Abstract
The fall armyworm of maize, Spodoptera frugiperda (J. E. Smith) (Lepidoptera; Noctuidae) is capable of causing a 100% yield loss due to its unforeseen occurrence from the seedling to the cob formation stage. To manage this serious pest, maize growers are tending to apply a high dosage of pesticides. This indiscriminate usage of pesticides has resulted in an unacceptable amount of insect resurgence in maize, harming maize production and consumption. In this review, we prepared a list of practical pest management options, including host plant resistance, agronomical, cultural, biological, botanical, chemical, and biotechnology approaches. It was found that cultivation of tolerant genotypes, adjusting sowing windows, and practicing specific intercultural and cropping systems measures in addition to chemical and non-chemical pest management strategies showed encouraging results for sustainable management of fall armyworm, which could protect the crop. This review highlights novel and successful management options advocated in various parts of the world. Recommendations documented in this paper would certainly pave the way for successful management of fall armyworm in maize and other concerned crops.
1. Introduction
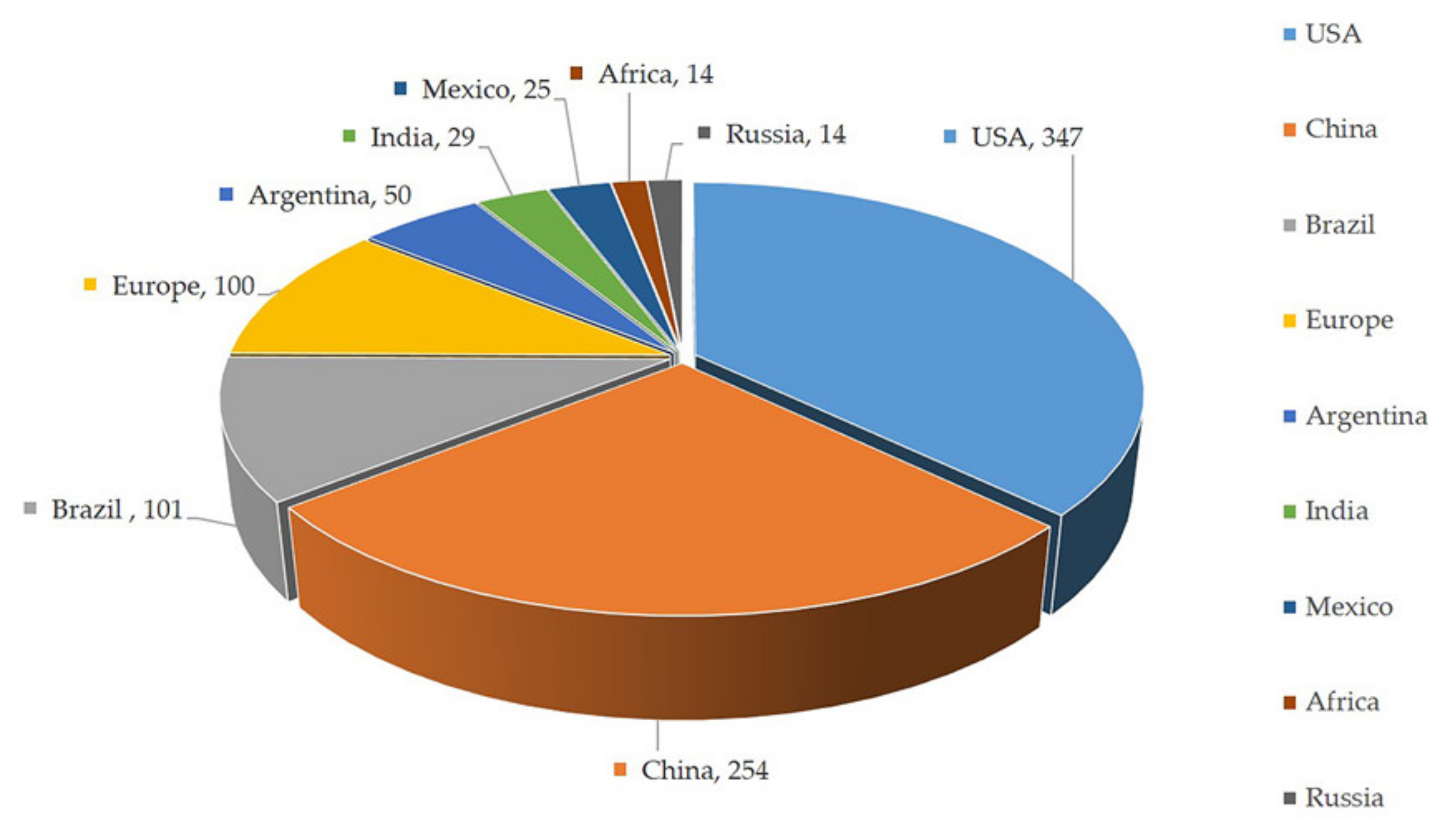
Maize (Zea mays L.) is an important cereal crop cultivated globally due to its versatile growth habit and higher productivity. It is gaining popularity among growers. Maize is grown on 193.7 million hectares worldwide, with a productivity of 5.75 tonnes per hectare. Currently, approximately 1147.7 million tonnes of maize kernels are being produced per year [1]. Maize is commercially grown in over 100 countries throughout the world, with the United States, Brazil, China, and India contributing significantly to production (Figure 1). Regardless of whether or not maize is a relatively new crop in many parts of the world, its unrivalled performance in both rainfed and irrigated ecosystems, as well as its ease of cultivation, entices growers to cultivate it. At present, maize has taken its place as the principal food crop next to rice and wheat due to its utilization in a wide variety of industrial production [2]. Importantly, maize is an indispensable source of raw material for the synthesis of corn oil, margarine, corn syrup, sweeteners, marmalade, and instant non-dairy coffee creamer besides its dedicated usage as animal and bird feed [3]. Corn is also used to make beverages, industrial chemicals, ethanol, fuel, plastics, and high-quality paper among other things [4,5]. Corn’s demand is always rising because of its many applications. To meet the ever-increasing demand, biotechnological discoveries paved the way for the discovery of elite cultivars with herbicide resistance, drought tolerance, high protein content, and many other features [6].
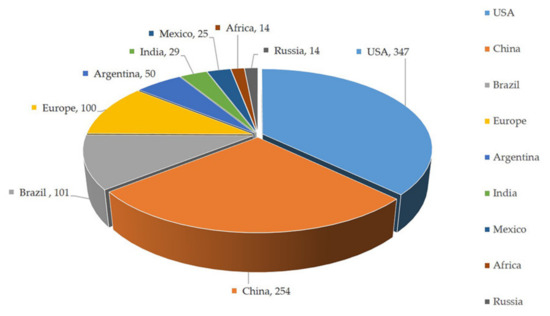
Figure 1.
Top ten maize-producing countries of the world. The number followed by country name indicates area in millions of hectares [Source: Food and Agriculture Organization, 2020] [1].
1.1. Distribution, Spread, and Host Plants
The fall armyworm (FAW), Spodoptera frugiperda is, is an invasive pest native to tropical and subtropical America that has expanded substantially across Western Africa and quickly into most Sub-Saharan African countries [7]. It has now spread to over 44 countries on the African continent [8,9]. The invasion began in India in 2018–2019, and within a year, it had expanded to most Asia-Pacific countries, including Korea, Japan, and Australia [10,11]. The fall armyworm’s massive migratory strength poses a serious threat to new habitats in Africa and Asia, as well as a considerable threat to crop plants’ economic viability [12]. Fall armyworm can harm over 350 plant species, including maize, sorghum, rice, sugarcane, cabbage, beet, groundnut, soybean, onion, cotton, pasture grasses, millets, tomato, potato, and cotton, to name a few [13]. Amongst the crops, maize is the most preferred one [14]. Fall armyworm is a gregarious pest that has both migratory and localised dispersal habits. It can move over 500 km before oviposition, just like any other Spodoptera moth [15]. Moths can travel up to 1600 km when the wind pattern is favourable [14]. In many parts of the world, it arrives seasonally, but in the event of alternate host availability and congenial weather, it can continue its generations throughout the year endemically and spread to adjacent areas. Fall armyworm is common in huge numbers, especially after a period of dry weather. The damaging stage of the pest is the larvae. The larvae have various feeding patterns; most of the time, they hide in the mouths of plants or the soil until sunset, when they emerge to feed. However, the day-feeding fall armyworm is active during the day as well. If fall armyworm is not controlled, maize yield losses could range from 8.3 to 20.6 million tonnes per year (21–53 percent of total production), according to Day et al. [16]. Similarly, earlier studies indicated severe yield loss, with 11.6% yield loss observed by Baudron et al. [17], 32–47% by Kumela et al. [18], and 22 and 67% by Day et al. [16].
Maize crops under irrigated ecosystems create congenial habitats that promote the survivability of insects, which usually build up and migrate to areas that are otherwise beyond their reach [15]. Furthermore, large-scale cultivation of other preferred hosts (sorghum, rice, sugarcane, cabbage, beet, groundnut, soybean, onion, cotton, pasture grasses, millets, tomato, potato, and cotton) in most maize-growing countries in Asia and Africa has exacerbated the general pest situation due to pest population shifts from one host to another (Figure 2). Therefore, over the years, the incidence of fall armyworm in maize has become a common phenomenon. As a result, developing management measures for this invasive insect pest of maize is critical to ensuring global food and nutritional security. Because of the frequency of fall armyworm and the significant potential losses it might cause, a variety of management approaches have been advocated around the world [18]. In the present review, efforts are being made to compile most of the sustainable management options available across the globe.
