Abstract
The implementation of new planting systems in almond (Prunus amygdalus (L.) Batsch, syn P. dulcis (Mill.)) orchards for more sustainable practices has made tree architecture increasingly relevant as an important selection trait in plant breeding. Multiple features define the three-dimensional structure of the tree, with shoot production being the most important. Shoots can develop after a period of rest (proleptic shoots) or immediately (sylleptic shoots), at the same time as the extension of the parent shoot. Scion/rootstock interactions may change the proportion of proleptic and sylleptic shoots and thus alter tree architecture. To study this effect, we analyzed shoot formation on three almond scion cultivars grafted onto five interspecific hybrid rootstocks. The type of shoot (proleptic or sylleptic) and internode number per shoot were collected for two-year-old branches on three-year-old unpruned trees. We found that rootstock genotypes can alter the pattern of shoot production, especially the number of sylleptic shoots formed. More information on the molecular response is needed to comprehend the biological processes behind these differential phenotypes, which appear to be modulated by both apical dominance and apical control.
1. Introduction
Several aspects define the three-dimensional structure of a tree. The combination of all these scion phenotype features can be affected by how the rootstock and scion interact [1,2,3,4,5]. Scion/rootstock interaction determines multiple aspects of tree development, such as flowering time, fruit quality, yield, tree vigor and tree architecture [6,7,8,9].
Tree architecture is defined by four major features: (i) temporal growth pattern; (ii) branching pattern; (iii) morphological differentiation of axes; and (iv) sexual differentiation of meristems [10]. One of these four features, branching pattern, is a key part of the definitive three-dimensional structure of the tree. The distribution of branches along the parent axis largely determines the tree architecture, differentiating between rhythmic branching (when branched nodes are followed by unbranched nodes), and diffuse branching (when branches are established uniformly through the axis) [11]. Bud ontogeny also determines branching patterns. If an axillary shoot develops immediately, without the parent meristem going through a period of rest or dormancy, and the shoot extends at the same time as the extension of the parent axis, then this forms a sylleptic shoot. However, if the parent meristem forms a bud that remains inactive and develops after a period of rest or dormancy, which in Prunus trees is typically in the following growing season, the bud is said to produce a proleptic shoot [11,12].
The dynamics between proleptic and sylleptic shoot development, and their effect on other architectural traits, including fruiting, have previously been studied in different almond (Prunus amygdalus (L.) Batsch, syn P. dulcis (Mill)) cultivars. Negrón et al. [13] reported that the pattern of shoot production and fruiting was determined by cultivar genotype. The formation of proleptic and sylleptic shoots has also been analyzed under different environments or experimental treatments, for example, water deficiency or pruning [14,15]. In peach (P. persica (L.) Batsch), it has been reported that more floral buds are produced on proleptic than sylleptic shoots [16]. Therefore, the overall proportion of these shoot types can largely alter the commercial viability of Prunus cultivars, including almond.
In this study, we analyze the effect of different hybrid rootstocks on the formation of proleptic and sylleptic shoots on two-year-old branches of almond trees. Our hypothesis is that different scion/rootstock combinations will change the patterns of the axillary shoot production of almond trees.
2. Materials and Methods
2.1. Tree Population
In this study, three almond cultivars grafted onto five different commercial rootstocks, resulting in a total of fifteen different combinations, were used. The scion cultivars selected were ‘Isabelona’ (syn. ‘Belona’), ‘Guara’ and ‘Lauranne’; each of these are important commercial cultivars in Spain and Portugal. The three cultivars present different vigor and branching architectures [17]. The rootstocks were selected to represent a range of vigor responses in the grafted scion: Pilowred® (GN8), ‘Densipac’ (Rootpac® 20), ‘Nanopac’ (Rootpac® 40), ‘Replantpac’ (Rootpac® R) and Garnem® (GN15). All were hybrid rootstocks from different origins. Garnem® and Pilowred® are both almond × peach (P. amygdalus (L.) Batsch, syn P. dulcis (Mill.). × P. persica (L.) Batsch) hybrid rootstocks, while the three others came from the commercial Rootpac® series including Rootpac® 40 (P. amygdalus (L.) Batsch, syn P. dulcis (Mill.). × P. persica (L.) Batsch), Rootpac® 20 (P. cerasifera × P. besseyi) and Rootpac® R (P. cerasifera × P. amygdalus (L.) Batsch, syn P. dulcis (Mill.)). Grafted plants were supplied by the Agromillora Iberia S.L. nursery in 2018 (Barcelona, Spain). Trees were budded in July 2018 and planted during October 2018 at the Centro de Investigación y Tecnología Agroalimentaria de Aragón (CITA) experimental orchard El Vedado Bajo el Horno (Zuera, Zaragoza, 41°51′46.5″ N 0°39′09.2″ W). Trees were planted with a single trunk supported by a wooden stake. Trees were then left without pruning so that they could express their natural growth habit, unaltered. Conventional orchard practices were used for weed control and drip irrigation. The soil type was calcareous with pH around 7–8.
