
Polyaspartic Acid Urea Increased Maize Yield by Enhancing Leaf N Turnover Efficiency and Soil Microbial Diversity


Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Sampling and Measurements
2.4. Calculations
2.5. Statistical Analysis
3. Results
3.1. Seed Yield
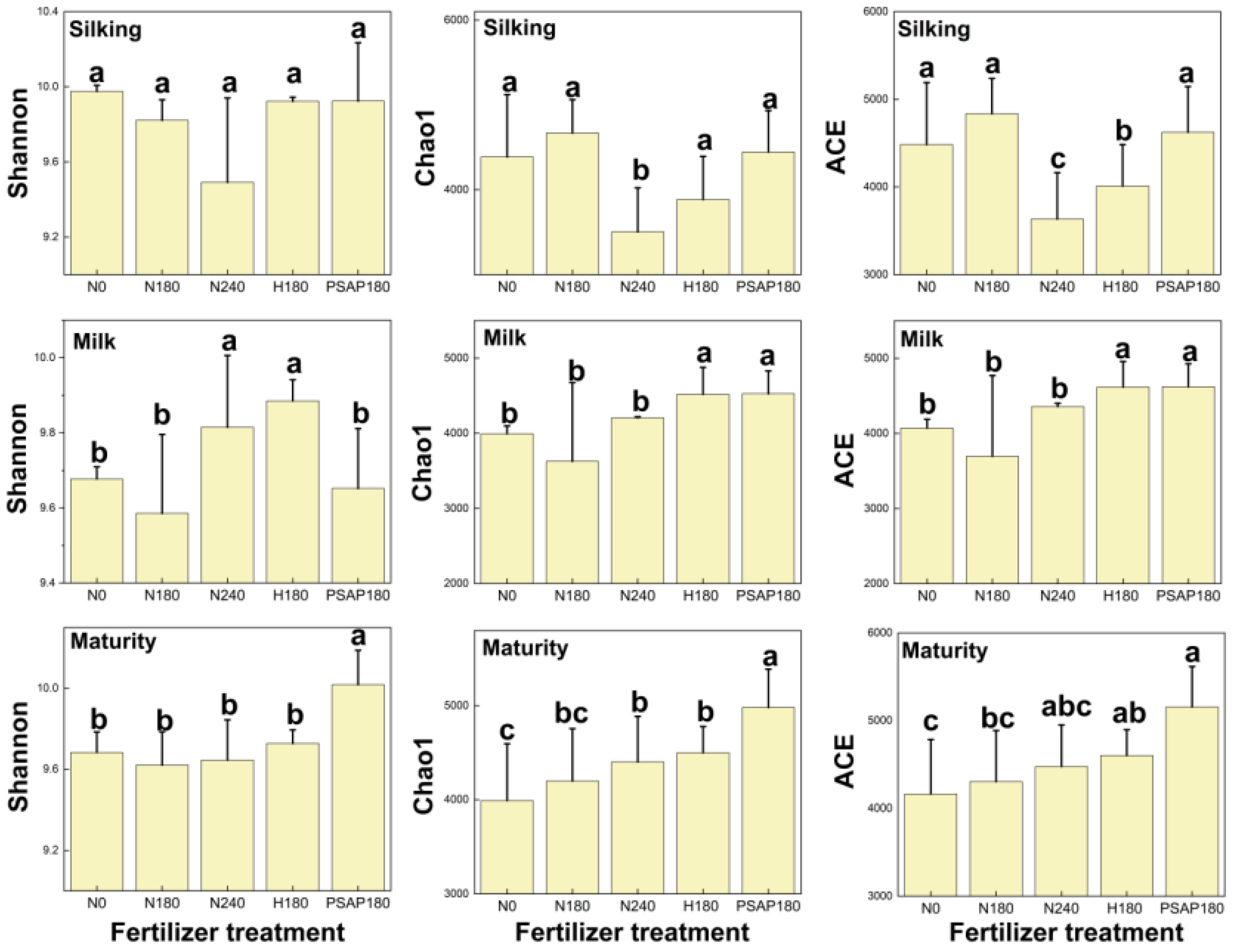
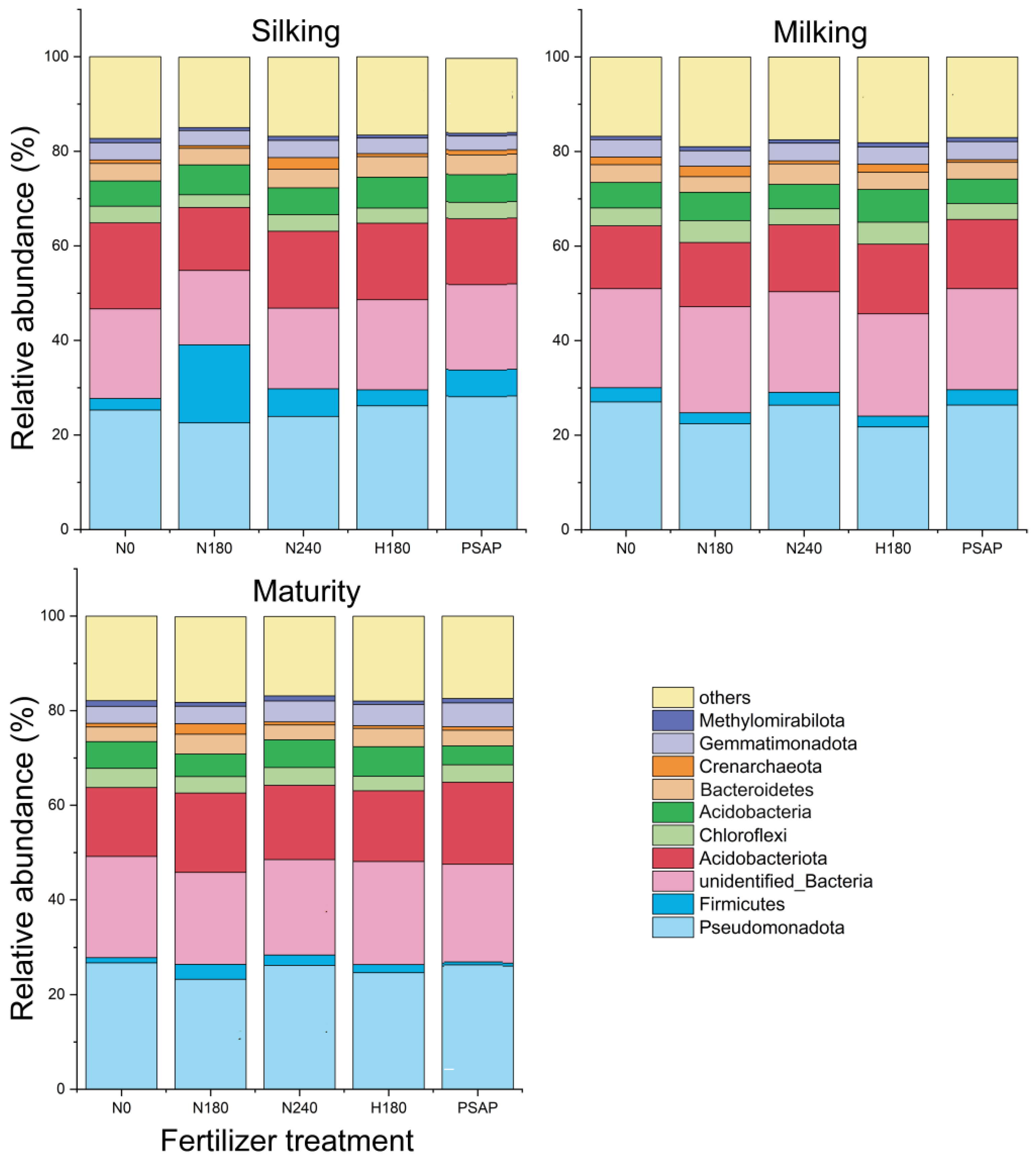
3.2. Soil Microbial α Diversity and Community Composition
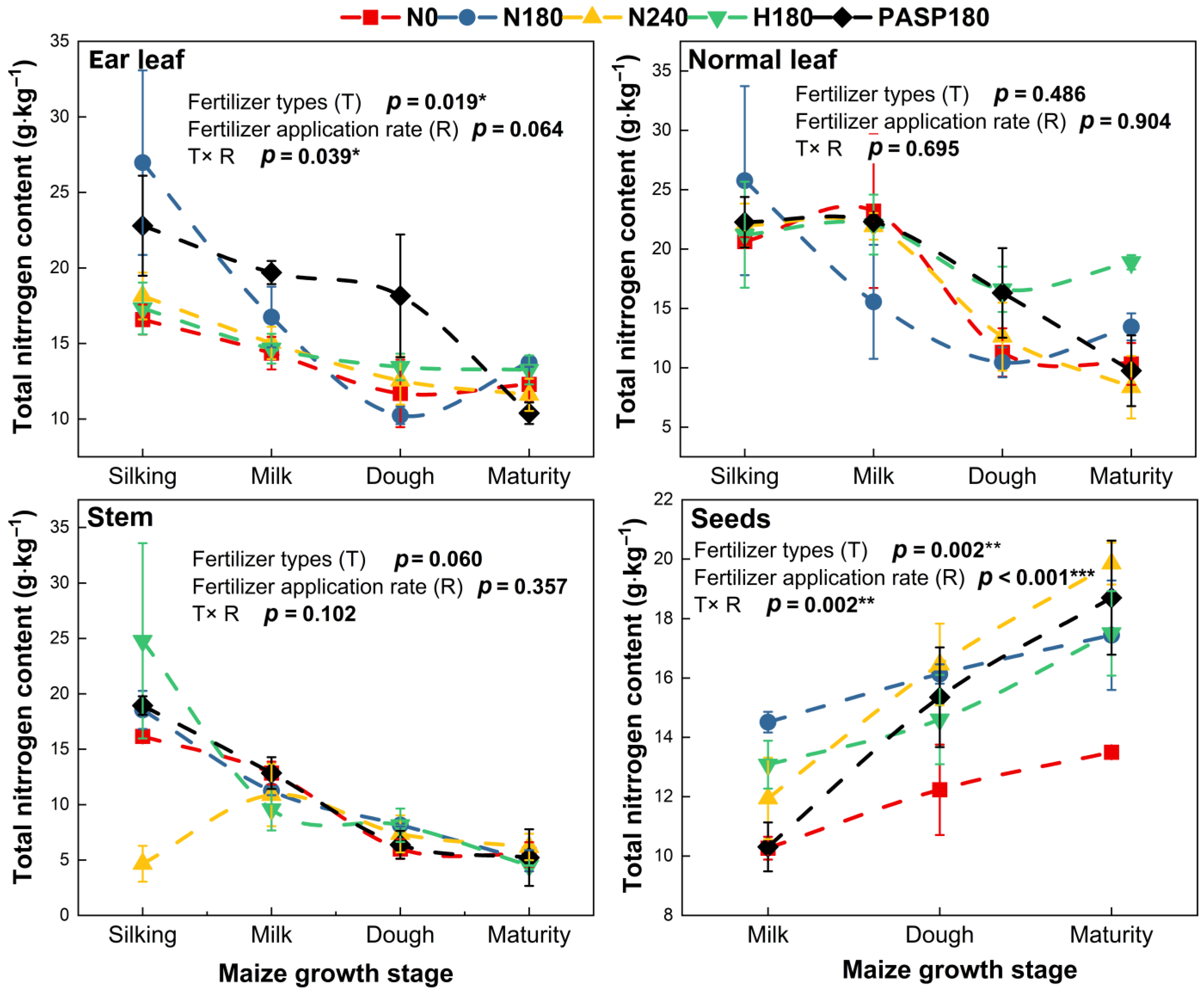
3.3. N Accumulation and Allocation
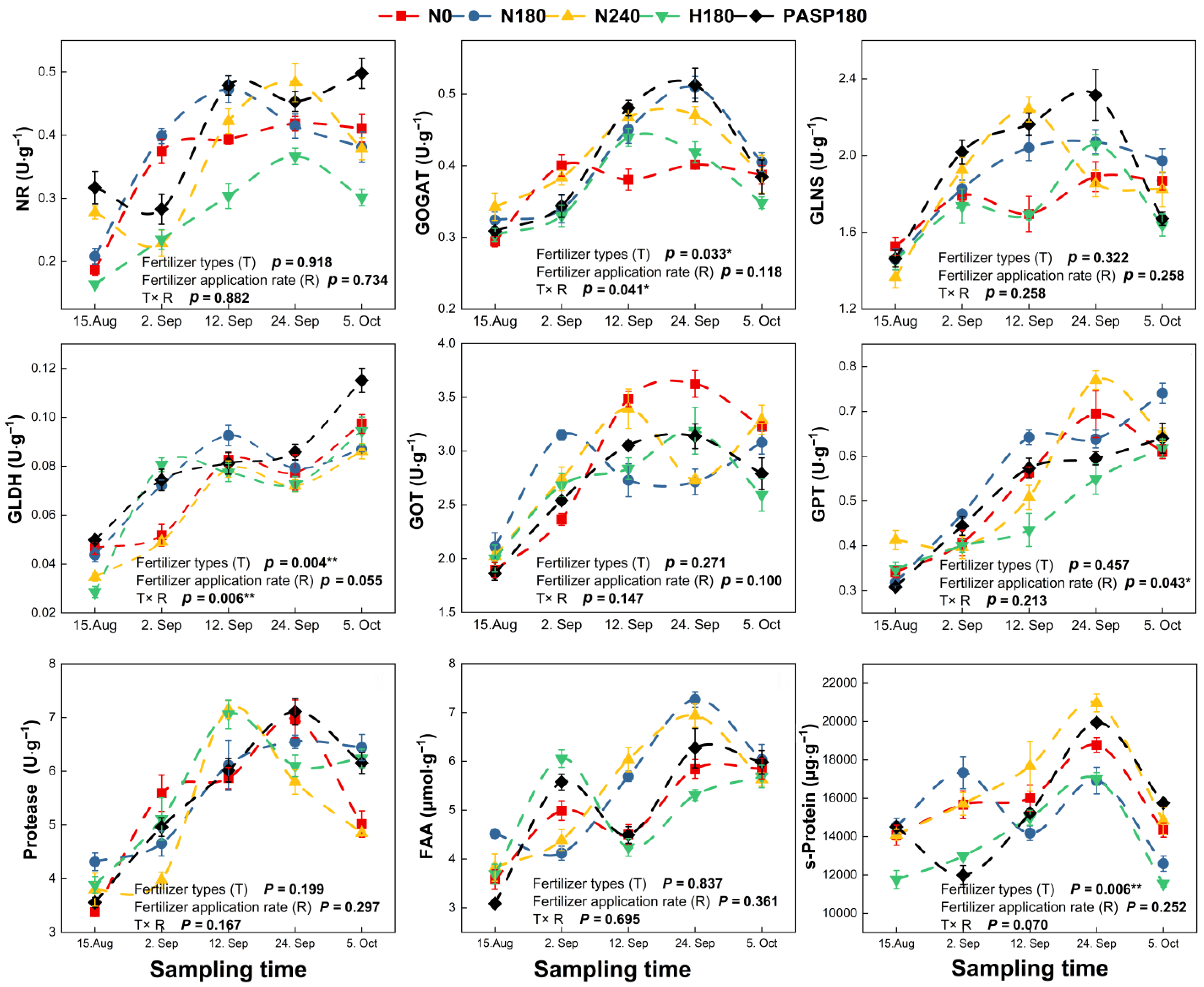
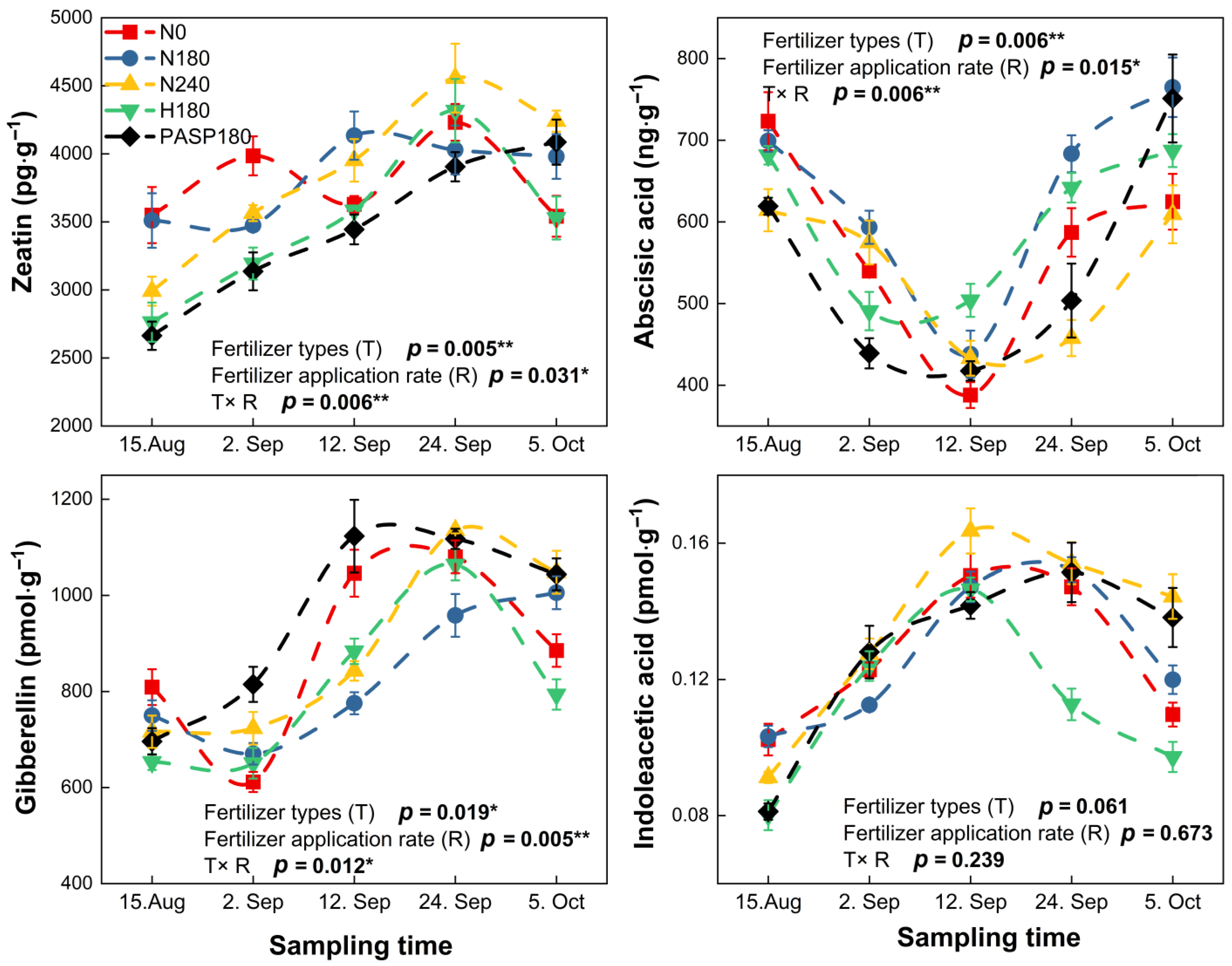
3.4. Leaf N-Converting Enzyme Activities and Phytohormones
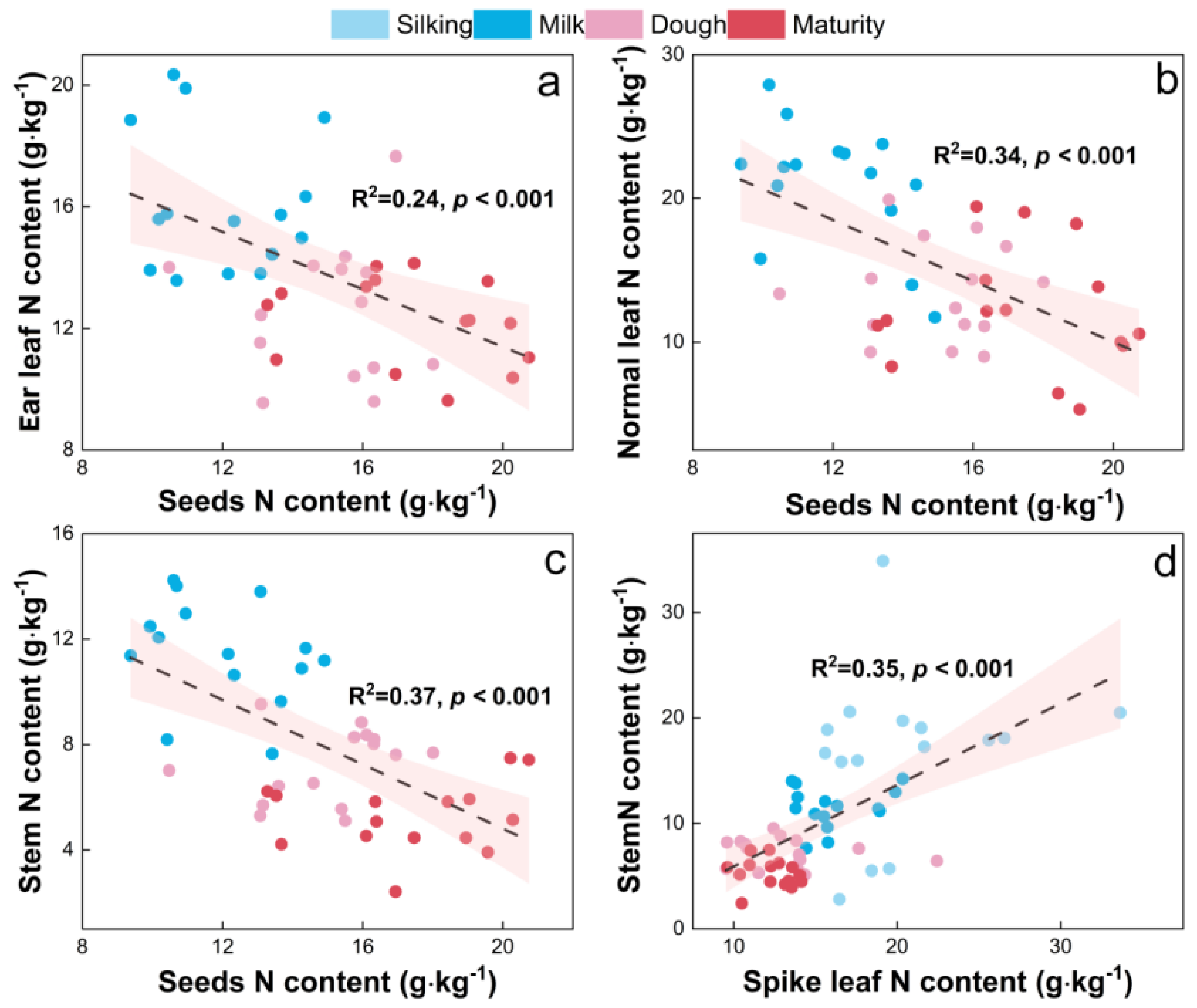
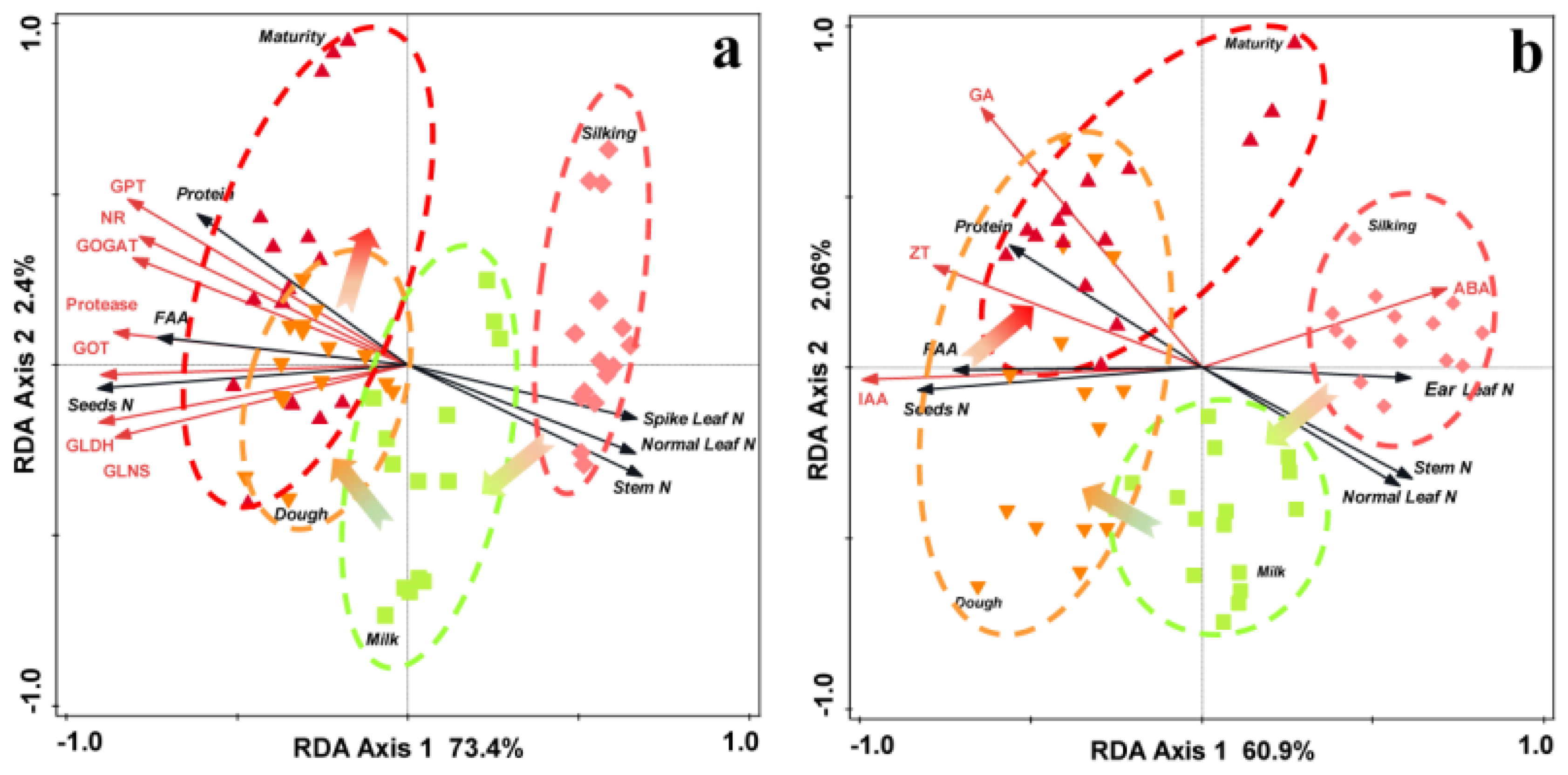
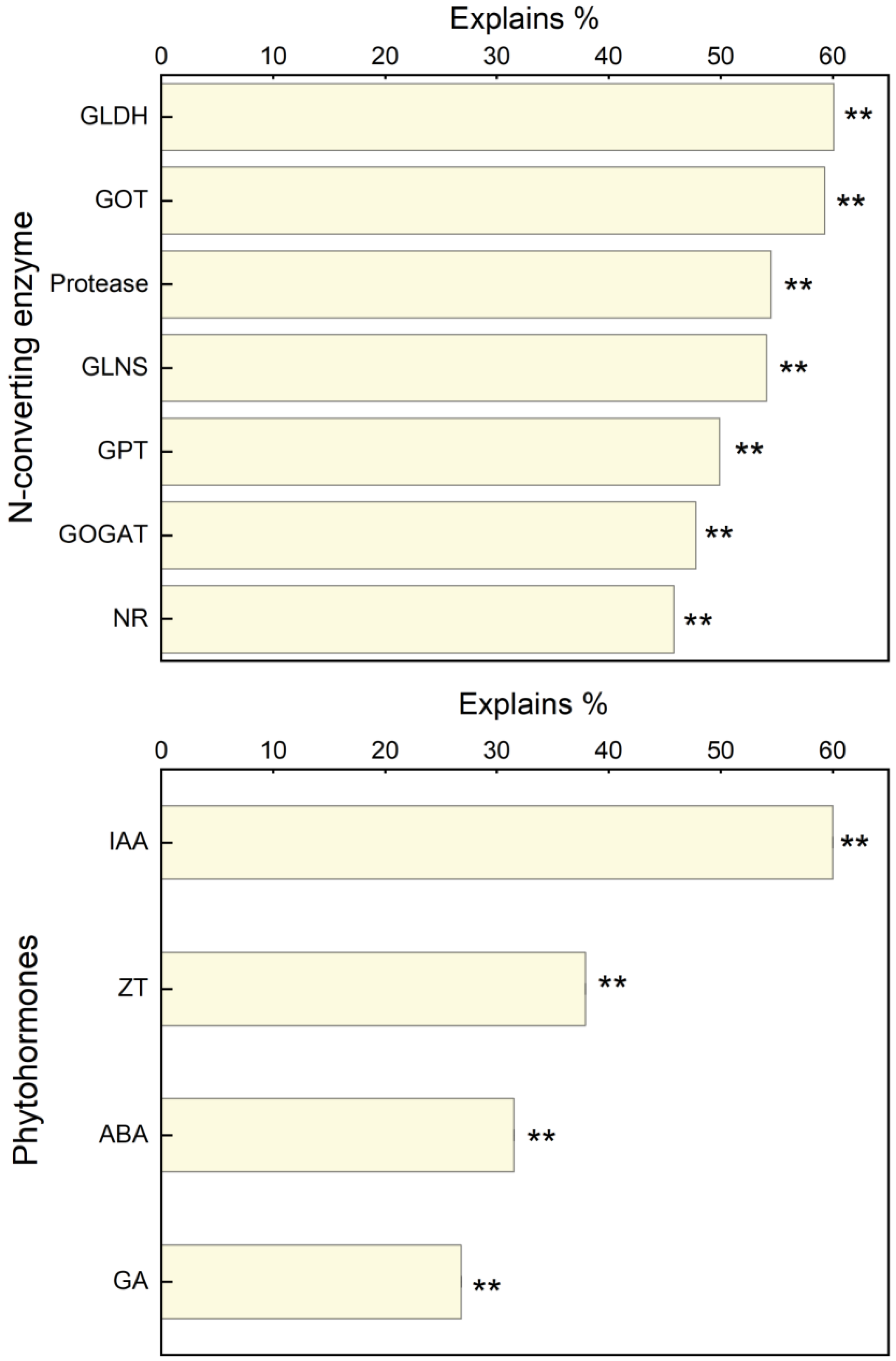
3.5. Relationship between Maize Yield and Leaf N Content and N-Converting Enzyme Activity
4. Discussion
4.1. Effect of Different Fertilizers on Seed Yield and Nitrogen Use Efficiency of Maize
4.2. Effect of Different Fertilizers on Leaf N Enzymes and Microbial Diversity and Composition
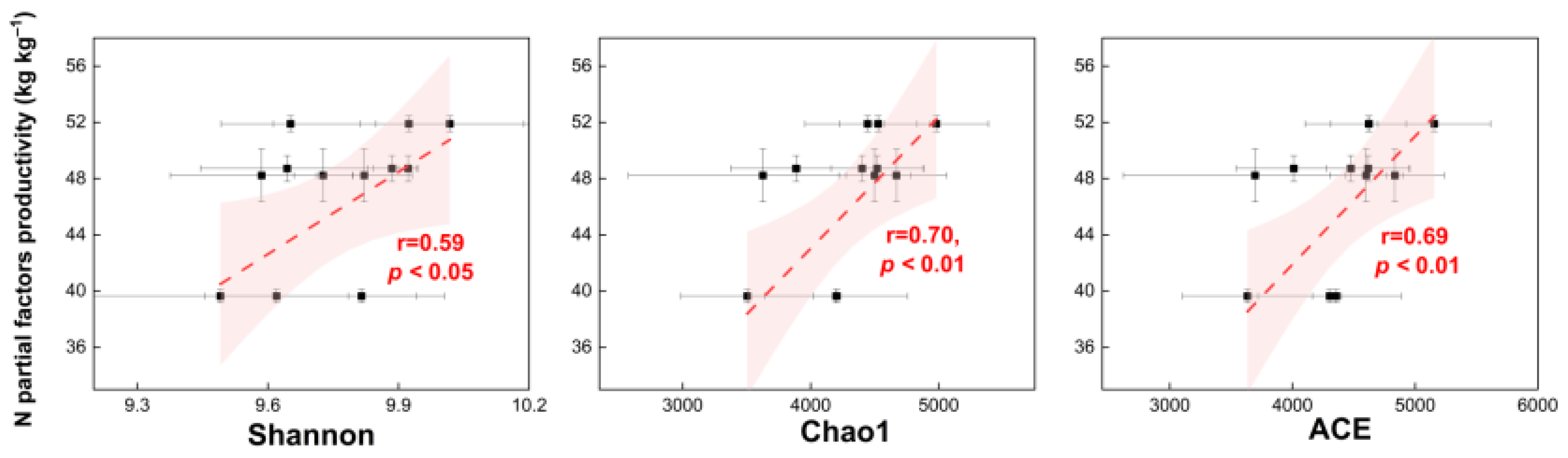
4.3. Relationship between the Leaf N Enzymes, Soil Microbial Diversity, and Seed Yield
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Guo, J.; Fan, J.; Zhang, F.; Yan, S.; Zheng, J.; Wu, Y.; Li, J.; Wang, Y.; Sun, X.; Liu, X.; et al. Blending urea and slow-release nitrogen fertilizer increases dryland maize yield and nitrogen use efficiency while mitigating ammonia volatilization. Sci. Total Environ. 2021, 790, 148058. [Google Scholar] [CrossRef] [PubMed]
- Lupwayi, N.Z.; Grant, C.A.; Soon, Y.K.; Clayton, G.W.; Bittman, S.; Malhi, S.S.; Zebarth, B.J. Soil microbial community response to controlled-release urea fertilizer under zero tillage and conventional tillage. Appl. Soil Ecol. 2010, 45, 254–261. [Google Scholar] [CrossRef]
