
Pollen Viability, and the Photosynthetic and Enzymatic Responses of Cowpea (Vigna unguiculata (L.) Walp., Fabaceae) in the Face of Rising Air Temperature: A Problem for Food Safety


 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phenological Cycle
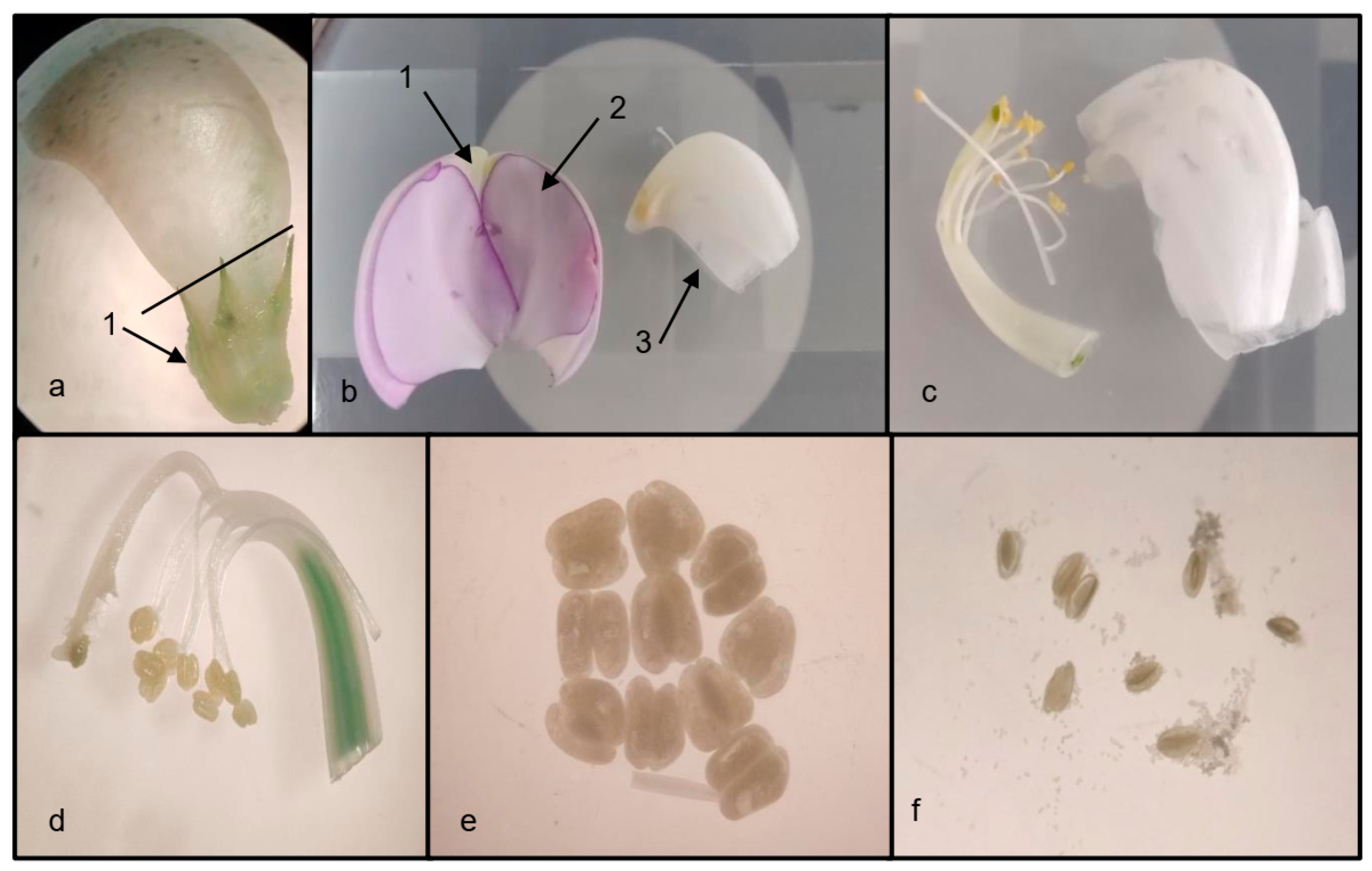
2.2. Pollen Viability
2.3. Physiological Evaluation and Enzymatic Activity
2.4. Productive Parameters
2.5. Statistical Analysis
3. Results
3.1. Phenological Cycle and Pollen Viability
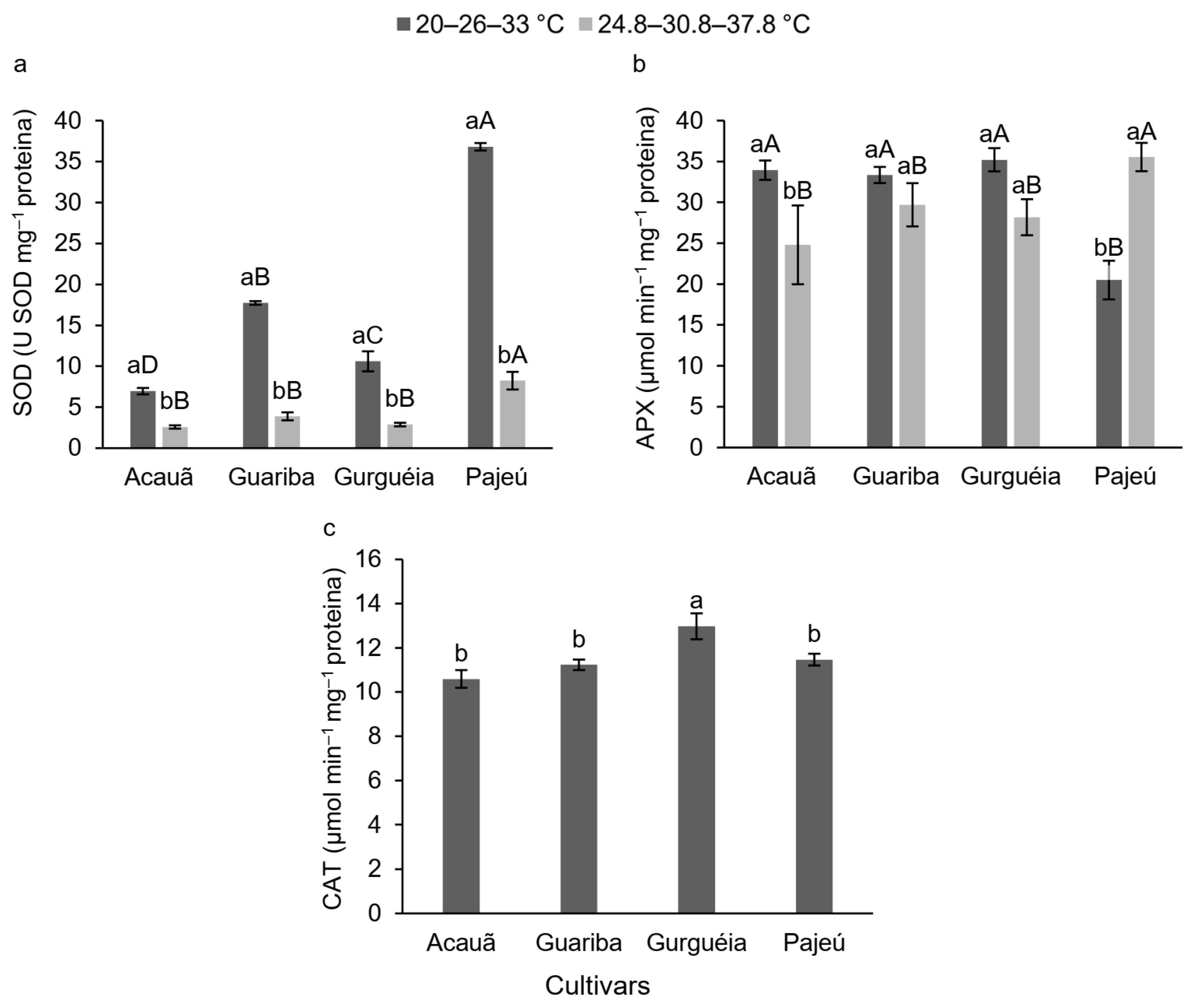
3.2. Physiological Evaluation and Enzymatic Activity
3.3. Productive Parameters
4. Discussion
4.1. Phenological Cycle
4.2. Pollen Viability
4.3. Physiological Evaluation and Enzymatic Activity
4.4. Productive Parameters
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lenaerts, B.; Collard, B.C.Y.; Demont, M. Review: Improving global food security through accelerated plant breeding. Plant Sci. 2019, 287, 110207. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef]
- Jameel, S.; Hameed, A.; Shah, T.M. Investigation of distinctive morpho-physio and biochemical alterations in desi chickpea at seedling stage under irrigation, heat, and combined stress. Front. Plant Sci. 2021, 12, 692745. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Nat. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef]
- Barros, J.R.A.; Guimarães, M.J.M.; Silva, R.M.; Rêgo, M.T.C.; Melo, N.F.; Chaves, A.R.M.; Angelotti, F. Selection of cowpea cultivars for high temperature tolerance: Physiological, biochemical and yield aspects. Physiol. Molec. Biol. Plants 2021, 27, 1–10. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Ed.; Cambridge University Press: Cambridge, UK, 2021; p. 42. [Google Scholar]
- Freire Filho, F.R.; Lima, J.A.A.; Ribeiro, V.Q. Feijão-Caupi: Avanços Tecnológicos; Embrapa Informações Tecnológicas: Brasília, Brazil, 2005; p. 519. [Google Scholar]
- Carvalho, M.; Castro, I.; Pereira, J.M.; Correia, C.; Cortines, M.E.; Matos, M.; Rosa, E.; Carnide, V.; Lino Neto, T. Evaluating stress responses in cowpea under drought stress. J. Plant Physiol. 2019, 241, 153001. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.S.; Silva, A.R.F.; Dutra, A.F.; Dutra, W.F.; Brito, M.E.B.; Silva, F.V. Photosynthetic efficiency and production of cowpea cultivars under deficit irrigation. Rev. Ambiente Água 2018, 13, 1–8. [Google Scholar] [CrossRef]