2.2. Data Collection
Data collection was carried out on two-year-old branches during winter 2021 from 150 trees with ten trees per scion/rootstock combination. Three parameters were collected for a whole branch of each tree. Total branch length (Length) and number of internodes (Nb_IN) were recorded from the trunk to the apex of the branch, and average internode length (IN_L) was calculated from those two measures. The fate of each node of the branch was annotated, distinguishing between blind nodes, proleptic shoots and sylleptic shoots. Data were collected for the whole branch after the second year of growth. Hence, while two phases of sylleptic shoot production were recorded on one- and two-year-old wood, proleptic shoot growth was only recorded on two-year-old wood, on the branch that grew during the first year.
2.3. Statistical Analysis
All statistical analyses were carried out on the R platform (https://cran.r-project.org/ (accessed on 10 November 2021). Significant differences in phenotypic data were evaluated using an ANOVA test. These were assessed with a Tukey’s test (p < 0.05) using the agricolae R package (https://CRAN.R-project.org/package=agricolae (accessed on 12 November 2021).
2.4. Shoot Probability
To represent the probability of a given node presenting a blind node, proleptic, or sylleptic shoot, the results obtained for the node fate description were translated to a branch with 100 internodes. For each relative node, probability was calculated based on the occurrence or not of a blind node, proleptic, or sylleptic shoot in each of the ten replicates annotated, consisting of ten different trees for each combination. This was undertaken to enable comparisons between different scion/rootstock combinations with highly variable internode numbers.
3. Results
3.1. Architectural Description of Two-Year-Old Branches
Rootstock influence on branch length (Length) was consistent across all cultivars (Table 1). Axillary shoots were longer when scions were grafted onto Garnem® rootstock than with the other rootstocks. ‘Lauranne’ and ‘Guara’ displayed an intermediate phenotype when grafted onto Rootpac® 40 and Rootpac® R, while combinations with Pilowred® and Rootpac® 20 tended to be shorter, although differences were only significant with ‘Guara’. When grafted onto Rootpac® 40, ‘Isabelona’ showed intermediate shoot length, whereas ‘Isabelona’ combinations with Rootpac® R presented a similar shoot length to those with Pilowred® or Rootpac® 20.

Table 1.
Analysis of architectural traits (shoot length, number and length of internodes) related to vigor of two-year-old branches of 15 scion/rootstock combinations of almond.
The effect of rootstock on the number of internodes (Nb_IN) per shoot presented a similar pattern to branch length. All cultivars had the highest number of internodes when grafted onto Garnem®, while displaying low Nb_IN when grafted onto Pilowred® (Table 1). Rootpac® series rootstocks presented small differences between each other in their influence of Nb_IN. The ‘Isabelona’/Rootpac® 40 combinations displayed a reduced number of internodes, while ‘Lauranne’, and significantly ‘Guara’, were higher when grafted onto Rootpac® 40. However, the two dwarfing rootstocks, Pilowred® and Rootpac® 20, showed a different influence on the number of internodes in combinations with ‘Isabelona’ and ‘Guara’, with both being less vigorous than ‘Lauranne’. In these, scions grafted onto ‘GN-8′ had less internodes than those grafted onto Rootpac® 20, pointing to a different mode of conferring a dwarfing response.