- Zheng, W.; Zhang, M.; Liu, Z.; Zhou, H.; Lu, H.; Zhang, W.; Yang, Y.; Li, C.; Chen, B. Combining controlled-release urea and normal urea to improve the nitrogen use efficiency and yield under wheat-maize double cropping system. Field Crops Res. 2016, 197, 52–62. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms—A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Roy, K.S.; Neogi, S.; Adhya, T.K.; Rao, K.S.; Manna, M.C. Effects of rice straw and nitrogen fertilization on greenhouse gas emissions and carbon storage in tropical flooded soil planted with rice. Soil Tillage Res. 2012, 124, 119–130. [Google Scholar] [CrossRef]
- Hamer, U.; Potthast, K.; Makeschin, F. Urea fertilisation affected soil organic matter dynamics and microbial community structure in pasture soils of Southern Ecuador. Appl. Soil Ecol. 2009, 43, 226–233. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, D.; Liu, S.; Liao, Y.; Han, J. Can controlled-release urea replace the split application of normal urea in China? A meta-analysis based on crop seed yield and nitrogen use efficiency. Field Crops Res. 2022, 275, 108343. [Google Scholar] [CrossRef]
- Zhang, W.; Liang, Z.; He, X.; Wang, X.; Shi, X.; Zou, C.; Chen, X. The effects of controlled release urea on maize productivity and reactive nitrogen losses: A meta-analysis. Environ. Pollut. 2019, 246, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Grant, C.A.; Wu, R.; Selles, F.; Harker, K.N.; Clayton, G.W.; Bittman, S.; Zebarth, B.J.; Lupwayi, N.Z. Crop yield and nitrogen concentration with controlled release urea and split applications of nitrogen as compared to non-coated urea applied at seeding. Field Crops Res. 2011, 127, 170–180. [Google Scholar] [CrossRef]
