = 4). Different letters next to columns represent significant differences among treatments, according to Tukey’s test (p ≤ 0.01).
It interesting to note the negative correlation between Put and Pro (r = −0.77; p ≤ 0.01), ethylene and Pro (r = −0.91; p ≤ 0.01), and ethylene and Spm (r = −0.90; p ≤ 0.01), and the positive correlation between Spm and Pro (r = 0.97; p ≤ 0.01), Cad and Pro (r = 0.93; p ≤ 0.01), and total PA and Pro (r = 0.90; p ≤ 0.01) (Table 5).
Table 5.
Correlation coefficient among all parameters studied in F. pulverulenta cultivated in saline and non-saline conditions (p ≤ 0.05 *; p ≤ 0.01 **).
4. Discussion
Exogenously applied PGRs enhance growth and abiotic stress tolerance in numerous species [14,31,32]. The application of PGRs under saline conditions alleviates the inhibitory effects of salt on growth (increases in the root:shoot ratio), morphological (leaf area, dry mass) and physiological (stomatal conductance, photosynthetic rate) characteristics, and productivity [33]. This research could provide useful information for the application of these PGRs when cultivating the halophyte Frankenia pulverulenta for ornamental and phytoremediation purposes. We focused on the application of auxins, cytokinins, and gibberellins, little studied in work on abiotic stress tolerance, and Spd and SA, two growth regulators with positive effects on saline stress tolerance.
In the salt-free pretreatments, all PGRs increased their values for both dry mass and water content compared to untreated plants (Table 1A). Growth regulators act on multiple cellular and physiological processes of growth and plant development. More specifically, CKs stimulated cell elongation (shoot and roots), cell division, and up-regulate gene CK production to delay senescence. Auxins cooperate with CKs in the regulation of cell cycles and cell expansion, and especially in the formation of lateral roots. GAs are implicated in seed germination, stem elongation, and fruit development [34,35,36]. These findings coincide with the greater growth obtained in F. pulverulenta cultivated with PGRs. Plant growth regulators may also allow plants to adapt to saline stress. In pretreatment PGRs + salt, Spd had the greatest effect of all the PGRs on water content and dry mass, and thus on photosynthetic activity and growth. Polyamines offer specific protection to photosynthetic tissues and play a key role in the structural organization and functional activity of thylakoids in the chloroplast of stressed tissues [37]. Salicylic acid was also effective in promoting the growth of F. pulverulenta. It is well known that SA ameliorates toxicity symptoms provoked by saline stress. Nevertheless, the effect of exogenous SA on growth is dependent on concentrations (concentrations of 1 mM are excessively high), type of stress, time of exposure, and the plant species, which suggests that each halophyte species will respond differently to SA treatment and that this type of treatment may benefit some plants but not others [15]. In general, few studies of the effect of PGRs on halophytes have ever been conducted. One of most studied plants is the extreme halophyte Mesembryanthemum crystallinum, in which the application of CKs or GAs increases the expression of the key enzyme PEPCase, which is correlated with increases in proline and other osmolytes [13,38]. By contrast, in the halophyte Prosopis strombulifera, Kinetin application varies with different types of salt and high salinity increases the levels of this hormone [39]. Indole acetic acid is also implicated in the stress response of Suaeda fruticosa and Atriplex halimus, in which an increase in NaCl significantly increased IAA levels in the roots of both species. Conversely, in Prosopis strombulifera, low IAA levels were observed. Therefore, for each halophyte studied, it will be necessary to investigate which is the best pretreatment for stimulating growth and improving productivity [11,40].