Since both Length and Nb_IN are influenced by rootstock, the mean length of the internodes (IN_L) was also influenced. While internode length of ‘Isabelona’ and ‘Guara’ were more affected by the rootstock, ‘Lauranne’ displayed less differences between combinations (Table 1). In the first two, combinations with Rootpac® 20 had the shortest internodes, whereas combinations with Pilowred®, Rootpac® 40 and Rootpac® R presented intermediate IN_L values. Combinations with Garnem® had the highest IN_L. Surprisingly, ‘Lauranne’ displayed the longest internodes when grafted onto Pilowred®, whereas when grafted onto ‘Garnem’, they had the shortest internodes of all combinations.
3.2. Number of Proleptic and Sylleptic Shoots on Two-Year-Old Branches
The occurrence of proleptic and sylleptic shoots has an important weight in determining tree architecture. Here, we reported the fate of each node in two-year-old branches, distinguishing between blind nodes (when no shoot was present), proleptic shoots (when the shoot was developed after a period of dormancy), and sylleptic shoots (when the shoot extended at the same time as the extension of the primary axis).
The number of proleptic shoots was influenced by rootstock, though this influence differed between cultivars. ‘Lauranne’ and ‘Guara’ displayed the highest number of proleptic shoots when grafted onto Garnem® (Table 2). In contrast, the ‘Isabelona’/Garnem® combination had a markedly reduced number of proleptic shoots. The effect of Pilowred® also varied depending on the grafted cultivar. ‘Guara’/Pilowred® had a reduced number of proleptic shoots, while ‘Isabelona’ and ‘Lauranne’ presented medium values when grafted onto Pilowred®. On the other hand, Rootpac® rootstocks showed a similar influence on all cultivars. Combinations with Rootpac® 20 had an elevated number of proleptic shoots, whereas those with Rootpac® 40 presented few proleptic shoots in general. Cultivars grafted onto Rootpac® R displayed an intermediate number of proleptic shoots.
Table 2.
Mean number of blind nodes, proleptic shoots, and sylleptic shoots on two-year-old branches of 15 scion/rootstock combinations of almond tree.
There was not much disparity among cultivars in the influence of rootstocks on the number of sylleptic shoots. In all cultivars, combinations with Garnem® and Rootpac® 40 showed the highest number of sylleptic shoots (Table 2). On the contrary, cultivars grafted onto Rootpac® 20 produced few or no sylleptic shoots. Combinations with Rootpac® R also displayed a reduced number of sylleptic shoots across all cultivars. Only with scions grafted onto Pilowred® were differences observed between cultivars. While ‘Isabelona’ and ‘Guara’ had few sylleptic shoots grafted onto this rootstock, ‘Lauranne’ presented a medium–high value.
3.3. Probability of Proleptic and Sylleptic Shoot Formation According to Nodal Position along the Branch
Proleptic shoots on all three scion cultivars were recorded on two-year-old wood, generally within the first 50 relative nodes, while sylleptic shoots were recorded on one- and two-year-old wood along the whole branch (Figure 1). Rootstock genotypes had a strong influence over branching probability, with little variation according to scion cultivar. Combinations with Rootpac® 20 rootstock had mostly proleptic shoots, with relatively few sylleptic shoots compared with scions on the other rootstocks. Scions grafted onto Rootpac® 40 had numerous sylleptic shoots located throughout the whole branch. Rootpac® R produced a scion with consistent branching all along the axis with proleptic shoots primarily at the lower nodal positions and sylleptic shoots at the higher nodal positions.
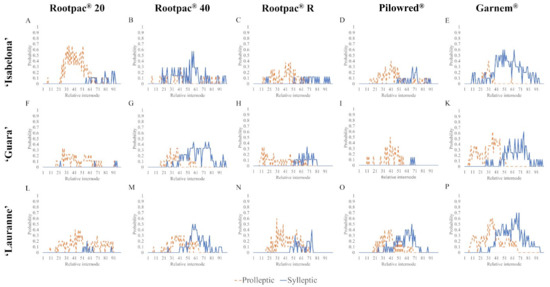
Figure 1.
Probability of proleptic and sylleptic shoot occurrence according to nodal position along two-year-old branches of 15 scion/rootstock combinations of almond trees. Location of sylleptic shoots was recorded on one- and two-year-old wood, while proleptic shoots were recorded only on two-year-old wood. The probabilities of proleptic and sylleptic shoot formation at each node were converted to their relative position and represented in a branch with 100 internodes. Internode 0 refers to the base of the branch and internode 100 refers to the apex.