- Li, G.; Zhao, B.; Dong, S.; Zhang, J.; Liu, P.; Lu, W. Controlled-release urea combining with optimal irrigation improved seed yield, nitrogen uptake, and growth of maize. Agric. Water Manag. 2020, 227, 105834. [Google Scholar] [CrossRef]
- Shaviv, A. Advances in controlled release of fertilizers. Adv. Agron. 2001, 71, 105834. [Google Scholar]
- Blackshaw, R.E.; Hao, X.; Brandt, R.N.; Clayton, G.W.; Harker, K.N.; O’Donovan, J.T.; Johnson, E.N.; Vera, C.L. Canola response to ESN and urea in a four-year no-till cropping system. Agron. J. 2011, 103, 92–99. [Google Scholar] [CrossRef]
- Deng, F.; Wang, L.; Ren, W.J.; Mei, X.F.; Li, S.X. Optimized nitrogen managements and polyaspartic acid urea improved dry matter production and yield of indica hybrid rice. Soil Tillage Res. 2015, 145, 1–9. [Google Scholar] [CrossRef]
- Deng, F.; Wang, L.; Ren, W.J.; Mei, X.F. Enhancing nitrogen utilization and soil nitrogen balance in paddy fields by optimizing nitrogen management and using polyaspartic acid urea. Field Crops Res. 2014, 169, 30–38. [Google Scholar] [CrossRef]
- Hu, M.; Dou, Q.; Cui, X.; Lou, Y.; Zhuge, Y. Polyaspartic acid mediates the absorption and translocation of mineral elements in tomato seedlings under combined copper and cadmium stress. J. Integ. Agric. 2019, 18, 1130–1137. [Google Scholar] [CrossRef]
- Liu, W.L.; Zhao, J.B.; Zeng, H.; Wan, C.X. Effect of polyaspartic acid on soil water storage, soil microbial diversity, cotton yield and fiber quality. Arch. Microbiol. 2022, 205, 35. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Wang, L.; Mei, X.F.; Li, S.X.; Pu, S.L.; Li, Q.P.; Ren, W.J. Polyaspartic acid (PASP)-urea and optimised nitrogen management increase the grain nitrogen concentration of rice. Sci. Rep. 2019, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yang, J.; Wang, H.; Chen, Y.; Ren, J.; Lin, X.; Zhao, J.; Chen, B.; Liu, H. Effects of molecular weight of polyaspartic acid on nitrogen use efficiency and crop yield. J. Sci. Food Agric. 2022, 102, 7343–7352. [Google Scholar] [CrossRef]
