Abstract
Melatonin is a new plant hormone involved in multiple physiological functions in plants. Its roles in germination, photosynthesis, growth, flowering, fruiting, and senescence are noteworthy. Most studies have focused on its protective role in different stress situations, both biotic and abiotic. Preliminary data on its promoting role in almond seed germination and seedling growth are presented in this work. The germination-promoting capacity of melatonin was studied in different conditions of light and temperature, with important differences with respect to untreated seeds. The data confirmed that, at specific concentrations, melatonin was capable of stimulating germination, avoiding the stratification process, and therefore, shortening germination times. Furthermore, the melatonin-primed seedlings showed a higher root and aerial growth versus the untreated seeds when light was applied. The possible agronomic impact on the propagation of stone fruit trees was also discussed.
1. Introduction
Prunus dulcis, syn. P. amygdalus, Fam. Rosaceae, is an arboreous species native to the Mediterranean area that produces edible fruits named almonds. In the world, around 3.3 million tons of almonds are produced per year, the United States of America being the largest producer of almonds with around 2 million tons per year. Spain ranks second with an annual production of around 200 thousand tons [1].
In general, almond trees are propagated via seeds, which gives to the crop excellent biological diversity with different genotypes showing different stress tolerance responses, seed dormancy and germination rates, and also distinct flowering times [2,3]. Dormancy refers to a developmental block period in many mature seeds in which they are not able to germinate despite favorable conditions. Generally, a break or alleviation of dormancy periods can be achieved by incubating seeds in the cold, a process known as cold stratification. Almond seeds, as do other Prunus species, usually require a stratification process to break seed dormancy [4]. Normally, the seeds were treated in the cold (7 °C) for several weeks (1–10) and then put in a substrate of ideal germination conditions at about 22 °C and high humidity, requiring about 2–4 weeks to germinate. The stratification response is highly dependent on the almond variety, obtaining high percentages of germination after 6–8 weeks of cold [5].
Several strategies have been tested to improve germination with varying degrees of success. Thus, different stratification techniques and chemical treatments have reported some progress in improving germination mainly due to the great intraspecific variability in the response of the seeds [3,6]. Plant growth regulators have also been used to enhance the germination percentage of seeds, including abscisic acid (ABA), and gibberellins (GA), and also GA-inhibitors in peach [7], wild almond [8], and in almond [9] and aged almond seeds [10].
Melatonin (N-acetyl-5-methoxytryptamine), which was discovered in plants in 1995 [11,12,13,14], has been shown as a multifunctional molecule with regulating functions in many physiological processes such as germination, growth, rooting, photosynthesis, fruiting, ripening, and senescence, among others [15,16]. Melatonin has been shown to be a key new plant hormone in responses to biotic and abiotic stress, improving plant tolerance to adverse situations [17,18,19,20]. In germination, melatonin generally improves the process mainly in the presence of contaminants and/or stressors such as drought, salinity, UV radiation, etc., through the regulation of the redox network, gibberellin and abscisic acid levels in seeds [21,22,23,24,25,26]. Melatonin also seems to have a relevant role in chrono-regulated responses, interacting with circadian rhythms and their responses [27]. For example, we know that melatonin delays flowering in Arabidopsis thaliana through the regulation of gibberellin level and via stabilizing DELLA [28].
However, the role of melatonin as a possible substitute agent for the stratification process has not been studied. In this work, almond seeds were subjected to treatments with melatonin and different environmental conditions (temperature and photoperiod) to gauge their success in germination, avoiding the stratification process. The preliminary results show that melatonin stimulates the germination process in almond seeds, but also that light and temperature conditions were decisive factors. Thus, optimal treatments with melatonin were proposed as a strategy to improve the germination of almond seeds, and to enhance the production of seedlings for propagation without stratification.
2. Material and Methods
Prunus dulcis (Mill.) D.A.Webb; syn. P. amygdalus Batsch seeds var. Garrigues were obtained from almond trees of a farm of Cehegín, in the northwest of the Region of Murcia, Spain (38°06′14.6″ N, 1°47′22.9″ W). The seeds were collected in September/October 2021 and conserved in a silo; prior to the experiments (March 2022), seeds were classified according to their size and healthy appearance, discarding damaged or small seeds. The seeds (without exocarp) were hydrated for 24 h, and then endocarp (hull) was eliminated. The kernels, maintaining their seed coat (tegument), were primed in different melatonin solutions (20, 100 and 200 µM; and control in water) for 24 h, at 20 °C in the dark. So, four groups of seeds were made, which were distributed in the chambers A, B, C, and D (Table 1). For each seed group (36 seeds), the seeds were planted individually in little pots (∅ 8.5 cm × 11 cm) with vermiculite as inert substrate, incubated in their respective chambers and irrigated (only distilled water) to maintain high moisture in the substrate for 30 days. Seed germination was observed every five days and was defined as the emergence of the radicle. The experiments were repeated three times and the total germination rate (%) and other parameters were calculated.