Pilowred® presented more differences between cultivars than any other scion/rootstock combination. While each of the three scion cultivars had similar branching probabilities for proleptic shoots, mainly located in the first 50 relative internodes, ‘Guara’ trees on this rootstock produced almost no sylleptic shoots, ‘Isabelona’ had a few more, and ‘Lauranne’ had numerous sylleptic shoots across the whole branch, especially in the section between relative nodes 50 and 70. Scions grafted onto Garnem® rootstock produced numerous sylleptic shoots, similar to Rootpac® 40 rootstock, but considerably more than the other rootstocks. While all cultivars had similar distributions of sylleptic shoots, there were some scion cultivar-specific differences in the distribution of proleptic shoots. ‘Isabelona’ trees on Garnem® rootstock had few proleptic shoots in the first 30 relative nodes, while ‘Guara’ and ‘Lauranne’ presented significantly more proleptic shoots in the first 50 relative nodes and beyond.
4. Discussion
The dynamics of shoot development on two-year-old branches of young almond trees grafted on a range of rootstocks has previously been described [17]. The original study included thirty different combinations of five rootstocks and six scion cultivars. In this current study, we selected combinations of five rootstocks and three cultivars (‘Isabelona’, ‘Guara’ and ‘Lauranne’) with contrasting phenotypes for further analysis. Of particular interest was the pattern of sylleptic and proleptic axillary shoot production along the two-year-old branches.
As expected, vigor-conferring rootstocks such as Garnem® and Rootpac® 40 increased branch length. Branch growth is controlled by hormones, with gibberellic acid (GA), ethylene (ET) and brassinosteroids (BRs) being essential in its regulation [18,19,20,21]. Here, a differential hormonal activity must be exerted by the rootstock, explaining the variations between combinations. Nutrient uptake (nitrogen, in particular) and transport to the scion could also be an explanation for this rootstock effect on vigor and branch development [22,23,24]. In contrast, low vigor-conferring rootstocks Pilowred® and Rootpac® 20 showed a different effect. Trees grafted onto Pilowred® rootstock produced fewer nodes (Nb_IN) but with a length (IN_L) similar to scions grafted onto non-dwarfing rootstocks such as Garnem®. Combinations with Rootpac® 20 had a similar number of nodes as the others, but they presented a reduced mean internode length (13.1, 13.9 and 17.1 mm). These differences in internode development may be created by differences in the hormonal profile of the scion cultivar, which is influenced by rootstock. Internode elongation is mainly controlled by GA activity, although other hormones such as strigolactones (SL) or BRs may be involved [25,26,27,28]. This rootstock effect on two-year-old branches is quite different to our previous data on axillary shoot production along the trunk of the same trees. In this case, scion cultivars grafted onto Pilowred® displayed shorter internodes than other combinations [17]. While this result from our previous study may contradict the results presented in this current study, it reflects the elevated complexity of tree architecture regulation according to tree age and shoot position, which might be related to different hormonal gradients within the tree, or a different effect of the environment on this hormonal profile. Apple (Malus × domestica) trees show a similar response where rootstocks that confer a more dwarfing response influence shoot growth on young trees while the less dwarfing rootstocks do not affect shoot growth until much later [5,29].
Sylleptic shoot production was correlated with rootstock conferred vigor. Vigorous rootstocks such as Rootpac® 40 and Garnem®, promoted the formation of sylleptic shoots. On the other hand, proleptic shoots were present in high numbers in both combinations with the vigor-conferring Garnem® and with the dwarfing rootstock Rootpac® 20. This contradictory situation may be explained by a combination of apical dominance and vigor, affecting shoot development. Apical dominance is where the shoot apical meristem inhibits lateral bud outgrowth, favoring the development of the main axis [19,30]. Sylleptic shoots develop/extend at the same time as the main axis, generally forming lower down on the parent shoot and, therefore, leaving nodes near the shoot apex in a vegetative state. In the next year or season of growth, the shoot apex distances itself from these dormant vegetative buds, releasing them to form proleptic shoots, as we observed in the ‘Isabelona’/Rootpac® 20 or ‘Lauranne’/Rootpac® 20 combinations. In other cases, such as the ‘Isabelona’/Garnem® or ‘Guara’/Rootpac® 40 combinations, we observed a high number of sylleptic shoots and a reduced number of proleptic shoots. These phenotypes might be explained by a process dominated by apical control, where distal shoots inhibit the formation of new branches [19,31]. In these combinations, sylleptic shoots are produced all along the two-year-old branch, inhibiting the development of proleptic shoots in the next year of growth. Only those combinations that conjugated high vigor and weak apical dominance had strong sylleptic shoot formation in the first year of growth, a process also described in peach [32].
