Abstract
The stem branch trait of the stolon enables the common wild rice to produce new individuals through vegetative reproduction habit. In order to understand the genetic mechanism of stem branch character, we developed introgression lines (ILs) with the irrigated rice variety Yundao1 (YD1, Oryza sativa) as the recipient parent and Yuanjiang common wild rice (YJCWR, O. rufipogon) as the donor parent for subsequent identification of the relevant genes. An IL named IL-J85 was selected, which can originate new individuals from stem branches on stem nodes. Furthermore, the newly formed individuals can survive cutting to bear normally and produce the same yield per plant as IL-J85, which saved the growth time and production cost. Two QTLs (quantitative traits loci) related to the stem branch trait, qSBR1 and qSBR5, were first mapped on chromosomes 1 and 5. The near isogenic lines NIL-Y37 and NIL-D1 in the background of Yunjing 37 and Dianjingyou 1 were cultivated, showing the same characteristics as IL-J85. Our results provide new insights into the underlying genetic mechanism of the stem branch trait in the common wild rice and have the value of breeding utilization using vegetative reproduction to fix heterosis and breed new rice varieties with the cutting characteristic.
1. Introduction
As an annual crop and one of the three major food crops, rice (Oryza sativa) provides staple food for 3 billion people worldwide and plays an essential role in world food security [1,2]. Its importance is even more evident in China due to its long history of cultivation, given that it is the most dominant staple food for over 60% of the Chinese [3]. The traditional cultivation modes require sufficient irrigation water and have caused the serious nutrient loss. Moreover, labor costs, seeds, and other production costs keep rising, which affect people’s enthusiasm and motivation to cultivate perennial rice [4]. Recently, researchers placed efforts into using the perennial habit of wild rice varieties to breed perennial rice/upland rice and fixing heterosis through vegetative reproduction to increase harvest area and reduce production costs and soil erosion, thereby protecting the environment [4,5].
Different perennial characteristics were performed in the species of Oryza. The common wild rice (O. rufipogon) grows perennially with stolons. However, O. longistaminata, O. rhizomatis, and O. australiensis grow permanently with rhizomes. Both O. longistaminata and O. rufipogon with the typical perenniality have the same AA genome as O. sativa, which are considered ideal donors for transplanting the perennial habit [6]. Rhizomes determine the perennial growth of O. longistaminata and are controlled by one pair of dominant complementary genes, Rhz2 and Rhz3, located on chromosomes 3 and 4, respectively [5]. Further studies have showed that a very complex gene regulatory network mediated the growth and development of rhizomes in O. longistaminata [7]. O. rufipogon is the ancestor of O. sativa [8], and there are almost no reproductive disorders between them. It is sensitive to photoperiod and has a high outcrossing rate, preferring to live near swamps, lakes, and ditches with perennial habit [9]. O. rufipogon has a prostrate growth for many years by branching stolons and functional roots to form new individuals and finally achieving perennial survival through vegetative reproduction [10].
Various important agronomical traits were selected during the domestication of cultivated rice, including seed shattering, erect plant architecture, panicle shape, awn length, grain size and quality, and hull color [11]. Some key genes controlling rice domestication are well-understood, including the shattering gene Sb1 and qSH1 [12,13]; awn length gene An-1, An-2, and RAE2 [14,15,16,17]; hull color gene Bh4 [18]; pericarp color gene Rc [19]; panicle shape and ligule development gene OsLG1 [20]; grain width gene GW5 [21,22]; and tiller angle gene PROG1 [23,24]. The selection and domestication of these genes resulted in dramatic morphological changes in cultivated rice in Asia, from wild varieties adapted to natural environments to cultivated varieties adapted to agricultural environments. In addition, about 40% of wild rice alleles, including perennial alleles in the process of domestication from the wild rice to the cultivated rice, were lost [25], which resulted in narrowing of the genetic basis of cultivated varieties and further restricted improvement and breakthrough of yield potential of rice varieties. Genetic and molecular analyses of the perennial habit of O. rufipogon are the basis for breeding utilization and also help reveal the origin and domestication process of cultivated rice. Creeping growth gene PROG1 and photoperiod sensitive gene Hd1 and Ehd1 at the heading stage were associated with the perennial habit of O. rufipogon. However, the formation of new individuals by stem branches and adventitious roots on stolons and the critical part of the perennial habit, is little known.
Tiller, the shoot branch in rice which is closely related to rice yield, has been extensively studied, and the formation process of tiller has been elucidated [26,27], including MOC1 [28], MOC3/TABLE1 [29,30], TAD1/TE [31,32], OSH1 [30], LAX1, and LAX2 [33,34], which influence tiller bud initiation. Furthermore, D27 [35], OsMAX1 [36], OsMADS57 [37], D53 [38], OsOTUB1 [39], IPI1 [40], IPA1 [41], OsTB1 [42,43], and DLT [44] have been demonstrated to participate in tiller bud outgrowth. Phytohormones also regulate tiller bud initiation and outgrowth. In rice, strigolactones (SLs), auxin, gibberellins (GAs), and abscisic acid (ABA) can repress bud outgrowth, whereas brassinosteroids (BRs) and cytokinins (CKs) promote bud outgrowth [45].
The above studies mainly focus on the tiller, a lateral shoot arising from an axillary bud at basal nodes in rice. However, the genetic and molecular mechanism of the stem branch, a lateral shoot arising from an axillary bud at high nodes aboveground, remains elusive. Herein, we investigated the stem branch trait of an introgression line, a cross between cultivated rice (O. sativa) and its wild relative, O. rufipogon. The introgression line formed new individuals from the branches on stem nodes with adventitious roots, which was the same as the O. rufipogon. Furthermore, these individuals can survive by cutting, bear normally, and produce the same yield per plant with the introgression line. This study will provide theoretical basis and practical value for rice breeding using the stem branch trait to fix heterosis and cultivate new rice varieties of vegetative reproduction.
2. Materials and Methods
2.1. Plant Materials and Natural Field Experiment Conditions
The japonica paddy rice varieties, including Yundao1 (YD1), Yunjing37 (YJ37), and Dianjingyou1 (DJY1), were used as the recurrent parents. The common wild rice (YJCWR) collected from Yuanjiang county, Yunnan Province, China was used as the donor parent. IL-J85 was an introgression line from the cross between YD1 and YJCWR. NIL-Y37 and NIL-D1 (BC3F5) were the near-isogenic lines by crossing/backcrossing between IL-J85 and YJ37, respectively. C-J85, C-Y37, and C-D1 were the cuttings from the stem branches of IL-J85, NIL-Y37, and NIL-D1, respectively. S-J85, S-Y37, and S-D1 were the seed seedlings of IL-J85, NIL-Y37, and NIL-D1, respectively. BC3F2 and BC3F3, derived from the cross between YD1 and IL-J85 were cultivated to map the QTLs responsible for the stem branch trait.
