Abstract
Combining ability along with heterosis was elucidated in the 21 F1 hybrid population of rice derived from a 7 × 7 diallel mating design. Furthermore, to formulate a strategy for marker-based prediction of heterosis, the molecular genetic diversity (GD) was then surveyed among the seven parental lines with RAPD markers. Analysis of combining ability revealed significant GCA and SCA variances and the ratio between the aforesaid genetic parameter was more than unity for most of the traits signifying the preponderance of additive gene action. Hybrid combination Double Rice × Pokkali was the best specific combiner for the traits pollen fertility percentage (%), panicle length (cm), filled grains/panicle (no.) and grain yield/hill (g) while Muktagacha × BRRI dhan29 and Basmati × Double Rice were the best specific combiners for grain length (mm) and 1000-grain weight (g), respectively. Regarding the magnitude of heterosis, Double Rice × Pokkali exhibited maximum heterobeltiosis for panicle length (33.33%) and the number of filled grains/panicle (144.01%). Furthermore, Pokkali × Chinigura was verified for producing the highest heterobeltiosis for tillers/hill (173.17%), as well as grain yield/hill (71.05%). The heatmap relying on molecular genetic diversity exposed the highest genetic distance between Chinigura and Pokkali followed by Muktagacha and Pokkali. Unweighted neighbor-joining trees clustered the parental lines into three groups indicating the presence of considerable genetic diversity among those seven parental genotypes. A positive and significant correlation was explored between molecular GD with specific combining ability as well as heterosis for all of the traits revealing the huge potentiality of hybrid performance prediction using RAPD markers with the present set of individuals. Thus, exploration of definite genomic regions underlying the candidate genes for respective traits relying on such RAPD-generated data following an advanced molecular marker approach would pave the path of mining out yielder heterotic combinations.
1. Introduction
For the majority of Asians, rice is a way of life. Rice has influenced the cultures, cuisines, and economics of millions of people worldwide [1]. It is a self-pollinated cereal crop having chromosome number 2n = 24 [2]. Rice is a member of the genus Oryza under the Poaceae family. This monocot genus comprises 27 species, 25 of which are wild type, and only two species are cultivated viz., Oryza sativa and Oryza glaberrima. Oryza sativa includes all variants found in Asia, America, and Europe, while Oryza glaberrima covers all types found in West Africa. Rice is a nutrient-dense cereal. It offers 20% of the calories and 15% of the protein that the world’s population consumes [3]. Rice is considered a staple food in the Bangladeshi diet as it is consumed by 164 million people in the country. It is grown on a large scale by the farmers of the country to meet the expected demand of consumption as well as the exportation of rice production accounting for half of Bangladesh’s agricultural gross domestic product (GDP) and one-sixth of the country’s national income [4]. Being the staple food of the population, its demand is increasing day by day. By 2050, the world’s rice demand will reach 943.6 million tons, requiring a 5.8-million-ton annual increase from current levels of production [5]. However, rice production is not increasing to keep pace with this gradual demand due to different constraints. Due to the construction of industries, factories, residences, roads, and other structures, total cultivable land is declining at a pace of more than 1% each year. Furthermore, agriculture is subjected to a variety of adverse conditions as a result of climate change, including drought, flood, salinity, and high temperatures. So, the demand for increasing rice production in sustainable ways remains an enormous challenge. Nowadays, quality parameters are becoming increasingly important in today’s world, particularly in areas where rice production is self-sufficient. People’s demand for high-quality rice is also increasing steadily as their per capita income and living standards improve. Therefore, in recent times, grain quality improvement should consider an important objective next to yield in rice breeding. In this circumstance, one of the main possibilities for meeting the increased demand is to develop and disseminate high-yielding and high-quality rice varieties. That’s why the current study focuses on the production of high-yielding and high-quality rice varieties through the selection of proper parents and identifying superior cross combinations. The parental lines are considered to be the backbone of rice breeding since the performance of the progenies mainly depends on the genetic potential of the parents [6]. In the case of a hybrid breeding program, the selection of parents based on phenotypic performance alone is not a sound proposition since phenotypically superior lines may yield poor recombination in the segregating generations. To overcome this challenge, combining ability is used in hybrid plant breeding programs to select parents and superior hybrid combinations [7,8]. Combining ability analysis is a useful approach for measuring a parent’s potential in order to create a superior hybrid. Combining ability can be used to assess nicking ability between genotypes while also explaining the nature and extent of gene effects [9]. Sprague and Tatum [8] were the first to define the concept of combining ability, which was later clarified by Griffing [10]. Among two types of combining ability, general combining ability (GCA) denotes a parental line’s average performance in hybrid combinations and reflects additive gene action, whereas Specific combining ability (SCA) indicates non-additive gene action and cross combinations with high SCA will be exploited to produce hybrid varieties [11]. Heterosis or hybrid vigor is another important factor related to hybrid performance which refers to the superior performance of hybrids over the parents. For decades, the use of heterosis has made a significant contribution to feeding an ever-increasing human population on depleting arable land [12]. Heterosis may be positive or negative where positive heterosis is desirable for grain yield and negative heterosis is preferred for earliness and plant height. With the advancement of biometrical genetics, numerous strategies for estimating combining ability and heterosis have been proposed. These include top cross, poly cross, diallel cross analysis by Griffing [10], and line × tester analysis by Kempthorne [13]. In this experiment, diallel mating design is followed to estimate combining ability and heterosis. The parental lines are crossed in all feasible combinations in diallel mating to identify parents as the best or poor general combiners by GCA and specific cross combinations by SCA. For many species, morphological descriptions are not sufficient for the detection of differences among themselves. Since the morphological characters are influenced by the environment, they often do not reliably portray genetic diversity. Compared with morphological variation, molecular polymorphism is generally considered more informative. Thus, screening inbred lines for genetic diversity using molecular markers could enhance the accuracy of parent selection for breeding operations [14]. Molecular techniques combined with biometrical methodologies have opened up new avenues for assessing input materials’ appropriateness as parents [15]. Since the 1990s, several pieces of research have been conducted to predict progeny performance based on genetic distance utilizing the basic theory of quantitative genetics. The utilization of molecular markers to forecast heterosis has ushered in a new era since its introduction in 1980 [16]. Much research on various crop plants, such as rice [16,17,18], wheat [19], and maize [20,21], has been undertaken to anticipate the link between genetic diversity and heterosis. Molecular markers have been proposed as a more efficient technique for selecting inbreds and superior hybrid combinations, as well as a way to reduce the number of multi-location trials of promising hybrids [22,23].