Table 1.
Conditions of the different chambers to the germination.
After 30 days, germinated seeds were transferred to pots (15 cm × 15 cm × 30 cm) with organic substrate/perlite (3:1) and developed in chamber E (Table 1). In the case of seeds germinated in chamber A or B, the length of the stem and the number of leaves by plant were measured for 20 more days (until day 50 from the start of the assay). In this assay, there were no additional melatonin treatments, but the differences were due to initial priming of seeds with melatonin (20, 100 or 200 μM).
The graphical analysis of the data was made using SigmaPlot program version 14 (SYSTAT Software Inc., Richmond, CA, USA). Analysis of variance was performed using IBM SPSS Statistics 22.0 (IBM, New York, NY, USA). The statistical significance was considered for p-values less than 0.05 in ANOVA and a post hoc with Duncan Test.
3. Results and Discussion
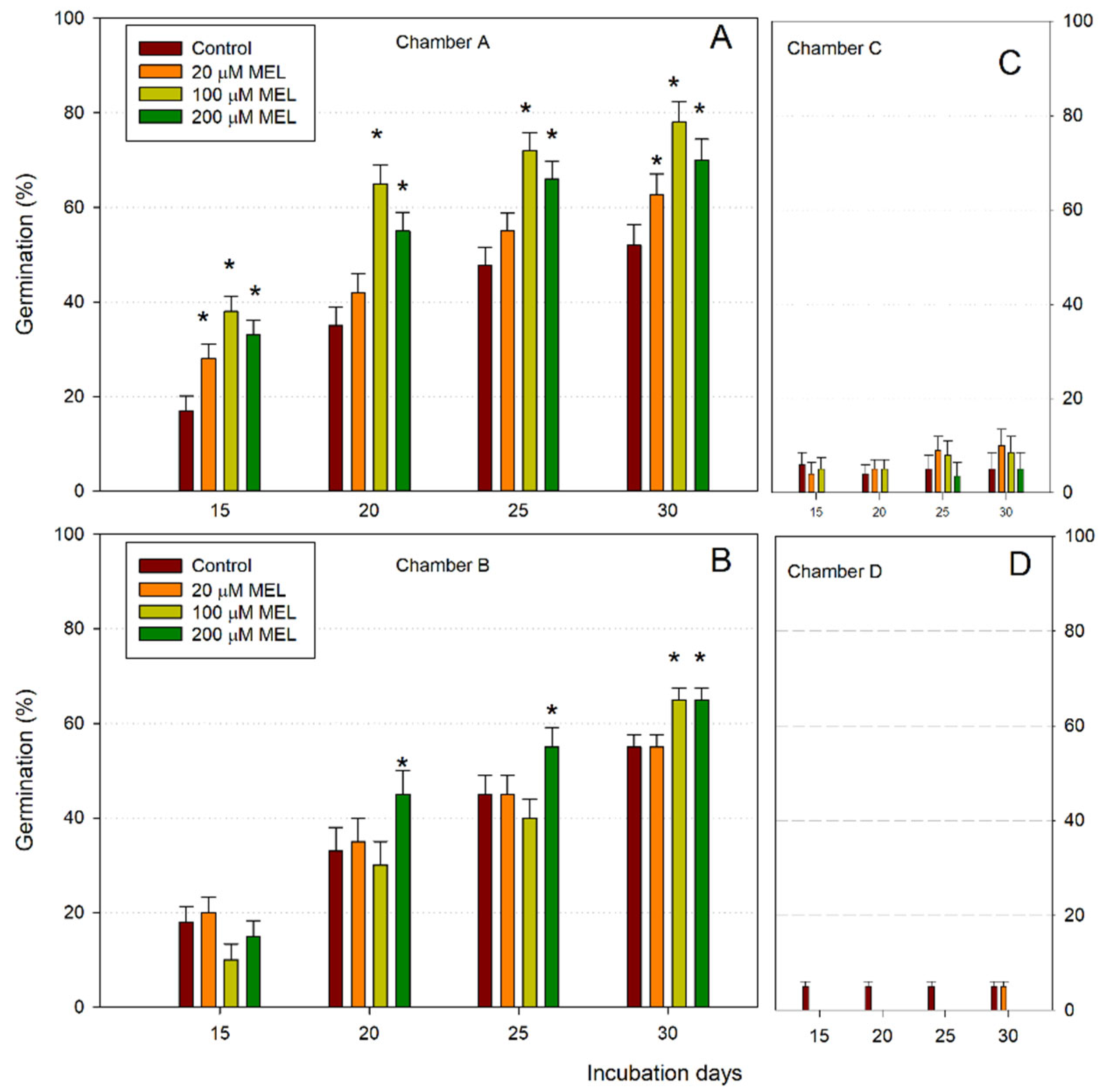
Figure 1 shows the germination response of unstratified almond seeds exposed to 10 °C and 22 °C in the conditions described in Table 1; the seeds were previously primed with different concentrations of melatonin. As can be seen, only the seeds in chambers A and B germinated successfully. Seeds exposed to 22 °C (chambers C and D) had a very low percentage of germination at 15 days, but also after 30 days, germination was less than 10%. Thus, cold conditions (A and B chambers) were clearly required for germination, in both light:dark (chamber A) and in the dark (chamber B) conditions. After 15 days at 10 °C, control (untreated) seeds showed germination rates of 17% at 10 °C in the 16:8 h photoperiod (chamber A) (Figure 1A) and 18% at 10 °C in the dark (chamber B) (Figure 1B). Under the 16:8h photoperiod at 10 °C (chamber A), the melatonin treatments improved germination after 15 days to about 21%, obtaining a maximum germination of about 80% after 30 days at 100 μM