Abstract
The reduction in biodiversity owing to agricultural intensification has brought negative effects on sustainable crop production in the agro-environment. Plant diversity can help regulate insect pests; however, research that demonstrates that genotypic diversity of multiple varieties with prominent agroecological practices to suppress insect pests and benefit crop yield at agricultural landscapes is limited. To examine the effects of diversity of wheat varieties on aphids and crop productivity, we performed field experiments by designing plots of single (resistant/susceptible) and cultivar mixtures in 2018–2020. The effects of population abundance of cereal aphids (Sitobion miscanthi and Rhopalosiphum padi) and crop yield on resistant and susceptible wheat varieties and mixture of wheat cultivars were determined to reveal the most dominant mixed broadcasting mode in Kaifeng of China. The results showed that cultivar mixture significantly decreased aphid abundance compared with the single varieties (resistant/susceptible). Cultivar mixture had a higher yield compared with the single varieties (resistant/susceptible). Additionally, cultivar mixture had significantly higher thousand-grain weights than susceptible and resistant wheat varieties in 2019 and 2020. Cultivar mixture enhanced wheat yield by decreasing cereal aphids, indicating a bottom-up or top-down effect from genetic diversity to pest abundance. Our results indicated that appropriate cultivar mixtures could manage insect pests to some extent and stabilize crop yield.
1. Introduction
Wheat is a staple food consumed by 35% of the world’s population, and it occupies an important position in the grain production of China []. Pest damage can cause an average of 15% reduction in crop production; cereal aphids, corn borers, and rice planthoppers all have unfavorable effects on crop yield [,]. Farmers strongly depend on pesticides and chemical fertilizers to improve the yield and quality of crops and use vast monocultures [,]. The long-term use of chemical inputs has caused ecological destruction and reduction in ecosystem function and has also led to high levels of pesticide resistance and resurgence of insect pests []. In monoculture landscapes, the reduction in crop diversity owing to agricultural intensification has increased the vulnerability of crops to abiotic and biotic stresses and necessitated the increased dependence on agrochemical inputs [,]. Moreover, the need for food security and production worldwide are becoming increasingly prominent owing to the growing population and increased food consumption []. Therefore, other farming practices are needed to ensure pest management and safe food production for sustainable environmental development [].
Biodiversity has been shown to have the potential of insect management through both improved bottom-up and top-down effects in forest ecosystems [] and agroecosystems []. Biodiversity includes mixing different cultivars of a crop species (genotypic diversity) or mixing different species of plants (species diversity) by intercropping, cover cropping, or using living mulches []. Compared to species diversity, genotypic diversity can regulate pest populations more simply, requiring few extra changes in farm practices or economic investments [,], which would have more potential to be adopted by conventional growers. Contrary to the widespread application of cultivar mixtures to effectively suppress key pathogens in developed and developing countries [,], limited studies have been performed on pest control using genotypic diversity. Some researchers have highlighted the importance of cultivar mixtures in reducing herbivorous pest abundance [,]. Mixtures of maize varieties significantly decreased population densities of the corn leafhopper Dalbulus maidis, versus monocultures of a single variety []. Similarly, laboratory-based studies demonstrated that wheat cultivar mixtures reduced pest infestation and damages []. Moreover, aphids that belong to Hemipterans share more attributes of pathogens than other insect species, so aphids may be more vulnerable to the effect of cultivar mixtures with different susceptibility to crop diseases, and it has been reported that cereal aphids limit the production of wheat in China. Therefore, it is reasonable to take wheat and cereal aphids as the research object to explore the effects of cultivar mixtures with different susceptibility to crop diseases on insect pests and crop yield [,].