- Janska, A.; Marsik, P.; Zelenkova, S.; Ovesna, J. Cold stress and acclimation: What is important for metabolic adjustment? Plant Biol. 2010, 12, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Hossain, M.A.; Silva, J.A.T.; Fujita, M. Plant response and tolerance to abiotic oxidative stress: Antioxidant defense is a key factor. In Crop stress and Its Management: Perspectives and Strategies; Springer: Dordrecht, The Netherlands, 2012; Volume 15, pp. 261–315. [Google Scholar]
- Driedonks, N.; Wolters-Arts, M.; Huber, H.; Boer, G.J.; Vriezen, W.; Mariani, C.; Rieu, I. Exploring the natural variation for reproductive thermotolerance in wild tomato species. Euphytica 2018, 214, 67. [Google Scholar] [CrossRef]
- Cavalcanti, F.J.d.A. Recomendações de Adubação para o Estado de Pernambuco; 2ª Apr. 3.ed.; IPA: Recife, Brazil, 2008; p. 212. [Google Scholar]
- Oliveira, M.G.C.; Oliveira, L.F.C.; Wendland, A.; Guimarães, C.M.; Quintela, E.D.; Barbosa, F.R.; Carvalho, M.C.S.; Lobo Junior, M.; Silveira, P.M. Conhecendo a Fenologia do Feijoeiro e Seus Aspectos Fitotécnicos; Embrapa Arroz e Feijão: Brasília, Brazil, 2018; p. 59. [Google Scholar]
- Vargas, P.J.; Souza, S.A.M.; Silva, S.D.A.; Bobrowski, V.L. Análise dos grãos de pólen de diferentes cultivares de manona (Ricinus Communis L., Euphorbiaceae): Conservação e viabilidade. Comun. Cien. 2009, 76, 1–6. [Google Scholar] [CrossRef]
- Rocha, F.M.R.; Mousinho, S.F.; Freire Filho, F.R.; Silva, S.M.d.S.E.; Bezerra, A.A.d.C. Aspectos da biologia floral do caupi (Vigna unguiculata (L.) Walp.). In Reunião Nacional de Pesquisa de Caupi, 5, Teresina; Avanços tecnológicos no feijão caupi Anais; Embrapa Meio-Norte: Teresina, Brazil, 2001; pp. 27–29. [Google Scholar]
- Gomes, A.D.; Rossi, A.A.B.; Dardengo, J.F.E.; Silva, B.M.; Silva, I.V. Razão sexual e viabilidade polínica de Mauritia flexuosa L. (Arecaceae). In Enciclopédia Biosfera, Centro Científico Conhecer, 9th ed.; CNPq/Museu Paraense Emílio Goeldi: Belém, Brazil, 2013; pp. 2864–2870. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of proteindye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplast. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Havir, E.A.; Mchale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.J.; Wanjura, D.V. Plant responses to temperature extremes. In Physiology of Cotton; Stewart, J., Oosterhuis, D.M., Heitholt, J.J., Mauney, J.R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 123–128. [Google Scholar]