MEL (Figure 1A). After 15, 20, 25 and 30 days, melatonin clearly promoted germination with respect to the control of between 21 and 30%. In the dark at 10 °C (chamber B), a significant positive effect on germination could be observed after 20, 25 and 30 days, mainly at 200 µM MEL, but also at 100 µM MEL at 30 days, improving the germination percentage a maximum of 10% with respect to the control in the last days (Figure 1B). However, comparing the percentage of germination in chambers A and B (both at 10 °C), in the presence of melatonin, the photoperiod applied significantly improved the germination rate (Figure 1A), being more irregular in the dark (Figure 1B). The seeds incubated at 22 °C (chambers C and D) had hardly germinated after 30 days and the presence of melatonin did not improve the process (Figure 1C,D).
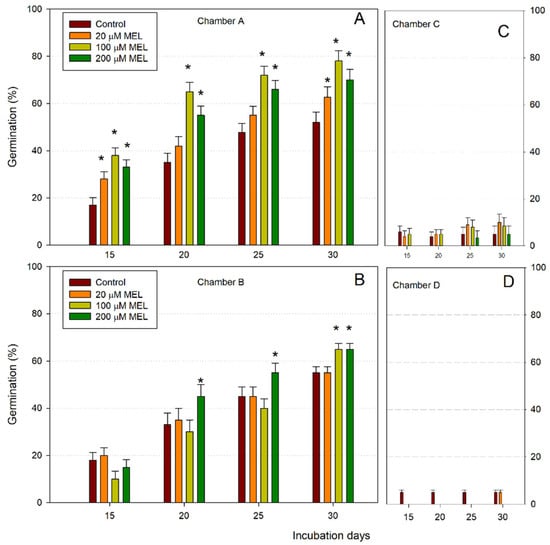
Figure 1.
Germination rates (%) of almond seeds primed with melatonin (MEL) at different concentrations after 30 days at 10 °C under 16:8h photoperiod (A) and in darkness (B), and at 22 °C under 16:8h photoperiod (C) and in darkness (D). Each bar represents the mean value (n = 36) ±SE obtained for each concentration used of three repeated experiments. Asterisks indicate significant differences with the control of that day based on a Duncan test with a significance level of p < 0.05.
Figure 2 shows some representative photographs of almond seeds germinated under the conditions of chamber A, where no anomalies or deformations in the radicle were observed, and similarly, seeds germinated in chamber B. In contrast, in chamber C (22 °C and photoperiod) there was hardly any germination (3–4 seeds of 36 in each assay), but those that germinated developed roots, stem and leaves with some deformations. In chamber D, the few seeds that germinated (1–2 seeds of 36 in each assay) hardly showed development under any of the conditions (data not shown).

Figure 2.