In general, while glycophyte limit sodium uptake from saline soil to the plant, however, most halophytic dicots accumulate toxic ions in the plants’ shoots to increase osmotic pressure and maintain cell turgor in the leaves [7]. Frankenia pulverulenta is a dicotyledonous halophyte that accumulates toxic ions (Na+ and Cl−) in its aerial parts. Part of the salts are excreted through its saline glands while another part accumulates in the vacuoles, which allows cell turgor to be maintained in the leaf tissue [18]. For this reason, we focused our study on the Pro, PAs, saline excretion, and ethylene on leaves. Among other strategies, halophytes tolerate salinity by accumulating osmolytes such as Pro, glycine-betaine, sugars, polyols, and PAs that help stabilize subcellular structures and scavenging ROS (inhibiting lipid peroxidation of membranes), and buffer the cellular redox potential, which favors osmotic adjustment and water absorption. Proline is the most frequent osmolyte found in halophytes [41]. In leaves of F. pulverulenta, high levels of Pro were detected with increases in salt levels (especially at 300 mM NaCl) [18]. Pretreatments without salt increased Pro levels (Figure 2A). High Pro levels can serve as a nitrogen source during growth and development, and its metabolism is linked to several key pathways such as pentose phosphate and tricarboxylic acid and contributes to purine synthesis and the phenylpropanoid pathway [42]. In plants pretreated with salt (Figure 2B), Pro levels were higher than in the salt-free pretreatments, and Spd (0.5 mM) + NaCl (300 mM) and SA (0.5 mM) + NaCl (300 mM) had better growth and water content. The accumulation of Pro may represent a major physiological adaptation in plants for osmotic adjustment and homeostasis redox. Some PGRs such as PA increase N levels and endogenous Pro content [43]. Salicylic acid application also boots Pro levels by increasing nitrogen assimilation and improving photosynthesis [44].
In general, halophytes are used for multiple purposes, including transgenic approaches that can identify potential genes for improving salt tolerance in glycophytes [45]. In addition, halophyte cultivation can also be used for desalinating degraded soils and revegetation [46,47,48,49], biosaline agriculture [50,51], biofuel-producing crops [52], forages and fodder, and the production of secondary metabolites with medicinal uses [3]. Salt excretion is an important desalinization mechanism that can contribute to revegetation using native halophytes and the rehabilitation of salt-affected land. We studied the effect of PGR pretreatments + salt on the excretion of saline solution in F. pulverulenta (Table 2). Most PGR pretreatments had higher values for the excretion of Na+ and Cl− through their saline glands than the controls (-PGRs + salt). However, only pretreatment Spd (0.5 mM) + NaCl (300 mM) showed any significant increase in salt excretion. More in-depth studies are still needed if we are to determine whether the amount of salt excretion can reduce the salt content of soils and create conditions for glycophyte cultivation (salt-sensitive plants). Ben Hassine et al. [53] have proposed that PAs (especially Spd and Spm) could be involved in saline excretion mechanisms and reduce the influx of Na+ into mesophyll cells by redirecting it towards the organs of saline excretion, which may explain our results. Subsequently, Pottosin and Shabala [54] discovered that PAs are involved in the transport of cations through the tonoplast and plasma membranes, as well as the regulation of ion homeostasis, under saline conditions. Salt excretion activity is mediated by multi-cells and multi-genes, and several exist strategies have been identified [55]. Nevertheless, more study is still required to determine the potential of applying other recretohalophytes to saline land.
The best pretreatment used in the halophyte cultivates was Spd + salt, which prompted us to analyze the endogenous PA content. In this sense, ornithine and arginine are precursors of Put (diamine), while Spd (triamine) and Spm (tetraamine) are synthetized from Put via Spd synthase and Spm synthase, respectively, by the addition of the amino-propyl groups generated by S-adenosylmethionine decarboxylase from SAM (S-adenosyl-L-methionine). Spermidine, Spm, and ethylene share a common precursor [56,57]. Salt modifies the accumulation of PAs in F. pulverulenta leaves and increases Spd, Spm, and Cad levels, whereas in non-saline conditions, only endogenous Spm increases (Table 3A,B). Obviously, exogenous Spd increases the content of this triamine and favors the formation of Spm. The drop in ethylene could contribute to an increase in the total PA levels through SAM (Table 4). Therefore, competition for the SAM could be between ethylene and PA biosynthetic pathways. On the other hand, exogenous Spd application could increase PA biosynthetic enzyme levels without altering PA degrading enzymes, thereby favoring PA accumulation (Spd and Spm) as occurs in zoysiagrass subjected to saline stress [27]. We have previously analyzed the function of PAs in halophytes [58] and noted the key role played by this amine in protecting the photosynthetic apparatus and membranes, maintaining ion balance, mitigating oxidative stress, and reducing the accumulation of heavy metals in certain salt-tolerant species. Pretreatment with Spd (0.5 mM) + salt exerts a protective effect by improving growth and other physiological characteristics in F. pulverulenta. It is well known that PA applications and transgenic plants with over produced PAs played protective roles under abiotic stress conditions, while reduced PA levels in vivo result in lower stress tolerance [56,59,60]. All these studies lead us to highlight the protection against salt stress provided by PAs. On the other hand, salt always increases Cad levels in F. pulverulenta [18]. The diamine Cad (1.5-diaminopentane) is a lysine catabolite related to plant growth and development, and its levels always increase in plants when exposed to drought and salt stress [61,62]. Under stress conditions, Cad accumulation has been proposed to compensate for the decrease in the content of the Put-family PAs [63]. Based on the correlation matrix (Table 5) and the analyzed parameters, it is worth underlining the negative correlation between Put, Pro, and ethylene, and the positive correlation between Pro, Spm, Cad, and total PAs. This indicates that PGRs + salt modulate the levels of these amines, thereby favoring the mechanisms of homeostasis redox and antioxidant activities, key roles played by these amines. Our results confirm the potential application of PAs to improve growth and benefits of F. pulverulenta against stress.