Gradziel [33] described that a desirable almond tree ideotype may present axillary shoots equally distributed along the axis. In peach trees, floral buds are developed in higher numbers on proleptic shoots than sylleptic shoots [16]. This suggests that a commercial ideotype should not produce too many sylleptic shoots as these might be at the expense of fruiting. To avoid this phenotype, combinations of low vigor-conferring rootstocks with weak apical dominance cultivars might be the best option. Although scions with strong apical dominance such as ‘Isabelona’ develop multiple proleptic shoots when grafted onto dwarfing rootstocks, these combinations typically develop few lateral branches and relocate resources to the main axis [17]. In contrast, combinations with weak apical dominance, such as ‘Lauranne’, with vigor-conferring rootstock, or Garnem® or Rootpac® 40, might naturally develop excessive sylleptic branching that would require detailed and expensive pruning to maintain high productivity in the orchard.
5. Conclusions
As with other aspects of almond tree architecture, proleptic and sylleptic shoot development is significantly influenced by rootstock genotype. Apical dominance and apical control have a crucial impact in differences observed between different scion/rootstock combinations. Although these processes are initiated in the shoots, they appear to be regulated by signals from the rootstock. Other processes related to vigor, such as hormonal activity or nutrient assimilation, could also be rootstock-mediated influences of bud fate. It is, therefore, important to study the biological processes occurring at the molecular level to better understand how the scion/rootstock interaction specifically effects the architecture of almond trees.
Author Contributions
Conceptualization, Á.M., J.G. and M.J.R.-C.; methodology, Á.M.; formal analysis, Á.M.; investigation, Á.M.; writing—original draft preparation, Á.M.; writing—review and editing, J.G. and M.J.R.-C.; visualization, Á.M.; supervision, J.G. and M.J.R.-C.; project administration, M.J.R.-C.; funding acquisition, M.J.R.-C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Research State Agency (AEI), grant numbers RTI-2018-094210-R-100 and FPI-INIA CPD2016-0056.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Balducci, F.; Capriotti, L.; Mazzoni, L.; Medori, I.; Albanesi, A.; Giovanni, B.; Giampieri, F.; Mezzetti, B.; Capocasa, F. The Rootstock Effects on Vigor, Production and Fruit Quality in Sweet Cherry (Prunus Avium L.). J. Berry Res. 2019, 9, 249–265. [Google Scholar] [CrossRef]
- Seleznyova, A.N.; Tustin, D.S.; Thorp, T.G. Apple Dwarfing Rootstocks and Interstocks Affect the Type of Growth Units Produced during the Annual Growth Cycle: Precocious Transition to Flowering Affects the Composition and Vigour of Annual Shoots. Ann. Bot. 2008, 101, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Tworkoski, T.; Fazio, G. Effects of Size-Controlling Apple Rootstocks on Growth, Abscisic Acid, and Hydraulic Conductivity of Scion of Different Vigor. Int. J. Fruit Sci. 2015, 15, 369–381. [Google Scholar] [CrossRef]
- Tworkoski, T.; Miller, S. Rootstock Effect on Growth of Apple Scions with Different Growth Habits. Sci. Hortic. 2007, 111, 335–343. [Google Scholar] [CrossRef]
