1. Introduction
Grape belongs to the family Vitaceae, Vitis, and woody vine. Its fruit can be used for fresh food, wine making, juice making, and drying, and it is greatly loved by the public because of its diverse flavors and delicious taste [
1]. Since the 21st century, China’s grape planting area, output, grape consumption, and import of grape products have ranked among the highest in the world [
2]. However, one factor contributing to China’s low grape exports may be the heavy use of chemical fertilizers and pesticides by some orchards to maximize yields, which leads to soil hardening and degradation, fertility decline, and grape quality deterioration [
3,
4]. In addition, due to land constraints and regional developmental needs, orchards are operated under continuous crop systems. Long-term irrational cultivation may lead to soil deterioration, the accumulation of self-toxic substances, and an imbalance in microbial communities, resulting in a nutrient imbalance in the soil, which affects crop yield and quality [
5]. In recent years, to obtain high-quality organic grapes, large amounts of organic fertilizers have been applied. However, due to the uneven quality of the organic fertilizers used and the improper application methods, the amount applied often far exceeds the amount actually required for grape growth, resulting in poor plant growth [
6,
7]. Studies have shown that the soil nutrient content of vineyards is generally at very high or extremely high levels at present [
8], and the fertilization methods should be adjusted as soon as possible for large-scale and high-quality vineyards to achieve sustainable and efficient utilization of nutrients and improve the economic and environmental benefits of vineyards. How to improve fertilizer utilization efficiency, enhance soil conditions, and strengthen the effective absorption and utilization of nutrients by grapevines, thereby promoting grape growth and fruit quality, is a critical issue that needs to be addressed in contemporary vineyard production.
Plant growth-promoting bacteria (PGPB) [
9] colonize the plant rhizosphere, which can improve plant growth, increase yield, enhance soil fertility, and reduce pathogens as well as biotic or abiotic stressors [
10]. PGPB help plants by producing plant growth phytohormones such as indole acetic acid (IAA), cytokinins, and gibberellins [
11], fixing nitrogen [
12], solubilizing inorganic phosphate [
13], and producing siderophores, antibiotics, and fungicidal compounds with antagonistic effects against phytopathogenic microorganisms [
14]. Liu et al. [
15] showed that
Priestia aryabhattai JY17 and
P. aryabhattai JY22B possessed a variety of PGP traits and can be used as biofertilizers to increase soil soluble phosphorus content. Rolli et al. [
16] indicated that beneficial bacteria rapidly and intimately colonize the rhizoplane and the root system of grapevine and can be successfully used to promote the growth of grapevines in the field. The use of PGPB in the form of biofertilizers or inoculants to reduce the application of fertilizers and pesticides, thereby improving the soil environment, and increase plant yield is currently a significant research area in the fields of agriculture, microbiology, and biotechnology [
17]. However, few studies have combined organic fertilizers with PGPB to improve plant growth.
Five PGPB,
Bacillus velezensis YH-18,
B. velezensis YH-20,
B. megaterium ZS-3,
P. aryabhattai SK1-7, and
Rahnella aquatilis JZ-GX1, were screened in the forest pathology laboratory of Nanjing Forestry University, and they showed good growth promotion, yield increases, insect resistance, and disease resistance in pot experiments [
18,
19,
20]. These microbial strains have substantial application potential. Nonetheless, studies have shown that many bacterial strains are able to exert beneficial effects in laboratory cultures, but fewer strains are successful in promoting disease resistance in laboratory greenhouses, and far fewer bacterial strains function under practical conditions, i.e., in commercial greenhouses or in fields, which is closely related to the complex soil environment of fields and weather [
21]. For example, high soil alkalinity and salinity, heavy metal contamination, poor nutrient availability, water scarcity, high ultraviolet radiation, and extreme temperature fluctuations can limit the growth of many PGPB. It is very important for the market expansion of microbial inoculants to apply successful strains from the laboratory to field environments and verify their effectiveness. Introducing PGPB to organic fertilizers for combined application may not only enhance the survival rate of microbial inoculants in the field but also improve the efficiency of organic fertilizer utilization.