5. Conclusions
Recretohalophyte culture can help reduce lower environmental temperatures, desalinize soils, and reestablish ecosystems. Exogenous PGR application stimulates growth and saline excretion in the halophyte F. pulverulenta and improves the physiological characteristics of this plant. The most efficient pretreatment in both non-saline and saline conditions is Spd. Spermidine application enhances stress tolerance up to 300 mM NaCl, by modulating cellular PA levels and has a demonstrable role in protecting against salinity. The pool of PAs is increased when adding exogenous Spd to nutritive solutions and favor the formation of Spm. Cadaverine is also increased and improves growth and mitigates the effects of saline stress. These PAs probably act via the maintenance of photosynthetic activity, the accumulation of osmolytes and the modulating of the homeostasis redox. In the case of SA, a positive effect was also observed in F. pulverulenta, as it mitigated the toxicity effect of saline stress, albeit to a lesser degree than with the PAs. Auxins, CKs and GAs stimulated growth less well than Spd and SA, above all under saline conditions. Therefore, PA application could further improve halophyte cultivation in saline and non-saline conditions and increase its potential uses in phytodesalination and as ornamental plant. Nevertheless, more work is still needed to determine which is the best pretreatment for stimulating growth and productivity in each halophyte considering the large number of applications that these plants have (phytoremediation, biosaline agriculture, biofuel-producing crops, forages, and medicinal uses).
Author Contributions
Conceptualization, M.d.P.C. and M.B.; methodology, M.d.P.C. and M.B.; software, M.d.P.C. and M.B.; validation, M.d.P.C. and M.B.; formal analysis, M.d.P.C. and M.B.; investigation, M.d.P.C. and M.B.; resources, M.B.; data curation, M.d.P.C. and M.B.; writing—Original draft preparation, M.B.; writing—Review and editing, M.B.; visualization, M.d.P.C. and M.B.; supervision, M.d.P.C. and M.B.; project administration, M.d.P.C.; funding acquisition, M.d.P.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by SPANISH MINISTRY OF SCIENCE AND INNOVATION, grant number CGL2006-08830, The involvement of growth regulators in plant response to salinity).
Acknowledgments
We want to thank to designer Sonia Reyes for her technical support to achieve the high-quality tables and figures presented in this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thornton, P.K.; Ericksen, P.J.; Herrero, M.; Challinor, A.J. Climate variability and vulnerability to climate change: A review. Glob. Chang. Biol. 2014, 20, 3313–3328. [Google Scholar] [CrossRef] [PubMed]
- Leisner, C.P. Review: Climate change impacts on food security-focus on perennial cropping systems and nutritional value. Plant Sci. 2020, 293, 110412. [Google Scholar] [CrossRef]
- Panta, S.; Flowers, T.; Lane, P.; Doyle, R.; Haros, G.; Shabala, S. Halophyte agriculture: Success stories. Environ. Exp. Bot. 2014, 107, 71–83. [Google Scholar] [CrossRef]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Bueno, M. Adaptation of halophytes to different habitats. In Seed dormancy and Germination, 1st ed.; Jiménez-López, J.C., Ed.; Intech Open: London, UK, 2020; Volume 1, pp. 97–119. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Fernanda Ortuño, M.; Bernal-Vicente, A.; Díaz-Vivancos, P.; Sánchez-Blanco, M.J.; Hernández, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Kumari, A.; Das, P.; Parida, A.K.; Agarwal, P.K. Proteomics, metabolomics, and ionomics perspectives of salinity tolerance in halophytes. Front. Plant Sci. 2015, 6, 537. [Google Scholar] [CrossRef]