- Shen, Y.; Lin, H.; Gao, W.; Li, M. The effects of humic acid urea and polyaspartic acid urea on reducing nitrogen loss compared with urea. J. Sci. Food Agric. 2020, 100, 4425–4432. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef] [PubMed]
- Bernard, S.M.; Habash, D.Z. The importance of cytosolic glutamine synthetase in nitrogen assimilation and recycling. New Phytol. 2009, 182, 608–620. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef]
- Yadav, A.N.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.; Dikilitas, M.; Abdel-Azeem, A.M.; Ahluwalia, A.S.; Saxena, A.K. Biodiversity, and biotechnological contribution of beneficial soil microbiomes for nutrient cycling, plant growth improvement and nutrient uptake. Biocatal. Agric. Biotechno. 2021, 33, 102009. [Google Scholar] [CrossRef]
- Hu, Y.; Zheng, Q.; Zhang, S.; Noll, L.; Wanek, W. Significant release and microbial utilization of amino sugars and D-amino acid enantiomers from microbial cell wall decomposition in soils. Soil Biol. Biochem. 2018, 1123, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zheng, Q.; Wanek, W. Flux analysis of free amino sugars and amino acids in soils by isotope tracing with a novel liquid chromatography/high resolution mass spectrometry platform. Anal. Chem. 2017, 89, 9192–9200. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Hu, Y.; Zhang, S.; Noll, L.; Böckle, T.; Dietrich, M.; Herbold, C.W.; Eichorst, S.A.; Woebken, D.; Richter, A.; et al. Soil multifunctionality is affected by the soil environment and by microbial community composition and diversity. Soil Biol. Biochem. 2019, 136, 107521. [Google Scholar] [CrossRef]