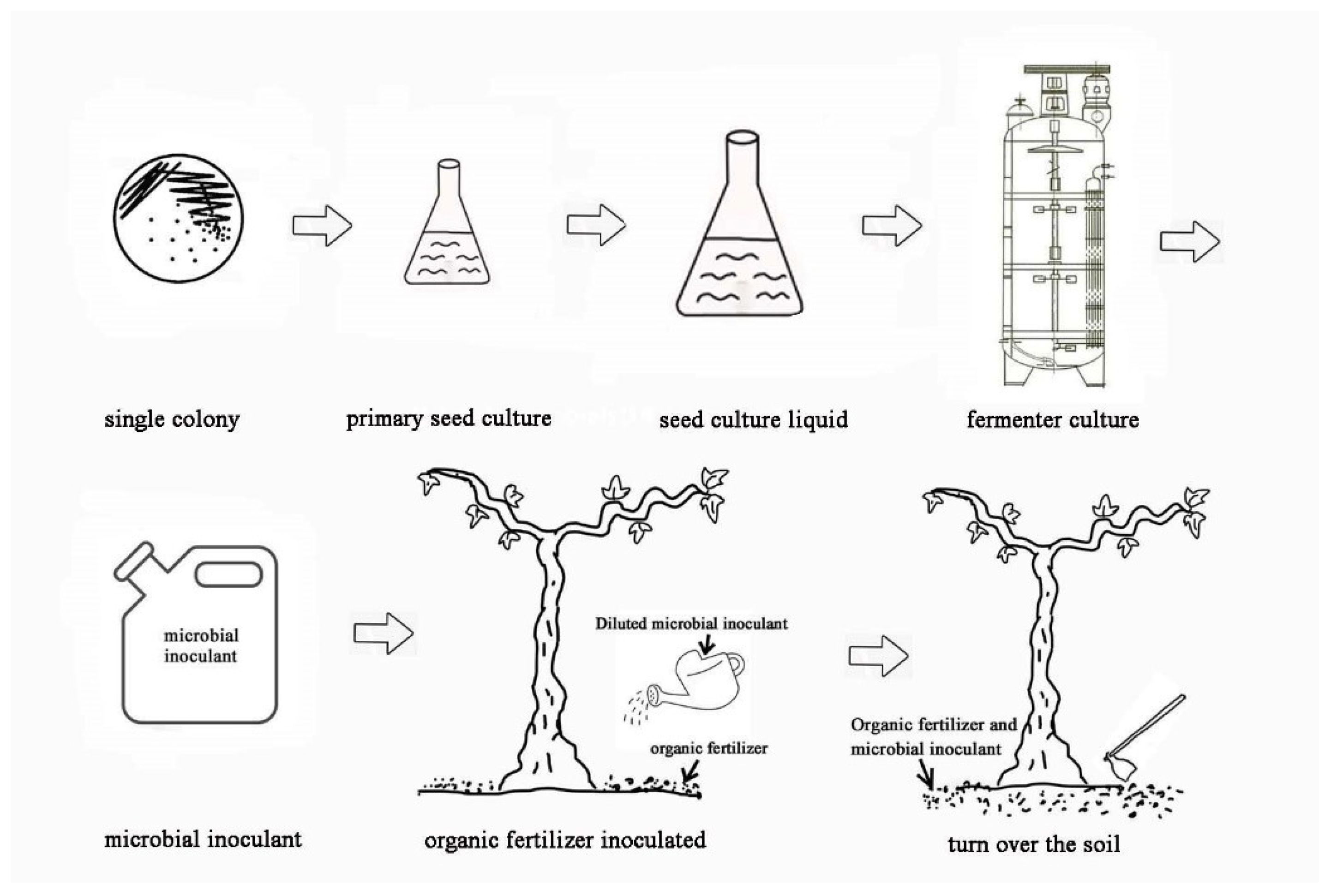
The main objectives of this study included the following: 1. to explore the PGP characteristics of the five PGPB and their possible growth-promoting mechanisms; 2. to investigate the effects of the five PGPB on the nutritional growth and fruit quality of grapes in combination with different organic fertilizers under field conditions and to screen for suitable microbial fertilizer types and successful application protocols for high-quality organic grape production; and 3. to investigate the stability of the PGPB in field trials by inoculating them for two consecutive years. The results of this study provide a theoretical and practical basis for the field application of five PGPB on grapes.
3. Results
3.1. Plant Growth-Promoting Characteristics of the Five Strains
The growth of bacteria in media in the presence of high salt concentrations is an important manifestation of their salt tolerance. YH-18, YH-20, and ZS-3 can grow on solid plates with NaCl contents in the range of 0 to 9%, but when the NaCl concentration is 9%, the growth is slow, and the number of colonies is small. SK1-7 and JZ-GX1 can grow on solid plates with NaCl contents in the range of 0 to 6%, but when the NaCl concentration is 6%, the growth is slow, and the number of colonies is small (
Table 3).
The growth of bacteria in a strongly alkaline medium is an important index that demonstrates their alkaline resistance. YH-20 can grow on a solid plate at a pH of 7–9. YH-18 grows on a solid plate at a pH of 7–10. ZS-3 and JZ-GX1 can grow on a plate at a pH of 7–11, but the strain grows slowly when the pH of the plate is 11. SK1-7 can grow on a plate at pH 7–12, and growth is still robust on a plate at pH 12 (
Table 3).
The five strains of bacteria were inoculated on a nitrogen-free medium, and strain growth was observed to preliminarily determine if they had nitrogen-fixing capacities. The results showed that all five strains could grow on a nitrogen-free medium, and all of them had certain nitrogen-fixing abilities (
Table 3).