2. Materials and Methods
Seven rice (Oryza sativa L.) varieties/landraces were selected from the rice germplasm pool of the Department of Genetics and Plant Breeding, Bangabandhu Sheikh Mujibur Rahman Agricultural University. The collection of germplasm was being conducted around April–May of 2019. Detailed information on the collected germplasm for this research is given in the following Supplementary Table S1. Flowering in all the parents took place more or less at the same time. The seven diverse genotypes were crossed in a 7 × 7 half-diallel fashion, resulting in 21 single crosses. Healthy and representative plants of the variety were removed from the experimental plot by cutting the mud around the base using a sickle. The plants were then placed in pots by adding soil and water as necessary and tagged with proper designation and brought to the laboratory for emasculation. In the afternoon, emasculation was performed by trimming the top 1/3rd and bottom 1/3rd portions of the targeted female parent’s panicle using scissors. Again, the top 1/3rd portion in each spikelet was trimmed off in a slanting position. The six anthers present in each spikelet were removed with the help of the Vacuum Emasculator. The emasculated spikelets were then wrapped with a butter paper bag to prevent contamination from external pollen. The blooming panicle from the intended male parent was taken the following morning (typically around 9.30 a.m.). Then, the butter paper bag was removed from the emasculated panicles of the female parent and pollen was dusted from the desired male parent. After confirming that plenty of pollen was distributed, the opened butter paper bag was sealed with a pin. In the second Aman season, both parental and F1 hybrid seeds were again set for germination. On 13 July 2020, seeds were sown in the pot after completing germination. Then, on 13 August 2020 thirty-one days, old seedlings were transplanted in the main field in a Randomized Complete Block Design (RCBD) with two replications. At this time, field preparation, application of fertilizers and manures, planting spacing, and application of different intercultural operations were the same as the previous season. Both parents and F1 hybrids were allowed self-pollination and left in the field for attaining maturity. After becoming fully matured, seeds were harvested and then dried in sun and made ready for data collection. Data were recorded on 10 randomly selected plants from parents and F1s plant samples. In this study, ten yield-contributing agronomic traits were estimated which included pollen fertility (%), days to first flowering, plant height (cm), tillers/hill, panicle length (cm), filled grains/panicle, grain length (mm), 1000-grain weight (g), grain test weight (g/5 mL), grain yield/hill (g) were recorded.
2.1. Analysis Combining Ability and Heterosis
Griffing’s [10] method 2, model 1 (fixed-effect model) was used for combining ability analysis for each of the traits. In model 1, the experimental material is regarded as the population about which inferences are to be made, the general objectives are to compare the combining abilities of the parents and, using parents as a tester, to identify better cross combination(s) [24]. The amount of heterosis was calculated by comparing the mean of F1 hybrids over mid-parental value and higher parent in respect of a particular character using the following formulae [25]. The percent of heterosis over mid parent (MP) = [(F1 − MP)/MP] × 100, percent of heterosis over higher parent (HP) = [(F1 − HP)/HP] × 100 and percent of heterosis over super parent (SP) = [(F1 − SP)/SP] × 100. The test of significance for heterosis was conducted by usual statistical practice using the error variance from the ANOVA of F1 and parental populations [24].
2.2. DNA Extraction and RAPD Marker
DNA has been extracted following the modified Cetyl Trimethyl Ammonium Bromide (CTAB) method [26]. After several successive centrifugations at projected rpm DNA pellets formed at the bottom of the tube were separated from the supernatants. The pellets were then rinsed with 70% ethanol and centrifuged for 5 min. at 12,000 rpm. Finally, the pellets were suspended in 150 μL 1X TE buffer and stored at −20 °C. A spectrophotometer was used to quantify the extracted DNA, while the DNAs were run on 1% agarose gel to determine their quality. Seven RAPD primers with clear amplifications were selected for genetic diversity analysis of seven parents of rice. Detailed information about the RAPD primers used in this study is presented in Supplementary Table S2.
2.3. PCR Mix Preparation and Amplification
PCR was performed in 14 μL reactions containing 7.5 μL master mixtures, 3.5 μL distilled water, 1 μL primer, and 2 μL of DNA sample (Supplementary Table S3). The DNA was transferred from the dilution plate to the PCR plate using a single-channel pipette. After initial denaturation for 1 min at 94 °C, each cycle comprises 30 s denaturation at 94 °C, 30 s annealing at 40 °C, and 1 min extension at 72 °C with a final extension for 5 min at 72 °C at the end of 38 cycles. The PCR product was preserved at 12 °C temperature in the thermal cycler in case of necessity.
2.4. Agarose Gel Electrophoresis and Visualization
The gel was prepared by dissolving the 1.2 g agarose powder in 100 mL TAE buffer. Then, 0.5 μL Ethidium Bromide was added for visualization. The agarose was distributed in the buffer before being heated to near boiling but not boiling temperatures. The melted agarose was allowed to cool completely before being poured into a cast, as a heated agarose solution could cause the cast to bend or crack. A comb was placed in the cast to create wells for loading the sample, and the gel should be completely set before use. After the gel was polymerized, the gasket was removed starting from one corner of the plate assembly. Around 500 mL of 1× TAE buffer was added to the base of the tank. Around 300 mL of 1× TAE buffer was again added on top of the tank and the comb was removed gently. Around 5 μL of the mixer was loaded in the wells of gel with the help of a pipette. Then the cover of the tank was put and the electrodes were connected to the power supply and the gel was run for about 30 min at 110 volts. It was noted that running time was dependent on the size of PCR fragments. The power supply unit was turned off and the plates were removed from the tank. The agarose gel was removed carefully and was put in the gel doc system. Then image analysis was performed on the connected computer. Here, the Ethidium Bromide protocol was run for image analysis. The amplified DNA band fragments of the RAPD assay were scored in binary characters, where 1 and 0 indicate the presence and absence of each band, respectively.
2.5. Analysis of Marker Data
Analysis of molecular variance (AMOVA) was used to partition the variation among and within-group (population) components. AMOVA was constructed by using the software GenAlEx6.5 [27]. Matrices of Roger’s genetic distance [28] was also calculated between each pair of line in the study using the same software. With the help of DARwin software (version 6.0, Montpellier CEDEX 5, France) a dendrogram was generated from the genetic distance matrix using the unweighted neighbor-joining algorithm [29]. The correlation of RAPD-generated genetic distance with the specific combining ability (SCA), mid-parent heterosis (MPH), better-parent heterosis (BPH), and standard-parent heterosis (SPH) was calculated by exploiting the ‘Data Analysis’ wing of the Microsoft Excel 2019 software and the significance test was performed with the same software assuming two samples with equal variance. A heatmap was generated with the help of a freely available web server named Heatmapper [30].
3. Results
3.1. Analysis of Variance (ANOVA)
The preliminary analysis of variance showed that the mean sum of squares for the parents and F1 hybrids were significant at p ≤ 0.01 for all the traits (Supplementary Table S4). These significant variances suggested that there was a wide diversity in the genotypes for the studied traits. Accordingly, analysis of variances for combining ability (Figure 1 and revealed that variances for both GCA and SCA variances were also significant for the studied traits. Moreover, the mean square values for the GCA were higher than the SCA for all the traits, signifying the preponderance of additive gene action for controlling the traits.
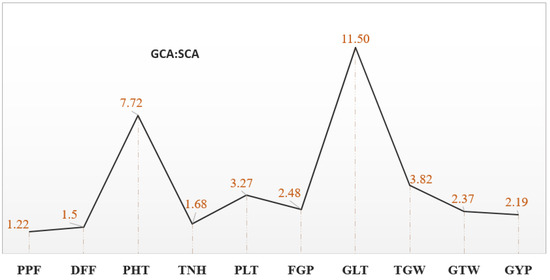
Figure 1.
General Combining Ability (GCA) and Specific Combining Ability (SCA) ratio for ten yield and yield contributing traits in a 7 × 7 diallel population of rice. PPF = Percent Pollen fertility, DFF = Days to first flowering, PHT = Plant height (cm), TNH = Tillers/hill, PLT = Panicle length (cm), FGP = Filled grains/panicle, GLT= Grain length (mm), TGW = 1000-grain weight (g), GTW = Grain test weight (g/5 mL), GYP = Grain yield/plant (g).