- Liang, C.; Amelung, W.; Lehmann, J.; Kästner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Global Change Biol. 2019, 25, 3578–3590. [Google Scholar] [CrossRef]
- Warren, C. What are the products of enzymatic cleavage of organic N? Soil Biol. Biochem. 2021, 154, 108152. [Google Scholar] [CrossRef]
- Chen, X.; Xia, Y.; Rui, Y.; Ning, Z.; Hu, Y.; Tang, H.; He, H.; Li, H.; Kuzyakov, Y.; Ge, T.; et al. Microbial carbon use efficiency, biomass turnover, and necromass accumulation in paddy soil depending on fertilization. Agric. Ecosyst. Environ. 2020, 292, 106816. [Google Scholar] [CrossRef]
- Wang, B.; Huang, Y.; Li, N.; Yao, H.; Yang, E.; Soromotin, A.V.; Kuzyakov, Y.; Cheptsov, V.; Yang, Y.; An, S. Initial soil formation by biocrusts: Nitrogen demand and clay protection control microbial necromass accrual and recycling. Soil Biol. Biochem. 2022, 167, 108607. [Google Scholar] [CrossRef]
- Wang, B.; An, S.; Liang, C.; Liu, Y.; Kuzyakov, Y. Microbial necromass as the source of soil organic carbon in global ecosystems. Soil Biol. Biochem. 2021, 162, 108422. [Google Scholar] [CrossRef]
- He, H.; Zhang, W.; Zhang, X.; Xie, H.; Zhuang, J. Temporal responses of soil microorganisms to substrate addition as indicated by amino sugar differentiation. Soil Biol. Biochem. 2011, 43, 1155–1161. [Google Scholar] [CrossRef]
- Zhang, W.; Cui, Y.; Lu, X.; Bai, E.; He, H.; Xie, H.; Liang, C.; Zhang, X. High nitrogen deposition decreases the contribution of fungal residues to soil carbon pools in a tropical forest ecosystem. Soil Biol. Biochem. 2016, 97, 211–214. [Google Scholar] [CrossRef]