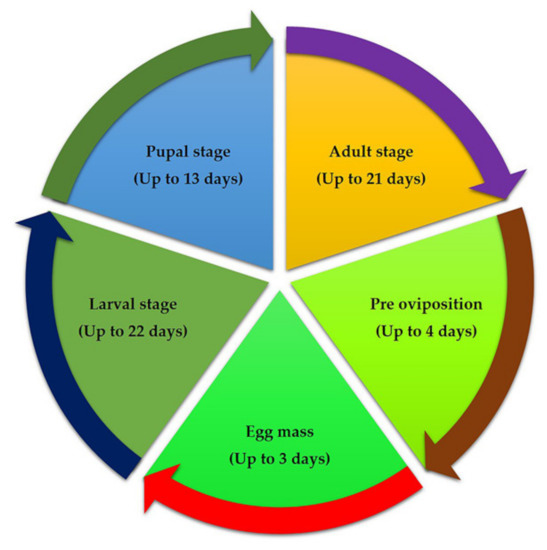
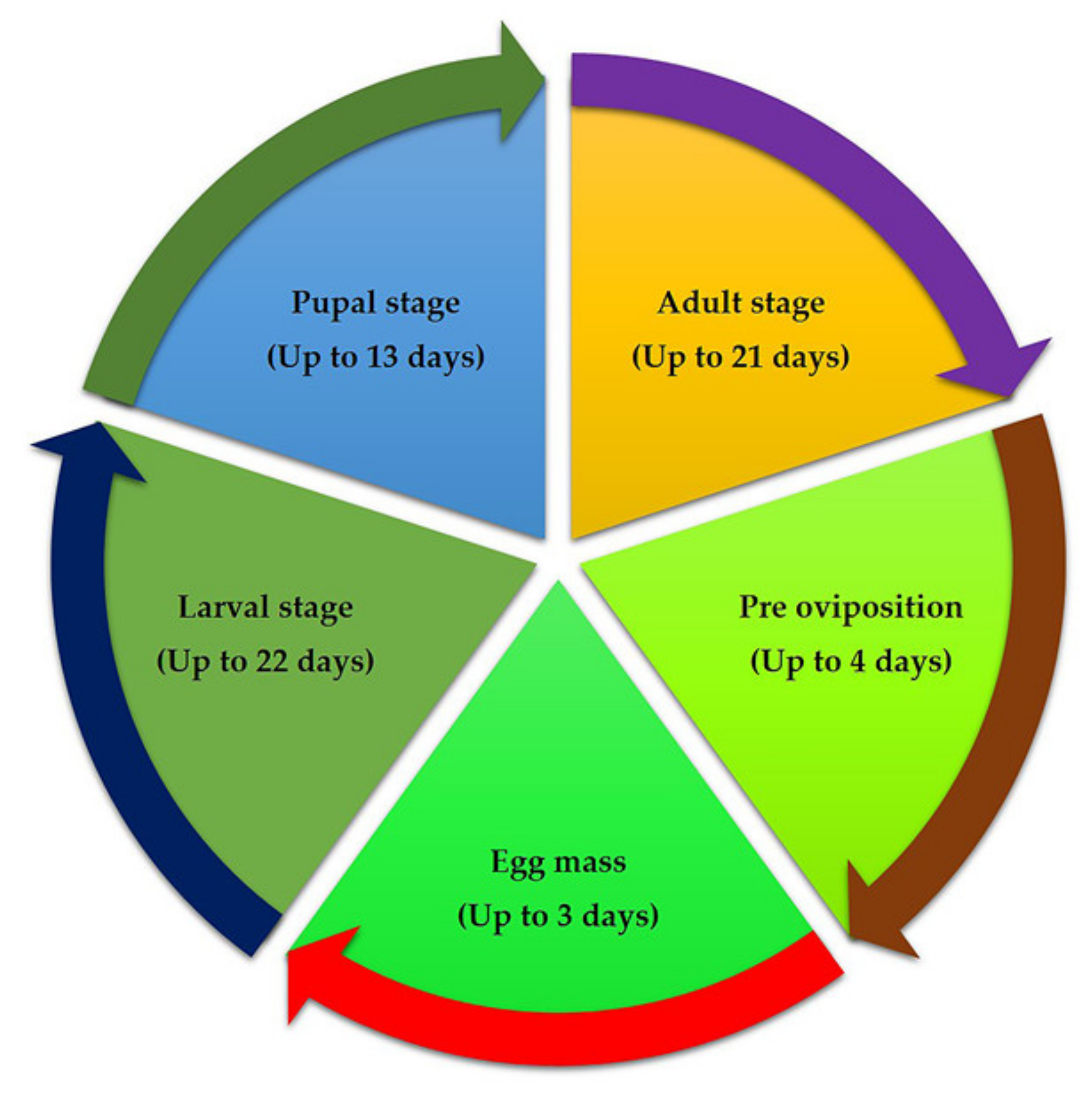
Figure 2.
Life cycle of fall armyworm of maize.
1.2. Biology
A thorough understanding of the life cycle of insect pests is necessary to devise sound management strategies. Fall armyworm’s life cycle varies depending on the environment in which it lives (Figure 2). In summer conditions in the USA, the insect completes its life cycle in around 30 days (at a daily temperature of ~28 °C), whereas in the spring and winter, it can take up to 60–90 days [10]. The minimum and maximum threshold for the development of FAW are 13.01 °C and 30 °C, respectively [19]. The number of generations of fall armyworm in a given area varies depending on the appearance of the dispersing adults and meteorological conditions [10,20]. Each female adult laid eggs in the group on either side of the leaf, and each female could lay about 1500–2000 eggs. Eggs are 0.3 to 0.4 mm in diameter and dome-shaped with an incubation period of 2–3 days. During the summer, six instars were observed; the immature larvae were greenish with black heads. Interestingly, Wu et al. (2021) [21] reported that invasive populations of S. frugiperda show bet-hedging strategy life history, enabling earlier reproduction and elongated reproductive lifespan, and that it promotes invasion success of the species. The mature larva has a brownish coloration and a reddish-brown head. A white inverted “Y”-shaped suture has been used to mark a fully mature larva. Pupation takes place in a cocoon in the soil at a depth of 2–8 cm, and the cocoon is usually oval in shape and measures 20–30 mm in diameter. The pupa is reddish-brown in colour and measures 14–18 mm in length and 4.5 mm in width, with an 8–9-day pupal phase. The adult has a grey and brown colour scheme and silvery-white hind wings with a small dark mark around the margins. The adult is nocturnal and only comes out in the evenings when the weather is hot and humid. The average adult lifespan ranges from seven to ten days [14].
1.3. Nature and Symptoms of Damage
The pest phase of the fall armyworm is the larvae. The immature larvae feed on the dorsal section of the leaf blade first, leaving the opposite epidermal layer unharmed. Larvae of the insect’s second or third instar bore holes in young leaves. As the larvae age, they start to graze from the edge of the leaf inward. In the initial phase of larvae, this affects the leaf in its whorl itself; during complete leaf expansion, the affected plants exhibit characteristic shot hole symptoms [22]. Though young larvae aggravate at the initial stages of their hatching, at later instars, larval densities usually reduce, and mostly one to two larvae can be seen per plant due to cannibalistic behaviour [23]. Larvae at later stages of their growth cause extensive defoliation [10]. In uncontrolled/unmanaged situations, larvae graze all the green leaves within a short period, leaving only leaf ribs and stalks that eventually look torn. The early whorl stage is the least sensitive, the mid-whorl stage is intermediate, and the late whorl stage is the most sensitive in terms of crop sensitivity. Mean densities of 0.2 to 0.8 larvae per plant during the late whorl stage can reduce production by 5 to 20%. At very high rates of infestation, larvae also move up to the ear and feed on kernels, causing complete yield loss.
The increased demand for maize kernels in the local, domestic, and international markets in many countries offered market assurance by proposing premium prices. Due to photo and thermal insensitivity coupled with a good market price and ease of production, maize has replaced several crops and cropping systems in most African and Asian countries. Of late, due to continuous monocropping and monoculture, maize has started to be affected by many pests. At present, maize is being affected by more than 25 different insect pests at various stages of crop growth. Fall armyworm—Spodoptera frugiperda, an invasive pest with a wide host range that causes severe yield loss across large areas of infection—is one of them [10,14,23,24,25,26].
2. Management Approaches
2.1. Agronomic Management
2.1.1. Sowing Window
Planting dates alter the growing environment of crop plants [27]. Pests are an integral part of the agro-ecosystem. Weather variables, such as temperature, relative humidity, rainfall, etc., have a significant impact on insect infestation and population growth. Insect pests’ survival, development, and reproductive capacity are all influenced by environmental variables [28]. Usually, fall armyworm appears late in the growing season. Hence, to avoid its infestation, late sowing in kharif crops should be avoided. Furthermore, seeding kharif maize should be limited to the earliest and narrowest sowing windows to disrupt the host range’s continuous availability because early harvest helps maize ears avoid the increased armyworm concentrations that emerge later in the season [29]. Furthermore, the study conducted by Teare et al. [30] also confirms the advantages of early sowing by stating that fall armyworm of maize was not a severe problem in early-planted maize. Kandel and Poudel [31] stated that fall armyworm is likely to affect the adoption of late-maturing hybrids and late-planted maize crops in Nepal. A recent study by Food and Agriculture Organizations (2018) [32] found considerable yield losses in maize due to the occurrence of fall armyworms in late-planted maize plots compared to nearby plots that were planted earlier. Therefore, early planting and adoption of short-duration genotypes are precise practices in endemic patches of fall armyworm in US corn belts [33]. According to Bhusal and Chapagain [34], planting maize earlier than the actual date will avoid the pest’s arrival time, allowing the crop to be protected or less afflicted. Based on the limited published work with regards to the influence of sowing windows on the pest reaction of fall armyworm in maize, it can be concluded that early sowing denies the entry and establishment of fall armyworm in the maize ecosystem so that possible yield loss can be avoided.