Representative photo of germinated almond seeds primed with different concentrations of melatonin (MEL) and incubated in chamber A after 30 days. Scale: each black or white mark represents 4 cm × 1 cm.
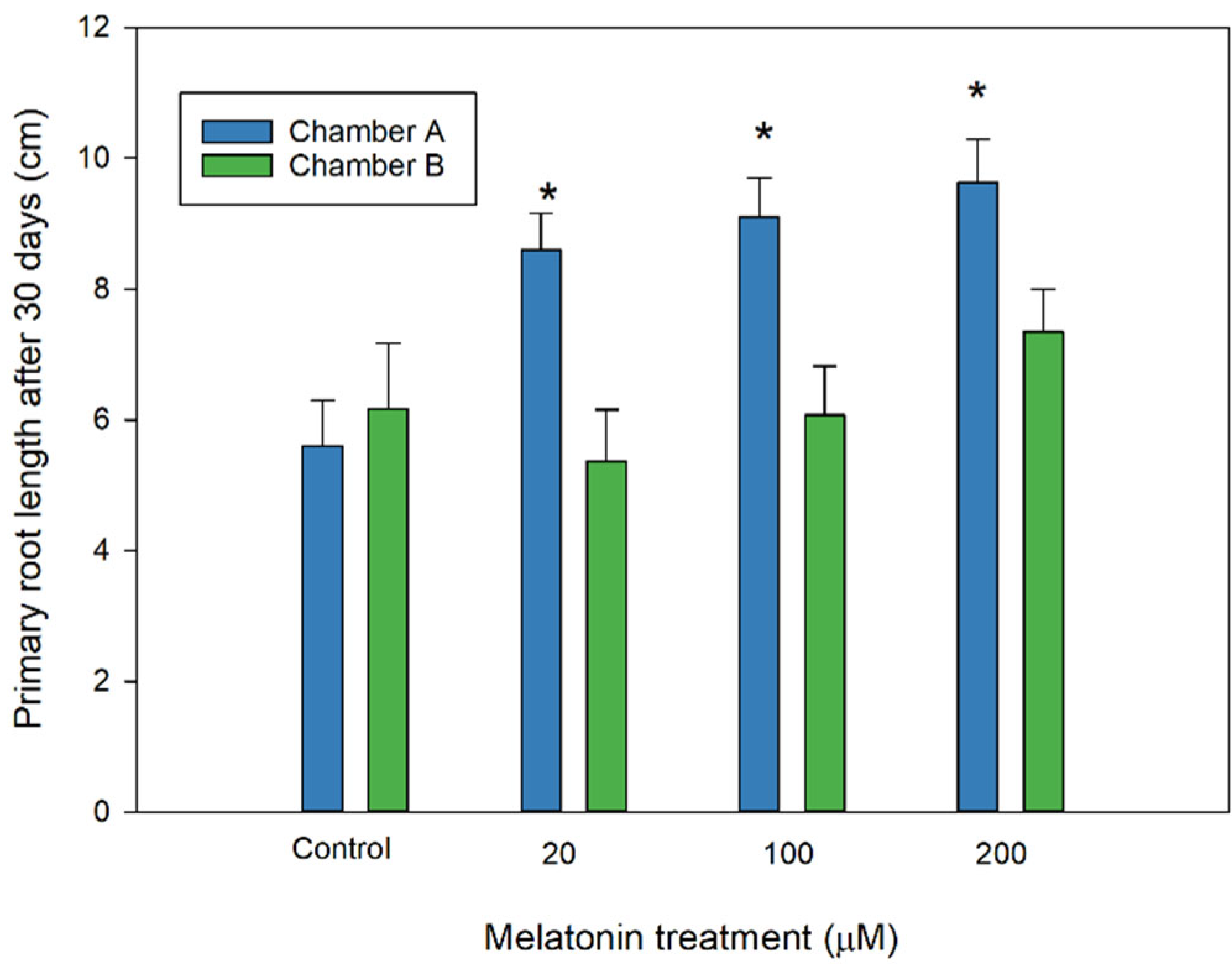
In addition, almond radicles from melatonin-treated seeds developed with a greater growth rate than those not treated, as can be seen in Figure 3, where the primary root length was promoted in melatonin-primed seeds, especially in the chamber at 10 °C with photoperiod (chamber A). In contrast, no improved effect was observed in chamber B.