Biodiversity can help agroecosystems improve tolerance to biotic stressors for better management of insect pests, and could be incorporated into crop production [,,]. Cultivar mixtures help in the regulation of abiotic stressors and plant biomass, and an abundance of floral visitors and pollination services may also benefit yield and yield stability in agricultural fields [,]. Plant genotypic diversity can increase adaptation to unpredictable environmental variability and resilience to agroecosystems compared to monocultures []. Genotypic diversity is confirmed to have greater resource-use efficiency on light, CO2, soil, water [], and more root biomass to buffer water deficits than monocultures, improving their tolerance to biotic and abiotic stress factors, and, therefore, may increase yield [,,]. A meta-analysis of 32 selected studies showed that the yield of wheat increased by 3.2% per added component variety under high disease pressure []. However, some reviews have reported that the yield of cultivar mixtures is equivalent [] or significantly lower than that of monoculture []. More studies focused on the effects of crop cultivar mixtures on disease suppression and have revealed the positive relationship with crop yield [,,]; however, limited studies and applications have been performed on wheat pest control and its possible correlation with wheat yield.
In this study, we conducted two-year field experiments to study how wheat cultivar mixtures with different susceptibility to wheat stripe rust disease (Puccinia striiformis f. sp. tritici) affect the naturally occurring populations of the dominant cereal aphids: Sitobion miscanthi (Hemiptera: Aphididae) and Rhopalosiphum padi Linnaeu. The effect of cultivar mixtures with different susceptibility to wheat stripe rust disease on the wheat performance of thousand-grain weights and wheat yield were also evaluated in this study. Our intention was to (1) investigate the population dynamics of cereal aphids with different wheat cultivar mixtures and its possible relationship with wheat performance on a large scale and (2) determine whether wheat cultivar mixtures could have a relationship with high yield and high resistance to pests, which provides a viable and sustainable management strategy using low chemical inputs.
2. Materials and Methods
2.1. Plants and Experimental Design
Field experiments were carried out using plots of five winter wheat genotypes grown from October 2018 to June 2020 at the Kaifeng Experimental Station of China Agricultural University, Henan Province, Northern China (34°46′ N, 114°16′ E). Five winter wheat cultivars with different susceptibility to Puccinia striiformis f. sp. tritici were chosen for this study: Miannong4 (high susceptibility), Mianyang31 (middle susceptibility), Mianmai1403 (high resistance), Chuanmai55 (high resistance), and Rongmai4 (high resistance). The winter wheat (Triticum aestivum cv. Zhou 22) was sown in the buffer zones and bare ground between plots and blocks. There were three treatments in this study; they were susceptible, resistant, and cultivar mixture (combination of resistant and susceptible). The cultivar mixture comprised two susceptible and three resistant varieties. There were eight replications for each treatment, indicating that 24 plots were used totally in our study. Completely randomized block experiments were conducted in a 27 × 68 m sized field. Each plot was 6 × 6 m, with 3 m buffer zones, 2 m wide bare ground between plots, and 3 m wide bare ground between blocks. Seeds were mixed completely with equal proportions of each cultivar, maintaining the same density of treatment, before seeding. Winter wheat was sown in October each year and harvested in early June 2019 or mid–late May 2020. The field was fertilized with urea (12.5 kg/ha/year), compound fertilizer (40 kg/ha/year), and farm manure (178 kg/ha/year), which functioned as nitrogen, phosphate, and potash source for wheat growth and grain tiller. Compound fertilizers and farm manure were applied in October, and in the following March urea was added to enhance soil fertility level and organic compost. The flood irrigation system was used to provide water to support the growth of wheat four times a year (on 6 November, the following 4 March, 7 April, and 7 May, respectively). No fungicides, pesticides, or herbicides were applied on the wheat during the experiments due to low chemical input farming regulations.
2.2. Aphid Abundance
A five-point sampling method (5 tillers at each point) was used to investigate aphid densities and count the total number of S. miscanthi and R. padi in 5 tillers at each point. Five points within the plot were chosen randomly. All experimental plots were sampled at a six-day interval from mid-April to the end of April, three samples each year.