All rice materials used for agronomic analysis were cultivated at the breeding base of the Xishuangbanna Botanical Garden, Chinese Academy of Sciences (21°56′ N, 101°15′ E) under natural field conditions twice a year in the spring and fall of 2021. The seeds were soaked for 48 h, and then germinated in a growth chamber at 28 °C for 24 h, subsequently sown on seedbeds. The seed seedlings were transplanted to paddy field at the four-leaf stage with 6 rows per plot and 12 plants per row. The density was 25 cm × 20 cm between each individual. For the breeding utilization of the stem branch trait, about 10 days before harvest of the NILs in the previous season, the seeds of the NILs were sowed normally on the seedling bed. Furthermore, the stem branches were cut and collected in 15 days later after harvest of the NILs, and then the cuttings (C-Y37 and C-D1) were placed in water for 3 days to enable the growth of the adventitious root. The seed seedlings and cuttings of NILs were transplanted to the paddy field simultaneously. The seed seedlings and cuttings of IL-J85 were treated and planted as mentioned above. To better measure phenotypes, all rice materials were planted in random three repetitions. The water in the paddy field was covered 2–5 cm above the soil before the rice seeds matured and supplemented with the compound NPK fertilizer (2 kg/100 m2).
2.2. Phenotypes Analysis
The materials were harvested in the center of each plot in each line to investigate the phenotypes. For the parent YD1 and IL-J85, the agronomic traits, including plant height, primary panicle length, flag leaf length, flag leaf width, flag leaf length–width ratio, the number of panicles, primary branches per panicle, secondary branches per panicle, 1000-grain weight, grain yield per plant, and internode length were investigated for 30 individuals. The number of stem branches and tillers per plant were conducted in 15 days later after harvest. For the analysis in the breeding utilization of the stem branch trait, the agronomic traits, including heading date, plant height, primary panicle length, flag leaf length, flag leaf width, the number of panicles, primary branches per panicle, secondary branches per panicle, 1000-grain weight, and grain yield per plant, were investigated for 30 individuals of the seed seedling and cutting of corresponding ILs/NILs respectively. Each experiment was performed for three biological repetitions. All rice seeds for phenotypes analyses were air-dried. Grain weight was measured with a Yield-Traits Scorer (YTS-5DS).
2.3. DNA Extraction
Rice genomic DNA from leaves was obtained according to the method described by Abdel-Latif et al. [46].
2.4. Mapping of qSBR1 and qSBR2
We hybridized IL-J85 with YD1 to obtain the BC3F1. The BC3F1 generation all performed the stem branch trait. BC3F2 was obtained from the selfing of BC3F1. We used 210 individuals of the BC3F2 segregating population to map qSBR1 and qSBR5 regions based on the number of stem branches per plant. A total of 100 recombinant individuals from BC3F3 segregating populations were used to fine map the locus of the stem branch trait through the targeted sequencing genotype detection technology of 10 K SNP chip (Shijiazhuang Molbreeding Biotechnology Co., Ltd., Shijiazhuang, China).
2.5. Statistical Analysis
QTL IciMapping 4.1 software was used to construct a genetic linkage map based on the genotypes and phenotypes, and Excel 2007 and SPSS 22 were used for data analysis. All samples were subjected to three replicate measurements. The data were presented as the mean ± standard deviation (SD). Student’s t-test or one/two-way ANOVA was performed to determine the significant difference, as indicated by * and ** at p < 0.05 and p < 0.01, respectively.
3. Results
3.1. Phenotypic Analysis of YD1 and IL-J85
The common wild rice (O. rufipogon; YJCWR) was collected from Yuanjiang County, Yunnan Province, China (Figure 1a,b). Yundao1 (O. sativa; YD1) a paddy japonica rice variety in Yunnan, was bred by Yunnan Academy of Agricultural Sciences (and Figure 1c). In order to explore the excellent traits from YJCWR, we constructed a set of introgression lines (ILs, BC2F8) with YJCWR as the donor parent and YD1 as the recurrent parent. Among these ILs, one introgression line IL-J85, showed stem branches at high nodes aboveground 15 days later after harvest, but the stem branch trait did not occur before maturity compared with YD1 (Figure 1c,d). We found that the stem could form more branches after crushing artificially or submerged naturally, which was similar to the trait of YJCWR (Figure 1e). These results suggest that the stem branch trait of introgression line IL-J85 was inherited from YJCWR.

Figure 1.
The phenotype of YJCWR, YD1, and IL-J85. (a,b) The phenotype of YJCWR. (c) YD1 and IL-J85 in the mature period. (d) IL-J85 15 days later after harvest. (e) IL-85 after crushing artificially or submerged naturally; the red arrow indicated the stem branches. Bar = 10 cm.
We conducted a comparative test on the main agronomic traits between IL-J85 and its recurrent parent, YD1. The results showed that plant height (PH), primary panicle length (PL), flag leaf length (FL), flag leaf width (FW), primary branches per panicle (PP), and secondary branches per panicle (SP) of IL-J85 decreased, but the number of panicles (NP) increased compared with those of YD1; whereas, there was no difference between YD1 and IL-J85 in flag leaf length–width ratio (FL/FW) (Table 1). We further analyzed (1st–5th) the internode length (INL) from top to bottom, and found that INL2, INL3, and INL4 of IL-J85 were significantly shorter than YD1, while INL1 and INL5 of IL-J85 had no significant difference from that of YD1 (Table 1). In addition, the number of stem branches per plant (TB) and tillers number per plant (TP) of two parents after harvest were investigated, and it was found that both TB and TP of IL-J85 were significantly higher than YD1 (Table 1). YD1 could produce a small amount of tillers after harvest, but did not produce stem branches which was different from IL-J85. The 1000-grain weight and grain yield per plant (GY) had no significant difference between YD1 and IL-J85, indicating that the stem branch trait after harvest did not affect the yield in IL-J85 compared with that in YD1 (Table 1). These results suggest that the stem branch trait of IL-J85 have the potential for breeding practice.

Table 1.
Investigation of agronomic traits in YD1 and IL-J85.