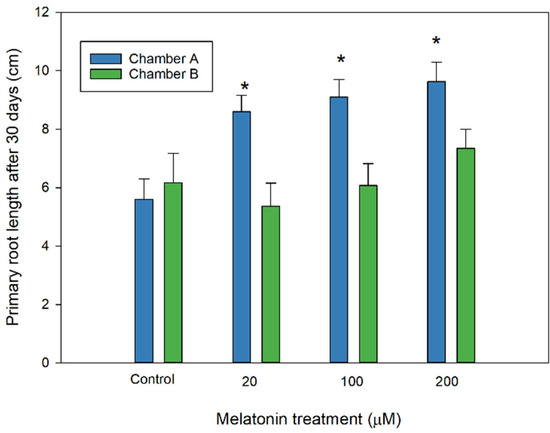
Figure 3.
Effect of melatonin in almond primary root length of primed seeds after 30 days in chamber A and B (see Table 1). Each bar represents the mean value (n = 36) ±SE obtained for each concentration used of three repeated experiments. Asterisks indicate significant differences with the control by each treatment based on a Duncan test with a significance level of p < 0.05.
After 30 days in their respective chambers (Table 1), the germinated seeds were transferred to pots with substrate:perlite, and into ideal growth conditions (24 °C and photoperiod, chamber E) to permit seedling development. Figure 4 shows photographs of seedlings from primed seeds with the different concentrations of melatonin, after 42 days of primed treatments.

Figure 4.
Representative photo of almond seedings 42 days old from primed seeds with different concentrations of melatonin (MEL), incubated in chamber A during germination (30 days) and then in chamber E (12 days). Scale: each black or white mark represents 4 cm × 1 cm.
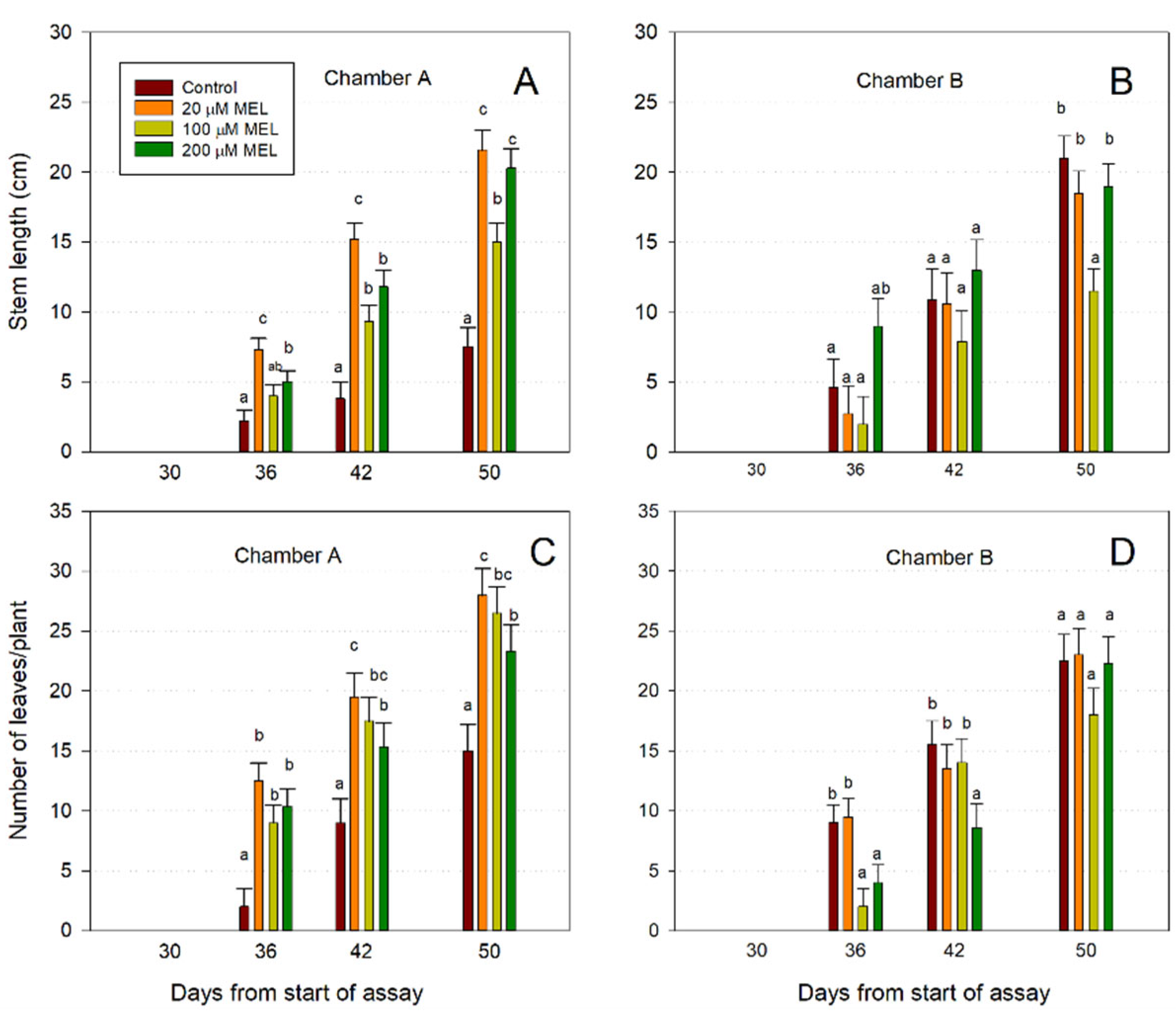
At the time of transfer (day 30), neither stem nor leaves existed on seedlings (Figure 5). After 50 days, the stems from the melatonin-primed seeds were up to 34.7% longer than control seedlings (Figure 5A). Concerning the number of leaves on seedlings, significant differences between seedlings from melatonin-primed seeds vs. control seedlings can be observed (Figure 5C). Thus, after 50 days, a differential of up to 53.5% appeared between control seedlings and those treated with 20 μM of melatonin in respect of the number of leaves by plant (Figure 5C). In respect of the germinated seeds in chamber B, the stem length of the melatonin-primed seeds did not show significant differences versus control, except in 100 μM melatonin at 50 days (Figure 5B); nor in the number of leaves per plant were there increases in the seeds primed with melatonin compared to control seeds, with even fewer leaves at 36 and 42 days in some treatments (Figure 5D). Therefore, while