- Bremner, J.M. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Wu, L.; Jiang, H.; Tao, N. Colorimetric determination of glu tamate oxaloacetate transaminase and glutamate pyruvate trans aminase activity in plants and its application. Chin. J. Rice Sci. 1998, 29, 136–138. (In Chinese) [Google Scholar]
- Yuan, M.M.; Guo, X.; Wu, L.; Zhang, Y.; Xiao, N.; Ning, D.; Shi, Z.; Zhou, X.; Wu Liyou Yang, Y.; Tiedje, J.M.; et al. Climate warming enhances microbial network complexity and stability. Nat. Rev. Earth Environ. 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. Qiime allows analysis of high-throughput community sequencing data intensity normalization improves color calling in solid sequencing. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Reyon, D.; Tsai, S.Q.; Khgayter, C.; Foden, J.A.; Sander, J.D.; Joung, J.K. FLASH assembly of TALENs for high-throughput genome editing. Nat. Biotechnol. 2012, 30, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Gao, Y.; Jia, X.; Wang, M.; Ding, J.; Cheng, L.; Bao, F.; Wu, B. Network analysis reveals the strengthening of microbial interaction in biological soil crust development in the Mu Us Sandy Land, northwestern China. Soil Biol. Biochem. 2020, 14, 10778. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. Uchime improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- De Wit, C.T. Resource use efficiency in agriculture. Agric. Syst. 1992, 40, 125–151. [Google Scholar] [CrossRef]
- Li, R.; Gao, Y.; Chen, Q.; Li, Z.; Gao, F.; Meng, Q.; Li, T.; Liu, A.; Wang, Q.; Wu, L.; et al. Blended controlled-release nitrogen fertilizer with straw returning improved soil nitrogen availability, soil microbial community, and root morphology of wheat. Soil Tillage Res. 2021, 212, 105045. [Google Scholar] [CrossRef]