- Bergamashi, H.; Bergonci, J.I. As Plantas e o Clima: Princípios e Aplicações; Agrolivros: Guaíba, Brazil, 2017; p. 352. [Google Scholar]
- Vale, J.C.; Bertini, C.; Borem, A. Feijão-Caupi: Do Plantio à Colheita; Editora UFV: Viçosa, Brazil, 2017; p. 267. [Google Scholar]
- Angelotti, F.; Barbosa, L.G.; Barros, J.R.A.; Santos, C.A.F. Cowpea (Vigna unguiculata) development under different temperatures and carbon dioxide concentrations. Rev. Pesq. Agropec. Trop. 2020, 50, 1–7. [Google Scholar] [CrossRef]
- Ndiso, J.B.; Olubayo, F.; Chemining’wa, G.N.; Saha, H.M. Effect of drought stress on canopy temperature, growth and yield performance of cowpea varieties. Int. J. Plant Soil Sci. 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Matoso, A.O.; Soratto, R.P.; Guarnieri, F.; Costa, N.R.; Abrahão, R.C.; Tirabassi, L.H. Sowing date effects on cowpea cultivars as a second crop in southeastern Brazil. Agron. J. 2018, 110, 1799–1812. [Google Scholar] [CrossRef]
- Chaudhary, S.; Devi, P.; Bhardwaj, A.; Jha, U.C.; Sharma, K.D.; Prasad, P.V.V.; Siddique, K.H.M.; Bindumadhava, H.; Kumar, S.; Nayyar, D. Identification and characterization of contrasting genotypes/cultivars for developing heat tolerance in agricultural crops: Current status and prospects. Front. Plant Sci. 2020, 11, 1505. [Google Scholar] [CrossRef] [PubMed]
- Begcy, K.; Nosenko, T.; Zhou, L.Z.; Fragner, L.; Weckwerth, W.; Dresselhaus, T. Male sterility in maize after transient heat stress during the tetrad stage of pollen development. Plant Physiol. 2019, 181, 683–700. [Google Scholar] [CrossRef] [PubMed]
- Echer, F.R.; Oosterhuis, D.M.; Loka, D.A.; Rosolem, C.A. High night temperatures during the floral bud stage increase the abscission of reproductive structures in cotton. J. Agron. Crop. Sci. 2014, 200, 191–198. [Google Scholar] [CrossRef]
- Kaushal, N.; Bhandari, K.; Siddique, K.H.M.; Nayyar, H. Food crops face rising temperatures: An overview of responses, adaptive mechanisms, and approaches to improve heat tolerance. Cogent Food Agric. 2016, 2, 1134380. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Kumar, S.; Nayyar, H. Food legumes and rising temperatures: Effects, adaptive functional mechanisms specific to reproductive growth stage and strategies to improve heat tolerance. Front. Plant Sci. 2017, 8, 1658. [Google Scholar] [CrossRef] [PubMed]