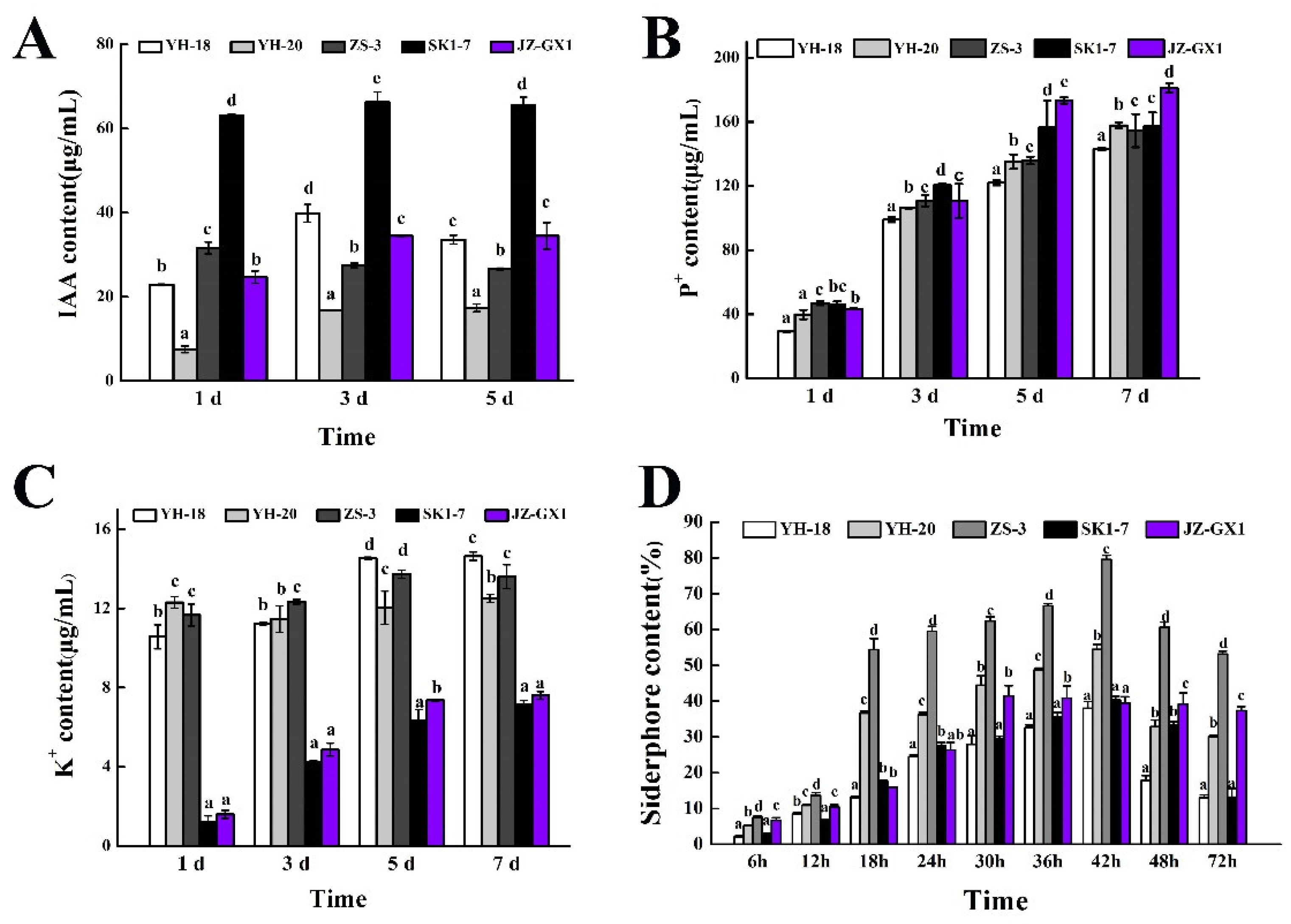
The IAA synthesis of the five strains was quantified by the Salkowski colorimetric method, and the results are shown in
Figure 3A. The IAA production of all five strains stabilized at approximately 3 days. At 5 days, the IAA production capacity of the five strains was SK1-7 > JZ-GX1 > YH-18 > ZS-3 > YH-20, and the amount of IAA produced at 5 days was 65.50, 34.45, 33.59, 26.63, and 17.40 μg/mL, respectively. In general studies, PGPB producing an IAA content of more than 30 μg/mL are considered to be high IAA-producing strains.
As shown in
Figure 3B, the amount of dissolved phosphorus in the culture containing the five strains increased rapidly during 1–3 days of culturing, and the amount of dissolved phosphorus increased gradually and steadily after 3 days of culturing. After 7 days of culturing, the phosphorus solubilization amounts of YH-18, YH-20, ZS-3, SK1-7, and JZ-GX1 were 143.07, 157.75, 154.54, 157.12, and 181.11 μg/mL, respectively.
Figure 3C indicates that the potassium-dissolving capacity of strains YH-18, YH-20, and ZS-3 increased rapidly on the first day of culture and then gradually stabilized, reaching 14.63, 12.50, and 13.6 μg/mL on the seventh day, respectively. The concentration of potassium ions in the SK1-7 and JZ-GX1 culture media increased rapidly after 1–5 days of culturing and then gradually stabilized after 5 days. By the seventh day, the amount of potassium released was 7.13 and 7.60 μg/mL, respectively.
Figure 3D shows that the content of siderophore produced by the five strains reached the highest after 42 h of culture, and the content of siderophore produced by ZS-3 and YH-20 was significantly higher than that of the other three strains.