The number of stem branches (TB) was significantly correlated with the number of panicles (NP); grain yield per plant (GY) were significantly correlated with NP and primary panicle length (PL), but not correlated with flag leaf related traits (Table 2); TB also affected plant architecture traits, including PH, PL, and flag leaf related traits, indicating that there may be one pleiotropic gene or linkage inheritance.
Table 2.
Correlative coefficients among different agronomic traits of IL-J85.
3.2. Genetics Analysis of Stem Branch Trait in IL-J85
To carry out a genetic analysis of the stem branch trait in IL-J85, we hybridized IL-J85 with the recurrent parent YD1 to obtain the BC3F1 population (Figure 2a). All of the BC3F1 generation showed a similar stem branch trait to IL-J85. Thus, we speculated that the stem branch trait in IL-J85 was effectively dominant. To further assess the characteristics of phenotypical variation in the stem branch trait of IL-J85, the BC3F2 population were obtained from the selfing of BC3F1. We found the distribution in the BC3F2 population based on the number of stem branches per plant presented a multimodal distribution (Figure 2b). These findings indicated that major QTLs controlled the stem branch trait.
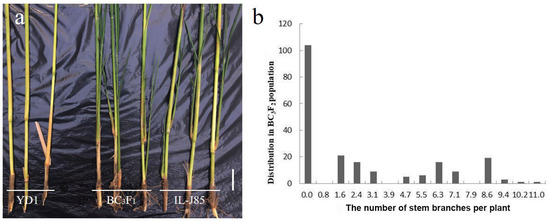
Figure 2.
The phenotype of BC3F1 in the mature period and the distribution in BC3F2 population. (a) YD1, BC3F1, and IL-J85 15 days later after harvest (bar = 10 cm). (b) The distribution in BC3F2 population based on the number of stem branches per plant (N = 210).
3.3. QTL Mapping for Stem Branch Trait
In order to map the stem branch trait, a total of 452 SSR markers, relatively uniformly distributed throughout the rice genomes, were used to assess polymorphism between IL-J85 and YD1. Sixteen SSR markers were polymorphic and distributed on nine chromosomes as nine segments except for chromosomes 2, 4, and 10 in IL-J85, indicating that they were introgressed from CWRYJ. To evaluate whether a QTL locus of stem branch was harbored in these nine chromosome regions, the genotypes of 210 individuals from the BC3F2 population were investigated using the 16 polymorphic SSR markers. The loci on chromosomes 1 and 5 had significant effects, whereas the effects of other introgressed segments were barely significant (Table 3). These results suggest that these two introgressed regions were responsible for the stem branch trait.
Table 3.
One-way ANOVA of stem branch presence or absence using marker genotypes as groups.
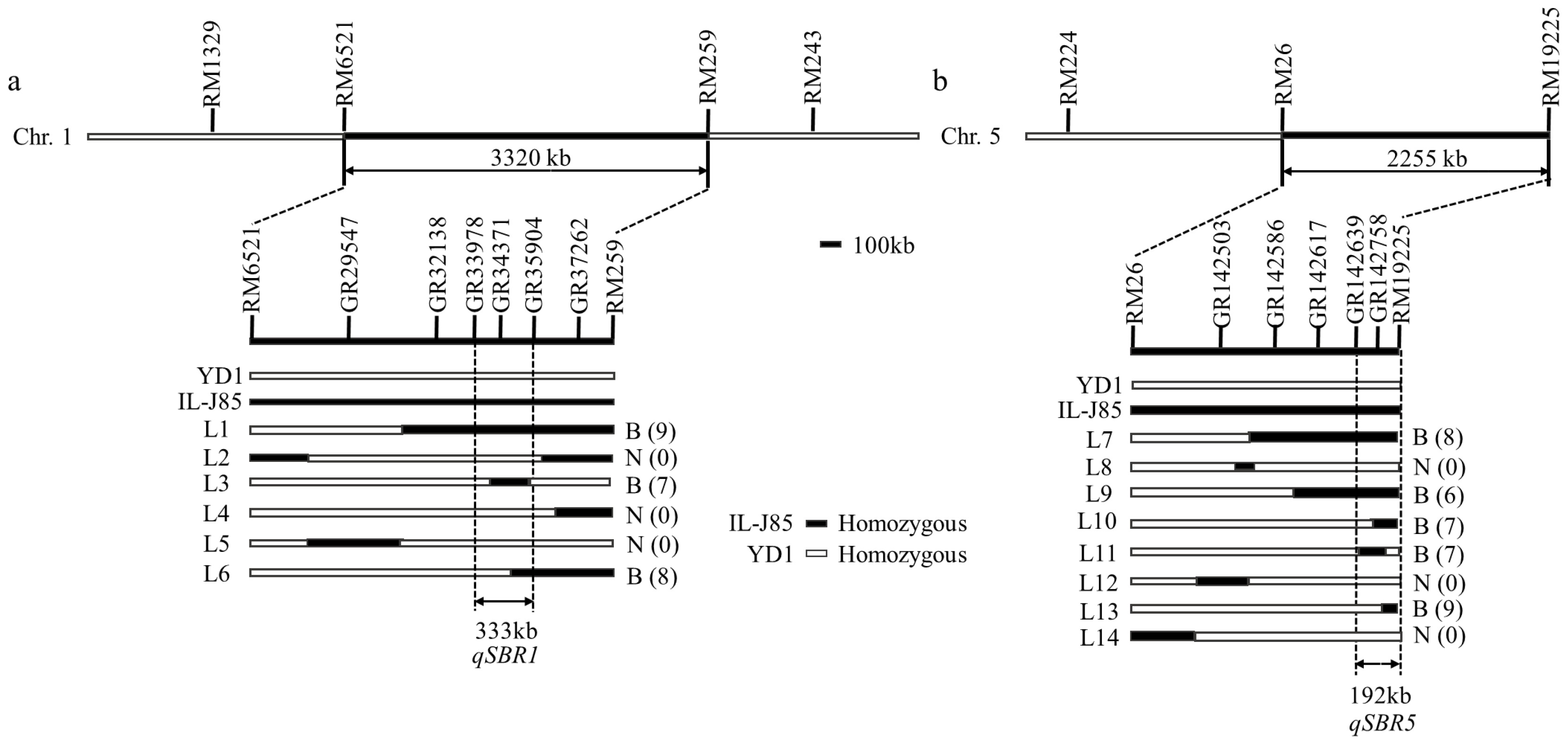
A two-way ANOVA was performed with the markers showing the most effect from these two regions (RM259 and RM26) (Figure 3). The results showed that the effect of these two regions alone and their interaction on the stem branch trait were significant. A QTL analysis showed that two QTLs on chromosomes 1 (in the interval between RM6521 and RM259) and 5 (in the interval between RM26 and RM19225) explained 15.65% and 13.45%, respectively (Table 4 and Table 5). We designated the locus on chromosome 1 as qSBR1 (Stem branch of Rice 1) and the locus on chromosome 5 as qSBR5 (Stem branch of Rice 5).