- Sita, K.; Sehgal, A.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Identification of high-temperature tolerant lentil (Lens culinaris Medik.) genotypes through leaf and pollen traits. Front. Plant Sci. 2016, 8, 744. [Google Scholar] [CrossRef]
- Paupière, M.J.; Haperen, P.V.; Rieu, I.; Visser, R.G.F.; Tikunov, Y.M.; Bovy, A.G. Screening for pollen tolerance to high temperatures in tomato. Euphytica 2017, 213, 130. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Muthurajan, R.; Oane, R.; Wheeler, T.R.; Heuer, S.; Bennett, J.; Craufurd, P.Q. Physiological and proteomic approaches to dissect reproductive stage heat tolerance in rice (Oryza sativa L.). J. Exp. Bot. 2010, 61, 143–156. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Bheemanahalli, R.; Jagadish, S.K. Field crops and the fear of heat stress—Opportunities, challenges and future directions. Field Crops Res. 2017, 200, 114–121. [Google Scholar] [CrossRef]
- Raja, M.M.; Vijayalakshmi, G.; Naik, M.L.; Basha, P.O.; Sergeant, K.; Hausman, J.F.; Khan, P.S.S.V. Pollen development and function under heat stress: From effects to responses. Acta Physiol. Plant 2019, 41, 47. [Google Scholar] [CrossRef]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. High temperature susceptibility of sexual reproduction in crop plants. J. Exp. Botany 2020, 71, 555–568. [Google Scholar] [CrossRef]
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, B. Carbohydrate accumulation in relation to heat stress tolerance in two creeping bentgrass cultivars. J. Am. Soc. Hortic. Sci. 2000, 125, 442–447. [Google Scholar] [CrossRef]
- Roitsch, T.; González, M.C. Function and regulation of plant invertases: Sweet sensations. Trends Plant Sci. 2004, 9, 606–613. [Google Scholar] [CrossRef]
- Li, M.; Ji, L.; Yang, X.; Meng, Q.; Guo, S. The protective mechanisms of CaHSP26 in transgenic tobacco to alleviate photoinhibition of PSII during chilling stress. Plant Cell Rep. 2012, 31, 1969–1979. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; Shaked, R.; Peet, M.; Pharr, D.; Zamski, E.; Rosenfeld, K. Pollen grains of heat tolerant tomato cultivars retain higher carbohydrate concentration under heat stress conditions. Sci. Hortic. 2006, 109, 212–217. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Fisiologia e Desenvolvimento Vegetal, 6th ed.; Artmed: Porto Alegre, Brazil, 2017. [Google Scholar]