3.2. Mean Performance of the Parents and Hybrids
The parent Chinigura (96.16%) exhibited the best performance for the trait pollen fertility (%), while the lowest performance was revealed by the parent Double Rice (61.56%) (Figure 2 and Supplementary Table S5). Consequently, in hybrid genotypes, pollen fertility ranged from 59.19% to 96.83%. The maximum value was observed in the cross-combination Basmati × Double Rice (96.83%). The parental genotype Pokkali took 79 days for the first flowering while the line Double Rice took the maximum time (98.5 days). Interestingly, in the case of hybrids, days to first flowering varied from 67 to 95 days where the minimum time was required for the cross combination Pokkali × BRRI dhan29 and maximum time was taken by cross named Bashmoti × Chinigura (95 days).
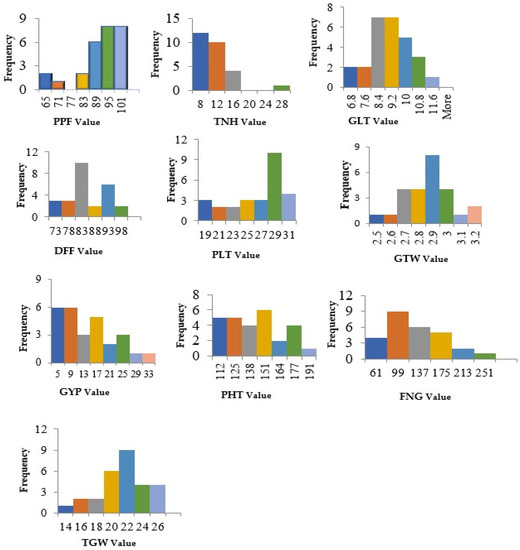
Figure 2.
Frequency distribution graph illustrating overall mean performance of the 7 × 7 diallel population of rice. PPF = Percent Pollen fertility, DFF = Days to first flowering, PHT = Plant height (cm), TNH = Tillers/hill, PLT = Panicle length (cm), FGN = Filled grain numbers/panicle, GLT = Grain length (mm), TGW = 1000-grain weight (g), GTW = Grain test weight (g/5 mL), GYP = Grain yield/plant (g).
In parents, panicle length ranged from 20.63 to 29.24 cm, however, the highest panicle length was observed as 30.95 cm in (Bashmoti × Muktagacha) considering the hybrid population. Filled grains/panicle ranged from 46.5 to 174.38 and 23 to 249.5, respectively, in parents and hybrid populations. Grain length, values ranged from 6.1 mm (Chinigura) to 10.96 mm (Muktagacha) in parent and the grain length of hybrids ranged from 6.55 (BRRI dhan29 × Chinigura) to 10.22 cm (Double Rice × Kotoktara). The hybrid Double Rice × Kotoktara (23.55 g) followed by Muktagacha × Kotoktara (23.05 g) was recorded as having the highest grain weight. Meanwhile, in the parental genotype, the highest value was found in Double Rice (25.42 g). Test weight is used as an indicator of general grain quality and is a measure of grain bulk density. It is also an important predictor for milling of yield and is used to compare grain densities. Farmers prefer genotypes of crops having high test weight. The cross-combination Basmati × Double Rice (3.22 g/5 mL) exhibited the highest value followed by Pokkali × Chinigura (3.14 g/5 mL). Finally, grain yield/plant ranged from 2.05 g to 18.95 g in parents and 1.34 g to 29.60 g in hybrid combinations where the parental genotype basmati (18.95 g) and the hybrid, Pokkali × Chinigura (29.6 g) were the best performers. Moreover, correlation coefficient estimation of yield components with the farmer desire trait, i.e., grain yield per plant relying on mean performance exposed that plant height, panicle length and filled grains per panicle were the most contributing traits toward the grain yield (Figure 3).
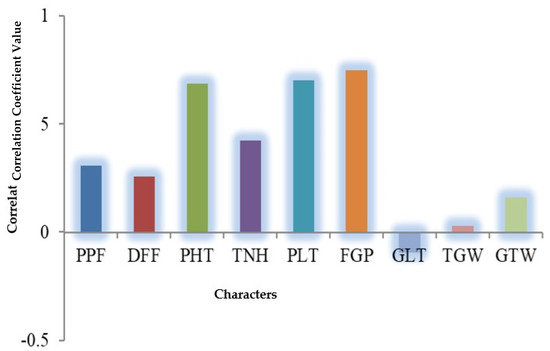
Figure 3.
Correlation coefficient values of different yield components with grain yield per plant. PPF = Percent Pollen fertility, DFF = Days to first flowering, PHT = Plant height (cm), TNH = Tillers/hill, PLT = Panicle length (cm), FGP = Filled grains/panicle, GLT = Grain length (mm), TGW = 1000-grain weight (g), GTW = Grain test weight (g/5 mL), GYP = Grain yield/plant (g).
3.3. Combining Ability Analysis (GCA and SCA)
Parental genotype BRRI dhan29 followed by Kotoktara (−15.71) showed significant results and the highest negative GCA effect for plant height signifying their potentiality for developing short stature hybrid variety(s) (Figure 4 and Supplementary Table S6). Parental line Muktagaccha was identified as a good combiner for exhibiting significant and the highest GCA effects for panicle length filled grains/panicle, and grain length. For days to first flowering, Pokkali was the best performer as it revealed the highest negative and significant GCA effects (−4.43) followed by BRRI dhan29. Double Rice was a good combiner for 1000-grain weight with a GCA effect of 2.62. In the case of test weight, Chinigura (0.16) exhibited the highest GCA effect. Chinigura could be considered as the best general combiner for grain yield/hill due to the highest positive (4.57) effects followed by Basmati (2.08).
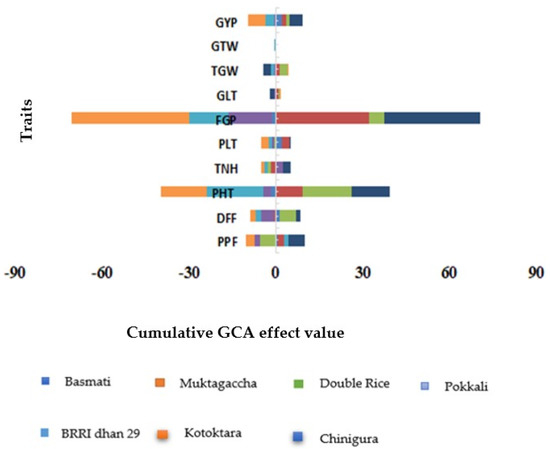
Figure 4.
General combining ability (GCA) effects for ten yield and yield related characters in a 7 × 7 diallel population of rice. PPF = Percent Pollen fertility, DFF = Days to first flowering, PHT = Plant height (cm), TNH = Tillers/hill, PLT = Panicle length (cm), FGP = Filled grains/panicle, GLT = Grain length (mm), TGW = 1000-grain weight (g), GTW = Grain test weight (g/5 mL), GYP = Grain yield/plant (g).
Rice hybrids that mature early are preferable because they are well fitted for multiple cropping. So, a negative SCA effect is desirable for days to flowering. The highest negative SCA effect (−15.44) was recorded for the cross Double Rice × BRRI dhan29 (Table 1) followed by Muktagacha × Chinigura (−10.67) which indicated that these combinations might be suitable for evolving short-duration rice genotypes. Shortening plant height is a desirable trait in rice crop cultivation as it enhances resistance to lodging and increases tillering ability (Saleem et al., 2008). So, the combination of Kotoktara × Chinigura (−34.79) showing a significant negative SCA effect was considered desirable regarding this trait.

Table 1.