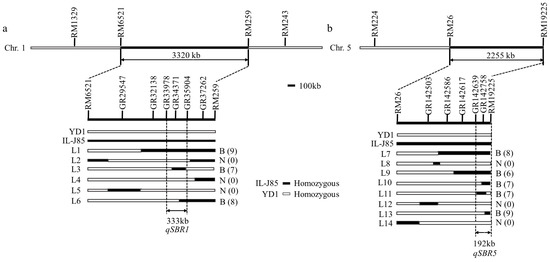
Figure 3.
Fine mapping of the qSBR1 and qSBR5 locus. (a) qSBR1 was initially mapped to the interval between the markers RM6521 and RM259, and then narrowed to a 333 kb region between the markers GR33978 and GR35904. (b) qSBR5 was initially mapped to the interval between the markers RM26 and RM19225, and then narrowed to a 192 kb region between the markers GR142639 and RM19225. B: stem branch; N: no-stem branch; the number of individuals is in parentheses; L1–L6: individuals that exchanged on chromosome 1; L7–L14: individuals that exchanged on chromosome 5.
Table 4.
Two-way ANOVA using one marker locus from each of the two putative regions related to stem branch trait.
Table 5.
QTLs controlling stem branch in rice.
In order to further fine map the two QTLs, we selected six individuals with heterozygous QTLs segments from the BC3F2 population to obtain the BC3F3 populations. Through the genotype analysis of 1461 individual plants, 50 recombinations of distinct stem branch individuals similar to IL-J85, and 50 non-stem branch individuals similar to YD1 were selected from the BC3F3 segregation population. The targeted sequencing genotype detection technology of the 10k SNP chip was used to analyze the genotype of these recombinant individuals. Through parental polymorphism analysis, a total of 67 and 132 polymorphic SNP markers were screened in the qSBR1 and qSBR5 intervals. The total length of qSBR1 interval was 18 cm with an average of 0.28 cm, and qSBR5 interval was 18.39 cm with an average of 0.14 cm. Compared with the phenotype and genotype of recombinants, we narrowed the qSBR1 and qSBR5 to chromosome 1 (18.64–19.94 cm, 333 kb) and chromosome 5 (117.54–118.30 cm, 192 kb) (Figure 3).
3.4. Effects of Different Genotype Combinations of Two QTLs on Stem Branch and Related Trait
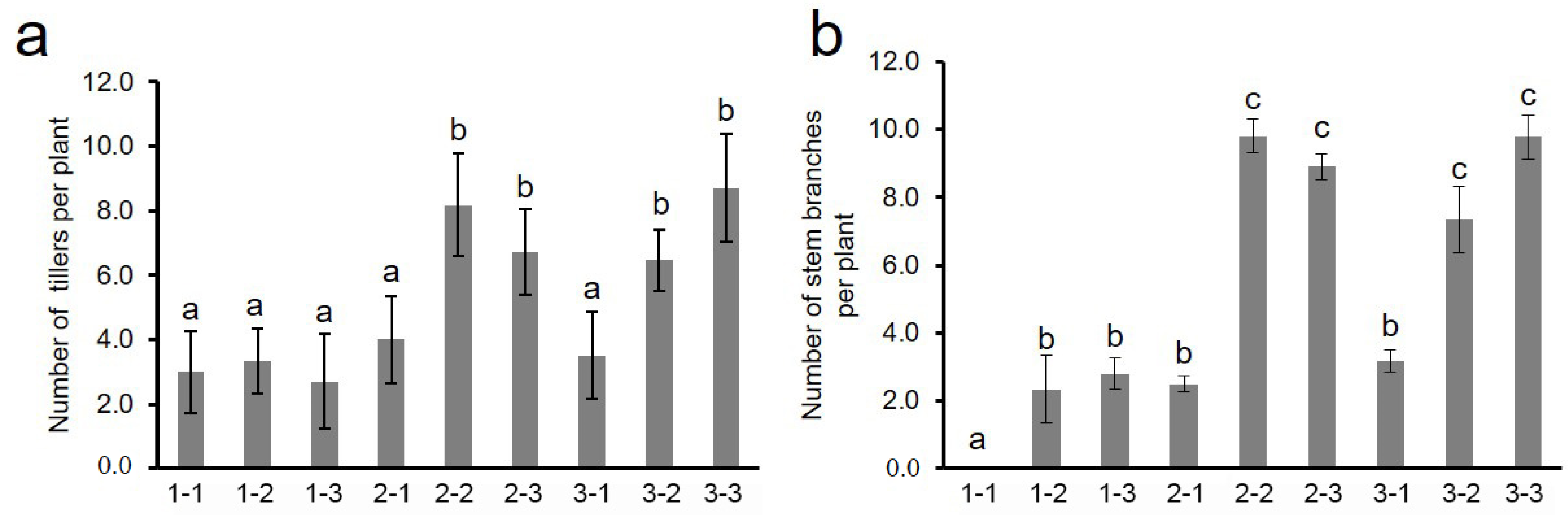
The investigation of the BC3F3 population found the basal regenerative tiller and stem branch trait 15 days later after harvest. We further analyzed the effects of different genotype combinations of two QTLs on the tiller and stem branch trait by QTL closely linked markers (qSBR1 Marker ID: GR34371; qSBR5 Marker ID: GR142758). The results showed that there was no significant difference in the number of regenerative tillers when two QTLs showed homozygous YD1 and one QTL showed heterozygous or homozygous introgression. However, with the combined introgression of two QTLs, whether heterozygous or homozygous, the number of regenerative tillers was significantly more than the YD1 genotype (Figure 4a). The same situation appeared on stem branch trait (Figure 4b). When the two QTLs were YD1 genotype, the stem branch trait did not occur, and one QTL introgression significantly increased the number of stem branches, and the simultaneous introgression of two QTLs significantly increased the number of stem branches compared with the YD1 genotype (Figure 4b). Interestingly, there was no significant difference between heterozygous and homozygous genotypes introgression of two QTLs. These results indicate that two QTLs simultaneously affected the growth of regenerated tiller and stem branch.