2.3. Wheat Performance
Three days before wheat maturity, the spike was selected by a five-point sampling method (5 tillers at each point) in each plot to weigh. After being harvested on 2–3 June in 2019 and 23–24 May in 2020, the wheat yield and grain weight of the plants were determined after air-drying in the sun for about five days, and thousand-grain weights (TGWs) were calculated using a hundred-grain counting board for each treatment with three replications.
2.4. Rust Disease Survey
At the jointing stage, the stripe rust races CYR32, CYR33, and CYR34 (1:1:1) were artificially inoculated. Mingxian 169 was planted in each plot center as the induction center. Slides were placed at different distances from each plot to the disease center, and 3 replicates were set with 3–10 sampling points. Spore sampler and disease assessment were carried out several times after onset. The number of diseased leaves and severity were recorded, and the severity evaluation criteria were divided according to the size of summer spore heap in leaf area, namely 0, 1%, 5%, 10%, 20%, 40%, 60%, 80%, and 100%. Then the incidence and disease index were calculated using the formula from Gu [].
2.5. Statistical Analysis
All statistical analysis was performed using SPSS V 26.0.0.0. A one-way ANOVA was used to compare the influences of various mixtures of wheat on aphid population numbers, thousand-grain weights, and wheat yield in 2019 and 2020, respectively.
3. Results
3.1. Aphid Populations
The aphid abundance in 2019 was significantly higher in the susceptible wheat treatment than in other treatments, and no significant difference was found in aphid abundance between the resistant wheat treatment and the 5-cultivar mixture at first and second investigations. As the wheat grew, no significant difference was found in aphid abundance among the three treatments (Figure 1A). The average aphid abundance was significantly higher in the susceptible wheat treatment than in the other treatments, and there was no significant difference found in the average aphid abundance between the resistant wheat treatment and the 5-cultivar mixture (Figure 1B).
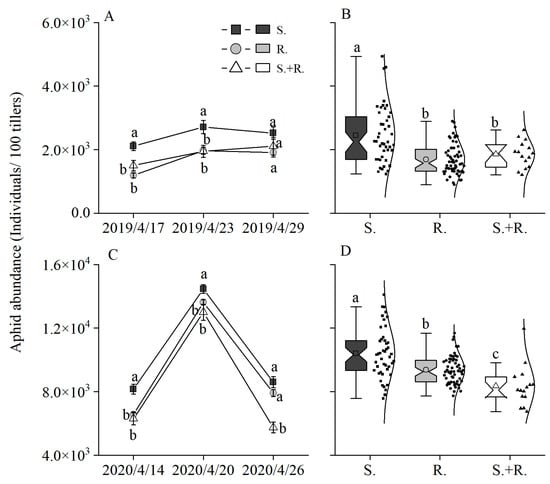
Figure 1.
Effect of genotypic diversity of wheat (S: susceptible varieties; R: resistant varieties; S+R: susceptible and resistant varieties) on aphid abundance (A,C) and average aphid abundance (B,D) in April 2019 and April 2020. Significantly different means within each treatment are indicated by different letters according to the SNK (Student–Newman–Keuls) test: (A) 2019/4/17 (F2, 117 = 20.182, p < 0.001); 2019/4/23 (F2, 117 = 6.758, p = 0.002); 2019/4/29 (F2, 117 = 3.518, p = 0.033); (B) (F2, 117 = 17.046, p < 0.001); (C) 2020/4/14 (F2, 117 = 14.201, p < 0.001); 2020/4/20 (F2, 117 = 6.134, p = 0.003); 2020/4/26 (F2, 117 = 12.111, p < 0.001); (D) (F2, 117 = 18.214, p < 0.001).
In 2020, the aphid abundance was significantly higher in the susceptible wheat treatment than in the other treatments; however, aphid abundance did not differ significantly between the resistant wheat treatment and the 5-cultivar mixture at first and second investigations. As the wheat grew, there was no significant difference between the susceptible and resistant wheat treatments; however, the 5-cultivar mixture showed significantly lower aphid abundance compared with susceptible or resistant wheat treatment (Figure 1C). The average aphid abundance in the 5-cultivar mixture was the lowest among all the treatments; it was significantly lower than that in the susceptible or resistant wheat treatment, and the average aphid abundance in the resistant wheat treatment was significantly lower than that in the susceptible wheat treatment (Figure 1D).