3.2. Effect of Five PGPB in Combination with Two Different Organic Fertilizers on Grape Growth and Fruit Quality in the First Year
As shown in
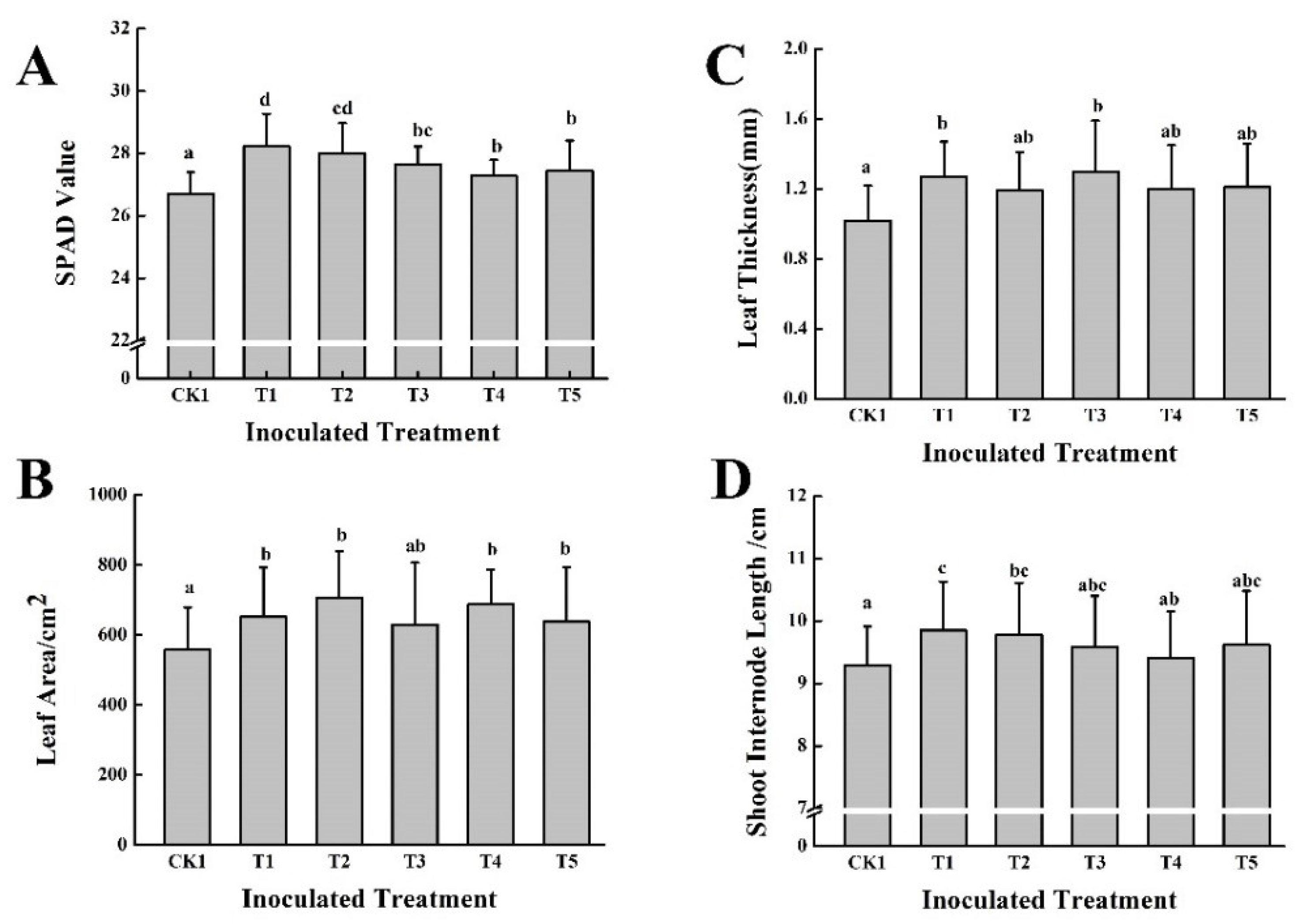
Table 4, the five treatments of cow dung fertilizer combined with microbial inoculant (T1~T5) showed no significant differences in SPAD values. The SPAD value of CK1 was significantly higher than that of CK2. In addition, the five treatments of distillers’ grain fertilizer combined with microbial inoculant (D1~D5) did not increase the SPAD value.
The results showed that the leaf area of the T1~T5 treatments was significantly higher than that of the CK1 treatment by 12.09–20.10%. The leaf area of CK2 was significantly higher than that of CK1. The D1~D5 treatments had no effect on the leaf area.
There was no significant effect of any of the treatments on leaf thickness.
Different treatments were applied to the grape vines, and the yield parameters of the fruits were determined (
Table 5). The T1~T5 treatments significantly promoted cluster biomass. Among them, the T3, T1, and T2 treatments were the most effective and increased fruit biomass by 73.4%, 66.3%, and 63.0%, respectively, relative to CK1. The cluster biomass of CK2 was significantly higher (39.1%) than that of CK1. Nevertheless, the D1~D5 treatments had no noticeable effect on cluster biomass.
The T1~T5 treatments increased the cross and longitudinal diameters of clusters and showed values significantly different from those of CK1; the cross diameters increased by 12.02–26.07%, and the longitudinal diameters increased by 9.28–18.90%. Compared with CK1, the cross and longitudinal diameters of the clusters of CK2 were higher; the cross diameter significantly increased by 8.66%. The cluster cross diameters of the D1~D5 treatments were significantly higher than that of CK2, while there was no significant difference in the cluster longitudinal diameter.
The T1~T5 treatments significantly increased berry biomass compared with CK1. Among them, the T1 and T2 treatments increased berry biomass most significantly, at 33.7% and 29.8% higher than CK1. The berry biomass of CK2 was significantly heavier than that of CK1, and the former increased by 20.74% compared with the latter. The berry biomass of the D1~D5 treatments was not significantly different from that of CK2.
The T1~T5 treatments showed no significant difference in the TSS content of grapes. There was no significant difference in TSS content between CK1 and CK2. Except for the D3 treatment, the D1~D5 treatments had no significant effect on the TSS content. Compared with CK2, the D3 treatment significantly increased the TSS content of fruit by 5.62%.