in the seeds germinated in chamber A (10 °C and photoperiod) a positive effect of melatonin was observed, this did not occur in the seeds germinated in chamber B (10 °C and dark), which indicates that photoperiod was necessary for the induction of growth by melatonin in terms of stem length (Figure 5A,B) and number of leaves (Figure 5C,D).
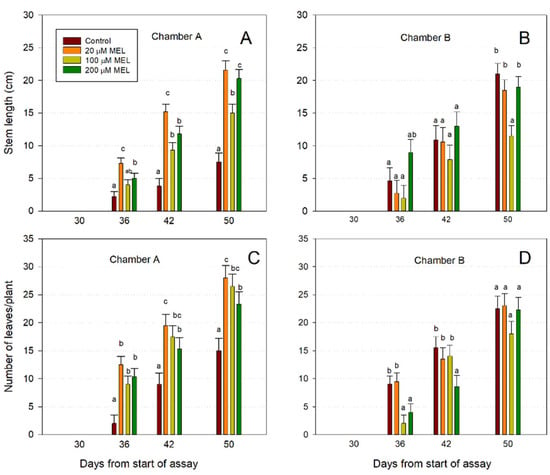
Figure 5.
Effect of melatonin on stem length (A,B) and number of leaves/plant (C,D) of primed almond seeds after 30 days in chambers A and B, and subsequent growth in chamber E. Each bar represents the mean value (n = 6) ±SE obtained for each concentration used of three repeated experiments. Superscript letters indicate significant differences with the control by each treatment based on a Duncan test with a significance level of p < 0.05.
In our assays without stratification period, melatonin-priming treatments saved time, in our case up to 4–5 weeks, in obtaining the same or higher percentages of germination than in the habitually stratified seeds [5]. In this way, germinated seeds with longer primary roots than control germinated seed can be transferred to habitual growth conditions (22–24 °C) much earlier than stratified ones, and with a better average success. In respect of the seedlings, melatonin-primed seeds presented a significant promoter effect on the length of the primary root and stem, and on the number of leaves by plants, a photoperiod treatment also being necessary to improve both the germination of the seeds and the seedling growth by melatonin.
4. Conclusions
These preliminary data showed that the stimulating effect of melatonin on germination in almond seeds was more strongly expressed during photoperiod exposure (chamber A) than in the dark (chamber B) at 10 °C. The seeds exposed to 22 °C barely germinated, with no melatonin having any promoting effect. The melatonin effect was manifested up to 50 days after priming the seeds, with a larger stem and a greater number of leaves in the seedlings from treated seeds than those untreated.