- Van Hooijdonk, B.M.; Woolley, D.J.; Warrington, I.J.; Tustin, D.S. Initial Alteration of Scion Architecture by Dwarfing Apple Rootstocks May Involve Shoot-Root-Shoot Signalling by Auxin, Gibberellin, and Cytokinin. J. Hortic. Sci. Biotechnol. 2010, 85, 59–65. [Google Scholar] [CrossRef]
- Albacete, A.; Martínez-Andújar, C.; Martínez-Pérez, A.; Thompson, A.J.; Dodd, I.C.; Pérez-Alfocea, F. Unravelling Rootstock×scion Interactions to Improve Food Security. J. Exp. Bot. 2015, 66, 2211–2226. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.M.; Celton, J.M.; Chagne, D.; Tustin, D.S.; Gardiner, S.E. Two Quantitative Trait Loci, Dw1 and Dw2, Are Primarily Responsible for Rootstock-Induced Dwarfing in Apple. Hortic. Res. 2015, 2, 15001. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.C.; Alcaraz-López, C.; Muries, B.; Mota-Cadenas, C.; Carvajal, M. Physiological Aspects of Rootstock-Scion Interactions. Sci. Hortic. 2010, 127, 112–118. [Google Scholar] [CrossRef]
- Warschefsky, E.J.; Klein, L.L.; Frank, M.H.; Chitwood, D.H.; Londo, J.P.; Von Wettberg, E.J.B.; Miller, A.J. Rootstocks: Diversity, Domestication, and Impacts on Shoot Phenotypes. Trends Plant Sci. 2016, 21, 418–437. [Google Scholar] [CrossRef]
- Costes, E.; Lauri, P.E.; Regnard, J.L. Analizing Fruit Tree Architecture: Implications for Tree Management and Fruit Production. Hort. Rev. 2006, 32, 1–59. [Google Scholar] [CrossRef]
- Barthélémy, D.; Caraglio, Y. Plant Architecture: A Dynamic, Multilevel and Comprehensive Approach to Plant Form, Structure and Ontogeny. Ann. Bot. 2007, 99, 375–407. [Google Scholar] [CrossRef]
- Costes, E.; Crespel, L.; Denoyes, B.; Morel, P.; Demene, M.-N.; Lauri, P.-E.; Wenden, B. Bud Structure, Position and Fate Generate Various Branching Patterns along Shoots of Closely Related Rosaceae Species: A Review. Front. Plant Sci. 2014, 5, 666. [Google Scholar] [CrossRef]
- Negrón, C.; Contador, L.; Lampinen, B.D.; Metcalf, S.G.; Dejong, T.M.; Guédon, Y.; Costes, E. Systematic Analysis of Branching Patterns of Three Almond Cultivars with Different Tree Architectures. J. Am. Soc. Hortic. Sci. 2013, 138, 407–415. [Google Scholar] [CrossRef]
- Negrón, C.; Contador, L.; Lampinen, B.D.; Metcalf, S.G.; Guédon, Y.; Costes, E.; Dejong, T.M. Differences in Proleptic and Epicormic Shoot Structures in Relation to Water Deficit and Growth Rate in Almond Trees (Prunus Dulcis). Ann. Bot. 2014, 113, 545–554. [Google Scholar] [CrossRef]
- Negrón, C.; Contador, L.; Lampinen, B.D.; Metcalf, S.G.; Guédon, Y.; Costes, E.; Dejong, T.M. How Different Pruning Severities Alter Shoot Structure: A Modelling Approach in Young “Nonpareil” Almond Trees. Funct. Plant Biol. 2015, 42, 325–335. [Google Scholar] [CrossRef]
- Fyhrie, K.; Prats-Llinàs, M.T.; López, G.; DeJong, T.M. How Does Peach Fruit Set on Sylleptic Shoots Borne on Epicormics Compare with Fruit Set on Proleptic Shoots? Eur. J. Hortic. Sci. 2018, 83, 3–11. [Google Scholar] [CrossRef]
- Montesinos, Á.; Thorp, G.; Grimplet, J.; Rubio-Cabetas, M. Phenotyping Almond Orchards for Architectural Traits Influenced by Rootstock Choice. Horticulturae 2021, 7, 159. [Google Scholar] [CrossRef]
- Hollender, C.A.; Hadiarto, T.; Srinivasan, C.; Scorza, R.; Dardick, C. A Brachytic Dwarfism Trait (Dw) in Peach Trees Is Caused by a Nonsense Mutation within the Gibberellic Acid Receptor PpeGID1c. New Phytol. 2016, 210, 227–239. [Google Scholar] [CrossRef]