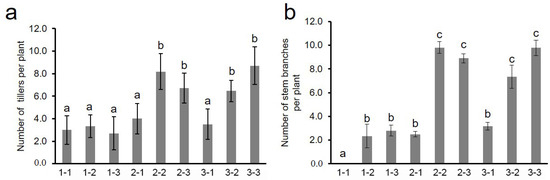
Figure 4.
Effects of different genotype combinations of two QTLs on branch trait. (a) Number of tillers per plant. (b) Number of stem branches per plant. 1, 2, 3 indicated homozygotes of YD1, and homozygotes and heterozygotes of IL-J85, respectively; the number in front and behind represent qSBR1 and qSBR5, respectively. The different letters above each bar indicated statistically significant differences as determined by one-way ANOVA followed by Tukey’s multiple comparison test (p < 0.05).
3.5. Breeding Utilization of Stem Branch Trait
In order to evaluate the breeding value of the stem branch trait, near-isogenic lines (NILs; NIL-Y37, NIL-D1) were cultivated using two japonica rice varieties Yunjing 37 (YJ37) and Dianjingyou 1 (DJY1) to cross and backcross IL-J85, respectively, through molecular markers assisted selection.
We further investigated the growth period of cuttings of ILs/NILs from transplanting to heading and found that the heading date (HD) of cuttings were shorter than that of seed seedlings (Table 6 and Figure 5). Other agronomic traits were investigated in the mature period. S-J85 was higher than C-J85 in plant height (PH), but the difference was not significant. However, S-Y37 was significantly higher than C-Y37 and S-D1 was significantly higher than C-D1; there was no significant difference of the primary panicle length (PL) between S-J85 and C-J85, but PL of S-Y37 was significantly shorter than that of C-Y37. Similarly, PL of S-D1 was shorter than that of C-D1; the flag leaf length (FL) of S-Y37 was significantly higher than that of C-Y37, while there was no significant difference between S-J85 and C-J85, and S-D1 and C-D1, respectively; in addition, there was no significant difference between S-J85 and C-J85, S-Y37 and C-Y37, and S-D1 and C-D1 for their flag leaf width (FW) and number of panicles (NP); both the primary branches per panicle (PP) and the secondary branches per panicle (SP) of S-Y37 were significantly higher than that of C-Y37; however, there was no significant difference between S-J85 and C-J85, S-D1 and C-D1, respectively; the 1000-grain weight of S-D1 was significantly higher than that of C-D1. Likewise, there was no significant difference between S-J85 and C-J85, and S-Y37 and C-Y37, respectively; the grain yield per plant (GY) of S-Y37 was significantly higher than that of C-Y37, but there was no significant difference between S-J85 and C-J85, and S-D1 and C-D1, respectively. These results showed that the cuttings can reduce the heading date compared with the seed seedlings in different backgrounds. Interestingly, there was no significant difference between the cuttings and the seed seedlings of S-J85 for their 1000-grain weight and GY. However, there was a partial difference between S-Y37 and C-Y37, and S-D1 and C-D1, in different backgrounds. These results suggest that the stem branch character can be used as vegetative reproduction in rice breeding utilization.
Table 6.
Investigating the agronomic traits of seed seedlings and cuttings from ILs/NILs in different backgrounds.

Figure 5.
Investigating plant architecture of seed seedling and cutting of ILs/NILs at the seedling and mature period. (a) S-Y37 and S-D1 are seed seedlings, C-Y37 and C-D1 are cuttings from the stem branches (bar = 5 cm). (b–d) S-J85 and C-J85, S-Y37 and C-Y37, S-D1 and C-D1 are at mature period; (bar = 10 cm).
4. Discussion
Rice is the staple food for half of the population worldwide. Breeders have placed efforts into improving output per unit area to fulfill the increasing demand of food. Double-cropping rice can be developed in tropical areas to increase rice production. Seed is the main expense of the cost of rice production, especially for hybrid rice. South China and the middle and lower reaches of the Yangtze River are the main areas for rice production. In most rice plant regions, the whole growth duration for a single cropping rice season was surplus but insufficient for double cropping rice. IL-J85 with the stem branch trait discovered in this research can be transplanted to cuttings after harvest and performed with a similar yield compared with YD1, which saved the growth time and production cost. A correlation analysis found that grain yield per plant was negatively correlated with the number of panicles; however, the number of stem branches was significantly correlated with the number of panicles. Furthermore, the number of stem branches affected plant agronomic traits including plant height, flag leaf length, flag leaf width, the number of panicles, and primary panicle length, indicating that there may be one pleiotropic gene or linkage inheritance. A genotypic analysis showed that the phenotype of the stem branch in the heterozygous state was consistent with that in the homozygous state, indicating that it can be broadly used in hybrid rice production. These results suggest that common wild rice has practical value for rice production by using the stem branch trait to fix heterosis in breeding rice varieties of vegetative reproduction.
The novel beneficial alleles and potential genetic diversity have been lost during wild rice domestication in cultivated rice. The wild rice species can better adapt to different environments and various abiotic and biotic stresses owing to the reservoir of novel genes/QTLs. Some agronomically important genes/QTLs for improving abiotic and biotic stresses, resistance, productivity, and grain quality characteristics were identified from AA genome donor wild species and labelled with breeder-friendly molecular markers to elite genetic backgrounds [47]. O. rufipogon (2n = 24, AA) was considered as the progenitor of O. sativa. There were several reports of using chromosomal segment substitution lines (CSSLs), backcross inbred lines (BILs), ILs, and NILs to introgress QTLs related to yield and grain quality from different O. rufipogon accessions into elite indica and japonica genetic backgrounds [48,49,50,51,52]. In this research, the introgression line IL-J85, constructed with YD1 and Yuanjiang common wild rice (O. rufipogon), had the stem branch trait and could be cut to form new individuals to be achieved a with similar yield of seed seedlings. The qSBR1 and qSBR5 responsible for the stem branch trait were first mapped on chromosomes 1 and 5. However, rice tiller is regulated by complex genes, hormones, and environmental factors. The molecular mechanism and regulatory network of the stem branch trait still need further elucidation.