3.2. Wheat Performance
In 2019, TGWs of the 5-cultivar mixture were significantly higher than that in other treatments; however, no significant difference was found in TGWs between the resistant wheat treatment and the susceptible wheat treatment (Figure 2). In 2020, the TGWs of resistant wheat were significantly lower than that of other treatments, and TGWs of 5-cultivar mixture were significantly higher than that of the susceptible wheat treatment (Figure 2).
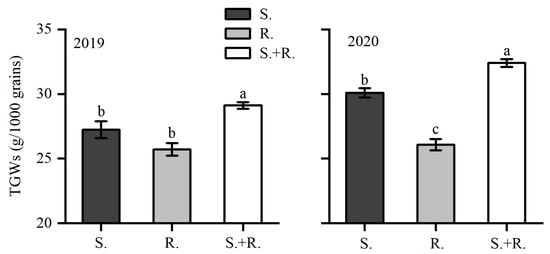
Figure 2.
Thousand-grain weights (TGWs) of wheat of susceptible varieties (S.), resistant varieties (R.), and susceptible and resistant varieties (S+R: 5-cultivar mixture) grown in 2019 and 2020. Significantly different means within each treatment are indicated by different lowercase letters according to the SNK test: 2019 (F2, 45 = 8.076, p = 0.001); 2020 (F2, 45 = 51.002, p < 0.001).
In 2019 and 2020, wheat yield did not show significant differences among all the treatments. In 2020, the yield of 5-cultivar mixture was 30.3% higher than that of the resistant wheat treatment and 5.7% higher than that of the susceptible wheat treatment. In 2019, the yield of 5-cultivar mixture was 42.5% higher than that of the resistant wheat treatment and 21.6% higher than that of the susceptible wheat treatment (Figure 3).
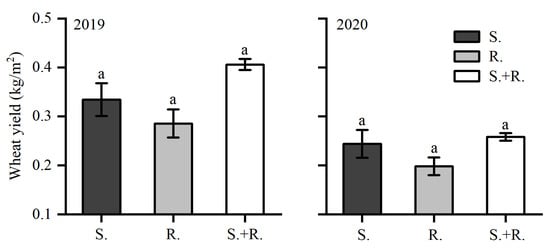
Figure 3.
Yield of wheat of susceptible varieties (S.), resistant varieties (R.), and susceptible and resistant varieties (S+R: 5-cultivar mixture) grown in 2019 and 2020. Not significantly different means within each treatment are indicated by the same lowercase letters according to the SNK test: 2019 (F2, 13 = 3.071, p > 0.05); 2020 (F2, 13= 2.021, p > 0.05).
3.3. Rust Disease Survey
In Kaifeng, the estimated AUDPC (the area under the disease progress stairs) of stripe rust susceptible varieties in the field was 123.87, actual AUDPC of resistant varieties was 0.42, estimated AUDPC of susceptible and resistant varieties was 50.15, actual AUDPC of susceptible and resistant varieties was 22.03, and the relative control efficacy was 56% (Table 1).

Table 1.
Rust disease survey.
4. Discussion
Genotypic diversity considerably suppressed aphid abundance, corroborating the report that genotypic diversity could enhance the resistance of crop to biotic pressure []. Plants synthesize defensive metabolites to cope with herbivores attacks, and genotypic diversity further improves the resistance of plants to herbivores []. The cultivar mixtures in this study, which comprised the susceptible and resistant varieties, harbored fewer aphids under high pest pressure, than the single variety, probably owing to the volatiles emitted by the different plants in the mixture that enhanced the defenses against aphids and transmitted information to the neighboring undamaged plants, thus changing the movement of the aphid. It had been shown that R. padi has low tolerance to volatiles from a mixture, corroborating the reallocation of biomass resources of barley plants exposed to undamaged plants of different cultivars in mixtures [,]. Moreover, volatile interactions between different species of the same crop are more attractive to the ladybugs and further regulate the aphids []. The different effects of genotype diversity on aphid regulation during the two years might be due to the increased attraction to natural enemies at high aphid density and the change in aphid migration at different population levels. Therefore, there is a need to conduct a long-term field study, based on years, with different pest densities and a proper combination of cultivars.