- Tomida, M.; Nakato, T.; Matsunami, S.; Kakuchi, T. Convenient synthesis of high molecular weight poly(succinimide) by acid-catalysed polycondensation of L-aspartic acid. Polymer 1997, 38, 4733–4736. [Google Scholar] [CrossRef]
- Dubois, F.; Tercé-Laforgue, T.; Gonzalez-Moro, M.B.; Estavillo, J.M.; Sangwan, R.; Gallais, A.; Hirel, B. Glutamate dehydrogenase in plants: Is there a new story for an old enzyme? Plant Physiol. Biochem. 2003, 41, 565–576. [Google Scholar] [CrossRef]
- Osaki, M.; Iyoda, M.; Tadano, T.; Iyoda, M. Ontogenetic changes in the contents of ribulose-1, 5-bisphosphate carboxylase/oxygenase, phosphoenolpyruvate carboxylase, and chlorophyll in individual leaves of maize. Soil Sci. Plant Nutr. 1995, 41, 285–293. [Google Scholar] [CrossRef]
- De Vries, F.T.; Shade, A. Controls on soil microbial community stability under climate change. Front. Microbiol. 2013, 4, 1–16. [Google Scholar] [CrossRef]
- Jiao, S.; Lu, Y.; Wei, G. Soil multitrophic network complexity enhances the link between biodiversity and multifunctionality in agricultural systems. Global Change Biol. 2021, 28, 140–153. [Google Scholar] [CrossRef]
- Fan, K.; Delgado-Baquerizo, M.; Guo, X.; Wang, D.; Zhu, Y.G.; Chu, H. Microbial resistance promotes plant production in a four-decade nutrient fertilization experiment. Soil Biol. Biochem. 2020, 141, 107679. [Google Scholar] [CrossRef]
- Fan, K.; Delgado-Baquerizo, M.; Guo, X.; Wang, D.; Zhu, Y.G.; Chu, H. Biodiversity of key-stone phylotypes determines crop production in a 4-decade fertilization experiment. Isme J. 2021, 15, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Li, B.B.; Roley, S.S.; Duncan, D.S.; Guo, J.; Quensen, J.F.; Yu, H.Q.; Tiedje, J.M. Long-term excess nitrogen fertilizer increases sensitivity of soil microbial community to seasonal change revealed by ecological network and metagenome analyses. Soil Biol. Biochem. 2021, 160, 108349. [Google Scholar] [CrossRef]