Estimates of specific combining ability (SCA) effects for ten yield and its related characters in a 7 × 7 diallel population of rice.
The crosses Double Rice × Pokkali, BRRI dhan29 × Chinigura, and Muktagacha × Pokkali were the best specific combinations due to their highest positive SCA effect (Table 1). The cross combinations Muktagaccha × BRRI dhan29 (0.82), Kotoktara × Chinigura (0.79), and Pokkali × Chinigura (0.58) were promising for long-grain as they possessed significant and positive SCA effect. 1000-grain weights are an important trait influencing yield and its positive SCA effect is desirable. For 1000-grain weight, positive and significant SCA effects with greater value were 48 registered in crosses viz., Muktagaccha × Chinigura (6.12) followed by Double Rice × Chinigura (4.73) and Basmati × Pokkali (1.86). The hybrids BRRI dhan29 × Chinigura (10.44), Double Rice × Pokkali (13.91), and Pokkali × Chinigura were good specific combiners for grain yield because they expressed high SCA effects for grain yield.
3.4. Estimation of Heterosis
The degree and nature of heterosis were found to vary depending on cross combinations and the characters under study (Table 2). For all of the attributes, the majority of the cross combinations showed considerable positive heterosis. Double Rice × Pokkali exhibited maximum heterobeltiosis for panicle length (33.33%) and filled grains/panicle (144.01%). Significant heterosis for filled grains/panicle was observed in almost all the hybrids. The value ranged from −78.73% to 144.01% better parent and −78.73% to 130.75% over standard parent which indicates the presence of both positive and negative heterosis. The highest heterosis (144.01%) over better parent heterosis was obtained from Double Rice × Pokkali. Longer grain length is considered an important trait regarding grain quality. Long grains have a good market value over short grains. So, heterosis in a positive direction is desirable for this trait. The heterotic effects for the character grain length ranged between 18.16% and −12.18% over the mid parent, 14.52% and −32.25% over the better parent, and 17.99% and −25.71% over the standard parent (Table 2). Nonetheless, the positive estimate of heterosis for 1000-grain weights is desirable as it is an important trait influencing yield but both positive and negative heterotic values for 1000-grain weight among different cross combinations were observed. The highest positive value of heterobeltiosis was demonstrated by Muktagacha × Chinigura, while the highest desirable positive estimate of standard heterosis was exhibited by Muktagacha × Chinigura. Furthermore, Pokkali × Chinigura was recorded for producing the highest heterobeltiosis for tillers/hill (173.17%) as well as grain yield/plant (71.05%). Furthermore, Double Rice × Kotoktara and Basmati × Double Rice revealed maximum standard heterosis for grain length (15.95%) and grain test weight (20.56%), respectively. A few hybrids also revealed higher heterotic values (Table 2) for grain yield per plant over the better and standard parent. For instance, superior heterobeltiosis was recorded in cross combination Double Rice × Pokkali (95.40%) followed by Pokkali × Chinigura (71.05%) while Pokkali × Chinigura revealed the topmost standard heterosis as 262.08% exploring its potentiality for higher yield.
Table 2.
Percentage of heterosis over mid, better, and standard parent for ten yield and yield-related traits in a 7 × 7 diallel population of rice.
3.5. Pairwise Genetic Distance of Parents Relying on Heatmap
To determine the pairwise difference between genotypes, the concept of dissimilarity may be used in a more general way (Figure 5). The dissimilarity matrix combined with heatmap is an effective method for visualizing genetic distance (GD) among genotypes. The pairwise genetic distance values were graphically represented by a heatmap in Figure 4, which were indicated by gradient colors from red (low value) to light green (high value). Observing the heatmap it is observed that the highest genetic distance was found between Chinigura and Pokkali followed by Muktagacha and Pokkali revealing a higher degree of dissimilarity among these genotypes. Due to having considerable genetic diversity, crossing between Chinigura and Pokkali, Muktagacha and Pokkali would yield potential progeny as there is a thumb rule that the more will be genetic diversity greater will be the possibility of getting a heterotic combination.
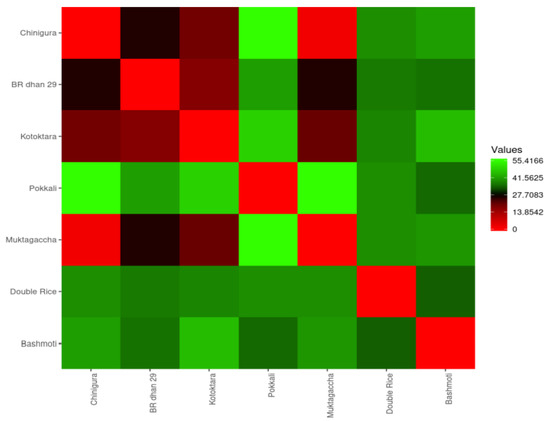
Figure 5.
Heatmap generated exploiting the dissimilarity matrix revealing the genetic distances among the genotypes. Light green color denotes the highest dissimilarity and red indicates the lowest genetic distance among the genotypes.
3.6. Cluster Analysis of Parental Genotypes Based on RAPD
The unweighted neighbor-joining tree exhibited was constructed using DARwin software 6.0 which revealed genetic relatedness among the seven rice genotypes using seven RAPD primers (Figure 6). The rice genotypes were grouped into three primary groups, demonstrating that the seven parental rice genotypes have a lot of genetic variabilities (Figure 6). In particular, cluster B (Muktagacha, Chinigura, Kotoktara) and cluster C (Double rice, Pokkali, Basmati) both contain the equal and the highest number of parents, whereas cluster A possesses only one (BRRI dhan29) genotype. Genetically similar genotypes clustered together, while dissimilar genotypes were placed in a separate cluster. Thus, cluster analysis revealed a significant genetic diversity among the exploited genotypes at the genotypic level. The possession of a significant amount of variability among inbred lines grouped into divergent clusters indicated their suitability for hybridization and breeding program to develop potential hybrids [31]. Genetic architecture explored by cluster analysis signifying that progeny derived from cross exploiting the individual from cluster B, Muktagacha with either individual from group C or A would lead to the accumulation of favorable genes underlying a prominent heterotic combination.
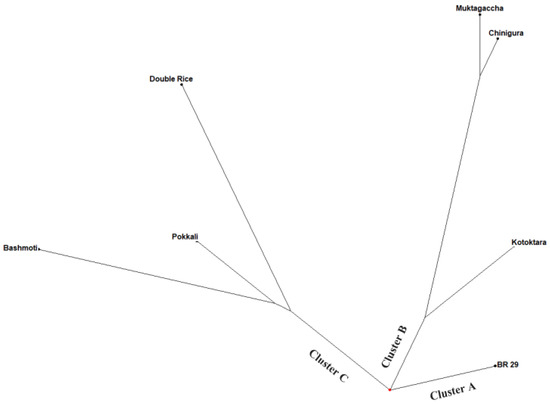
Figure 6.
An unweighted neighbor-joining tree showing the genetic relationships among seven rice genotypes (parental line) based on data obtained from 7 RAPD markers.
3.7. Correlation between Markers Generated GD with SCA, MPH, BPH, and SPH
The genetic distance (GD) among the rice parental lines demonstrated a positive and significant relationship with a specific combining ability (SCA) for all of the traits under study where maximum correlation (Figure 7) was observed for the traits 1000-grain weight (0.38) and tillers/hill (0.31), respectively. Furthermore, the magnitudes of correlation of genetic distances with mid-parent and better parent, as well as standard parent heterosis for yield and other traits particularly tillers/hill and 1000-grain weight were found highly significant. As a result, this highly significant correlation between RAPD-generated genetic distance and heterosis in exploited rice genotypes indicates that the accuracy of RAPD markers in predicting hybrid performance with the present set of individuals is very high.