In this study, cereal aphid dynamics varied significantly with years. The annual variation in aphid population numbers was associated with temperature according to the 24 years long time-series analysis of aphid abundance on winter wheat by Brabec []. The number of aphids in April 2020 was more than four times that in April 2019. The higher temperature in 2020 improved the overwintering survival rate of cereal aphids and the earlier spring immigration relieved the limitation of the length of aphid presence in wheat, thus lengthening the time of aphid damage and increasing the density of aphids in spring and summer. The outbreaks of cereal aphids were also found in the northern United States due to the higher winter temperature in 1991, which increased the number of aphid overwintering []. Moreover, the rainfall in April 2019 in Kaifeng was significantly higher than that in April 2020. The moderate rain was negatively correlated with the aphid peak in late April and the rainfall could have strongly reduced the aphid population on wheat. Therefore, long-term dynamics observation studies of cereal aphids are needed to investigate the abundance of aphids from different years and predict the aphid dynamics more precisely through corresponding climatic conditions and accumulated statistics for many years.
In our study, the TGWs of wheat varied with different treatments. The lower TGWs of resistant wheat treatments might be correlated with the increased defense resources to aphid attack and corresponding reduced resources for growth. The highest TGWs of mixtures might be associated with more light interception and higher photosynthesis efficiency owing to the increased number of cultivars in the mixtures [,]. The effects of genotypic diversity on wheat yield were similar among different treatments and years. There was no significant yield advantage in the mixed planting of winter wheat, probably owing to the small number of mixed planting varieties. Consistent with our hypothesis, some studies have shown that greater yield advantages were more likely to occur when the number of cultivars in the mixtures increased by more than two [,]. Thus, further study is needed to investigate the underlying mechanisms of genotypic diversity effects on wheat performance and utilize cultivar mixtures properly.
Unexpectedly, more aphid populations were observed in the disease-susceptible varieties than the other varieties while the susceptible wheat varieties had a relativity high yield; this outcome is a deviation from a previous study that suggested that some varietal blends might be endowed with additional superior attributes and offered notable yield advantage over susceptible cultivars []. Herbivores usually have a negative influence on crop yield; the opposite results gained in this study might be due to the relatively high level of aphid feeding that led to the self-compensation effect of the two susceptible wheat varieties, which stimulated the growth of wheat. It is possible that the relatively low yield of high resistance cultivars was related to the increased synthesis of metabolites for defense response against aphid feeding and the corresponding reduction in resources for growth. Moreover, the yields of the treatments in 2020 were lower than the average value of each combination in 2019, indicating that the number of cereal aphids was four times more than that in 2019, corroborating the study that showed that crop production was reduced by high pest pressure that was more than the economic threshold []. The number of aphids on susceptible wheat varieties was high and the yield of wheat was relatively higher compared to when the number of aphids on resistant wheat varieties was relatively low and the yield of wheat was lower. The five-cultivar mixtures composed of two susceptible varieties and three resistant varieties showed relatively lower aphid density, highest TGWs, and increased biodiversity. Although the yield of wheat cultivar mixtures was lower than that of wheat with pesticides, the price of green food could be higher than the purchase price of common wheat. Therefore, it is recommended to consider more than two cultivars in mixtures with the appropriate proportion of susceptible and resistant varieties. Moreover, breeding approaches are needed to create cultivar mixtures that possess increased abilities for the variance of relevant traits and foster complementarity and selection effects, meeting the demand for pest management and yield advantage [].