- Bueno, M.; Cordovilla, M.P. Ecophysiology and uses of halophytes in diverse habitats. In Handbook of Halophytes: From Molecular to Ecosystems towards Biosaline Agriculture, 1st ed.; Grigore, M.N., Ed.; Springer Nature: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Torre-González, A.; Navarro-León, E.; Albacete, A.; Blasco, B.; Ruíz, J.M. Study of phytohormone profile and oxidative metabolism as key process to identification of salinity response in tomato commercial genotypes. J. Plant Physiol. 2017, 216, 164–173. [Google Scholar] [CrossRef]
- Zheng, L.; Ma, H.; Jiao, Q.; Ma, C.; Wang, P. Phytohormones: Important participators in plant salt tolerance. Int. J. Agric. Biol. 2020, 24, 319–332. [Google Scholar] [CrossRef]
- Szepesi, A. Halotropism: Phytohormonal aspects and potential applications. Front. Plant Sci. 2020, 11, 571025. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Llanes, A.; Reginato, M.; Devinar, G.; Luna, V. What is known about phytohormones in halophytes? A review. Biologia 2018, 73, 727–742. [Google Scholar] [CrossRef]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small amines with large effects on plant abiotic stress tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic acid in plant salinity stress signalling and tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Consejería de Medio Ambiente. Flora Vascular de Andalucía Oriental, 1st ed.; Junta de Andalucía: Sevilla, Spain, 2009. [Google Scholar]
- Pérez Cuadra, V.; Cambi, V. Morphoanatomical functional traits in xerophytic species of a saline environment. Int. J. Exp. Bot. 2014, 83, 389–396. [Google Scholar]
- Bueno, M.; Lendínez, M.L.; Calero, J.; Cordovilla, M.P. Salinity responses of three halophytes from inland saltmarshes of Jaén (southern Spain). Flora 2020, 266, 151589. [Google Scholar] [CrossRef]
- Easton, I.C.; Kleindorfer, S. Effects of salinity levels and seed mass on germination in Australian species of Frankenia L. (Frankeniaceae). Environ. Exp. Bot. 2009, 65, 345–352. [Google Scholar] [CrossRef]
- Veldkornet, D.A.; Potts, A.J.; Adams, J.B. The distribution of salt marsh macrophyte species in relation to physicochemical variables. S. Afr. J. Bot. 2016, 107, 84–90. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Velasquez Espino, C.; Lao, M.T. Effects of leachate fertigation and the addition of hydrogen peroxide on growth and nutrient balance in Dracaena deremensis potted plants. Agronomy 2021, 11, 127. [Google Scholar] [CrossRef]
- Ben Sassi, A.; Harzallah-Skhiri, F.; Bourgougnon, N.; Aouni, M. Antiviral activity of some Tunisian medicinal plants against Herpes simplex virus type 1. Nat. Prod. Res. 2008, 22, 53–65. [Google Scholar] [CrossRef]
- Lopes, A.; Rodrigues, M.J.; Pereira, C.; Oliveira, M.; Barreira, L.; Varela, J.; Trampetti, F.; Custódio, L. Natural products from extreme marine environments: Searching for potential industrial uses within extremophile plants. Ind. Crop Prod. 2016, 94, 299–307. [Google Scholar] [CrossRef]
- Ben Mansour, R.; Wided, M.K.; Cluzet, S.; Krisa, S.; Richard, T.; Ksouri, R. LC-MS identification and preparative HPLC isolation of Frankenia pulverulenta phenolics with antioxidant and neuroprotective capacities in PC12 cell line. Pharm. Biol. 2017, 55, 880–887. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Bull. 1950, 347, 1–32. [Google Scholar]
- Tounekti, T.; Hernández, I.; Müller, M.; Khemira, H.; Munné-Bosch, S. Kinetin applications alleviate salt stress and improve the antioxidant composition of leaf extracts in Salvia officinalis. Plant Physiol. Biochem. 2011, 49, 1165–1176. [Google Scholar] [CrossRef]