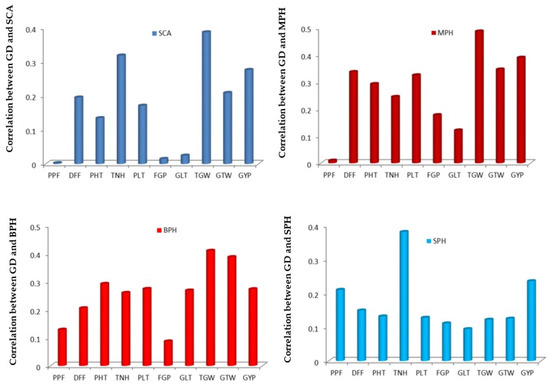
Figure 7.
Correlation between RAPD marker generated genetic distance (GD) with SCA, MPH, BPH and SPH of ten yield and yield related traits in rice. SCA = Specific Combining Ability, MPH = Mid- parent heterosis, BPH = Better-parent heterosis, SPH = Standard-parent heterosis, PPF = Percent pollen fertility, DFF = Days to first flowering, PHT = Plant height (cm), TNH = Tillers/hill, PLT = Panicle length (cm), FGP = Filled grains/panicle, GLT = Grain length (mm), TGW = 1000-grain weight (g), GTW = Grain test weight (g/5 mL), GYP = Grain yield/plant (g).
3.8. Maker-Based Prediction of Yield Heterosis
The unweighted neighbor-joining tree revealed that the exploited seven rice parental lines used for the development of F1 hybrids were clustered into three major groups indicating the existence of considerable genetic diversity among these parental lines. Moreover, progeny derived from the cross exploiting the individual from the different groups will lead to the accumulation of more favorable genes leading to a prominent heterotic combination. The correlation between RAPD marker-generated genetic distance (GD) and specific combining ability was found highly significant in almost all of the agronomic traits. The genetic distance (GD) among the rice parental lines was also significantly correlated with mid-parent and better parent, as well as standard parent heterosis for yield and other traits signifying the possibility of mining favorable heterotic combinations using RAPD markers. Moreover, the correlation of RAPD-based genetic diversity with SCA or heterosis imposed to assume that there might be a putative candidate gene for those trait(s) because of nonspecific amplification of RAPD marker definite physical position is unknown. So, converting this nonspecific marker to a location-specific marker such as SSR is recommended to explore the specific position of genes of studied traits.
4. Discussion
Knowledge of genetic interactions between crosses is crucial for plant breeding, as it contributes to the fundamental criteria for selecting combinations. Selection solely based on desirable traits is insufficient to create progeny with high genetic potential. The identification of high combining ability genitors is necessary for maximum heterosis expression. The strategies adopted in breeding programs depend on the genetic analysis of the traits of interest because they lead to a better understanding of the genetic relationship between crossed lineages; thereby the diallel crosses are useful for predicting the best combinations between parents and segregating populations.
The analysis of variance for parents and F1 hybrids was significant at p ≤ 0.01 for all the traits. These significant variances suggested that there was a wide diversity in the genotypes for the studied traits. Similar variability and diversity were also observed in different rice cultivars [32,33,34,35,36,37,38,39,40,41], vegetable amaranth [42,43,44,45,46,47,48,49] and cauliflower [50]. Again, analysis of variances for combining ability (GCA and SCA) revealed noteworthy variations for the studied traits. Moreover, the mean square values for the GCA were higher than the SCA for all the traits, signifying the preponderance of additive gene action for controlling the traits which are corroborative to the previous studies in rice [51,52,53].
The goal of the diallel analysis is to divide the data’s total variation into GCA and SCA of parents and crosses, respectively. It also explains how the quantity of additive and non-additive gene action affects parents’ ability to generate superior offspring [54]. For all traits studied, the GCA and SCA effects were profound, implying the presence of large genetic variants. Nevertheless, GCA variances were considerably higher in magnitude than SCA variances for all of the characters in our analysis, indicating a prevalence of additive gene effects for the acquisition of these traits. Therefore, as additive gene action is predominant, selection during the early generations would be effective. Similarly, if non-additive gene effects are present, selection should be carried out in later generations, when these effects would have been fixed in homozygous lines [55]. In this manner, the importance of both GCA and SCA variances for yield and yield-contributing traits in rice has been reported [56,57]. Moreover, the specific combining ability (SCA) of a cross is the estimation and understanding of the non-additive gene action of a trait signifying the selection of a superior hybrid combination. we have observed the generation of promising F1 hybrids for different traits derived from high × high, high × low, low × high, average × low, and low × low general combiner parents. Several previous studies have shown good general combiner parents from high × high, high × low, low × high, average × low, and low × low general combiner parents for grain yield and its related traits in rice which conformed to the current results [58,59,60]. In general, an appreciable amount of the SCA effects expressed by low × low crosses might be ascribed to the dominance × dominance type of non-allelic gene action produced over-dominance and are non-fixable. It appears that the superior performance of most hybrids may be largely due to epistatic interaction [61]. According to the SCA effect estimates, none of the cross combinations in this experiment had substantial and desired SCA effects for all of the factors at the same time, indicating that no single combination was desirable for all attributes. These results are in complete agreement with earlier findings [62].
Although, the SCA effect in a positive direction for grain yield performance is desirable as well as superior SCA effects for grain yield in rice are very common [6,56] only eight out of twenty-one crosses revealed a significant and positive SCA effect. This phenomenon suggested a more careful selection of inbred lines for the generation of the Diallel population. In addition, most of the high-yielding hybrids (BRRI dhan29 × Chinigura), (Double Rice × Pokkali), and (Pokkali × Chinigura) originated from high × low GCA combinations signifying that the genetic regulator of grain yield underlying the phenomena of additive × dominance gene interaction.
Molecular markers are an effective tool for determining genetic variability and diversity within landraces, populations, and species. Several attempts have been made to use molecular markers to analyze the hybrid parents’ genetic variation and heterosis in their offspring. In the present study, a cluster analysis was performed to measure genetic diversity and closeness among the parents. In the cluster analysis, parental genotypes are divided into three groups indicating that the parental genotypes have a lot of genetic diversity among themselves. Thus, cluster analysis revealed a significant genetic diversity among the exploited genotype at both phenotypic and genetic levels, however, such analysis fails to cluster the base material into seven unique clades according to their origin of development while it was expected that founder parents would be significantly different from each other as their phenotypic performance was significantly different from each other. Furthermore, genetic architecture revealed by cluster analysis relying on RAPD marker-generated genetic distance signifying that parents derived from the diverse group would lead to the accumulation of favorable genes leading to a prominent heterotic combination.