The data presented here lead us to study whether the positive effect of melatonin on the growth of seedlings was also reflected in the parameters of oxidative potential (redox network), that is, a greater tolerance to stress situations in the young seedlings, especially against drought, salinity and excess solar radiation, common stressors in almond seedbeds. More extensive work in different varieties of almond is also projected. These results could significantly change the strategies for obtaining seedlings from seeds that require stratification, such as in stone fruit trees and others, by avoiding the long periods of time and infrastructure required by stratification processes. In addition, it will be necessary to verify the ability of melatonin to avoid the stratification process of seeds in other plant species of agronomic interest.
Author Contributions
M.B.A. conceived and designed work. S.G.-S. performs the experiments. M.B.A., A.C. and J.H.-R. analyzed the data. M.B.A. wrote the manuscript. M.B.A. and S.G.-S. critically revised the final version of manuscript. M.B.A. and J.H.-R. read and approved the final article. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been funded through the project of the Ministry of Science and Innovation “R + D + I Projects”, State Program for the Generation of Knowledge and Scientific and Technological Strengthening of the R + D + I System and R + D + I Oriented to the Challenges of Society of the State Plan for Scientific and Technical Research and Innovation 2017–2020, Grant PID2020-113029RB-I00 funded by MCIN/AEI/10.13039/501100011033.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Thanks are due to the “Finca Parada Beltrán (Cehegín, Murcia) ”farm, and in especial to Antonio García Abril for providing almond seeds for our studies. More information in: https://www.um.es/en/web/phytohormones/, accessed on 17 August 2022 (Phytohormones & Plant Development Lab).
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Halstead, T. Tree Nuts: World Markets and Trade. USDA Foreign Agric. Serv. 2022. Available online: https://www.fas.usda.gov/data/tree-nuts-world-markets-and-trade (accessed on 17 August 2022).
- Raouf, A.; Salih, K.; Mohammed, A. Examination of Some Nut Traits and Release from Dormancy Along with Germination Capacity in Some Bitter Almond Genotypes. Tikrit J. Agric. Sci. 2022, 21, 25–32. [Google Scholar]
- García-Gusano, M.; Martínez-García, P.J.; Dicenta, F. Seed Germination Time as a Criterion for the Early Selection of Late-Flowering Almonds. Plant Breed. 2010, 129, 578–580. [Google Scholar] [CrossRef]
- Zigas, R.P.; Coombe, B.G. Seedling Development in Peach, Prunus persica (L.) Batsch. I. Effects of Testas and Temperature. Funct. Plant Biol. 1977, 4, 349–358. [Google Scholar] [CrossRef]
- García-Gusano, M.; Martínez-Gómez, P.; Dicenta, F. Breaking Seed Dormancy in Almond (Prunus dulcis (Mill.) D.A. Webb). Sci. Hort. 2004, 99, 363–370. [Google Scholar] [CrossRef]
- García-Gusano, M.; Martínez-Gómez, P.; Dicenta, F. Influence of Stratification, Heat and Removal of Teguments on Breaking of Seed Dormancy in Almond. In Proceedings of the XIII GREMPA Meeting on Almonds and Pistachios; Options Méditerranéennes: Série A. Séminaires Méditerranéens; n. 63; Oliveira, M.M., Cordeiro, V., Eds.; Centre International de Hautes Études Agronomiques Méditerranéennes (CIHEAM): Zaragoza, Spain, 2005; Volume 63, pp. 373–377. [Google Scholar]
- Zigas, R.P.; Coombe, B.G. Seedling Development in Peach, Prunus persica (L.) Batsch. II. Effects of Plant Growth Regulators and Their Possible Role. Funct. Plant Biol. 1977, 4, 359–369. [Google Scholar] [CrossRef]
- Rahemi, A.; Taghavi, T.; Fatahi, R.; Ebadi, A.; Hassani, D.; Chaparro, J.; Gradziel, T. Seed Germination and Seedling Establishment of Some Wild Almond Species. Afr. J. Biotech. 2011, 10, 7780–7786. [Google Scholar] [CrossRef]
- Koukourikou-Petridou, M. Paclobutrazol Affects Growth of Almond Fruits and Germination of Almond Seeds. Plant Growth Regul. 1996, 20, 267–269. [Google Scholar] [CrossRef]