Table 6 shows that the T1~T5 treatments all promoted internode length, and the maximum value occurred in the T1 treatment (10.81 cm). There was no significant difference in internode length between CK1 and CK2. Other than the internode length of the D1 treatment, which increased by 13.80%, the other treatments had no marked difference from that of CK2.
The T1~T5 treatments increased shoot internode thickness, among which the T4 treatment had a marked effect, at 14.22% higher than CK1. There was no significant difference in shoot internode thickness between CK1 and CK2. Except for the internode thickness of the D5 treatment, which increased by 13.24%, the other treatments showed no marked difference from that of CK2.
The stem–pith ratio indicates the degree of lignification and maturity of the grape branches. The T1~T5 treatments significantly increased the stem–pith ratio, which was 7.64–18.78% higher than that of CK1. Compared with CK1, the stem–pith ratio of CK2 was improved, but the effect was not significant. The D1–D5 treatments had no significant difference compared with CK2 except D2, which significantly reduced the stem–pith ratio.
In this study, the T1~T5 treatments all improved the roundness of primary shoots, among which the T5 treatment showed the most significant improvement: 3.92% higher than that of CK1. Compared with CK1, the roundness of primary shoots in CK2 was also significantly improved: 4.65% higher than that of CK1. There was no difference in roundness between the D1~D5 treatments and CK2.
3.3. Effect of the Five PGPB in Combination with Cow Dung Organic Fertilizers on Grape Growth and Fruit Quality over Two Consecutive Years
The experimental results from 2019 to 2020 showed that the fruit quality using microbial inoculant combined with cow dung fertilizer was better than that of distillers’ grain fertilizer; additionally, the cost of microbial inoculant combined with cow dung fertilizer was lower and the benefit was greater, which was more in line with the needs of agricultural production. Therefore, during the second year, we continued to trace the effects of combining cow manure fertilizer with microbial inoculant.
After two years of continuous application of microbial inoculant with cow dung, the growth of grape seedlings was effectively promoted.