2.1.2. Tillage and Land Preparation
Tillage and soil management have a significant impact on the agro–pest ecosystem’s dynamics [35]. It is a fact that continuous and intensive land disturbance has deleterious impacts on soil by favouring erosion, organic matter depletion, and negative impacts on overall soil organisms [36]. Despite the fact that tillage has a number of negative effects on agroecosystem sustainability, it is the most widely used agro-technique in crop production worldwide to prepare fields for planting and manage pests [37]. This is especially true in the case of the management of the fall armyworm in maize. The grown larvae fall to the ground to pupate, making a reddish-brown oval cocoon that is about 2–3 cm long. In light soil, they can burrow 2–8 cm, whereas in hard soil, they can spin a webbed cocoon under leaf detritus. These pest propagules in turn contribute to life cycle completion and further multiplication. Kumar and Mihm [38] reported a 30–60% reduction in fall armyworm infestations under undisturbed soil conditions as compared to conventional tillage methods, which could be due to the availability of important predators. Clark [39] discovered that carabid beetles, rove beetles, spiders, and ants were key predators of fall armyworm and were found in greater numbers in reduced-tillage maize fields than in standard tillage fields. They also discovered that the farms with the most mulch coverage and the least tillage intensity had the highest predator numbers. Similar results of reduced fall armyworm infestation with zero tillage were also reported by Baudron et al. [17] and Roberts and All [40]. Rivers et al. [41] found considerably higher spider activity in no-till maize and with residue retention in wheat, as well as increased predation of fall armyworm following planting in maize, in a maize–wheat cycle conservation agriculture system in central Mexico. For example, research performed by Kumar et al. [38] found that zero–tillage consistently reduced fall armyworm damage and boosted yields by roughly 10% when compared to traditional tillage systems. According to preliminary findings from Zimbabwe, minimum tillage strategies reduce fall armyworm infestation rates in Africa’s maize belts [28].
Based on the findings of other studies, tillage may be a viable fall armyworm control strategy with minimal soil disturbance and residue retention that can be easily integrated with existing fall armyworm control efforts while also promoting sustainable intensification and climate change adaptation through conservation agriculture [42,43]. Furthermore, Harrison et al. [44] also reviewed that agroecological approaches to pest management, such as land tilling, bund planting, and soil fertility management, offer appropriate and low-cost pest control strategies that can be readily integrated into existing efforts of pest management strategies.
2.1.3. Nutrient Management
Balanced crop nutrition is a key factor in producing healthy plants and crops, and a healthy plant and vigorous crop growth usually resist biotic and abiotic stresses [27,44]. However, imbalanced crop nutrition paves the way for increased pest damage [45]. Morales et al. [46] reported that the introduction of inorganic fertilisers encouraged herbivorous pests in maize fields, including fall armyworm. A study conducted by Morales et al. [46] confirmed that crops fertilised with inorganic fertilizers had a higher incidence of aphids than organically fertilized crops. Inorganic sources of plant nutrients, such as chemical fertilizer amendments, assure the instantaneous supply of plant nutrients immediately after their application, especially with nitrogen. This spike in nitrogen content in the plant tissues results in attractiveness to insect herbivores and was evident from several studies conducted [45,46]. A higher incidence of fall armyworm in nitrogen-applied plots was also documented in the maize belts of Eastern Zimbabwe by Baudron et al. [17]. In a study conducted on Bermuda grass [47], it was found that nitrogen favours and potassium discourages the growth and development of fall armyworm. Maize is a nutrient-intensive crop that belongs to the C4 family. It always lures the grower to apply an elevated dosage of nitrogenous fertilizer, which imparts an imbalance in crop nutrition [48]. This could be the most pertinent factor that encourages the sporadic incidence of fall armyworm in maize over the years. Hence, care should be taken to assure balanced crop nutrition during integrated management of fall armyworm in particular and other biotic stress in general. Altieri and Nicholls [45] observed a lower abundance of insect herbivores and active soil biology in soils with high organic matter than in degraded soils with low organic carbon content. Therefore, it is imperative to amend the soil with organic manures, crop rotation with restorative field crops, green manuring, and inclusion of bio-inoculate-mediated crop nutrient modules to efficiently manage the fall armyworm.
2.1.4. Cropping System Measures
Growing crops in combination or succession is a vague practice in modern intensive agriculture. Cultivation of different crop species on the same piece of land imparts ecological diversity and prosperity. Bi-cropping is the practice of growing two crops with dissimilar growth habits on the same piece of land in a given period [49]. In addition to its numerous advantages over solitary cropping, bi-cropping practices in the traditional farming system give insurance against pests and unusual weather [48]. By increasing soil quality, stimulating vigorous plant development [50], restricting insect migration [51], obstructing egg-laying through visual or chemical disruption [52], and providing habitat for natural enemies, bi-cropping can lessen pest damage [53]. In research done by Altieri et al. [54], maize intercropped with beans reduced fall armyworm infestation by 23 percent as compared to solitary maize farming. Altieri [55] discovered that planting beans up to one month before maize reduced fall armyworm infestation significantly. Similarly, in another study in Nicaragua, maize intercropped with beans reduced fall armyworm infestations by 20–30% [51]. This reduction in pest load on maize under intercropped situations could be due to reduced larval dispersal [44] and is obvious because the diversified component crops trapped the ballooning instar larvae. Furthermore, the availability of natural enemies in agro-ecosystems is aided by the species diversity of crop plants in the same location, as evidenced by Van Huis [51], who observed higher densities of natural enemies, such as earwigs and spiders, feeding on early larval instars under maize polyculture systems. Maize growers in East Africa frequently intercrop maize with plants that emit pest-repellent volatiles, such as Desmodium spp. (Family: Fabaceae), as a measure of weed control, which is also found to be successful in the management of fall armyworm. In this system, the reduction in fall armyworm incidence was as much as 86%. As a result, it increased maize grain yields 2.7-fold in drought-prone areas of East Africa [53]. Similarly, bund-planted elephant grass (Pennisetum purpureum Schumach) (Family: Poaceae) attracts fall armyworm more than maize. This is also used in focused pesticide application [56]. In comparison to mono-cropped maize, intercropping maize with leguminous crops resulted in a considerable reduction in fall armyworm, notably during the early growth stages of the maize up to tasselling. Hence, in devising integrated management strategies against fall armyworm utilisation of plant species that produce semiochemicals may be incorporated into the cropping system. Furthermore, numerous research studies from around the world have found that leguminous intercrops are beneficial in controlling fall armyworm infestations in maize [53,57].
2.1.5. Trap Cropping
Trap crops are designed to attract pests away from cash crops, protecting them from assault [58]. The trap crop can be from the same family group as the main crop or from a separate one. Trap crops can be planted in two ways: perimeter trap cropping and row intercropping. The planting of trap crops around the perimeter of the primary cash crop is known as perimeter trap cropping (border trap cropping). It guards against pest attacks from all sides of the field. It is most effective against pests that are found along the farm’s perimeter. The trap crop is planted in alternating rows with the main crop in intercropped rows. Trap cropping has several pest management advantages. Because harm to main crops is reduced when trap crops effectively attract pest populations, major crops rarely require insecticide treatments [58]. Insect pests can be treated in a confined region rather than the entire field when they are highly concentrated in trap crops. Savings from reduced pest attack and insecticide use may far surpass the cost of sustaining non-profitable crops. The fall armyworm is a relatively recent pest in most of the world’s maize-growing regions. Information on trap cropping practice for the management of fall armyworm is scarce. On the other hand, Mooventhan et al. [59] advocate sowing 3–4 rows of Napier grass surrounding maize fields and spraying with 5% neem seed kernel extract or azadirachtin 1500 ppm as soon as the trap crop displays symptoms of fall armyworm damage. Furthermore, Gueraet al. [60] noticed more oviposited eggs on Brachiaria hybrid cv. Mulato II (Family: Poaceae), Panicum maximum cv. Mombasa (Family: Poaceae), and Panicum maximum cv. Tanzania (Family: Poaceae) than on maize. Trap crops could also be used in the “Push–Pull” cropping method, which involves intercropping pest-repellent (“push”) plant species (e.g., Desmodium spp.) with a pest-attractive trap (“pull”) plant species on the borders (e.g., napier grass (Pennisetum purpureum Schumach.) or Brachiaria spp.). Farmers in East Africa who used the Push–Pull technique completely saw an 86% reduction in fall armyworm infestation and crop loss as well as a 2.7-fold increase in production [53].
2.1.6. Pheromones Traps
Pheromones are scents produced by males or females that activate one or more behavioural reactions in the opposite sex, attracting males and females to mate. The use of pheromones primarily reduces insect populations in the given locality by way of lowering their reproduction rate because these chemicals confuse insects and get them trapped [61]. The female pheromone of S. frugiperda which attracts male moths consists of the major component (Z)-9-tetradecenyl acetate (Z9–14:Ac) and the minor component (Z)-7-dodecenyl acetate (Z7–12:Ac) [62]. The minor component (E)-7-dodecenyl acetate (E7–12:Ac) shows geographic variation and has so far only been found in females from Brazil. Regarding pest management, it is vital to remember that fall armyworm is made up of two strains (corn and rice) that have different pheromones [13]. In the case of fall armyworm, the female’s sex pheromone is commercially available in numerous countries [63], and pheromones have long been used to monitor the male population [64]. Monitoring with pheromone traps is useful because pest infestation varies from farm to farm and over time. Knowing when and where the adult pest is active and plentiful gives an early warning system that allows for field sampling and treatment. The grower was able to prevent wasteful pesticide applications or time-consuming samples by learning about the presence or absence of pests, and he was also given the warning to safeguard crops when moth flight was first identified [63]. Adult male moths are caught in traps, but larvae cause plant harm. As a result, we cannot simply count the number of moths in the traps while ignoring other elements, such as temperature, crop stage, and even natural control. Wind speed and temperature are favourably connected to trap captures, but relative humidity is adversely correlated. Monitoring fall armyworm adults with pheromone traps is the most effective way to determine the number of pesticide applications required to manage the pest in maize [63].