- Li, S.; Jin, H.; Zhang, Q. The effect of exogenous spermidine concentration polyamine metabolism and salt tolerance in Zoysiagrass (Zoysia japonica Steud) subjected to short-term salinity stress. Front. Plant Sci. 2016, 7, 1221. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Zouhaier, B.; Najla, T.; Abdallah, A.; Wahbi, D.; Wided, C.; Chedly, A.; Abderrazak, S. Salt stress response in the halophyte Limoniastrum guyonianum Boiss. Flora 2015, 217, 1–9. [Google Scholar] [CrossRef]
- Bueno, M.; Lendínez, M.L.; Aparicio, C.; Cordovilla, M.P. Effect of salinity on polyamines and ethylene in Atriplex prostrata and Plantago coronopus. Biol. Plant 2015, 59, 596–600. [Google Scholar] [CrossRef]
- Amir, R.; Munir, F.; Khan, M.; Iqbal, T. Use of plant hormones for the improvement of plant growth and production under salt stress. In Salt Stress, Microbes, and Plant Interactions: Causes and Solution, 2nd ed.; Akhtar, M.S., Ed.; Springer Nature: Singapore, 2019; Volume 1, pp. 59–90. [Google Scholar]
- Morsi, M.M.; Abdelmigid, H.M.; Aljoudi, N.G.S. Exogenous salicylic acid ameliorates the adverse effects of salt stress on antioxidant system in Rosmarinus officinalis L. Egypt. J. Bot. 2018, 58, 249–263. [Google Scholar] [CrossRef]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; García-Sánchez, F. Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Rivero, R.M.; Gimeno, J.; Van Deynze, A.; Walia, H.; Blumwald, E. Enhanced cytokinin synthesis in tobacco plants expressing PSARK::IPT prevents the degradation of photosynthetic protein complexes during drought. Plant Cell Physiol. 2010, 51, 1929–1941. [Google Scholar] [CrossRef]
- Pearce, S.; Huttly, A.K.; Prosser, I.M.; Li, Y.D.; Vaughan, S.P.; Gallova, B.; Phillips, A.L. Heterologous expression, and transcript analysis of gibberellin biosynthetic genes of grasses reveals novel functionality in the GA3ox family. Plant Biol. 2015, 15, 130. [Google Scholar] [CrossRef]
- Ashfaq, M.; Khan, S. Role of phytohormones in improving the yield of oilseed crops. In Oils Seed Crops: Yield and Adaptation under Environmental Stress, 1st ed.; Ahmad, P., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; Volume 9, pp. 165–183. [Google Scholar] [CrossRef]
- Ghabriche, R.; Ghnaya, T.; Mnasri, M.; Zaier, H.; Baioui, R.; Vromman, D.; Abdelly, C.; Lutts, S. Polyamine and tyramine involvement in NaCl-induced improvement of Cd resistance in the halophyte Inula chrithmoides L. J. Plant Physiol. 2017, 216, 136–144. [Google Scholar] [CrossRef]
- Thomas, J.C.; Bohnert, H.J. Salt stress perception and plant growth regulators in the halophyte Mesembryanthemum crystallinum. Plant Physiol. 1993, 103, 1299–1304. [Google Scholar] [CrossRef][Green Version]
- Llanes, A.; Bertazza, G.; Palacio, G.; Luna, V. Different sodium salts cause different solute accumulation in the halophyte Prosopis strombulifera. Plant Biol. 2013, 15, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Llanes, A.; Pinamonti, P.; Iparraguirre, J.; Bertazza, G.; Luna, V. Abscisic acid alters carbohydrate accumulation induced by differential response to sodium salts in the halophyte Prosopis strombulifera. Plant Biosyst. 2019, 154, 337–347. [Google Scholar] [CrossRef]
- Tipirdamaz, R.; Gagneul, D.; Duhazé, C.; Aïnouche, A.; Monnier, C.; Özkum, D.; Larher, F. Clustering of halophytes from an inland salt marsh in Turkey according to their ability to accumulate sodium and nitrogenous osmolytes. Environ. Exp. Bot. 2006, 57, 139–153. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A.; Iqbal, M.; Khan, R. A new perspective of phytohormones in salinity tolerance: Regulation of proline metabolism. Environ. Exp. Bot. 2014, 100, 34–42. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Singh Sidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef]
- Pathak, M.R.; Teixeira, J.A.; Wani, S.H. Polyamines in response to abiotic stress tolerance through transgenic approaches. GM Crop. Food 2014, 5, 87–96. [Google Scholar] [CrossRef]