- Kamal, M.; Saleem, M.F.; Wahid, M.A.; Shakeel, A. Effects of ascorbic acid on membrane stability and yield of heat-stressed BT cotton. J. Anim. Plant Sci. 2017, 27, 192–199. [Google Scholar]
- Begcy, K.; Weiger, A.; Egesa, A.; Dresselhaus, T. Compared to Australian cultivars, European summer wheat (Triticum aestivum) overreacts when moderate heat stress is applied at the pollen development stage. Agronomy 2018, 8, 99. [Google Scholar] [CrossRef]
- Sharma, L.; Priva, M.; Bindumadhava, H.B.; Nair, R.M.; Navvar, H. Influence of high temperature stress on growth, phenology and yield performance of mungbean [Vigna radiata (L.) Wilczek] under managed growth conditions. Sci. Hortic. 2016, 213, 379–391. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhou, L.; Liu, J.; Cao, Z.; Du, X.; Huang, F.; Pan, G.; Cheng, F. Involvement of CAT in the detoxification of HT-induced ROS burst in rice anther and its relation to pollen fertility. Plant Cell Rep. 2018, 37, 741–757. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Iba, K. Acclimative response to temperature stress in higher plants: Approaches of gene engineering for temperature tolerance. Annu. Rev. Plant Biol. 2002, 53, 225–245. [Google Scholar] [CrossRef]
- Yang, H.; Gu, X.; Ding, M.; Lu, W.; Lu, D. Heat stress during grain filling affects activities of enzymes involved in grain protein and starch synthesis in waxy maize. Sci. Rep. 2018, 8, 15665. [Google Scholar] [CrossRef]
- Barros, J.R.A.; Guimarães, M.J.M.; Simões, W.L.; Melo, N.F.; Angelotti, F. Water restriction in different phenological stages and increased temperature affect cowpea production. Rev. Ciênc. Agrotec. 2020, 45, e022120. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Pisipati, S.R.; Ristic, Z.; Bukovnik, U.; Fritz, A. Impact of nighttime temperature on physiology and growth of spring wheat. Crop Sci. 2008, 48, 2372–2380. [Google Scholar] [CrossRef]