Although there has been a doubling of major grain crop yields since the 1950s, people subjected to malnutrition prevail worldwide [53]. However, the crop yield increase cannot fulfill the demand for food with an ever-growing population [54]. Global food security depends on annual grains:cereals, oilseeds, and legumes, which damage essential ecosystem services, making some beyond sustainable boundaries [55,56]. The development of perennial varieties of important grain crops can expand choices of farmers to produce grains under less favorable circumstances, which can ensure food and ecosystem security [4]. Perennial crops are superior to annuals in sustaining important ecosystem functions, specifically on marginal landscapes or where resources are limited [57]. Perennial crops tend to have longer growing seasons and deeper rooting depths than annual counterparts. In addition, they intercept, retain, and utilize more precipitation [4,58,59]. Breeders are devoted to pursuing of high yield, high quality, high resistance, and other valuable characteristics of rice varieties. In this research, IL-J85 in YD1 background performed the stem branch trait and can be cut to form new individuals with similar yield of seed seedlings, which save rice seed cost, reduce seedling raising time and water usage. In addition, the cultivated near isogenic lines NIL-Y37 and NIL-D1 in the two backgrounds Yunjing 37 and Dianjingyou 1 perform the same characteristics as IL-J85, indicating the stem branch trait can be used in rice production and breeding utilization of rice varieties of vegetative reproduction.
5. Conclusions
This study found that IL-J85 can form new individuals by branches and adventitious roots on stem nodes, which can survive by cutting and bear the same yield per plant as IL-J85, thereby saving the growth time and production cost. The stem branch trait QTLs, namely qSBR1 and qSBR5, were first mapped on chromosomes 1 and 5 by mapping, MBS, and the targeted sequencing genotype detection technology of 10k SNP chip. The NIL-Y37 and NIL-D1 in the background YJ37 and DJY1 performed the same characteristics as IL-J85, which has the stem branch trait and could be cut to form new individuals. Our results provide new insights into the genetic mechanism of the stem branch characteristic in common wild rice, and have practical value using the stem branch trait to fix heterosis in breeding rice varieties of vegetative reproduction.
Author Contributions
P.X. and D.Y. conceived the project and designed the experiments. F.W. and P.X. wrote the manuscript and performed the experiments with assistance from C.L., J.W., J.Y., L.L., F.Z., Z.W., X.Z. and G.C. All authors have discussed the results and contributed to the drafting of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Science Foundation of China (Grant Nos. U1702231). The Science and technology projects of Yunnan Province, China (Grant Nos. 202003AD150007), Strategic Leading Science and Technology Program of Chinese Academy of Sciences (Grant Nos. XDA24030301), Natural Science Foundation of Yunnan, China (Grant Nos. 202101AT070062).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank public technology service center, crops protection and breeding base of Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences for technical and field support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sasaki, T.; Burr, B. International Rice Genome Sequencing Project: The effort to completely sequence the rice genome. Curr. Opin. Plant Biol. 2000, 3, 138–141. [Google Scholar] [CrossRef]
- Elert, E. Rice by the numbers: A good grain. Nature 2014, 514, S50–S51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Deng, Y.; Ding, Y.; Guo, J.; Qiu, J.; Wang, B.; Wang, C.; Xie, Y.; Zhang, Z.; Chen, J.; et al. Rice functional genomics: Decades’ efforts and roads ahead. Sci. China Life Sci. 2021, 65, 33–92. [Google Scholar] [CrossRef] [PubMed]
- Glover, J.D.; Reganold, J.P.; Bell, L.W.; Borevitz, J.; Brummer, E.C.; Buckler, E.S.; Cox, C.M.; Cox, T.S.; Crews, T.E.; Culman, S.W.; et al. Increased Food and Ecosystem Security via Perennial Grains. Science 2010, 328, 1638–1639. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.Y.; Tao, D.Y.; Sacks, E.; Fu, B.Y.; Xu, P.; Li, J.; Yang, Y.; McNally, K.; Khush, G.S.; Paterson, A.H.; et al. Convergent evolution of perenniality in rice and sorghum. Proc. Natl. Acad. Sci. USA 2003, 100, 4050–4054. [Google Scholar] [CrossRef] [Green Version]
- Tao, D.; Xu, P.; Hu, F.; Yang, Y.; Li, J.; Zhou, J. Hybrid sterility among near-isogenic lines derived from interspecific hybrid between cultivated rice species and Oraza glaberrima. Chin. J. Rice Sci. 2002, 16, 106–110. [Google Scholar]
- Hu, F.; Wang, D.; Zhao, X.; Zhang, T.; Sun, H.; Zhu, L.; Zhang, F.; Li, L.; Li, Q.; Tao, D.; et al. Identification of rhizome-specific genes by genome-wide differential expression Analysis in Oryza longistaminata. BMC Plant Biol. 2011, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Khush, G. S. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol. 1997, 35, 25–34. [Google Scholar] [CrossRef]
- Li, C.; Zhou, A.; Sang, T. Genetic analysis of rice domestication syndrome with the wild annual species, Oryza nivara. New Phytol. 2006, 170, 185–194. [Google Scholar] [CrossRef]
- Grillo, M.A.; Li, C.; Fowlkes, A.M.; Briggeman, T.M.; Zhou, A.; Schemske, D.W.; Sang, T. Genetic architecture for the adaptive origin of annual wild rice, Oryza nivara. Evol. Int. J. Org. Evol. 2009, 63, 870–883. [Google Scholar] [CrossRef]
- Chen, E.; Huang, X.; Tian, Z.; Wing, R.A.; Han, B. The Genomics of Oryza Species Provides Insights into Rice Domestication and Heterosis. Annu. Rev. Plant Biol. 2019, 70, 639–665. [Google Scholar] [CrossRef] [PubMed]