- Shaygan, M.; Mulligan, D.; Baumgartl, T. The potential of three halophytes (Tecticornia pergranulata, Sclerolaena longicuspis, and Frankenia serpyllifolia) for the rehabilitation of brine-affected soils. Land Degrad. Dev. 2018, 29, 2002–2014. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhowmik, P.C.; Hossain, M.A.; Rahman, M.M.; Vara Prasad, M.N.; Ozturk, M.; Fujita, M. Potential use of halophytes to remediate saline soils. BioMed Res. Int. 2014, 2014, 589341. [Google Scholar] [CrossRef]
- Jesus, J.M.; Danko, A.S.; Fiúza, A.; Borges, M.T. Phytoremediation of salt-affected soils: A review of processes, applicability, and the impact of climate change. Environ. Sci. Poll. Res. 2015, 22, 6511–6525. [Google Scholar] [CrossRef]
- Atzori, G. The potential of edible halophytes as new crops in saline agriculture. The ice plant (Mesembryanthemum crystallinum L.) case study. In Future of Sustainable Agriculture in Saline Environments, 1st ed.; CRC Press: Boca Raton, NY, USA, 2021; pp. 443–460. [Google Scholar] [CrossRef]
- Nikalje, G.C.; Srivastava, A.K.; Pandey, G.K.; Suprasanna, P. Halophytes in biosaline agriculture: Mechanism, utilization, and value addition. Land Degrad. Dev. 2018, 29, 1081–1095. [Google Scholar] [CrossRef]
- Duarte, B.; Caçador, I. Iberian halophytes as agroecological solutions for degraded lands and biosaline agriculture. Sustainability 2021, 13, 1005. [Google Scholar] [CrossRef]
- Christiansen, A.H.C.; Lyra, D.A.; Jørgensen, H. Increasing the value of Salicornia bigelovii green biomass grown in a desert environment through biorefining. Ind. Crop Prod. 2021, 160, 113105. [Google Scholar] [CrossRef]
- Ben Hassine, A.; Ghanem, M.E.; Bouzid, S.; Lutts, S. Abscisic acid has contrasting effects on salt excretion and polyamine concentrations of an inland and a coastal population of the Mediterranean xero-halophyte species Atriplex halimus. Ann. Bot. 2009, 104, 925–936. [Google Scholar] [CrossRef]
- Pottosin, I.; Shabala, S. Polyamines control of cation transport across plant membranes: Implications for ion homeostasis and abiotic stress signaling. Front. Plant Sci. 2014, 5, 154. [Google Scholar] [CrossRef]
- Yuan, F.; Leng, B.; Wang, B. Progress in studying salt secretion from the salt glands in recretohalophytes: How do plants secrete salt? Front. Plant Sci. 2016, 7, 977. [Google Scholar] [CrossRef]
- Liu, J.H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef]
- Majumdar, R.; Barchi, B.; Turlapati, S.A.; Gagne, M.; Minocha, R.; Long, S.; Minocha, S.C. Glutamate, ornithine, arginine, proline, and polyamine metabolic interactions: The pathway is regulated at the post-transcriptional levels. Front. Plant Sci. 2016, 7, 78. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.; Cordovilla, M.P. Polyamines in halophytes. Front. Plant Sci. 2019, 10, 439. [Google Scholar] [CrossRef] [PubMed]
- Minocha, R.; Majumdar, R.; Minocha, S.C. Polyamines and abiotic stress in plants: A complex relationship. Front. Plant Sci. 2014, 5, 175. [Google Scholar] [CrossRef]
- Pál, M.; Szalai, G.; Janda, T. Speculation: Polyamines are important in abiotic stress signaling. Plant Sci. 2015, 237, 16–23. [Google Scholar] [CrossRef]
- Tomar, P.C.; Lakra, N.; Mishra, S.N. Effect of cadaverine on Brassica juncea (L.) under multiple stress. Indian J. Exp. Biol. 2013, 51, 758–763. [Google Scholar]
- Jancewicz, A.L.; Gibbs, N.M.; Masson, P.H. Cadaverine’s functional role in plant development and environmental response. Front. Plant Sci. 2016, 7, 870. [Google Scholar] [CrossRef]
- Reginato, M.; Abdala, G.I.; Miersch, O.; Ruíz, O.A.; Moschetti, E.; Luna, V. Changes in the levels of jasmonates and free polyamines induced by Na2SO4 and NaCl in roots and leaves of the halophyte Prosopis strombulifera. Biologia 2012, 67, 689–697. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).