Figure 4 indicates that the T1~T5 treatments significantly increased the SPAD value of leaves, which was 5.69%, 4.83%, 3.52%, 2.13%, and 2.77% higher than that of CK1, respectively. Compared with CK1, the T1 and T3 treatments significantly increased the leaf thickness of grapes by 24.51% and 27.45%, respectively. The leaf area of the T1~T5 treatments also increased, and the leaf area effects of T1, T2, T4, and T5 were the most significant, increasing by 16.88%, 26.72%, 23.23%, and 14.41%, respectively, compared with CK1. The T1~T5 treatments increased the shoot internode length to different degrees, among which the T1 and T2 treatments increased shoot internode length most significantly, by 6.03% and 5.27% compared with CK1, respectively.
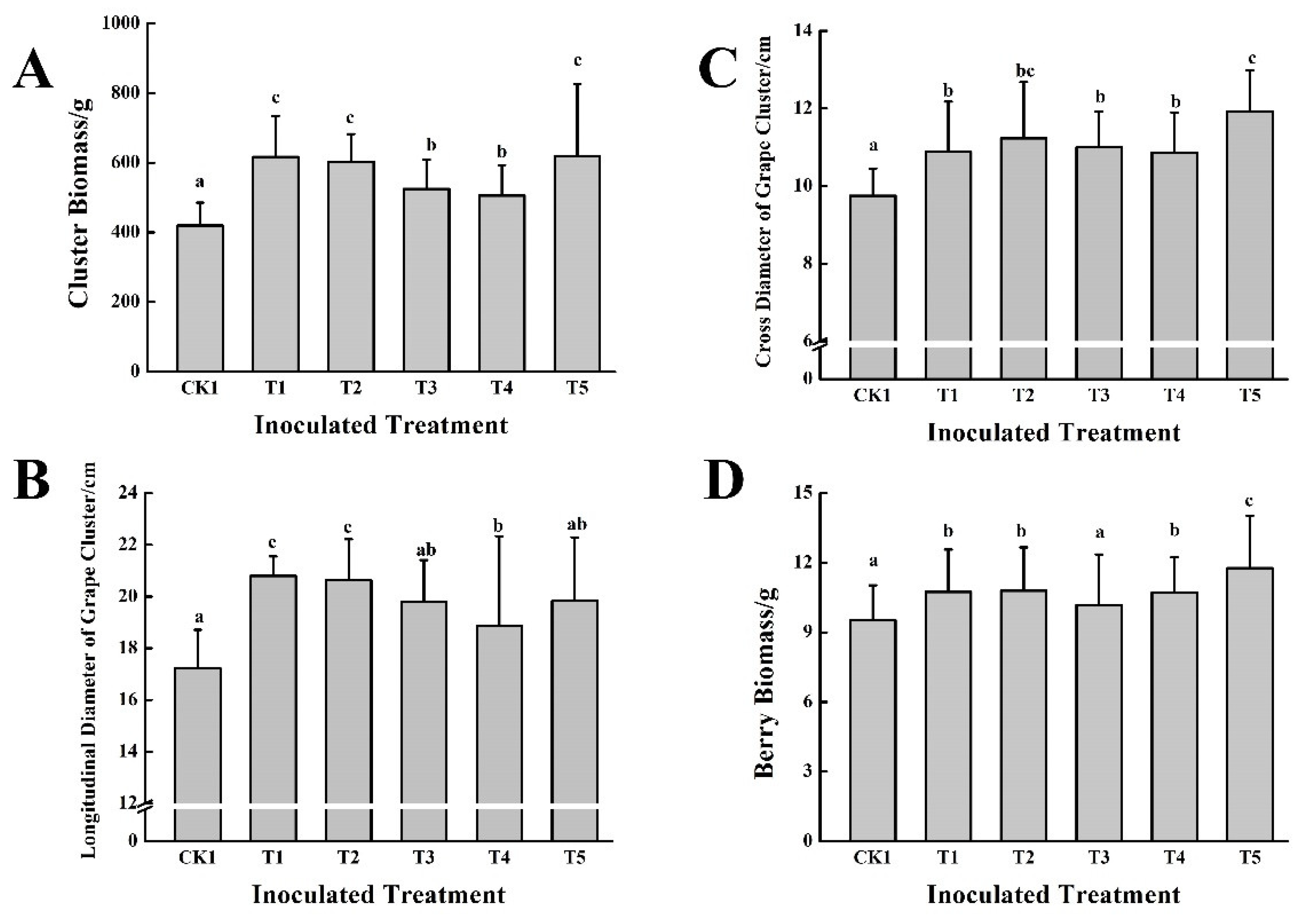
After two years of continuous application of microbial inoculants with cow dung fertilizer, the yield of grapes was effectively increased (
Figure 5). The T1~T5 treatments significantly increased the cluster biomass, which increased by 20.41~47.51% compared with CK1, and the T1, T2, and T5 treatments were significantly better than the other treatments. Compared with CK1, the cross diameter and longitudinal diameter of grape clusters in the T1~T5 treatments were significantly increased by 11.49~22.36% and 12.83~20.79%, respectively. Other than the T3 treatment, the berry biomass was significantly increased; the T1, T2, T4, and T5 treatments significantly increased the berry biomass by 12.70%, 13.22%, 12.49%, and 23.50%, respectively, compared with CK1.
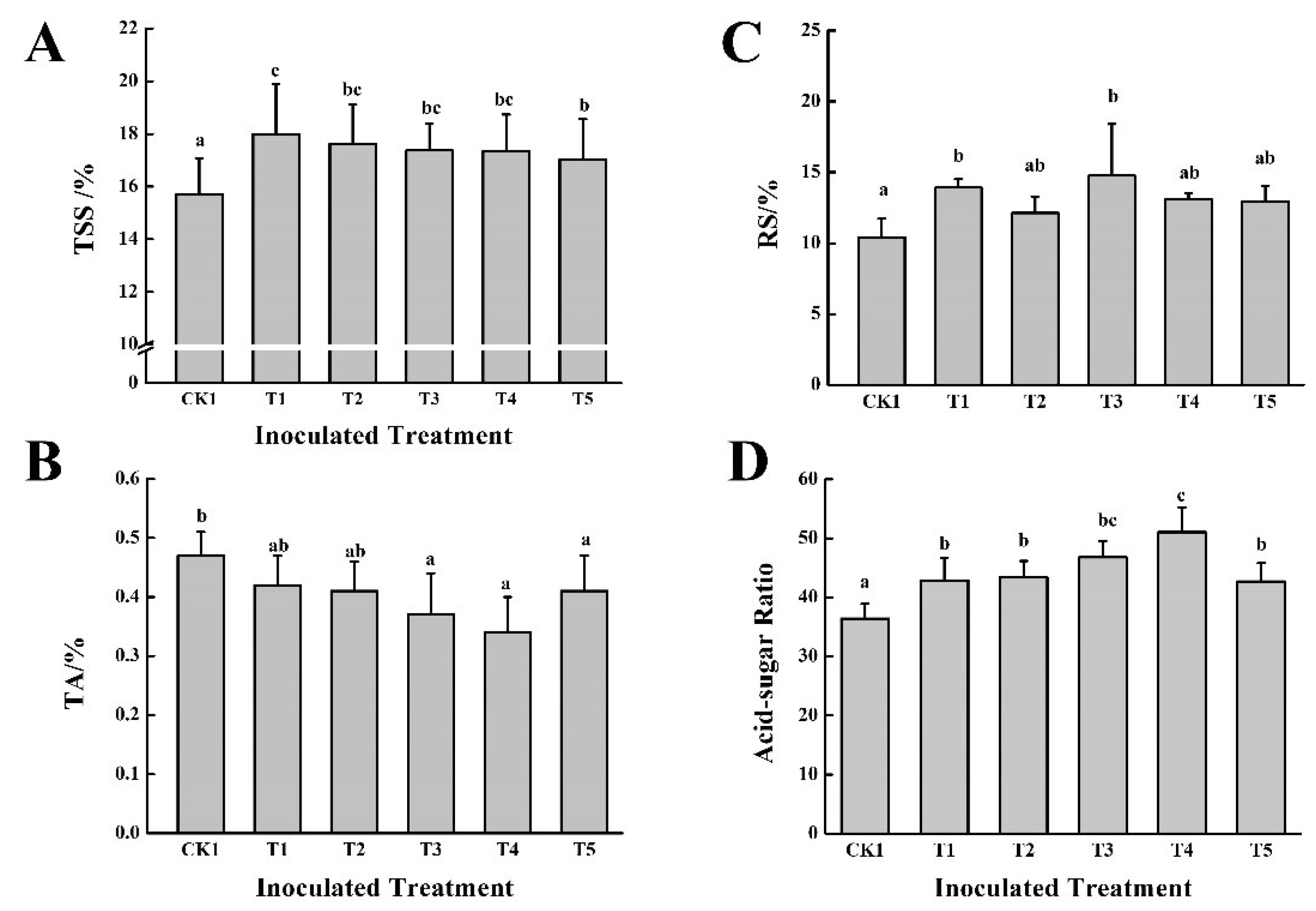
After two years of continuous application of microbial inoculants with cow dung fertilizer, the quality of grapes effectively improved (
Figure 6). The content of TSS in fruits that received the T1~T5 treatments increased significantly, by 14.58%, 12.22%, 10.63%, 10.44%, and 8.40%, respectively, compared with the control. The content of RS also increased to some extent, among which the T1 and T3 treatments showed the most significant increases, at 33.72% and 41.79%, respectively, compared with CK1. Compared with CK1, the titratable acid content of all treatments decreased, especially those of the T3 and T4 treatments, which decreased by 21.28% and 27.66%, respectively. The results of the solid-acid ratio showed that T4 > T3 > T2 > T1 > T5 > CK1.