- Konishi, S.; Izawa, T.; Lin, S.Y.; Ebana, K.; Fukuta, Y.; Sasaki, T.; Yano, M. An SNP Caused Loss of Seed Shattering During Rice Domestication. Science 2006, 312, 1392–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Zhou, A.; Sang, T. Rice Domestication by Reducing Shattering. Science 2006, 311, 1936–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessho-Uehara, K.; Wang, D.R.; Furuta, T.; Minami, A.; Nagai, K.; Gamuyao, R.; Asano, K.; Angeles-Shim, R.B.; Shimizu, Y.; Ayano, M.; et al. Loss of function at RAE2, a previously unidentified EPFL, is required for awnlessness in cultivated Asian rice. Proc. Natl. Acad. Sci. USA 2016, 113, 8969–8974. [Google Scholar] [CrossRef] [Green Version]
- Hua, L.; Wang, D.R.; Tan, L.; Fu, Y.; Liu, F.; Xiao, L.; Zhu, Z.; Fu, Q.; Sun, X.; Gu, P.; et al. LABA1, a Domestication Gene Associated with Long, Barbed Awns in Wild Rice. Plant Cell 2015, 27, 1875–1888. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Liu, H.; Zhou, T.; Gu, B.; Huang, X.; Shangguan, Y.; Zhu, J.; Li, Y.; Zhao, Y.; Wang, Y.; et al. n-1 encodes a basic helix-loop-helix protein that regulates awn development, grain size, and grain number in rice. Plant Cell 2013, 25, 3360–3376. [Google Scholar] [CrossRef] [Green Version]
- Yano, K.; Yamamoto, E.; Aya, K.; Takeuchi, H.; Lo, P.-C.; Hu, L.; Yamasaki, M.; Yoshida, S.; Kitano, H.; Hirano, K.; et al. Genome-wide association study using whole-genome sequencing rapidly identifies new genes influencing agronomic traits in rice. Nat. Genet. 2016, 48, 927–934. [Google Scholar] [CrossRef]
- Zhu, B.-F.; Si, L.; Wang, Z.; Zhu, Y.Z.J.; Shangguan, Y.; Lu, D.; Fan, D.; Li, C.; Lin, H.; Qian, Q.; et al. Genetic Control of a Transition from Black to Straw-White Seed Hull in Rice Domestication. Plant Physiol. 2011, 155, 1301–1311. [Google Scholar] [CrossRef] [Green Version]
- Gross, B.L.; Steffen, F.T.; Olsen, K.M. The molecular basis of white pericarps in African domesticated rice: Novel mutations at the Rc gene. J. Evol. Biol. 2010, 23, 2747–2753. [Google Scholar] [CrossRef] [Green Version]
- Ishii, T.; Numaguchi, K.; Miura, K.; Yoshida, K.; Thanh, P.T.; Htun, T.M.; Yamasaki, M.; Komeda, N.; Matsumoto, T.; Terauchi, R.; et al. OsLG1 regulates a closed panicle trait in domesticated rice. Nat. Genet. 2013, 45, 462–465. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.; Zheng, X.; Wu, F.; Lin, Q.; Heng, Y.; Tian, P.; Cheng, Z.; Yu, X.; Zhou, K.; et al. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice. Nat. Plants 2017, 3, 17043. [Google Scholar] [CrossRef] [PubMed]
- Shomura, A.; Izawa, T.; Ebana, K.; Ebitani, T.; Kanegae, H.; Konishi, S.; Yano, M. Deletion in a gene associated with grain size increased yields during rice domestication. Nat. Genet. 2008, 40, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Li, X.; Liu, F.; Sun, X.; Li, C.; Zhu, Z.; Fu, Y.; Cai, H.; Wang, X.; Xie, D.; et al. Control of a key transition from prostrate to erect growth in rice domestication. Nat. Genet. 2008, 40, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Huang, W.; Gao, J.P.; Yang, J.; Shi, M.; Zhu, M.Z.; Luo, D.; Lin, H.X. Genetic control of rice plant architecture under domestication. Nat. Genet. 2008, 40, 1365–1369. [Google Scholar] [CrossRef]
- Sun, C.; Wang, X.; Yoshimura, A.; Doi, K. Genetic differentiation for nuclear, mitochondrial and chloroplast genomes in common wild rice (Oryza rufipogon Griff.) and cultivated rice (Oryza sativa L.). Theor. Appl. Genet. 2002, 104, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.-H.; Shang, F.; Lin, Q.-T.; Lou, C.; Zhang, J. Tillering and panicle branching genes in rice. Gene 2014, 537, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Wang, J.; Zhu, X.; Hao, W.; Wang, L.; Li, Q.; Zhang, L.; He, W.; Lu, B.; Lin, H.; et al. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat. Genet. 2008, 40, 1370–1374. [Google Scholar] [CrossRef]
- Li, X.; Qian, Q.; Fu, Z.; Wang, Y.; Xiong, G.; Zeng, D.; Wang, X.; Liu, X.; Teng, S.; Hiroshi, F.; et al. Control of tillering in rice. Nature 2003, 422, 618–621. [Google Scholar] [CrossRef]
- Lu, Z.; Shao, G.; Xiong, J.; Jiao, Y.; Wang, J.; Liu, G.; Meng, X.; Liang, Y.; Xiong, G.; Wang, Y.; et al. MONOCULM 3, an ortholog of WUSCHEL in rice, is required for tiller bud formation. J. Genet. Genom. 2015, 42, 71–78. [Google Scholar] [CrossRef]
- Tanaka, W.; Ohmori, Y.; Ushijima, T.; Matsusaka, H.; Matsushita, T.; Kumamaru, T.; Kawano, S.; Hirano, H.-Y. Axillary Meristem Formation in Rice Requires the WUSCHEL Ortholog TILLERS ABSENT1. Plant Cell 2015, 27, 1173–1184. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Wang, Y.; Yu, Y.; Duan, J.; Liao, Z.; Xiong, G.; Meng, X.; Liu, G.; Qian, Q.; Li, J. Degradation of MONOCULM 1 by APC/CTAD1 regulates rice tillering. Nat. Commun. 2012, 3, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; Wang, D.; Dong, H.; Gu, S.; Cheng, Z.; Gong, J.; Qin, R.; Jiang, L.; Li, G.; Wang, J.L.; et al. Rice APC/C (TE) controls tillering by mediating the degradation of MONOCULM 1. Nat. Commun. 2012, 3, 752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsua, M.; Maekawab, M.; Shimamotoa, K.; Kyozuka, J. The LAX1 and FRIZZY PANICLE 2 Genes Determine the Inflorescence Architecture of Rice by Controlling Rachis-Branch and Spikelet Development. Dev. Biol. 2001, 231, 364–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabuchi, H.; Zhang, Y.; Hattori, S.; Omae, M.; Shimizu-Sato, S.; Oikawa, T.; Qian, Q.; Nishimura, M.; Kitano, H.; Xie, H.; et al. LAX PANICLE2 of Rice Encodes a Novel Nuclear Protein and Regulates the Formation of Axillary Meristems. Plant Cell 2011, 23, 3276–3287. [Google Scholar] [CrossRef] [Green Version]