2.2. Biotechnological Approach
2.2.1. Plant Resistance
Host plant resistance is a low-cost and potentially effective method of insect pest control. It is generally inexpensive, durable, non-polluting, and locally adaptable, which helps in sustainable production. Fall-armyworm-resistant genotypes have been developed using a variety of plant morphological traits that contribute to antixenosis. Among them, a classic study by Sanches et al. [65] found the larval phase, metabolised food, and insect stool bulk as critical factors that contribute to fall armyworm resistance. From their study, maize genotypes BOZM 260, PA 091, and PARA 172 have emerged as promising sources of resistance to the fall armyworm. Similarly, Chen et al. [66] stated that maize accession Mp708 and FAW7050 were resistant to fall armyworm due to enhanced defence protein, greater amino acid and glucose content, and constitutive jasmonic acid accumulation. A study by Smith et al. [61] indicated that (E)-β-caryophyllene, a terpenoid released constitutively in a maize line, Mp708 is responsible for the demonstration of fall armyworm resistance. Furthermore, a study conducted by Ni et al. [67] also identified maize germplasm Mp708 and FAW7061 as highly resistant accession to fall armyworm infestation. Many fall-armyworm-resistant sources have been identified by researchers all over the world; these resistance sources can be used as possible parents in the development of resistant/tolerant varieties and hybrids in the future breeding programme to reduce seedling harm. Various academics and renowned research organizations, such as the Food and Agriculture Organization (FAO) and the International Maize and Wheat Improvement Center, have discovered some of the International Maize and Wheat Improvement Center (CIMMYT), have discovered some of the elite sources of maize resistant to fall armyworm which are summarized in Table 1.

Table 1.
Maize and sorghum germplasm showed resistance to fall armyworm.
2.2.2. Genetic Engineering
Employing synthetic chemical pesticides is a commonly observed phenomenon in normal cultivation practices. In chemical control, pests come into contact with the lethal substance and are killed. The chemicals are applied only when characteristic symptoms are noticed on the plant. This technique is not normally effective for fall armyworm management because caterpillars are usually found in corn whorls, where they are generally protected from insecticide treatments [71]. The fall armyworm, on the other hand, is resistant to over 30 active components of insecticides from all main classes on a global scale. Therefore, the utilization of genetically engineered maize that resists fall armyworm is a feasible method of pest management [72]. The efficacy of using genetically engineered crops in managing the fall armyworm was reported in China [73]. To control caterpillar pests, genetically engineered crop types expressing insecticidal crystalline (Cry) or vegetative insecticidal proteins (Vip) generated from Bacillus thuringiensis (Bt) that are selectively poisonous to distinct insect species are planted in many regions of the world, with adoption rates of over 80% [74]. Mass adoption, together with high control efficacy provided by genetic engineering, has led to area-wide population reductions in some pests that were already established [72]. Furthermore, the use of Bt corn hybrids is widely used to manage fall armyworm in America [71,72]. Growing Bt crops has helped to minimize pollution since they can reduce the use of chemical insecticides and assist natural pest management due to the narrow spectrum of activity of the deployed Cry and Vip proteins [74]. Genetically modified maize, according to Li et al. [73], should not be viewed as a single instrument for managing fall armyworm, but rather as a complement to other measures, such as biological control, cultural controls, and judicious use of chemical insecticides. Studies in this line have been meagre until now. This might be the one potential option for the management of fall armyworm which could be incorporated by taking into consideration only the biosafety and policy issues of respective maize-growing regions. A good insect resistance management plan, particularly for the fall armyworm, is critical and can be achieved through gene pyramiding approaches [73,75].
2.2.3. Gene Editing Approach (CRISPR-Cas System)
Even though transgenic (Bt) crops have provided significant crop protection benefits, the technology has been plagued by insect resistance, leading to the development of newer biotechnological approaches to insect pest management, such as gene editing (RNA interference (RNAi); gene drives, and, most recently, the CRISPR-Cas9 system) [76,77]. Gene editing technology can be used to validate gene activity, allowing researchers to better understand the resistance mechanism and design new pest management strategies. In 2013, CRISPR-Cas application in plants was successfully achieved in the lab [78].
Wu [79] revealed CRISPR/Cas9-mediated deletion of the abdominal-A homeotic gene in the fall armyworm, implying that the CRISPR/Cas9 technology is highly efficient in editing the fall armyworm genome based on his findings. For genome editing in the fall armyworm Spodoptera frugiperda, CRISPR/Cas9-mediated site-specific mutagenesis of three target genes—two marker genes (Biogenesis of lysosome-related organelles complex 1 subunit 2(BLOS2) and tryptophan 2, 3-dioxygenase(TO)), and a developmental gene, E93 (a key ecdysone-induced transcription factor that promotes adult development)—was performed. These mutational studies demonstrated there is a need to improve genome editing in lepidopteran and other non-model insects by using different approaches [80].
2.3. Chemical Management
Poison Baits
Poisonous baits are pesticides that can be mixed with some nontoxic food additives and have a lethal effect on targeted pests. In fall armyworm management, the application of poison bait in the whorls at vegetative and broadcasting in the mature crop has also shown a good response to controlling infestation [33]. In India, Patil et al. [58] elaborated on the protocol for the preparation of poison bait, wherein 5.0 kg of jaggery was mixed with 4–5 L of water. To this solution, 625.0 mL of monocrotophos 36 SL was added. This solution was further mixed with 50 kg of rice or wheat bran and packed in gunny or plastic bags and allowed to ferment for 48 h. Application of this fermented bait either through broadcasting or placing in maize whorls, preferably in the evening hours, significantly reduced fall armyworm incidence in maize.
Insecticides
The application of varieties of chemicals in the management of crop pests is an ambiguous practice [81,82,83]. Based on the mode of action, insecticides are divided into systemic and contact insecticides. The fall armyworm larva eats by remaining in the whorl of maize, avoiding contact with insecticides that have been administered. Therefore, several systemic insecticides were studied against the fall armyworm of maize. Among them, the application of emamectin benzoate 5 SG showed the highest acute toxicity, followed by chlorantraniliprole 18.5 SC and spinetoram 11.7 SC, whereas toxicities of flubendiamide 480 SC, indoxacarb 14.5 SC, lambda-cyhalothrin5 EC, and novaluron10 EC were at par according to the leaf-dip bioassay. However, at field evaluation application, chlorantraniliprole 18.5 SC, followed by emamectin benzoate 5 SG, spinetoram 11.7 SC, flubendiamide 480 SC, indoxacarb 14.5 SC, lambda-cyhalothrin 5 EC, and novaluron 10 EC were found better [10]. Profenophos + cypermethrin and spinosad were shown to be the most efficient in killing sixth-instar larvae in whorls in another investigation, followed by profenophos + lambda cyhalothrin and indoxacarb. In another study conducted in Ethiopia by Sisay et al. [84], synthetic insecticides, such as Lambda-cyhalothrin 5 EC, chlorantraniliprole 20 SC, Spinetoram 120 SC, Dimethoate 40 percent, Tracer 480 SC, and Ampligo 150 SC, significantly reduced fall armyworm larval mortality, reduced leaf damage, and increased biomass in maize when compared to an untreated control. Based on these recent studies, it could be inferred that the application of systemic insecticides appears to be the most promising component of integrated pest management plans for fall armyworm. At present, in countries such as India, which the pest recently invaded, since there are no recommended insecticides in place, the recommendation has been made by Central Insecticide Board and Registration Committee. The committee recommends the use of chlorantraniliprole 18.5 SC, thiamethoxam 12.6% + lambda-cyhalothrin 9.5% ZC, and spinetoram 11.7 SC for fall armyworm management. However, potential large-scale use of these chemical insecticides may cause hormesis effects, pest resurgence, and resistance development in target insect pests [85,86,87,88,89]. Additionally, these insecticides may have multiple negative impacts on human health and non-target organisms [90,91,92].
2.4. Biological Approaches
2.4.1. Biological Control
Biological control appears as a potential alternative to the chemical management of fall armyworm [93,94]. In various places, many natural enemies have been discovered to be related to this pest. Molina-Ochoa et al. [95] documented 150 species of parasitoids and parasites associated with fall armyworm from the Americas and the Caribbean basin. In India, Shylesha et al. [20] also reported egg, larval, and larval–pupal parasitoids and predators attacking different stages of this pest on maize. Among the predators, various ground beetles (Coleoptera: Carabidae); the striped earwig, Labidura riparia (Pallas) (Dermaptera: Labiduridae); the spined soldier bug, Podisus maculiventris (Say) (Hemiptera: Pentatomidae); and the insidious flower bug, Orius insidiosus (Say) (Hemiptera: Anthocoridae) were found to be effective against fall armyworm of maize as a means of biocontrol [96]. While predators have a significant impact on fall armyworm survival and development, parasitoids, which are more effective in causing mortality in fall armyworm populations, significantly outnumber predators. Solitary parasitoids of the Hymenoptera genera Chelonus and Campoletis were recovered from S. frugiperda larvae [97] and Trichogramma parasitoids could be high potential biocontrol agents for developing inundative biological control programs [98,99,100]. In Africa, five species of parasitoids were recorded from the fall armyworm in three East African countries in 2017 [84]. The Campoletis species is a larval endoparasitoid that plays an important role in regulating S. frugiperda [95]. The fall armyworm is a natural host of the parasitoid C. flavicincta, the larvae of the parasitoid. The larvae of S. frugiperda starts making its cocoon, which allows it to continue its development until its adult emergence. At this juncture, prime emphasis must be given to identifying the potential natural enemies and disease-causing pathogens for biological control of S. frugiperda. Biocontrol and biopesticides approaches are eco-friendly, sustainable, and appropriate alternatives to chemical insecticides [101]. These approaches form a strong base, and they are the key component of any integrated pest management program (IPM). Fall armyworm can effectively control insect populations by introducing naturally occurring disease-causing infections as natural regulatory agents [102]. Several microbial infections have been investigated in hopes of controlling fall armyworm numbers. More than 53 parasitic species, comprising 43 genera and 10 families, have been discovered as attacking fall armyworm [103]. Although no commercial Bt isolates have been created to combat fall armyworm, the Cry1F protein is thought to be more toxic to fall armyworm than any other Cry protein [104]. Likewise, many predators that attack fall armyworm eggs and larvae were also reported.