4. Discussion
Many PGPB synthesize IAA, which is responsible for increasing numbers of adventitious roots that help plants uptake large volumes of nutrients and absorb water [
30]. In our study,
P. aryabhattai SK1-7 had the strongest IAA-producing capacity. Similarly, Pallab Kumar Gosh et al. [
31] found that
P. aryabhattai MCC3374 has the capacity to produce IAA efficiently and could be exploited for stress amelioration and plant growth enhancement in rice cultivars. Moreover, Li et al. [
32] from the Laboratory of Forest Protection Pathology of Nanjing Forestry University, showed that
R. aquatilis JZ-GX1 could secrete indole acetic acid and directly effectively promote seed germination and root length of maize and could secrete phytase and indirectly promote the growth of corn. Therefore, the other strains used in this study could also directly promote grape growth by producing IAA.
The availability of phosphorus in soil is usually limited, so the presence of phosphorus-solubilizing bacteria may be essential [
33]. The growth of phosphorus-increasing bacteria (PSB) often leads to soil acidification, which leads to the solubilization of phosphorus. However, microorganisms dissolve phosphorus and improve its uptake from the soil. Therefore, a continuous action of this type will lead to a reduction in the presence of phosphorus in the soil so it must be replenished on an ongoing basis from other sources (such as organic fertilizers) as a means of cycling nutrients through the soil. Torres et al. [
34] found that
B. velezensis XT1 could dissolve soil organic phosphorus and inorganic phosphorus and promote the growth of tomato stem and root biomass, plant height, and leaf number. Xiang et al. [
35] showed that
B. megaterium DSM3228, which is salt-tolerant and has phosphate-solubilizing functions, can effectively improve the uptake of phosphorus from soil by plants. The five PGPB tested in this study have strong phosphorus-solubilizing abilities and can be used as good PSB during agricultural production.
More than 90% of the potassium in soil exists in a slow-release state within silicate-rich minerals such as potassium feldspar and mica and cannot be directly absorbed and utilized by plants, resulting in poor growth of plants due to potassium deficiencies [
36]. Potassium-solubilizing bacteria (KSB) can dissolve aluminosilicate and apatite minerals and transform insoluble potassium, phosphorus, silicon, and other elements in soil into soluble forms, thus promoting absorption and utilization by plants [
37]. Li et al. [
38] found that
B. velezensis BA-26 can effectively improve soil nutrients and increase the content of available potassium in soil, thus increasing potato growth and reducing the use of chemical fertilizers. Chen et al. [
20] confirmed that SK1-7 has good growth-promoting effects on poplar and can effectively improve the available potassium content of poplar rhizosphere soil. In this study, the good potassium-releasing functions of the five PGP strains provided strong assurance of their ability to promote plant growth.
It has been suggested by Saha et al. [
39] that the production of siderophores is an important factor in phytopathogen antagonism and for the developmental growth of plants. In our study, we found that the five PGP strains were positive for siderophore production, and
B. megaterium ZS-3 had the highest productivity. Similarly, Chakraborty [
40] found that
B. megaterium DE BABY TRS-4 can reduce the occurrence of brown root rot in tea plants and promote the growth of tea plants by producing siderophores, antifungal metabolites and other active compounds. In addition, Kong et al. [
41] confirmed that inoculation with
R. aquatilis JZ-GX1 can induce camphor leaves to show higher levels of active iron and enhance rhizosphere acidification capacity and iron chelate reductase activity, thus treating iron deficiency chlorosis in camphor trees.
The results of the field trials showed that the effect of using only distillers’ grain fertilizer, which contained high organic matter content, on grape growth and fruit quality was significantly better than that of using only cow manure fertilizer, which contained low organic matter content. Grapes need sufficient amounts and various types of nutrients during their process of growth and development [
42]. The nutrients present in the highly decomposed distillers’ grain fertilizer are easily absorbed and utilized by vines, while the nutrients present in cow dung fertilizer, which is not highly decomposed, are not easily absorbed by vines [
43], resulting in large differences in tree growth and fruit quality. Therefore, the selection of high-quality organic fertilizer can reduce the excessive soil accumulation of fertilizer materials that cannot be degraded and absorbed and can also prevent root burn and seedling damage.