Our tested genotypes showed high heterosis for grain yield and yield contributing traits. High heterotic hybrids for grain yield and yield contributing traits were also reported in different cultivars of rice [63,64] which confirmed our present findings. Previous research has also shown that heterosis is mostly determined by the genetic variation and features of the parents’ interaction; the more the genetic variation and genetic distance, the greater the apparent hybrid vigor [65]. However, there is now a plethora of research testing the utility of DNA markers for predicting heterosis and hybrid performance. There are two opposing views about heterosis prediction: some scientists believe that molecular markers can be utilized to forecast heterosis [20,65,66], while others believe that molecular markers cannot predict heterosis [67]. The genetic distance (GD) between parental lines was significantly correlated with a specific combining ability (SCA) for all of the traits under study. GD exhibited a positive and significant correlation with mid-parent heterosis (MPH), better-parent heterosis (BPH), and also for standard-parent heterosis for all the traits. Consequently, such a significant correlation among the aforesaid occurrences in exploited rice genotypes indicated that the prediction of hybrid performance using RAPD markers with the present set of individuals was very efficient. Other findings on the association between genetic distance and heterosis, particularly in hybrid rice, support this conclusion [16,65]. In this study, molecular markers were found to be effective in predicting hybrid performance for the majority of attributes and marker-based genetic distance displayed a substantial relation with 1000-grain weight heterosis in hybrid combination. The information generated from this study would be useful for future rice breeding strategies.
5. Conclusions
Highly significant (p < 0.01) differences were observed among the genotypes for all of the traits investigated and the hybrids reflected the superiority over their parental lines in most of the yield and yield-contributing traits. Double Rice × Pokkali was the best specific combiner for pollen fertility (%), panicle length (cm), filled grains/panicle, and grain yield/plant (g) while Muktagacha × BRRI dhan29 and Basmati × Double Rice were the best specific combiners for grain length (mm) and 1000-grain weight (g), respectively. Moreover, Double Rice × Pokkali was accounted for producing maximum heterobeltiosis regarding the traits panicle length (cm) and filled grains/panicle. The unweighted neighbor-joining tree clustered the exploited seven parental lines into three major groups indicating the presence of considerable genetic diversity among them. The correlation between RAPD-generated genetic distance and mid-parent, better-parent, as well as standard parent heterosis regarding all of the agronomic traits, was significant. Thus, exploration of definite genomic regions underlying the candidate genes for respective traits relying on such RAPD-generated data following an advanced molecular marker approach would pave the path of mining out yielder heterotic combinations.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy12081797/s1, Table S1: Characteristic features of the rice parental genotypes used in the experiment; Table S2: Marker used for the genotyping par-ents; Table S3: PCR ingredients for RAPD Markers; Table S4: Analysis of variance (ANOVA) of the ten yield and yield contributing traits in a 7 × 7 diallel population of rice; Table S5: Mean values of ten yield and yield-related traits of the parents and F1 hybrids in a 7 × 7 diallel population of rice; Table S6: Estimates of general combining ability (GCA) effects for ten yield and yield related characters in a 7 × 7 diallel population of rice.
Author Contributions
Conceptualization, M.S.R. and U.S.; methodology, M.S.R. and U.S.; software, M.S.R., U.S., S.A. and M.H.S.; validation, M.S.R., U.S., M.A.H.S., S.O., S.A. and M.H.S.; formal analysis, M.M.R.; investigation, M.M.R., resources, M.S.R. and M.M.R.; data curation, M.M.R., writing—original draft preparation, M.M.R., U.S., S.A. and M.H.S.; writing—review and editing, M.S.R., U.S., M.A.H.S., S.O., S.A. and M.H.S.; visualization, M.S.R., U.S. and M.A.H.S.; supervision, M.S.R., U.S. and M.A.H.S.; project administration, M.S.R. and M.H.S.; funding acquisition M.S.R., S.A. and M.H.S. All authors have read and agreed to the published version of the manuscript.
Funding
Research Monitoring Committee (RMC) fund of the Bangabandhu Sheikh Mujibur Rahman Agricultural University, Gazipur-1706, Bangladesh. The study was also supported by the Researchers Supporting Project number (RSP-2021/194), King Saud University, Riyadh, Saudi Arabia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data recorded in the current study are available in all Tables and Figures of the manuscript.
Acknowledgments
We would like to acknowledge the financial support provided by the RMC, Grant SI No. 001 (2018–2021). The authors would like to extend their sincere appreciation to the Researchers Supporting Project number (RSP-2021/194), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Gnanamanickam, S.S. Rice and Its Importance to Human Life. In Biological Control of Rice Diseases; Springer: Dordrecht, The Netherlands, 2009; pp. 1–11. [Google Scholar]
- Hooker, J.D. The Flora of British India; Reeve Co.: Stowmarket, UK, 1979; Volume 2L. [Google Scholar]
- Roy, R.N.; Misra, R.V. Economic and Environmental Impact of Improved Nitrogen Management in Asian Rice-Farming Systems; Sustainable rice production for food security. In Proceedings of the 20th Session of the International Rice Commission, Bangkok, Thailand, 23–26 July 2002; pp. 171–180. [Google Scholar]
- Bangladesh Bureau of Statistics (BBS). Monthly Statistical Bulletin, Statistics & Informatics Division (SID); Ministry of Planning Government of The People’s Republic of Bangladesh: Dhaka, Bangladesh, 2018; pp. 49–69.
- Food and Agricultural Organization (FAO). Rice Market Monitor; Food and Agricultural Organization (FAO): Bangkok, Thailand, 2017; pp. 1–42.
- Gramaje, L.V.; Caguiat, J.D.; Enriquez, J.O.S.; Dela Cruz, Q.D.; Millas, R.A.; Carampatana, J.E.; Tabanao, D.A.A. Heterosis and Combining Ability Analysis in CMS Hybrid Rice. Euphytica 2020, 216, 14. [Google Scholar] [CrossRef]