Insect pathogenic viruses are also one of the important biocontrol agents of FAW. There are different viruses known to infect FAW larvae, such as Ascoviruses, Baculoviruses, Densoviruses, Rhabdoviruses, and Partiti-like viruses [105]. Among these, Baculoviruses are more promising. The marketability of virus products for the management of various insect pests around the world has improved as a result of recent technological advancements. Spodoptera frugiperda multiple nucleopolyhedrovirus (SfMNPV), a baculovirus infecting S. frugiperda, has become commercially and registered in several countries for FAW management [106,107].
Understanding the ecological and toxicological connection between genetically modified cultivars (GMCs) and biological control agents is critical for GM cultivar compatibility and integrated pest resistance management strategies. To support this, Souza et al. [108] reported the efficiency of biological control for fall armyworm resistance to the protein Cry1F. They studied the search behaviour and predatory capabilities of Oriusinsidiosus (Say) (Hemiptera: Anthocoridae) and Doru luteipes (Scudder) (Dermaptera: Forficulidae) on fall armyworm eggs and caterpillars resistant to the expressed protein Cry1F [108].
2.4.2. Bird Perches
A diverse group of insectivorous birds has been found to feed on crop insect pests [109]. These insectivorous birds have been known to reduce the larval population by as much as 84% [110]. According to Jones et al. [111], many birds occurring in cropped fields actively forage for and consume caterpillars in crop vegetation. Black drongo, house sparrows, blue jays, cattle egret, rosy pastor, and mynah are frequent insectivorous birds that feed on a large number of lepidopteran insects among predatory birds. Insectivore birds show a strong tendency of attracting large size larvae [111]. In the case of environmentally friendly pest management, fall armyworm larvae that have escaped parasitism grow faster than those parasitized by Euplectrus wasps [111]. Boundary trees (such as fodder, fuelwood, and shelter trees) provide perches and roosts for birds and bats and also increase the structural variety of the farm ecosystem by providing shade and shelter [44]. These bird species are capable of extracting fall armyworm larvae from maize plant whorls and husks. Based on these results, it was known that insectivorous birds are also a potential tool for the ecofriendly and sustainable management of fall armyworm of maize. Under field conditions, insectivorous birds normally attract large larvae, although red-winged blackbirds were equally likely to eat parasitized and non-parasitized fall armyworm prey of the same body size [111]. Maize is a non-branching crop plant, which discourages birds from sitting and staying for extended periods. Therefore, suitable live bird perches in the maize ecosystem should be established. In order to facilitate bird visits, several fast-growing plants that provide rigid support for perching insectivorous birds from the vegetative stage until crop maturity should be grown in the maize field.
2.4.3. Use of Botanicals
In nature, many plants exhibit insecticidal properties. Preparations made out of those plant products are called botanical insecticides. These are insect poisons that are found in nature and are extracted or generated from plants or minerals. Increased interest in the eco-friendly management of pests in most edible crops led to exploring the use of botanicals in pest management. Botanicals employed for the management of pests are safe for humans and animals with reduced environmental impact. Fall armyworm tends to become highly destructive in maize cultivation; synthetic chemicals used in maize production greatly impair the quality of maize kernels and fodder values. Therefore, the utilization of botanicals is reported to be a potential strategy in pest management [112]. Similarly, many studies were conducted in various parts of the world reported that botanicals found to be an alternative and potential strategy in fall armyworm management [113]. The greatest strength of botanical extracts is their specificity, as most are essentially nontoxic and non-pathogenic to animals and humans [114]. Various plant species have shown insecticidal properties against fall armyworm; for example, extracts of neem, Azadirachta indica (Family: Meliaceae), Argemone ochroleuca Sweet (Family: Papaveraceae) [115], Boldo, Peumusboldus Molina (Family: Monimiaceae), Jabuticaba, and Myrciaria cauliflora (Mart.) O. Berg (Family: Myrtaceae).
There is sufficient evidence, as shown by the research findings, showing there are several prospects for the use of botanical extracts in the management of fall armyworm. However, due to several operational constraints, utilization of these potentials is limited [116,117].
3. Conclusions
Fall armyworm of maize is a destructive insect pest intensively affecting most of the maize-growing tract of world. It has invaded the United States and all maize-producing countries of Africa and Asia. Isolated efforts of its management could not accomplish the intended pest management due to the pest’s high fecundity rates and fast-spreading nature. Therefore, integration of all possible tools and techniques of pest management is necessary to achieve satisfactory pest management in maize. In this review paper, most of the classical work performed in the field of fall armyworm management was brought together and discussed. Based on the work published, it was observed that the life cycle of fall armyworm varies with environmental factors. Importantly, the life cycle is prolonged by a decrease in temperature, and with the availability of a congenial environment, the insect completes 4–6 generations per annum. With respect to the management of field conditions, early planting in kharif has always allowed the crop to avoid higher rates of infestation. Maize cultivars developed by using resistant lines, such as Mp708, FAW7050, Mp708, and FAW7061, were found promising in withstanding outbreaks of fall armyworm. Besides these, practicing seed treatment, intercultural operations, cropping systems, cultural, chemical, non-chemical, biological, and biotechnological approaches to pest management proved to be excellent in the sustainable management of FAW with very high scientific relevance.
Author Contributions
Conceptualization and literature search, R.M.K., Y.M., N.N. and V.P.; writing—reviewing and editing, R.M.K., Y.M., B.-G.G., V.P., P.K. and F.U. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analysed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
Authors declare no potential conflict of interest.
References
- FAOSTAT. Statistics; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Andorf, C.; Beavis, W.D.; Hufford, M.; Smith, S.; Suza, W.P.; Wang, K.; Woodhouse, M.; Yu, J.; Lübberstedt, T. Technological advances in maize breeding: Past, present and future. Theor. Appl. Genet. 2019, 132, 817–849. [Google Scholar] [CrossRef] [PubMed]
- Kaul, J.; Jain, K.; Olakh, D. An overview on role of yellow maize in food, feed and nutrition security. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 3037–3048. [Google Scholar] [CrossRef]
- Naz, S.; Fatima, Z.; Iqbal, P.; Khan, A.; Zakir, I.; Noreen, S.; Younis, H.; Abbas, G.; Ahmad, S. Agronomic crops: Types and uses. In Agronomic Crops; Springer: Singapore, 2019; pp. 1–18. [Google Scholar]
- Gamage, A.; Liyanapathiranage, A.; Manamperi, A.; Gunathilake, C.; Mani, S.; Merah, O.; Madhujith, T. Applications of Starch Biopolymers for a Sustainable Modern Agriculture. Sustainability 2022, 14, 6085. [Google Scholar] [CrossRef]
- Malenica, N.; Dunić, J.A.; Vukadinović, L.; Cesar, V.; Šimić, D. Genetic approaches to enhance multiple stress tolerance in maize. Genes 2021, 12, 1760. [Google Scholar] [CrossRef] [PubMed]
- Brévault, T.; Ndiaye, A.; Badiane, D.; Bal, A.B.; Sembène, M.; Silvie, P.; Haran, J. First records of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), in Senegal. Entomol. Gen. 2018, 37, 129–142. [Google Scholar] [CrossRef]
- Bueno, R.C.O.d.F.; Carneiro, T.R.; Bueno, A.d.F.; Pratissoli, D.; Fernandes, O.A.; Vieira, S.S. Parasitism capacity of Telenomus remus Nixon (Hymenoptera: Scelionidae) on Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) eggs. Braz. Arch. Biol. Technol. 2010, 53, 133–139. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Goergen, G.; Tounou, K.A.; Agboka, K.; Koffi, D.; Meagher, R.L. Analysis of strain distribution, migratory potential, and invasion history of fall armyworm populations in northern Sub-Saharan Africa. Sci. Rep. 2018, 8, 2710. [Google Scholar] [CrossRef]