- Alder, A.; Jamil, M.; Marzorati, M.; Bruno, M.; Vermathen, M.; Bigler, P.; Ghisla, S.; Bouwmeester, H.; Beyer, P.; Al-Babili, S. The path from β-carotene to carlactone, a strigolactone-like plant hormone. Science 2012, 335, 1348–1351. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.M.; Ai-Ping, Z.; Ping, L.I.; Wang, S.Q.; Liang, Y.Y.; Ling, L.I. Identification and cloning of tillering-related genes osmax1 in rice. Rice Sci. 2015, 22, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Xu, Y.; Liu, H.; Mao, Z.; Zhang, C.; Ma, Y.; Zhang, Q.; Meng, Z.; Chong, K. The interaction between OsMADS57 and OsTB1 modulates rice tillering via DWARF14. Nat. Commun. 2013, 4, 1566. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Liu, X.; Xiong, G.; Liu, H.; Chen, F.; Wang, L.; Meng, X.; Liu, G.; Yu, H.; Yuan, Y.; et al. DWARF 53 acts as a repressor of strigolactone signalling in rice. Nature 2013, 504, 401–405. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wu, K.; Qian, L.; Liu, Q.; Li, Q.; Pan, Y.; Ye, Y.; Liu, X.; Wang, J.; Zhang, J.; et al. Non-canonical regulation of SPL transcription factors by a human OTUB1-like deubiquitinase defines a new plant type rice associated with higher grain yield. Cell Res. 2017, 27, 1142–1156. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yu, H.; Xiong, G.; Lu, Z.; Jiao, Y.; Meng, X.; Liu, G.; Chen, X.; Wang, Y.; Li, J. Tissue-Specific Ubiquitination by IPA1 INTERACTING PROTEIN1 Modulates IPA1 Protein Levels to Regulate Plant Architecture in Rice. Plant Cell 2017, 29, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.; Li, J.; Xie, D. Recent advances in molecular basis for strigolactone action. Sci. China Life Sci. 2017, 61, 277–284. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Xue, D.; Wang, J.; Yan, M.; Liu, G.; Dong, G.; Zeng, D.; Lu, Z.; Zhu, X.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef]
- Lu, Z.; Yu, H.; Xiong, G.; Wang, J.; Jiao, Y.; Liu, G.; Jing, Y.; Meng, X.; Hu, X.; Qian, Q.; et al. Genome-Wide Binding Analysis of the Transcription Activator Ideal Plant Architecture1 Reveals a Complex Network Regulating Rice Plant Architecture. Plant Cell 2013, 25, 3743–3759. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.; Jin, Y.; Liu, W.; Li, F.; Fang, J.; Yin, Y.; Qian, Q.; Zhu, L.; Chu, C. DWARF AND LOW-TILLERING, a new member of the GRAS family, plays positive roles in brassinosteroid signaling in rice. Plant J. 2009, 58, 803–816. [Google Scholar] [CrossRef]
- Liu, X.; Hu, Q.; Yan, J.; Sun, K.; Liang, Y.; Jia, M.; Meng, X.; Fang, S.; Wang, Y.; Jing, Y.; et al. ζ-Carotene Isomerase Suppresses Tillering in Rice through the Coordinated Biosynthesis of Strigolactone and Abscisic Acid. Mol. Plant 2020, 13, 1784–1801. [Google Scholar] [CrossRef]
- Abdel-Latif, A.; Osman, G. Comparison of three genomic DNA extraction methods to obtain high DNA quality from maize. Plant Methods 2017, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Gaikwad, K.B.; Singh, N.; Kaur, P.; Rani, S.; Babu, H.P.; Singh, K. Deployment of wild relatives for genetic improvement in rice (Oryza sativa L.). Plant Breed 2021, 140, 23–52. [Google Scholar] [CrossRef]
- Jin, F.X.; Kim, D.M.; Ju, H.G.; Ahn, S.N. Mapping quantitative traits loci for awnness and yield component traits in isogenic lines derived from an Oryza sativa/O. rufipogon cross. J. Crop Sci. Biotechnol. 2009, 12, 9–15. [Google Scholar] [CrossRef]
- Yuan, P.-R.; Kim, H.-J.; Chen, Q.-H.; Ju, H.-G.; Ji, S.-D.; Ahn, S.-N. Mapping QTLs for grain quality using an introgression line population from a cross between Oryza sativa and O. rufipogon. J. Crop Sci. Biotechnol. 2010, 13, 205–212. [Google Scholar] [CrossRef]
- Luo, X.; Wu, S.; Tian, F.; Xin, X.; Zha, X.; Dong, X.; Fu, Y.; Wang, X.; Yang, J.; Sun, C. Identification of heterotic loci associated with yield-related traits in Chinese common wild rice (Oryza rufipogon Griff.). Plant Sci. Int. J. Exp. Plant Biol. 2011, 181, 14–22. [Google Scholar] [CrossRef]
- Liang, Y.; Yan, C.; Zheng, J.; Nan, W.; Qin, X.; Zhang, H. Locating QTL associated with spike traits of Dongxiang wild rice (Oryza rufipogon Griff.). Euphytica 2019, 215, 26. [Google Scholar] [CrossRef]
- Guttikonda, H.; Thummala, S.R.; Agarwal, S.; Mangrauthia, S.K.; Ramanan, R.; Neelamraju, S. Genome-wide transcriptome profile of rice hybrids with and without Oryza rufipogon introgression reveals candidate genes for yield. Sci. Rep. 2020, 10, 4873. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations (FAO). The State of Food Insecurity in the World 2009; Progress Report; FAO: Rome, Italy, 2009. [Google Scholar]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Monfreda, C.; Ramankutty, N.; Foley, J.A. Farming the planet: 2. Geographic distribution of crop areas, yields, physiological types, and net primary production in the year 2000. Glob. Biogeochem. Cycles 2008, GB1022. [Google Scholar] [CrossRef]
- Pimm, S. L. The World According to Pimm; McGraw-Hill: New York, NY, USA, 2001. [Google Scholar]
- Tilman, D.; Socolow, R.; Foley, J.A.; Hill, J.; Larson, E.; Lynd, L.; Pacala, S.; Reilly, J.; Searchinger, T.; Somerville, C.; et al. Beneficial Biofuels—The Food, Energy, and Environment Trilemma. Science 2009, 325, 270–271. [Google Scholar] [CrossRef] [Green Version]
- Randall, G.W.; Huggins, D.R.; Russelle, M.P.; Fuchs, D.J.; Nelson, W.W.; Anderson, J.L. Nitrate Losses through Subsurface Tile Drainage in Conservation Reserve Program, Alfalfa, and Row Crop Systems. J. Environ. Qual. 1997, 26, 1240–1247. [Google Scholar] [CrossRef]
- Dohleman, F.G.; Long, S.P. More Productive Than Maize in the Midwest: How Does Miscanthus Do It? Plant Physiol. 2009, 150, 2104–2115. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).