5. Conclusions
Continuous agricultural intensification and agricultural expansion have greatly affected biodiversity and ecological conditions owing to the pressure of an increasing population globally, leading to great challenges in the pursuit of increasing crop yield and protecting the environment in the future. The use of pesticides will reach at least 1.5 times the present amount by 2050 if there are no measures to change the current situation [] and reduce the diversity and quantity of beneficial insects such as natural enemies and pollinators and increase the economic burden of growers. Phytophagous insects were predicted to remain on host plants growing in pure stands []. As a potential agronomic measure to control pests and increase yield, a deeper understanding of the mechanisms of cultivar mixtures on pest regulation and yield increase is needed. Long-term field experiments are needed to determine the mechanism and appropriate ratio of cultivar mixtures to reduce the uncertainty caused by frequent variability in field tests, optimizing the efficiency of genotypic diversity and minimizing the change of agricultural measures by reducing chemical input cost and increasing the amount of safe food for sustainable agricultural development.
Author Contributions
Conceptualization, F.G. and Z.Z.; formal analysis, X.D., F.G. and Z.Z.; investigation, X.D., M.F., B.C. and Z.M.; methodology, F.G. and Z.Z.; writing (original draft), X.D. and S.P.; writing (review and editing), F.G. and Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
National Natural Science Foundation of China, Grant Number: 31770453.
Data Availability Statement
The data presented in this study are available within the article.
Acknowledgments
We thank Rui Wang and Qi Zhao at the Experimental Station of China Agricultural University in Kaifeng for their assistance with the harvesting of wheat and allocation of workers. The present study is supported by the National Natural Science Foundation of China (grant number: 31770453).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Adu-Gyamfi, P.; Mahmood, T.; Trethowan, R. Can wheat varietal mixtures buffer the impacts of water deficit? Crop Pasture Sci. 2015, 66, 757–769. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.Y.; Wang, L.Y.; Reddy, G.V.P.; Zhao, Z.H. Mild drought facilitates the increase in wheat aphid abundance by changing host metabolism. Ann. Entomol. Soc. Am. 2021, 114, 79–83. [Google Scholar] [CrossRef]
- Jin, F.; Wang, J.; Shao, H.; Jin, M.J. Pesticide use and residue control in China. J. Pestic. Sci. 2010, 35, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.L.; Zhang, F.X.; Liu, C.; Yu, H.; Cao, B.G.; Tian, S.Q.; Liao, Y.C.; Siddique, K.H.M. Wheat yield improvements in China: Past trends and future directions. Field Crops Res. 2015, 177, 117–124. [Google Scholar] [CrossRef]
- Robertson, G.P.; Swinton, S.M. Reconciling agricultural productivity and environmental integrity: A grand challenge for agriculture. Front. Ecol. Environ. 2005, 3, 38–46. [Google Scholar] [CrossRef]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef]
- Tilman, D.; Fargione, J.; Wolff, B.; D’Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting agriculturally driven global environmental change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Grettenberger, I.M.; Tooker, J.F. Moving beyond resistance management toward an expanded role for seed mixtures in agriculture. Agric. Ecosyst. Environ. 2015, 208, 29–36. [Google Scholar] [CrossRef]
- Andow, D.A. Vegetational diversity and arthropod population response. Annu. Rev. Entomol. 1991, 36, 561–586. [Google Scholar] [CrossRef]