- San, B.; Yildirim, A.N. Seed and in Vitro Embryo Germination in Aged Almond. Seed Sci. Technol. 2009, 37, 365–371. [Google Scholar] [CrossRef]
- Hattori, A.; Migitaka, H.; Iigo, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of Melatonin in Plants and Its Effects on Plasma Melatonin Levels and Binding to Melatonin Receptors in Vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in Edible Plants Identified by Radioimmunoassay and by HPLC-MS. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef]
- van Tassel, D.L.; Roberts, N.; O’Neill, S.D. Melatonin from Higher Plants: Isolation and Identification of N-Acetyl-5-Methoxytryptamine. Plant Physiol. 1995, 108, 101. [Google Scholar]
- Kolar, J.; Machackova, I.; Illnerova, H.; Prinsen, E.; van Dongen, W.; van Onckelen, H. Melatonin in Higher Plant Determined by Radioimmunoassay and Liquid Chromatography-Mass Spectrometry. Biol. Rhythm Res. 1995, 26, 406–409. [Google Scholar]
- Arnao, M.B.; Cano, A.; Hernández-Ruiz, J. Phytomelatonin: An Unexpected Molecule with Amazing Performances in Plants. J. Exp. Bot. 2022; in press. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Mora-Poblete, F.; Arnao, M.B.; Naz, S.; Anwar, M.; Altaf, M.M.; Shahid, S.; Shakoor, A.; et al. Phytomelatonin: An Overview of the Importance and Mediating Functions of Melatonin against Environmental Stresses. Physiol. Plant. 2021, 172, 820–846. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Regulatory Role of Melatonin in the Redox Network of Plants and Plant Hormone Relationship in Stress. In Hormones and Plant Response; Plant in Challenging Environments; Gupta, D.K., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 235–272. ISBN 978-3-030-77477-6. [Google Scholar]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin as a Regulatory Hub of Plant Hormone Levels and Action in Stress Situations. Plant Biol. 2021, 23, 7–19. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin Against Environmental Plant Stressors: A Review. Curr. Prot. Pept. Sci. 2022, 22, 413–429. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zhang, N.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin Promotes Seed Germination under High Salinity by Regulating Antioxidant Systems, ABA and GA4 Interaction in Cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Bai, Y.; Xiao, S.; Zhang, Z.; Zhang, Y.; Sun, H.; Zhang, K.; Wang, X.; Bai, Z.; Li, C.; Liu, L. Melatonin Improves the Germination Rate of Cotton Seeds under Drought Stress by Opening Pores in the Seed Coat. PeerJ. 2020, 8, e9450. [Google Scholar] [CrossRef]
- Xiao, S.; Liu, L.; Wang, H.; Li, D.; Bai, Z.; Zhang, Y.; Sun, H.; Zhang, K.; Li, C. Exogenous Melatonin Accelerates Seed Germination in Cotton (Gossypium hirsutum L.). PLoS ONE 2019, 14, e0216575. [Google Scholar] [CrossRef]
- Hu, W.; Zhang, J.; Yan, K.; Zhou, Z.; Zhao, W.; Zhang, X.; Pu, Y.; Yu, R. Beneficial Effects of Abscisic Acid and Melatonin in Overcoming Drought Stress in Cotton (Gossypium hirsutum L.). Physiol. Plant. 2021, 173, 2041–2054. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Pan, J.; Wang, H.; Reiter, R.; Li, X.; Zongmin, M.; Zhang, J.; Yao, Z.; Zhao, D.; Yu, D. Melatonin Inhibits Seed Germination by Crosstalk with Abscisic Acid, Gibberellin, and Auxin in Arabidopsis. J. Pineal Res. 2021, 70, e12736. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Bai, Y.-L.; Gong, C.; Song, W.; Wu, Y.; Ye, T.; Feng, Y.-Q. The Phytomelatonin Receptor PMTR1 Regulates Seed Development and Germination by Modulating Abscisic Acid Homeostasis in Arabidopsis thaliana. J. Pineal Res. 2022, 72, e12797. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wei, J.; Peng, Z.; Ma, W.; Yang, Q.; Song, Z.; Sun, W.; Yang, W.; Yuan, L.; Xu, X.; et al. Daily Rhythms of Phytomelatonin Signaling Modulate Diurnal Stomatal Closure via Regulating Reactive Oxygen Species Dynamics in Arabidopsis. J. Pineal Res. 2020, 68, e12640. [Google Scholar] [CrossRef]
- Shi, H.; Wei, Y.; Wang, Q.; Reiter, R.J.; He, C. Melatonin Mediates the Stabilization of DELLA Proteins to Repress the Floral Transition in Arabidopsis. J. Pineal Res. 2016, 60, 373–379. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).