- Ahangar, L.; Ranjbar, G.A.; Nouroozi, M. Estimation of Combining Ability for Yield and Yield Components in Rice (Oryza sativa L.) Cultivars Using Diallel Cross. Pak. J. Biol. Sci. 2008, 11, 1278–1281. [Google Scholar] [CrossRef]
- Sprague, G.F.; Tatum, L.A. General vs. Specific Combining Ability in Single Crosses of Corn. J. Agron. 1942, 34, 923–932. [Google Scholar] [CrossRef]
- Veeresha, B.A.; Hanamaratti, N.G.; Salimath, P.M. Heterosis and Combining Ability Studies for Yield and Productivity Traits in Rice. Int. J. Agric. Res. 2015, 4, 120–126. [Google Scholar]
- Griffing, B. Concept of General and Specific Combining Ability in Relation to Diallel Crossing Systems. Aust. J. Biol. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, H.; Xie, W.; Xia, D.; Gao, G.; Zhang, Q.; He, Y. Genome-Wide Association Analyses Reveal the Genetic Basis of Combining Ability in Rice. Plant Biotechnol. 2019, 17, 2211–2222. [Google Scholar] [CrossRef]
- Masood, J.; Ghulam, H.; Iftikhar, K.; Raziuddin, N. Estimates of Heterosis and Heterobeltiosis for Morphological Traits in Wheat (Triticum aestivum L.). Pak. J. Biol. Sci. 2005, 8, 1261–1264. [Google Scholar]
- Kempthorne, O. An Introduction of Genetic Statistics; John Willey & Sons Inc.: New York, NY, USA, 1957; pp. 468–473. [Google Scholar]
- Melchinger, A.E.; Messmer, M.M.; Lee, M.; Woodman, W.L.; Lamkey, K.R. Diversity and Relationships among US Maize Inbreds Revealed by Restriction Fragment Length Polymorphisms. Crop Sci. 1991, 31, 669–678. [Google Scholar] [CrossRef]
- Krystkowiak, K.; Adamski, T.; Surma, M.; Kaczmarek, Z. Relationship Between Phenotypic and Genetic Diversity of Parental Genotypes and the Specific Combining Ability and Heterosis Effects in Wheat (Triticum aestivum L.). Euphytica 2009, 165, 419–434. [Google Scholar] [CrossRef]
- Xiao, J.; Li, J.; Yuan, L.; McCouch, S.R.; Tanksley, S.D. Genetic Diversity and Its Relationship to Hybrid Performance and Heterosis in Rice as Revealed by PCR-Based Markers. Theor. Appl. Genet. 1996, 92, 637–643. [Google Scholar] [CrossRef]
- Zhao, M.F.; Li, X.H.; Yang, J.B.; Xu, C.G.; Hu, R.Y.; Liu, D.J.; Zhang, Q. Relationship Between Molecular Marker Heterozygosity and Hybrid Performance in Intra and Inter-Subspecific Crosses of Rice. Plant Breed. 1999, 118, 139–144. [Google Scholar] [CrossRef]
- Wang, M.; Zhu, Z.; Tan, L.; Liu, F.; Fu, Y.; Sun, C.; Cai, H. Complexity of Indica-Japonica Varietal Differentiation in Bangladesh Rice Landraces Revealed by Microsatellite Markers. Breed. Sci. 2013, 63, 227–232. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Martin, J.M.; Talbert, L.E.; Lanning, S.P.; Blake, N.K. Hybrid Performance in Wheat as Related to Parental Diversity. Crop Sci. 1995, 35, 104–108. [Google Scholar] [CrossRef]
- Smith, O.S.; Smith, J.S.C.; Bowen, S.L.; Tenborg, R.A.; Wall, S.J. Similarities Among a Group of Elite Maize Inbreds as Measured by Pedigree, F1 Grain Yield, Grain Yield, Heterosis, and RFLPs. Theor. Appl. Genet. 1990, 80, 833–840. [Google Scholar] [CrossRef]
- Betrán, F.J.; Ribaut, J.M.; Beck, D.; De León, D.G. Genetic Diversity, Specific Combining Ability, and Heterosis in Tropical Maize Under Stress and Nonstress Environments. Crop Sci. 2003, 43, 797–806. [Google Scholar] [CrossRef]
- Menkir, A.; Melake-Berhan, A.; Ingelbrecht, I.; Adepoju, A. Grouping of Tropical Mid-Altitude Maize Inbred Lines on the Basis of Yield Data and Molecular Markers. Theor. Appl. Genet. 2004, 108, 1582–1590. [Google Scholar] [CrossRef]
- Barata, C.; Carena, M.J. Classification of North Dakota Maize Inbred Lines into Heterotic Groups Based on Molecular and Testcross Data. Euphytica 2006, 151, 339–349. [Google Scholar] [CrossRef]
- Raihan, M.S.; Newaz, M.A. Combining Ability for Quantitative Attributes in Lablab Bean (Lablab purpureus L.). Bangladesh J. Plant Breed. Genet. 2008, 21, 29–34. [Google Scholar] [CrossRef]
- Rai, B. Heterosis breeding. Agrobiol. Publ. 1979, 4, 65. [Google Scholar]
- Ferrari, C.D.S.; Valente, L.L.; Brod, F.C.A.; Tagliari, C.; Sant’Anna, E.S.; Arisi, A.C.M. Evaluation of Polymerase Chain Reaction and DNA Isolation Protocols for Detection of Genetically Modified Soybean. Int. J. Food Sci. Technol. 2007, 42, 1249–1255. [Google Scholar] [CrossRef]
- Peakall, R.O.D.; Smouse, P.E. GENALEX 6: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research. Mol. Ecol. 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–291. [Google Scholar] [CrossRef]
- Perrier, X.; Jacquemoud-Collet, J.P. Dissimilarity Analysis and Representation for Windows. 2006. Available online: https://darwin.cirad.fr/darwin (accessed on 11 March 2022).
- Heatmapper. Available online: http://www.heatmapper.ca/ (accessed on 17 May 2022).
- Kumari, R.; Singh, A.K.; Sharma, V.K. Genetic Divergence Studies for Morpho-Agronomical Traits in Maize (Zea mays L.) Inbreds. Int. J. Curr. Microbiol. 2018, 2, 3589–3594. [Google Scholar]
- Hasan-Ud-Daula, M.; Sarker, U. Variability, Heritability, Character Association, and Path Coefficient Analysis in Advanced Breeding Lines of Rice (Oryza sativa L.). Genetika 2020, 52, 711–726. [Google Scholar] [CrossRef]
- Ali, M.A.; Sarker, U.; Mian, M.A.K.; Islam, M.A.; Johora, F.T. Estimation of Genetic Divergence in Boro Rice (Oryza sativa L.). Int. J. BioRes. 2014, 16, 28–36. [Google Scholar]
- Rai, P.K.; Sarker, U.; Roy, P.C.; Islam, A. Character Association in F4 Generation of Rice (Oryza sativa L.). Bangladesh J. Plant Breed. Genet. 2013, 26, 39–44. [Google Scholar] [CrossRef]
- Nath, J.K.; Sarker, U.; Mian, M.A.K.; Hossain, T. Genetic Divergence in T. Aman Rice. Ann. Bangladesh Agric. 2008, 12, 51–60. [Google Scholar]
- Siddique, M.N.A.; Sarker, U.; Mian, M.A.K. Genetic diversity in restorer line of rice. In Proceedings of the International Conference on Plant Breeding and Seed for Food Security, Dhaka, Bangladesh, 12 September 2009; Bhuiyan, M.S.R., Rahman, L., Eds.; Plant Breeding and Genetics Society of Bangladesh: Dhaka, Bangladesh, 2009; pp. 137–142. [Google Scholar]
- Hasan, M.R.; Sarker, U.; Hossain, M.A.; Huda, K.M.K.; Mian, M.A.K.; Hossain, T.; Zahan, M.S.; Mahmud, M.N.H. Genetic Diversity in Micronutrient Dense Rice and Its Implication in Breeding Program. Ecofriendly Agril. J. 2012, 5, 168–174. [Google Scholar]
- Ganapati, R.K.; Rasul, M.G.; Mian, M.A.K.; Sarker, U. Genetic Variability and Character Association of T-Aman Rice (Oryza sativa L.). Int. J. Plant Biol. Res. 2014, 2, 1–4. [Google Scholar]