In this study, inoculation of cow manure fertilizer with PGPB was effective in enhancing tree vigor, increasing grape leaf area, and improving both fruit yield and quality. This is consistent with the findings of Wei et al. [
44], who showed that inoculation of biochar with
Pseudomonas putida improved grape fruit quality and altered bacterial diversity. Their results showed that inoculation with live microorganisms played a key role. The mechanisms by which the PGPB enhanced the morphological and yield parameters of grape are hypothesized to be their PGP traits [
45], which include nitrogen fixation [
46], phosphorus solubilization [
47], potassium dissolution, IAA, and siderophore production [
48]. Unlike the study by Wei et al. [
44], who used biochar as the carrier for PGPB, our study directly inoculated PGPB into organic fertilizer, using the organic fertilizer itself as the carrier. When PGPB are inoculated, the living microorganisms contact the available carbon and nitrogen sources in organic fertilizer, and they do not starve quickly because nutrients are readily available; this is conducive to bacterial reproduction, prolonged survival times, and improved colonization rates of strains in plant roots [
49,
50]. Notably, it is advantageous to have an initial high N content that promotes bacterial multiplication, but in subsequent stages with high N availability, the efficiency of atmospheric N fixation will be low. And free-living bacteria only release plant-available N after their death. The introduced microorganisms transform the nutrients that are difficult to absorb by plants in cow dung fertilizer and soil into nutrients that can be directly absorbed and utilized by plants via their PGP properties, which greatly increases the contents of organic matter and various bioactive substances in the soil, improves the fertilizer efficiency and utilization rate of organic fertilizer [
51], improves the soil ecological environment and flora structure, and provides sufficient nutrients and good habitats for plants [
52]. Previous research conducted by our team has also demonstrated that the inoculation of strains investigated in this study can improve soil nutrients and soil microecology. Additionally, there exists a significant positive correlation between soil nutrients and plant growth as documented in previous studies [
20,
53,
54]. This is in accordance with the research conducted by Asghar et al. [
55] wherein various PGPB they identified demonstrated increased wheat yield, improved nutritional and quality parameters, and enhanced availability of soil nutrients when employed in conjunction with organic and chemical fertilizers.
However, the effects of cow dung fertilizer inoculated with the five PGPB on the yield and quality of grapes were different, reflecting that different PGPB have different PGP abilities in the field and different fertilizer release abilities. Different strains might also have different colonization abilities in the rhizosphere, resulting in different final growth-promoting effects [
56,
57]. The five cow dung fertilizer inoculation treatments were more effective and less costly than treatment with distillers’ grain fertilizer alone. In practice, it may be possible to reduce the amount of organic fertilizer (with medium degrees of decomposition) needed by inoculating the fertilizer with PGPB.
After distillers’ grain fertilizer reaches high degrees of decomposition, it forms stable humus [
58]. There was no significant difference in grape yield and quality when distillers’ grain fertilizer was used in combination with PGPB compared with when distillers’ grain fertilizer was applied alone. This differs from the results of a study by da Silva, who showed that humic substances combined with PGP bacteria can enhance crop growth [
59]. It is also different from the results of the present study in which the treatments of cow dung fertilizer inoculated with PGPB significantly improved grape growth and fruit quality. We speculate that because the nutrients in the distillers’ grain fertilizer could be directly absorbed and used by plants, there was little room for material transformation via the added exogenous microorganisms. Grapes, on the other hand, are a very fertilizer-demanding species, and most of their nutrients come from applied fertilizers. The nutrients directly available from the fertilizer are much greater in quantity than those transformed by PGP bacteria, so inoculation with PGPB does not produce a significant efficiency boost. While cow dung fertilizer is not fully fermented, inoculation of PGPB can transform the unavailable fertilizing effect of the fertilizer into a large number of nutrients that can be directly absorbed by plants, thus significantly promoting the growth of grapes. One reason may be that distillers’ grain fertilizer is not suitable as the substrate for microbial inoculants, and the two cannot play a synergistic role. The residual alcohol, organic acids, and phenolic compounds in distillers’ grain fertilizer may exert inhibitory effects on inoculated PGPB, thereby restricting the growth of microorganisms. There may be an additional reason; when used in conjunction with high-nutrient-content distillers’ grain fertilizer, grapevines may experience nutrient excess and physiological imbalance, thereby limiting the performance of PGPB. However, in this study, field soil nutrients and soil physical and chemical properties were not determined, so the nutrient and improvement status of the soil could not be evaluated. Subsequently, a more in-depth investigation can be conducted into the nutrient changes following the inoculation of organic fertilizers with PGPB, aiming to better understand its underlying mechanisms.
Factors such as the climate and environment in fields can lead to unstable test results, so experiments are often carried out for many years in a row. In our study, it was found that cow manure fertilizer inoculation with the same PGPB for two consecutive years continued to significantly improve vine growth and fruit quality during the second year, which is consistent with the findings of Gong [
60] and others, who showed that a continuous application of bioorganic fertilizer significantly increased cucumber biomass and yield. Grape fruit qualities, including TSS, RS, and TA, are most important for fruit growers and are key factors that promote consumer purchases. During the second year, in addition to effective increases in fruit yield and fruit size, there were significant improvements in TSS and RS, reductions in TA, and enhancements in fruit taste, which makes for very good fruit for growers to sell. Fruit trees store nutrients in the fall and winter through their woody structure and root distribution, which work together to allow the tree to grow more effectively the following year [
61]. This also indicates that continuous soil fertility management is important for maintaining quality production and land health and that inoculation with PGPB improves soil microbial community structure and soil fertility, contributing to sustainable orchard management [
62,
63]. The use of organic fertilizers in combination with PGPB can be an option for sustainable agriculture.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}