- Pogue, M.G. A World Revision of the Genus Spodoptera Guenée: (Lepidoptera: Noctuidae); American Entomological Society: Philadelphia, PA, USA, 2002; Volume 43. [Google Scholar]
- Prasanna, B.; Huesing, J.; Eddy, R.; Peschke, V. Fall Armyworm in Africa: A Guide for Integrated Pest Management; USAID; CIMMYT: Mexico City, Mexico, 2018. [Google Scholar]
- Deshmukh, S.; Pavithra, H.; Kalleshwaraswamy, C.; Shivanna, B.; Maruthi, M.; Mota-Sanchez, D. Field efficacy of insecticides for management of invasive fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) on maize in India. Fla. Entomol. 2020, 103, 221–227. [Google Scholar] [CrossRef]
- Kenis, M.; Benelli, G.; Biondi, A.; Calatayud, P.; Day, R.; Desneux, N.; Harrison, R.; Kriticos, D.; Rwomushana, I.; van den Berg, J. Invasiveness, biology, ecology, and management of the fall armyworm, Spodoptera frugiperda. Entomol. Gen. 2022. [Google Scholar]
- Cock, M.J.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef]
- Navik, O.; Shylesha, A.; Patil, J.; Venkatesan, T.; Lalitha, Y.; Ashika, T. Damage, distribution and natural enemies of invasive fall armyworm Spodoptera frugiperda (JE smith) under rainfed maize in Karnataka, India. Crop Prot. 2021, 143, 105536. [Google Scholar] [CrossRef]
- Hafeez, M.; Li, X.; Ullah, F.; Zhang, Z.; Zhang, J.; Huang, J.; Khan, M.M.; Chen, L.; Ren, X.; Zhou, S. Behavioral and physiological plasticity provides insights into molecular based adaptation mechanism to strain shift in Spodoptera frugiperda. Int. J. Mol. Sci. 2021, 22, 10284. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; He, P.-C.; Hu, L.; Chi, X.-L.; Keller, M.A.; Chu, D. Host selection and adaptation of the invasive pest Spodoptera frugiperda to indica and japonica rice cultivars. Entomol. Gen. 2022, 42, 403–411. [Google Scholar] [CrossRef]
- Westbrook, J.; Fleischer, S.; Jairam, S.; Meagher, R.; Nagoshi, R. Multigenerational migration of fall armyworm, a pest insect. Ecosphere 2019, 10, e02919. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Baudron, F.; Zaman-Allah, M.A.; Chaipa, I.; Chari, N.; Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera frugiperda J.E. Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in Eastern Zimbabwe. Crop Prot. 2019, 120, 141–150. [Google Scholar] [CrossRef]
- Kumela, T.; Simiyu, J.; Sisay, B.; Likhayo, P.; Mendesil, E.; Gohole, L.; Tefera, T. Farmers’ knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya. Int. J. Pest Manag. 2019, 65, 1–9. [Google Scholar] [CrossRef]
- Du Plessis, H.; Schlemmer, M.-L.; van den Berg, J. The effect of temperature on the development of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2020, 11, 228. [Google Scholar] [CrossRef]
- Shylesha, A.; Jalali, S.; Gupta, A.; Varshney, R.; Venkatesan, T.; Shetty, P.; Ojha, R.; Ganiger, P.C.; Navik, O.; Subaharan, K. Studies on new invasive pest Spodoptera frugiperda (J.E. Smith)(Lepidoptera: Noctuidae) and its natural enemies. J. Biol. Control 2018, 32, 1–7. [Google Scholar] [CrossRef]
- Wu, P.; Ren, Q.; Wang, W.; Ma, Z.; Zhang, R. A bet-hedging strategy rather than just a classic fast life-history strategy exhibited by invasive fall armyworm. Entomol. Gen. 2021, 41, 337–344. [Google Scholar] [CrossRef]
- Wan, J.; Huang, C.; Li, C.; Zhou, H.; Ren, Y.; Li, Z.; Xing, L.; Zhang, B.; Xi, Q.; Bo, L. Biology, invasion and management of the agricultural invader: Fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Integr. Agric. 2021, 20, 646–663. [Google Scholar] [CrossRef]
- Kumar, R.M.; Nadagouda, B.; Hiremath, S. Studies on farmers perception about maize based cropping system in irrigated ecosystem of Gataprabha left bank cannel. Plant Arch. 2015, 15, 959–961. [Google Scholar]
- Rahmathulla, V.; Kumar, C.; Angadi, B.; Sivaprasad, V. Association of climatic factors on population dynamics of leaf roller, Diaphania pulverulentalis hampson (Lepidoptera: Pyralidae) in mulberry plantations of sericulture seed farm. Psyche 2012, 2012, 186214. [Google Scholar]
- Mitchell, E.R. Relationship of planting date to damage by earworms in commercial sweet corn in north central Florida. Fla. Entomol. 1978, 61, 251–255. [Google Scholar] [CrossRef]
- Teare, I.; Wright, D.; Sprenkel, R. Early planting reduces fall armyworm problems in no-till tropical corn. In Conservation Tillage for Agriculture in the 1990’s; North Carolina State University: Raleigh, NC, USA, 1990; p. 38. [Google Scholar]
- Kandel, S.; Poudel, R. Fall armyworm (Spodoptera Frugiperda) in maize: An emerging threat in Nepal and its management. Int. J. Appl. Sci. Biotechnol. 2020, 8, 305–309. [Google Scholar] [CrossRef]
- FAO. Integrated Management of the Fall Armyworm on Maize: A Guide for Farmer Field Schools in Africa; Food and Agriculture Organization: Rome, Italy, 2018. [Google Scholar]
- Tippannavar, P.; Talekar, S.; Mallapur, C.; Kachapur, R.; Salakinkop, S.; Harlapur, S. An outbreak of fall armyworm in Indian subcontinent: A new invasive pest on maize. Maydica 2019, 64, 10. [Google Scholar]
- Bhusal, S.; Chapagain, E. Threats of fall armyworm (Spodoptera frugiperda) incidence in Nepal and it’s integrated management—A review. J. Agric. Nat. Resour. 2020, 3, 345–359. [Google Scholar] [CrossRef]
- Alyokhin, A.; Nault, B.; Brown, B. Soil conservation practices for insect pest management in highly disturbed agroecosystems—A review. Entomol. Exp. Appl. 2020, 168, 7–27. [Google Scholar] [CrossRef]
- Rowen, E.K.; Regan, K.H.; Barbercheck, M.E.; Tooker, J.F. Is tillage beneficial or detrimental for insect and slug management? A meta-analysis. Agric. Ecosyst. Environ. 2020, 294, 106849. [Google Scholar] [CrossRef]
- Lal, R.; Reicosky, D.C.; Hanson, J.D. Evolution of the plow over 10,000 years and the rationale for no-till farming. Soil Tillage Res. 2007, 93, 1–12. [Google Scholar] [CrossRef]
- Kumar, H.; Mihm, J.A. Fall armyworm (Lepidoptera: Noctuidae), southwestern corn borer (Lepidoptera: Pyralidae) and sugarcane borer (Lepidoptera: Pyralidae) damage and grain yield of four maize hybrids in relation to four tillage systems. Crop Prot. 2002, 21, 121–128. [Google Scholar] [CrossRef]
- Clark, M.S. Generalist Predators in Reduced-Tillage Corn: Predation on Armyworm, Habitat Preferences, and a Method to Estimate Absolute Densities. Ph.D. Thesis, Virginia Tech, Blacksburg, VA, USA, 1993. [Google Scholar]
- Roberts, P.M.; All, J.N. Hazard for fall armyworm (Lepidoptera: Noctuidae) infestation of maize in double-cropping systems using sustainable agricultural practices. Fla. Entomol. 1993, 76, 276–283. [Google Scholar] [CrossRef]
- Rivers, A.; Barbercheck, M.; Govaerts, B.; Verhulst, N. Conservation agriculture affects arthropod community composition in a rainfed maize–wheat system in central Mexico. Appl. Soil Ecol. 2016, 100, 81–90. [Google Scholar] [CrossRef]
- Thierfelder, C.; Rusinamhodzi, L.; Ngwira, A.R.; Mupangwa, W.; Nyagumbo, I.; Kassie, G.T.; Cairns, J.E. Conservation agriculture in Southern Africa: Advances in knowledge. Renew. Agric. Food Syst. 2015, 30, 328–348. [Google Scholar] [CrossRef] [Green Version]
- Thierfelder, C.; Rusinamhodzi, L.; Setimela, P.; Walker, F.; Eash, N.S. Conservation agriculture and drought-tolerant germplasm: Reaping the benefits of climate-smart agriculture technologies in central Mozambique. Renew. Agric. Food Syst. 2016, 31, 414–428. [Google Scholar] [CrossRef]
- Harrison, R.D.; Thierfelder, C.; Baudron, F.; Chinwada, P.; Midega, C.; Schaffner, U.; van den Berg, J. Agro-ecological options for fall armyworm (Spodoptera frugiperda JE Smith) management: Providing low-cost, smallholder friendly solutions to an invasive pest. J. Environ. Manag. 2019, 243, 318–330. [Google Scholar] [CrossRef]
- Altieri, M.A.; Nicholls, C.I. Soil fertility management and insect pests: Harmonizing soil and plant health in agroecosystems. Soil Tillage Res. 2003, 72, 203–211. [Google Scholar] [CrossRef]
- Morales, H.; Perfecto, I.; Ferguson, B. Traditional fertilization and its effect on corn insect populations in the Guatemalan highlands. Agric. Ecosyst. Environ. 2001, 84, 145–155. [Google Scholar] [CrossRef]