- Hollender, C.A.; Dardick, C. Molecular Basis of Angiosperm Tree Architecture. New Phytol. 2015, 206, 541–556. [Google Scholar] [CrossRef]
- Liu, J.; Moore, S.; Chen, C.; Lindsey, K. Crosstalk Complexities between Auxin, Cytokinin, and Ethylene in Arabidopsis Root Development: From Experiments to Systems Modeling, and Back Again. Mol. Plant 2017, 10, 1480–1496. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Xue, H.; Zhang, L.; Zhang, F.; Ou, C.; Wang, F.; Zhang, Z. Involvement of Auxin and Brassinosteroid in Dwarfism of Autotetraploid Apple (Malus × Domestica). Sci. Rep. 2016, 6, 26719. [Google Scholar] [CrossRef]
- Costa-Broseta, Á.; Castillo, M.; León, J. Nitrite Reductase 1 Is a Target of Nitric Oxide-Mediated Post-Translational Modifications and Controls Nitrogen Flux and Growth in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 7270. [Google Scholar] [CrossRef]
- Krouk, G.; Ruffel, S.; Gutiérrez, R.A.; Gojon, A.; Crawford, N.M.; Coruzzi, G.M.; Lacombe, B. A Framework Integrating Plant Growth with Hormones and Nutrients. Trends Plant Sci. 2011, 16, 178–182. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Cheng, Y.H.; Chen, K.E.; Tsay, Y.F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef]
- De Saint Germain, A.; Ligerot, Y.; Dun, E.A.; Pillot, J.-P.; Ross, J.J.; Beveridge, C.A.; Rameau, C. Strigolactones Stimulate Internode Elongation Independently of Gibberellins. Plant Physiol. 2013, 163, 1012–1025. [Google Scholar] [CrossRef]
- Depuydt, S.; Hardtke, C.S. Hormone Signalling Crosstalk in Plant Growth Regulation. Curr. Biol. 2011, 21, R365–R373. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Z.L.; Park, J.; Tyler, L.; Yusuke, J.; Qiu, K.; Nam, E.A.; Lumba, S.; Desveaux, D.; McCourt, P.; et al. The ERF11 Transcription Factor Promotes Internode Elongation by Activating Gibberellin Biosynthesis and Signaling. Plant Physiol. 2016, 171, 2760–2770. [Google Scholar] [CrossRef]
- Zhu, L.; Hu, J.; Zhu, K.; Fang, Y.; Gao, Z.; He, Y.; Zhang, G.; Guo, L.; Zeng, D.; Dong, G.; et al. Identification and Characterization of SHORTENED UPPERMOST INTERNODE 1, a Gene Negatively Regulating Uppermost Internode Elongation in Rice. Plant Mol. Biol. 2011, 77, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Van Hooijdonk, B.; Woolley, D.; Warrington, I.; Tustin, S. Rootstocks modify scion architecture, endogenous hormones, and root growth of newly grafted ‘Royal Gala’ apple trees. J. Am. Soc. Hortic. Sci. 2011, 136, 93–102. [Google Scholar] [CrossRef]
- Wang, B.; Smith, S.M.; Li, J. Genetic Regulation of Shoot Architecture. Annu. Rev. Plant Biol. 2018, 69, 437–468. [Google Scholar] [CrossRef] [PubMed]
- Hearn, D.J. Perennial Growth, Form and Architecture of Angiosperm Trees. In Comparative and Evolutionary Genomics of Angiosperm Trees; Plant Genetics and Genomics: Crops and Models; Groover, A., Cronk, Q., Eds.; Springer: Cham, Switzerland, 2016; Volume 21, pp. 179–204. [Google Scholar] [CrossRef]
- Génard, M.; Pagès, L.; Kervella, K. Relationship between sylleptic branching and components of parent shoot development in the peach tree. Ann. Bot. 1994, 74, 465–470. [Google Scholar] [CrossRef]
- Gradziel, T.M. The utilization of wild relatives of cultivated almond and peach in modifying tree architecture for crop improvement. Acta Hortic. 2012, 948, 271–278. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).