- Camejo, D.; Jimenez, A.; Alarcon, J.J.; Torres, W.; Gomez, J.M.; Sevilla, F. Changes in photosynthetic parameters and antioxidant activities following heat-shock treatment in tomato plants. Funct. Plant Biol. 2006, 33, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, N.; Awasthi, R.; Gupta, K.; Gaur, P.; Siddique, K.H.M.; Nayyar, H. Heat-stress-induced reproductive failures in chickpea (Cicer arietinum) are associated with impaired sucrose metabolism in leaves and anthers. Funct. Plant Biol. 2013, 40, 1334–1349. [Google Scholar] [CrossRef]
- Kaur, R.; Bains, T.S.; Bindumadhava, H.; Nayyar, H. Responses of mungbean (Vigna radiata L.) genotypes to heat stress: Effects on reproductive biology, leaf function and yield traits. Sci. Hortic. 2015, 197, 527–541. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Djanaguiraman, M.; Perumal, R.; Ciampitti, I.A. Impact of high temperature stress on floret fertility and individual grain weight of grain sorghum: Sensitive stages and thresholds for temperature and duration. Front. Plant Sci. 2015, 6, 820. [Google Scholar] [CrossRef]
- Yuan, L.; Yuan, Y.; Liu, S.; Wang, J.; Zhu, S.; Chen, G.; Hou, J.; Wang, C. Influence of high temperature on photosynthesis antioxidative capacity of chloroplast and carbono assimilation among heat-tolerant and heat-susceptible genotypes of nonheading chinese cabbage. HortScience 2017, 52, 1464–1470. [Google Scholar] [CrossRef]
- Bheemanahalli, R.; Sunoj, V.S.J.; Saripalli, G.; Prasad, P.V.V.; Balyan, H.S.; Gupta, P.K.; Grant, N.; Gill, K.S.; Jagadish, S.V.K. Quantifying the impact of heat stress on pollen germination, seed set, and grain filling in spring wheat. Crop Sci. 2019, 59, 684–696. [Google Scholar] [CrossRef]
- Krishnamurthy, L.; Gaur, P.M.; Basu, P.S.; Chaturvedi, S.K.; Tripathi, S.; Vadez, V.; Rathore, A.; Varshney, R.K.; Gowda, L.L.C. Large genetic variation for heat tolerance in the reference collection of chickpea (Cicer arietinum L.) germplasm. Plant Genet. Resour. 2011, 9, 59–69. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature Regimes | Time/Temperature (°C) | |||
|---|---|---|---|---|
| 20 h to 6 h | 6 h to 10 h | 10 h to 15 h | 15 h to 20 h | |
| T1 (20–26–33 °C) | 20 | 26 | 33 | 26 |
| T2 (24.8–30.8–37.8 °C) | 24.8 | 30.8 | 37.8 | 30.8 |
| Phenological Cycle (Average of Days) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | Temperature | V0 | V1 | V2 | V3 | V4 | R5 | R6 | R7 | R8 | R9 | Ciclo |
| BRS Acauã | 20–26–33 °C | 1.0 a | 1.0 a | 8.0 a | 10.3 a | 26.5 b | 2.0 a | 1.0 b | 9.0 a | 7.3 a | 3.0 a | 69.0 |