- Tooker, J.F.; Frank, S.D. Genotypically diverse cultivar mixtures for insect pest management and increased crop yields. J. Appl. Ecol. 2012, 49, 974–985. [Google Scholar] [CrossRef]
- Barot, S.; Allard, V.; Cantarel, A.; Enjalbert, J.; Gauffreteau, A.; Goldringer, I.; Lata, J.C.; Le Roux, X.; Niboyet, A.; Porcher, E. Designing mixtures of varieties for multifunctional agriculture with the help of ecology. Agron. Sustain. Dev. 2017, 37, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.Y.; Chen, H.R.; Fan, J.H.; Wang, Y.Y.; Li, Y.; Chen, J.B.; Fan, J.X.; Yang, S.S.; Hu, L.P.; Leung, H.; et al. Genetic diversity and disease control in rice. Nature 2000, 406, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Mundt, C.C. Use of multiline cultivars and cultivar mixtures for disease management. Annu. Rev. Phytopathol. 2002, 40, 381–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Power, A.G. Leafhopper response to genetically diverse maize stands. Entomol. Exp. Appl. 1988, 49, 213–219. [Google Scholar] [CrossRef]
- Koricheva, J.; Hayes, D. The relative importance of plant intraspecific diversity in structuring arthropod communities: A meta-analysis. Funct. Ecol. 2018, 32, 1704–1717. [Google Scholar] [CrossRef] [Green Version]
- Shoffner, A.V.; Tooker, J.F. The potential of genotypically diverse cultivar mixtures to moderate aphid populations in wheat (Triticum aestivum L.). Arthropod Plant Interact. 2013, 7, 33–43. [Google Scholar] [CrossRef]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef]
- Kaczmarek, S.; Matysiak, K. Wheat cultivars, their mixtures and reduced herbicide doses as a practical solution in integrated weed management. Rom. Agric. Res. 2017, 34, 363–370. [Google Scholar]
- Vidal, T.; Boixel, A.L.; Durand, B.; de Vallavieille-Pope, C.; Huber, L.; Saint-Jean, S. Reduction of fungal disease spread in cultivar mixtures: Impact of canopy architecture on rain-splash dispersal and on crop microclimate. Agric. For. Meteorol. 2017, 246, 154–161. [Google Scholar] [CrossRef]
- Genung, M.A.; Lessard, J.P.; Brown, C.B.; Bunn, W.A.; Cregger, M.A.; Reynolds, W.N.; Felker-Quinn, E.; Stevenson, M.L.; Hartley, A.S.; Crutsinger, G.M.; et al. Non-additive effects of genotypic diversity increase floral abundance and abundance of floral visitors. PLoS ONE 2010, 5, e8711. [Google Scholar] [CrossRef]
- Qin, X.L.; Li, Y.Z.; Shi, C.X.; Song, D.P.; Wen, X.X.; Liao, Y.C.; Siddique, K.H.M. The number of cultivars in varietal winter-wheat mixtures influence aboveground biomass and grain yield in North China. Plant Soil 2019, 439, 131–143. [Google Scholar] [CrossRef]
- Jump, A.S.; Marchant, R.; Penuelas, J. Environmental change and the option value of genetic diversity. Trends Plant Sci. 2009, 14, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xu, B.C.; Liu, L.; Gu, Y.J.; Liu, Q.Q.; Turner, N.C.; Li, F.M. Does a mixture of old and modern winter wheat cultivars increase yield and water use efficiency in water-limited environments? Field Crops Res. 2014, 156, 12–21. [Google Scholar] [CrossRef]
- Borg, J.; Kiaer, L.P.; Lecarpentier, C.; Goldringer, I.; Gauffreteau, A.; Saint-Jean, S.; Barot, S.; Enjalbert, J. Unfolding the potential of wheat cultivar mixtures: A meta-analysis perspective and identification of knowledge gaps. Field Crops Res. 2018, 221, 298–313. [Google Scholar] [CrossRef]
- Dai, J.; Wiersma, J.J.; Nolen, D.L. Performance of hard red spring wheat cultivar mixtures. Agron. J. 2012, 104, 17–21. [Google Scholar] [CrossRef]
- Kiaer, L.P.; Skovgaard, I.M.; Ostergard, H. Grain yield increase in cereal variety mixtures: A meta-analysis of field trials. Field Crops Res. 2009, 114, 361–373. [Google Scholar] [CrossRef]