- Singh, G. Improving Integrated Pest Management Strategies for the Fall Armyworm (Lepidoptera: Noctuidae) in Turfgrass. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2020. [Google Scholar]
- Kumar, R.M.; Girijesh, G. Yield potential, biological feasibility, economic viability of maize (Zea mays L.) and local field bean (Dolichos lablab L.) intercropping system in southern transitional zone of Karnataka. Res. Environ. Life Sci. 2015, 8, 27–30. [Google Scholar]
- Cannon, N.D.; Kamalongo, D.M.; Conway, J.S. The effect of bi-cropping wheat (Triticum aestivum) and beans (Vicia faba) on forage yield and weed competition. Biol. Agric. Hortic. 2020, 36, 1–15. [Google Scholar] [CrossRef]
- Sida, T.S.; Baudron, F.; Kim, H.; Giller, K.E. Climate-smart agroforestry: Faidherbia albida trees buffer wheat against climatic extremes in the Central Rift Valley of Ethiopia. Agric. For. Meteorol. 2018, 248, 339–347. [Google Scholar] [CrossRef]
- Van Huis, A. Integrated Pest Management in the Small Farmer’s Maize Crop in Nicaragua; Wageningen University and Research: Wageningen, The Netherlands, 1981. [Google Scholar]
- Khan, Z.R.; Midega, C.A.; Bruce, T.J.; Hooper, A.M.; Pickett, J.A. Exploiting phytochemicals for developing a ‘push–pull’crop protection strategy for cereal farmers in Africa. J. Exp. Bot. 2010, 61, 4185–4196. [Google Scholar] [CrossRef] [PubMed]
- Midega, C.A.; Khan, Z.R.; Pickett, J.A.; Nylin, S. Host plant selection behaviour of Chilo partellus and its implication for effectiveness of a trap crop. Entomol. Exp. Appl. 2011, 138, 40–47. [Google Scholar] [CrossRef]
- Altieri, M.A.; Francis, C.A.; Van Schoonhoven, A.; Doll, J.D. A review of insect prevalence in maize (Zea mays L.) and bean (Phaseolus vulgaris L.) polycultural systems. Field Crops Res. 1978, 1, 33–49. [Google Scholar] [CrossRef]
- Altieri, M. Diversification of corn agroecosystems as a means of regulating fall armyworm [Spodoptera frugiperda] populations. Fla. Entomol. 1980, 63, 450–456. [Google Scholar] [CrossRef]
- Khan, Z.R.; Midega, C.A.; Hassanali, A.; Pickett, J.A.; Wadhams, L.J. Assessment of different legumes for the control of Striga hermonthica in maize and sorghum. Crop Sci. 2007, 47, 730–734. [Google Scholar] [CrossRef]
- Tanyi, C.B.; Nkongho, R.N.; Okolle, J.N.; Tening, A.S.; Ngosong, C. Effect of intercropping beans with maize and botanical extract on fall armyworm (Spodoptera frugiperda) infestation. Int. J. Agron. 2020, 2020, 4618190. [Google Scholar] [CrossRef]
- Patil, S.B.; Goyal, A.; Chitgupekar, S.S.; Kumar, S.; El-Bouhssini, M. Sustainable management of chickpea pod borer. A review. Agron. Sustain. Dev. 2017, 37, 20. [Google Scholar] [CrossRef]
- Mooventhan, P.; Baskaran, R.; Kaushal, J.; Kumar, J. Integrated Management of Fall Armyworm in Maize; ICAR-National Institute of Biotic Stress Management: Raipur, India, 2019; p. 225. [Google Scholar]
- Guera, O.G.M.; Castrejón-Ayala, F.; Robledo, N.; Jiménez-Pérez, A.; Sánchez-Rivera, G. Plant selection for the establishment of push–pull strategies for zea mays–spodoptera frugiperda pathosystem in Morelos, Mexico. Insects 2020, 11, 349. [Google Scholar] [CrossRef]
- Islam, M.A. Pheromone use for insect control: Present status and prospect in Bangladesh. Int. J. Agric. Res. Innov. Technol. 2012, 2, 47–55. [Google Scholar] [CrossRef]
- Jiang, N.J.; Mo, B.T.; Guo, H.; Yang, J.; Tang, R.; Wang, C.Z. Revisiting the sex pheromone of the fall armyworm Spodoptera frugiperda, a new invasive pest in South China. Insect Sci. 2022, 29, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Cruz, I.; Figueiredo, M.d.L.C.; Silva, R.B.d.; Silva, I.F.d.; Paula, C.d.; Foster, J.E. Using sex pheromone traps in the decision-making process for pesticide application against fall armyworm (Spodoptera frugiperda [Smith] [Lepidoptera: Noctuidae]) larvae in maize. Int. J. Pest Manag. 2012, 58, 83–90. [Google Scholar] [CrossRef]
- Batista-Pereira, L.G.; Stein, K.; de Paula, A.F.; Moreira, J.A.; Cruz, I.; Figueiredo, M.d.L.C.; Perri, J.; Corrêa, A.G. Isolation, identification, synthesis, and field evaluation of the sex pheromone of the Brazilian population of Spodoptera frugiperda. J. Chem. Ecol. 2006, 32, 1085–1099. [Google Scholar] [CrossRef] [PubMed]
- Sanches, R.E.; Suzukawa, A.K.; Contreras-Soto, R.I.; Rizzardi, D.A.; Kuki, M.C.; Zeffa, D.M.; Albuquerque, F.A.d.; Scapim, C.A. Multivariate analysis reveals key traits of fall armyworm resistance in tropical popcorn genotypes. Bragantia 2019, 78, 175–182. [Google Scholar] [CrossRef]
- Chen, Y.; Ni, X.; Buntin, G.D. Physiological, nutritional, and biochemical bases of corn resistance to foliage-feeding fall armyworm. J. Chem. Ecol. 2009, 35, 297–306. [Google Scholar] [CrossRef]
- Smith, W.; Shivaji, R.; Williams, W.; Luthe, D.; Sandoya, G.; Smith, C.; Sparks, D.; Brown, A. A maize line resistant to herbivory constitutively releases (E)-β-caryophyllene. J. Econ. Entomol. 2012, 105, 120–128. [Google Scholar] [CrossRef]
- Ni, X.; Chen, Y.; Hibbard, B.E.; Wilson, J.P.; Williams, W.P.; Buntin, G.D.; Ruberson, J.R.; Li, X. Foliar resistance to fall armyworm in corn germplasm lines that confer resistance to root-and ear-feeding insects. Fla. Entomol. 2011, 94, 971–981. [Google Scholar] [CrossRef]
- Alvarez, M.d.P.; Miranda Filho, J.B.d. Diallel crossing among maize populations for resistance to fall armyworm. Sci. Agric. 2002, 59, 731–741. [Google Scholar] [CrossRef]
- Kasoma, C.; Shimelis, H.; Laing, M.; Shayanowako, A.I.; Mathew, I. Screening of inbred lines of tropical maize for resistance to fall armyworm, and for yield and yield-related traits. Crop Prot. 2020, 136, 105218. [Google Scholar] [CrossRef]
- De Souza Tavares, W.; Cruz, I.; Petacci, F.; de Assis Júnior, S.L.; de Sousa Freitas, S.; Zanuncio, J.C.; Serrão, J.E. Potential use of Asteraceae extracts to control Spodoptera frugiperda (Lepidoptera: Noctuidae) and selectivity to their parasitoids Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) and Telenomus remus (Hymenoptera: Scelionidae). Ind. Crops Prod. 2009, 30, 384–388. [Google Scholar] [CrossRef]
- Hruska, A.J. Fall armyworm (Spodoptera frugiperda) management by smallholders. CAB Rev. 2019, 14, 1–11. [Google Scholar] [CrossRef]
- Li, Y.; Hallerman, E.M.; Wu, K.; Peng, Y. Insect-resistant genetically engineered crops in China: Development, application, and prospects for use. Annu. Rev. Entomol. 2020, 65, 273–292. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Romeis, J. Managing the invasive fall armyworm through biotech crops: A Chinese perspective. Trends Biotechnol. 2021, 39, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Romeis, J.; Naranjo, S.E.; Meissle, M.; Shelton, A.M. Genetically engineered crops help support conservation biological control. Biol. Control 2019, 130, 136–154. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef]
- Li, J.-J.; Shi, Y.; Wu, J.-N.; Li, H.; Smagghe, G.; Liu, T.-X. CRISPR/Cas9 in lepidopteran insects: Progress, application and prospects. J. Insect Physiol. 2021, 135, 104325. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Gul, H.; Tariq, K.; Hafeez, M.; Desneux, N.; Gao, X.; Song, D. RNA interference-mediated silencing of ecdysone receptor (EcR) gene causes lethal and sublethal effects on melon aphid, Aphis gossypii. Entomol. Gen. 2022. [Google Scholar] [CrossRef]
- Books, A. New Biotechnological Approaches to Insect Pest Management and Crop Protection; Gene Editing Approach (CRISPR-Cas System); University of Nebraska—Lincoln: Lincoln, NE, USA, 2019. [Google Scholar]
- Wu, K. Management strategies of fall armyworm (Spodoptera frugiperda) in China. Plant Prot. 2020, 46, 1–5. [Google Scholar]
- Zhu, G.-H.; Chereddy, S.C.; Howell, J.L.; Palli, S.R. Genome editing in the fall armyworm, Spodoptera frugiperda: Multiple sgRNA/Cas9 method for identification of knockouts in one generation. Insect Biochem. Mol. Biol. 2020, 122, 103373. [Google Scholar] [CrossRef]
- Baranek, J.; Banaszak, M.; Lorent, D.; Kaznowski, A.; Konecka, E. Insecticidal activity of Bacillus thuringiensis Cry1, Cry2 and Vip3 toxin combinations in Spodoptera exigua control: Highlights on synergism and data scoring. Entomol. Gen. 2021, 41, 71–82. [Google Scholar] [CrossRef]
- Liang, H.-Y.; Yang, X.-M.; Sun, L.-J.; Zhao, C.-D.; Chi, H.; Zheng, C.-Y. Sublethal effect of spirotetramat on the life table and population growth of Frankliniella occidentalis (Thysanoptera: Thripidae). Entomol. Gen. 2021, 41, 219–231. [Google Scholar] [CrossRef]