- Karim, D.; Sarker, U.; Siddique, M.N.A.; Mian, M.A.K.; Hasnat, M.Z. Variability and Genetic Parameter Analysis in Aromatic Rice. Int. J. Sustain. Crop Prod. 2007, 2, 15–18. [Google Scholar]
- Karim, D.; Siddique, M.N.A.; Sarker, U.; Hasnat, Z.; Sultana, J. Phenotypic and Genotypic Correlation Co-Efficient of Quantitative Characters and Character Association of Aromatic Rice. J. Biosci. Agric. Res. 2014, 1, 34–46. [Google Scholar] [CrossRef]
- Hasan, M.J.; Kulsum, M.U.; Majumder, R.R.; Sarker, U. Genotypic Variability for Grain Quality Attributes in Restorer Lines of Hybrid Rice. Genetika 2020, 52, 973–989. [Google Scholar] [CrossRef]
- Sarker, U.; Azam, M.G.; Talukder, M.Z.A. Genetic Variation in Mineral Profiles, Yield Contributing Agronomic Traits, and Foliage Yield of Stem Amaranth. Genetika 2022, 54, 91–108. [Google Scholar] [CrossRef]
- Rashad, M.M.I.; Sarker, U. Genetic Variations in Yield and Yield Contributing Traits of Green Amaranth. Genetika 2020, 52, 393–407. [Google Scholar] [CrossRef]
- Sarker, U.; Iqbal, M.A.; Hossain, M.N.; Oba, S.; Ercisli, S.; Muresan, C.C.; Marc, R.A. Colorant Pigments, Nutrients, Bioactive Components, and Antiradical Potential of Danta Leaves (Amaranthus lividus). Antioxidants 2022, 11, 1206. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Alsanie, W.F.; Gaber, A. Characterization of Phytochemicals, Nutrients, and Antiradical Potential in Slim Amaranth. Antioxidants 2022, 11, 1089. [Google Scholar] [CrossRef]
- Sarker, U.; Rabbani, M.G.; Oba, S.; Eldehna, W.M.; Al-Rashood, S.T.; Mostafa, N.M.; Eldahshan, O.A. Phytonutrients, Colorant Pigments, Phytochemicals, and Antioxidant Potential of Orphan Leafy Amaranthus Species. Molecules 2022, 27, 2899. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Ercisli, S.; Assouguem, A.; Alotaibi, A.; Ullah, R. Bioactive Phytochemicals and Quenching Activity of Radicals in Selected Drought-Resistant Amaranthus tricolor Vegetable Amaranth. Antioxidants 2022, 11, 578. [Google Scholar] [CrossRef]
- Hossain, M.N.; Sarker, U.; Raihan, M.S.; Al-Huqail, A.A.; Siddiqui, M.H.; Oba, S. Influence of Salinity Stress on Color Parameters, Leaf Pigmentation, Polyphenol and Flavonoid Contents, and Antioxidant Activity of Amaranthus lividus Leafy Vegetables. Molecules 2022, 27, 1821. [Google Scholar] [CrossRef]
- Sarker, U.; Lin, Y.P.; Oba, S.; Yoshioka, Y.; Ken, H. Prospects and Potentials of Underutilized Leafy Amaranths as Vegetable Use for Health Promotion. Plant Physiol. Biochem. 2022, 182, 104–123. [Google Scholar] [CrossRef]
- Prodhan, M.M.; Sarker, U.; Hoque, M.A.; Biswas, M.S.; Ercisli, S.; Assouguem, A.; Ullah, R.; Almutairi, M.H.; Mohamed, H.R.H.; Najda, A. Foliar Application of GA3 Stimulates Seed Production in Cauliflower. Agronomy 2022, 12, 1394. [Google Scholar] [CrossRef]
- Azad, A.K.; Sarker, U.; Ercisli, S.; Assouguem, A.; Ullah, R.; Almeer, R.; Sayed, A.A.; Peluso, I. Evaluation of Combining Ability and Heterosis of Popular Restorer and Male Sterile Lines for the Development of Superior Rice Hybrids. Agronomy 2022, 12, 965. [Google Scholar] [CrossRef]
- Azam, M.G.; Sarker, U.; Hossain, M.A.; Iqbal, M.S.; Islam, M.R.; Hossain, M.F.; Ercisli, S.; Kul, R.; Assouguem, A.; AL-Huqail, A.A.; et al. Genetic Analysis in Grain Legumes [Vigna radiata (L.) Wilczek] for Yield Improvement and Identifying Heterotic Hybrids. Plants 2022, 11, 1774. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Biswas, P.S.; Prasad, B.; Mian, M.A.K. Heterosis and Genetic Analysis in Rice Hybrids. Pak. J. Biol. Sci. 2002, 5, 1–5. [Google Scholar] [CrossRef]
- Bidhendi, M.Z.; Choukan, R.; Darvish, F.; Mostafavi, K.; Hervan, E.M. Determination of Combining Abilities and Heterotic Patterns of Fourteen Medium to Late Maturing Iranian Maize Inbred Lines Using Diallel Mating Design. Afr. J. Biotechnol. 2011, 10, 16854–16865. [Google Scholar]
- Geleta, L.F.; Labuschagne, M.T. Combining Ability and Heritability for Vitamin C and Total Soluble Solids in Pepper (Capsicum annuum L.). J. Sci. Food Agric. 2006, 86, 1317–1320. [Google Scholar] [CrossRef]
- Sari, W.K.; Nualsri, C.; Junsawang, N.; Soonsuwon, W. Combining Ability and Heritability for Yield and Its Related Traits in Thai Upland Rice (Oryza sativa L.). Agric. Nat. Resour. 2020, 54, 229–236. [Google Scholar]
- Sravan, T.; Jaiswal, H.K.; Waza, S.A.; Priyanka, K. Heterosis for Yield and Yield Attributes in Rice (Oryza sativa L.). J. Appl. Nat. Sci. 2016, 8, 622–625. [Google Scholar] [CrossRef]
- Alam, A.S.M.S.; Sarker, U.; Mian, M.A.K. Line × Tester Analysis in Hybrid Rice (Oryza sativa L.). Ann. Bangladesh Agric. 2007, 11, 37–44. [Google Scholar]
- Sarker, U.; Mian, M.A.K. Line × Tester Analysis for Yield and Its Components in Rice (Oryza sativa L.). J. Asiat. Soc. Bangladesh Sci. 2002, 28, 71–81. [Google Scholar]
- Sarker, U.; Rasul, M.G.; Mian, M.A.K. Combining Ability of CMS and Restorer Lines in Rice. Bangladesh J. Plant Breed. Genet. 2003, 16, 1–7. [Google Scholar]
- Begum, S.; Alam, S.S.; Omy, S.H.; Amiruzzaman, M.; Rohman, M.M. Inheritance and Combing Ability in Maize Using a 7 × 7 Diallel Cross. J. Plant Breed. Crop Sci. 2018, 10, 239–248. [Google Scholar]
- Kahani, F.; Hittalmani, S.; Erfani, R.; Haradari, C. Heterotic Effects and Combining Ability for Yield Traits in Rice Developed for Semi-Dry Aerobic Cultivation. SABRAO J. Breed. Genet. 2018, 50, 46–61. [Google Scholar]
- Sarker, U.; Rasul, M.G.; Mian, M.A.K. Heterosis and Combining Ability in Rice. Bangladesh J. Plant Breed. Genet. 2002, 15, 17–26. [Google Scholar]
- Islam, M.M.; Sarker, U.; Rasul, M.G.; Rahman, M.M. Heterosis in Local Boro Rice (Oryza sativa L.). Bangladesh J. Plant Breed. Genet. 2010, 23, 19–30. [Google Scholar] [CrossRef]
- Zhang, Q.; Gao, Y.J.; Yang, S.H.; Ragab, R.A.; Maroof, M.S.; Li, Z.B. A Diallel Analysis of Heterosis in Elite Hybrid Rice Based on RFLPs and Microsatellites. Theor. Appl. Genet. 1994, 89, 185–192. [Google Scholar] [CrossRef]
- Jaikishan, I.; Rajendrakumar, P.; Ramesha, M.S.; Viraktamath, B.C.; Balachandran, S.M.; Neeraja, C.N.; Sundaram, R.M. Prediction of Heterosis for Grain Yield in Rice Using ‘Key‘ Informative EST-SSR Markers. Plant Breed. 2010, 129, 108–111. [Google Scholar] [CrossRef]
- Joshi, S.P.; Bhave, S.G.; Chowdari, K.V.; Apte, G.S.; Dhonukshe, B.L.; Lalitha, K.; Gupta, V.S. Use of DNA Markers in Prediction of Hybrid Performance and Heterosis for a Three-Line Hybrid System in Rice. Biochem. Genet. 2001, 39, 179–200. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).