| 24.8–30.8–37.8 °C | 1.5 a | 1.3 a | 4.5 b | 9.8 a | 32.0 a | 2.3 a | 23.0 a | 7.0 a | 6.3 a | 3.0 a | 90.5 | |
| BRS Guariba | 20–26–33 °C | 1.0 a | 1.5 a | 8.0 a | 7.5 b | 26.3 a | 2.0 a | 1.3 b | 10.0 a | 7.0 a | 3.0 a | 67.5 |
| 24.8–30.8–37.8 °C | 1.0 a | 1.0 a | 5.3 b | 11.0 a | 25.8 a | 2.0 a | 7.3 a | 8.8 b | 5.5 a | 3.0 a | 70.5 | |
| BRS Gurguéia | 20–26–33 °C | 1.0 a | 1.5 a | 7.5 a | 8.5 a | 43.8 a | 2.0 a | 1.3 a | 8.8 a | 4.3 a | 3.0 a | 79.5 |
| 24.8–30.8–37.8 °C | 1.0 a | 1.0 a | 6.8 a | 9.0 a | 44.8 a | 2.0 a | 2.0 a | 10.5 a | 5.0 a | 3.0 a | 81.7 | |
| BRS Pajeú | 20–26–33 °C | 1.0 a | 1.8 a | 8.5 b | 12.5 a | 34.0 a | 2.0 a | 2.5 b | 10.3 a | 6.0 a | 3.0 a | 81.5 |
| 24.8–30.8–37.8 °C | 1.0 a | 1.0 a | 6.0 a | 9.5 b | 34.8 a | 2.3 a | 19.5 a | 8.5 a | 5.3 a | 3.0 a | 90.8 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barros, J.R.A.; dos Santos, T.C.; Silva, E.G.F.; da Silva, W.O.; Guimarães, M.J.M.; Angelotti, F. Pollen Viability, and the Photosynthetic and Enzymatic Responses of Cowpea (Vigna unguiculata (L.) Walp., Fabaceae) in the Face of Rising Air Temperature: A Problem for Food Safety. Agronomy 2024, 14, 463. https://doi.org/10.3390/agronomy14030463
Barros JRA, dos Santos TC, Silva EGF, da Silva WO, Guimarães MJM, Angelotti F. Pollen Viability, and the Photosynthetic and Enzymatic Responses of Cowpea (Vigna unguiculata (L.) Walp., Fabaceae) in the Face of Rising Air Temperature: A Problem for Food Safety. Agronomy. 2024; 14(3):463. https://doi.org/10.3390/agronomy14030463
Chicago/Turabian StyleBarros, Juliane Rafaele Alves, Tatiane Cezario dos Santos, Elioenai Gomes Freire Silva, Weslley Oliveira da Silva, Miguel Julio Machado Guimarães, and Francislene Angelotti. 2024. "Pollen Viability, and the Photosynthetic and Enzymatic Responses of Cowpea (Vigna unguiculata (L.) Walp., Fabaceae) in the Face of Rising Air Temperature: A Problem for Food Safety" Agronomy 14, no. 3: 463. https://doi.org/10.3390/agronomy14030463
APA StyleBarros, J. R. A., dos Santos, T. C., Silva, E. G. F., da Silva, W. O., Guimarães, M. J. M., & Angelotti, F. (2024). Pollen Viability, and the Photosynthetic and Enzymatic Responses of Cowpea (Vigna unguiculata (L.) Walp., Fabaceae) in the Face of Rising Air Temperature: A Problem for Food Safety. Agronomy, 14(3), 463. https://doi.org/10.3390/agronomy14030463