- Cox, C.M.; Garrett, K.A.; Bowden, R.L.; Fritz, A.K.; Dendy, S.P.; Heer, W.F. Cultivar mixtures for the simultaneous management of multiple diseases: Tan spot and leaf rust of wheat. Phytopathology 2004, 94, 961–969. [Google Scholar] [CrossRef] [Green Version]
- Gigot, C.; de Vallavieille-Pope, C.; Huber, L.; Saint-Jean, S. Using virtual 3-D plant architecture to assess fungal pathogen splash dispersal in heterogeneous canopies: A case study with cultivar mixtures and a non-specialized disease causal agent. Ann. Bot. 2014, 114, 863–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.L.; Chu, B.Y.; Wang, C.C.; Li, L.F.; Zhou, Y.L.; Luo, Y.; Ma, Z.H. Spore concentrations of Blumeria graminis f. sp. tritici in relation to weather factors and disease development in Gansu, China. Can. J. Plant Pathol. 2020, 42, 52–61. [Google Scholar] [CrossRef]
- Power, A.G. Virus spread and vector dynamics in genetically diverse plant-populations. Ecology 1991, 72, 232–241. [Google Scholar] [CrossRef]
- Kessler, A.; Halitschke, R. Specificity and complexity: The impact of herbivore-induced plant responses on arthropod community structure. Curr. Opin. Plant Biol. 2007, 10, 409–414. [Google Scholar] [CrossRef]
- Ninkovic, V.; Olsson, U.; Pettersson, J. Mixing barley cultivars affects aphid host plant acceptance in field experiments. Entomol. Exp. Appl. 2002, 102, 177–182. [Google Scholar] [CrossRef]
- Ninkovic, V. Volatile communication between barley plants affects biomass allocation. J. Exp. Bot. 2003, 54, 1931–1939. [Google Scholar] [CrossRef] [Green Version]
- Glinwood, R.; Ahmed, E.; Qvarfordt, E.; Ninkovic, V.; Pettersson, J. Airborne interactions between undamaged plants of different cultivars affect insect herbivores and natural enemies. Arthropod Plant Interact. 2009, 3, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Brabec, M.; Honek, A.; Pekar, S.; Martinkova, Z. Population dynamics of aphids on cereals: Digging in the time-series data to reveal population regulation caused by temperature. PLoS ONE 2014, 9, e106228. [Google Scholar] [CrossRef]
- Archer, T.L.; Johnson, G.D.; Peairs, F.B.; Pike, K.S.; Kroening, M.K. Effect of plant phenology and climate on Russian wheat aphid (Homoptera: Aphididae) damage to winter wheat. Environ. Entomol. 1998, 27, 221–231. [Google Scholar] [CrossRef]
- Wall, G.W.; Kanemasu, E.T. Carbon dioxide exchange rates in wheat canopies. Part I. Influence of canopy geometry on trends in leaf-area index, light interception and instantaneous exchange-rates. Agric. For. Meteorol. 1990, 49, 81–102. [Google Scholar] [CrossRef]
- Mundt, C.C.; Brophy, L.S.; Schmitt, M.S. Disease severity and yield of pure-line wheat cultivars and mixtures in the presence of eyespot, yellow rust, and their combination. Plant Pathol. 1995, 44, 173–182. [Google Scholar] [CrossRef]
- Newton, A.C.; Ellis, R.P.; Hackett, C.A.; Guy, D.C. The effect of component number on Rhynchosporium secalis infection and yield in mixtures of winter barley cultivars. Plant Pathol. 1997, 46, 930–938. [Google Scholar] [CrossRef]
- Vera, C.L.; Fox, S.L.; DePauw, R.M.; Smith, M.A.H.; Wise, I.L.; Clarke, F.R.; Procunier, J.D.; Lukow, O.M. Relative performance of resistant wheat varietal blends and susceptible wheat cultivars exposed to wheat midge, Sitodiplosis mosellana (Gehin). Can. J. Plant Sci. 2013, 93, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Cui, H.Y.; Chang, X.Y.; Zhu, M.M.; Zhao, Z.H. Increased nitrogen fertilization inhibits biocontrol activity promoted by the intercropping partner plant. Insect Sci. 2021, 28, 1179–1190. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).