- Gul, H.; Ullah, F.; Hafeez, M.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Sublethal concentrations of clothianidin affect fecundity and key demographic parameters of the chive maggot, Bradysia odoriphaga. Ecotoxicology 2021, 30, 1150–1160. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Malusi, P.; Likhayo, P.; Mendesil, E.; Elibariki, N.; Wakgari, M.; Ayalew, G.; Tefera, T. First report of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), natural enemies from Africa. J. Appl. Entomol. 2018, 142, 800–804. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Desneux, N.; Gao, X.; Song, D. Imidacloprid-induced hormesis effects on demographic traits of the melon aphid, Aphis gossypii. Entomol. Gen. 2019, 39, 325–337. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Thiamethoxam induces transgenerational hormesis effects and alteration of genes expression in Aphis gossypii. Pestic. Biochem. Physiol. 2020, 165, 104557. [Google Scholar] [CrossRef]
- Wang, X.; Xu, X.; Ullah, F.; Ding, Q.; Gao, X.; Desneux, N.; Song, D. Comparison of full-length transcriptomes of different imidacloprid-resistant strains of Rhopalosiphum padi (L.). Entomol. Gen. 2021, 41, 289–304. [Google Scholar] [CrossRef]
- Cutler, G.C.; Amichot, M.; Benelli, G.; Guedes, R.N.C.; Qu, Y.; Rix, R.R.; Ullah, F.; Desneux, N. Hormesis and insects: Effects and interactions in agroecosystems. Sci. Total Environ. 2022, 825, 153899. [Google Scholar] [CrossRef] [PubMed]
- Gowda, G.B.; Sahu, M.; Ullah, F.; Patil, N.B.; Adak, T.; Pokhare, S.; Mahendiran, A.; Rath, P.C. Insecticide-induced hormesis in a factitious host, Corcyra cephalonica, stimulates the development of its gregarious ecto-parasitoid, Habrobracon hebetor. Biol. Control 2021, 160, 104680. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Babin, A.; Lemauf, S.; Rebuf, C.; Poirié, M.; Gatti, J.-L. Effects of Bacillus thuringiensis kurstaki bioinsecticide on two non-target Drosophila larval endoparasitoid wasps. Entomol. Gen. 2022, 42, 611–620. [Google Scholar] [CrossRef]
- Akhtar, Z.R.; Tariq, K.; Handler, A.M.; Ali, A.; Ullah, F.; Ali, F.; Zang, L.-S.; Gulzar, A.; Ali, S. Toxicological risk assessment of some commonly used insecticides on Cotesia flavipes, a larval parasitoid of the spotted stem borer Chilo partellus. Ecotoxicology 2021, 30, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.-Y.; Xu, W.; Desneux, N.; Nkunika, P.O.; Bao, H.-P.; Zang, L.-S. Spodoptera frugiperda egg mass scale thickness modulates Trichogramma parasitoid performance. Entomol. Gen. 2022, 42, 589–596. [Google Scholar] [CrossRef]
- Gowda, B.; Pandi, G.G.P.; Ullah, F.; Patil, N.B.; Sahu, M.; Adak, T.; Pokhare, S.; Yadav, M.K.; Mahendiran, A.; Mittapelly, P.; et al. Performance of Trichogramma japonicum under field conditions as a function of the factitious host species used for mass rearing. PLoS ONE 2021, 16, e0256246. [Google Scholar]
- Molina-Ochoa, J.; Carpenter, J.E.; Heinrichs, E.A.; Foster, J.E. Parasitoids and parasites of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas and Caribbean Basin: An inventory. Fla. Entomol. 2003, 86, 254–289. [Google Scholar] [CrossRef]
- Pair, S.; Gross, H., Jr. Field mortality of pupae of the fall armyworm, Spodoptera frugiperda (JE Smith), by predators and a newly discovered parasitoid, Diapetimorpha introita. J. Ga. Entomol. Soc. 2012, 19, 22–26. [Google Scholar]
- Tendeng, E.; Labou, B.; Diatte, M.; Djiba, S.; Diarra, K. The fall armyworm Spodoptera frugiperda (JE Smith), a new pest of maize in Africa: Biology and first native natural enemies detected. Int. J. Biol. Chem. Sci. 2019, 13, 1011–1026. [Google Scholar] [CrossRef]
- Huang, N.-X.; Jaworski, C.; Desneux, N.; Zhang, F.; Yang, P.-Y.; Wang, S. Long-term, large-scale releases of Trichogramma promote pesticide decrease in maize in northeastern China. Entomol. Gen. 2020, 40, 331–335. [Google Scholar] [CrossRef]
- Zang, L.-S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status and perspectives. Annu. Rev. Entomol. 2021, 66, 463–484. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.-C.; Du, W.-M.; Zang, L.-S.; Ruan, C.-C.; Zhang, J.-J.; Zou, Z.; Monticelli, L.S.; Harwood, J.D.; Desneux, N. Multi-parasitism: A promising approach to simultaneously produce Trichogramma chilonis and T. dendrolimi on eggsof Antheraea pernyi. Entomol. Gen. 2021, 41, 627–636. [Google Scholar] [CrossRef]
- Santoiemma, G.; Tonina, L.; Marini, L.; Duso, C.; Mori, N. Integrated management of Drosophila suzukii in sweet cherry orchards. Entomol. Gen. 2020, 40, 297–305. [Google Scholar] [CrossRef]
- Akutse, K.S.; Khamis, F.M.; Ambele, F.C.; Kimemia, J.W.; Ekesi, S.; Subramanian, S. Combining insect pathogenic fungi and a pheromone trap for sustainable management of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Invertebr. Pathol. 2020, 177, 107477. [Google Scholar] [CrossRef] [PubMed]
- Assefa, F. Status of fall armyworm (Spodoptera frugiperda), biology and control measures on maize crop in Ethiopia: A review. Int. J. Entomol. Res. 2018, 6, 75–85. [Google Scholar] [CrossRef]
- Franz, L.; Raming, K.; Nauen, R. Recombinant expression of ABCC2 variants confirms the importance of mutations in extracellular loop 4 for Cry1F resistance in fall armyworm. Toxins 2022, 14, 157. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.G.; Wennmann, J.T.; Goergen, G.; Bryon, A.; Ros, V.I. Viruses of the fall armyworm Spodoptera frugiperda: A review with prospects for biological control. Viruses 2021, 13, 2220. [Google Scholar] [CrossRef]
- Valicente, F.H. Entomopathogenic viruses. In Natural Enemies of Insect Pests in Neotropical Agroecosystems; Springer: Cham, Switzerland, 2019; pp. 137–150. [Google Scholar]
- Lei, C.; Yang, S.; Lei, W.; Nyamwasa, I.; Hu, J.; Sun, X. Displaying enhancing factors on the surface of occlusion bodies improves the insecticidal efficacy of a baculovirus. Pest Manag. Sci. 2020, 76, 1363–1370. [Google Scholar] [CrossRef]
- Souza, C.; Silveira, L.; Souza, B.; Nascimento, P.; Damasceno, N.; Mendes, S. Efficiency of biological control for fall armyworm resistant to the protein Cry1F. Braz. J. Biol. 2020, 81, 154–163. [Google Scholar] [CrossRef]
- Nyffeler, M.; Şekercioğlu, Ç.H.; Whelan, C.J. Insectivorous birds consume an estimated 400–500 million tons of prey annually. Sci. Nat. 2018, 105, 47. [Google Scholar] [CrossRef]
- Patil, S.D.; Gaikwad, A.B. Awareness and knowledge of fall armyworm pest amongst maize growers in Dhule district. J. Entomol. Zool. Stud. 2021, 9, 122–126. [Google Scholar]
- Jones, G.A.; Sieving, K.E.; Avery, M.L.; Meagher, R.L. Parasitized and non-parasitized prey selectivity by an insectivorous bird. Crop Prot. 2005, 24, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Badshah, K.; Ullah, F.; Ahmad, B.; Ahmad, S.; Alam, S.; Ullah, M.; Jamil, M.; Sardar, S. Management of Lycoriella ingenua (Diptera: Sciaridae) on oyster mushroom (Pleurotus ostreatus) through different botanicals. Int. J. Trop. Insect Sci. 2021, 41, 1435–1440. [Google Scholar] [CrossRef]
- Rioba, N.B.; Stevenson, P.C. Opportunities and scope for botanical extracts and products for the management of fall armyworm (Spodoptera frugiperda) for smallholders in Africa. Plants 2020, 9, 207. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.; Ullah, F.; Khan, M.A.; Ahmad, S.; Jamil, M.; Sardar, S.; Tariq, K.; Ahmed, N. Efficacy of various natural plant extracts and the synthetic insecticide cypermethrin 25EC against Leucinodes orbonalis and their impact on natural enemies in brinjal crop. Int. J. Trop. Insect Sci. 2022, 42, 173–182. [Google Scholar] [CrossRef]
- Martínez, A.M.; Aguado-Pedraza, A.J.; Viñuela, E.; Rodríguez-Enríquez, C.L.; Lobit, P.; Gómez, B.; Pineda, S. Effects of ethanolic extracts of Argemone ochroleuca (Papaveraceae) on the food consumption and development of Spodoptera frugiperda (Lepidoptera: Noctuidae). Fla. Entomol. 2017, 100, 339–345. [Google Scholar] [CrossRef]
- Mkenda, P.; Mwanauta, R.; Stevenson, P.C.; Ndakidemi, P.; Mtei, K.; Belmain, S.R. Extracts from field margin weeds provide economically viable and environmentally benign pest control compared to synthetic pesticides. PLoS ONE 2015, 10, e0143530. [Google Scholar] [CrossRef]
- Mkindi, A.G.; Tembo, Y.L.; Mbega, E.R.; Smith, A.K.; Farrell, I.W.; Ndakidemi, P.A.; Stevenson, P.C.; Belmain, S.R. Extracts of common pesticidal plants increase plant growth and yield in common bean plants. Plants 2020, 9, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).