- Inselsbacher, E.; Hinko-Najera Umana, N.; Stange, F.C.; Gorfer, M.; Schüller, E.; Ripka, K.; Zechmeister-Boltenstern, S.; Hood-Novotny, R.; Strauss, J.; Wanek, W. Short-term competition between crop plants and soil microbes for inorganic N fertilizer. Soil Biol. Biochem. 2010, 42, 360–372. [Google Scholar] [CrossRef]
- Sun, R.; Wang, F.; Hu, C.; Liu, B. Metagenomics reveals taxon-specific responses of the nitrogen-cycling microbial community to long-term nitrogen fertilization. Soil Biol. Biochem. 2021, 156, 108214. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Treatment | Grain Yield (kg hm−2) | N-Agronomic Efficiency (kg kg−1) | N-Partial Factors (kg kg−1) |
|---|---|---|---|---|
| 2019 | N0 | 8679.5 c | - | |
| N180 | 10,364.8 b | 8.25 b | 57.58 b | |
| N240 | 10,956.3 b | 12.65 a | 58.09 b | |
| H180 | 10,260.3 b | 7.67 b | 55.89 b | |
| PASP180 | 11,332.1 a | 14.74 a | 62.96 a | |
| 2020 | N0 | 8549.4 d | - | |
| N180 | 10,170.7 c | 9.01 c | 58.17 b | |
| N240 | 11,159.0 ab | 14.50 b | 61.99 b | |
| H180 | 10,112.3 c | 8.13 c | 55.62 b | |
| PASP180 | 12,627.2 a | 22.65 a | 70.15 a | |
| Source of variation | ||||
| Year | NS | NS | NS | |
| N | ** | * | * | |
| Year × N | NS | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, P.; Peng, Y.; Li, X.; Wang, R.; Tao, P.; Zhang, Y. Polyaspartic Acid Urea Increased Maize Yield by Enhancing Leaf N Turnover Efficiency and Soil Microbial Diversity. Agronomy 2024, 14, 796. https://doi.org/10.3390/agronomy14040796
Ji P, Peng Y, Li X, Wang R, Tao P, Zhang Y. Polyaspartic Acid Urea Increased Maize Yield by Enhancing Leaf N Turnover Efficiency and Soil Microbial Diversity. Agronomy. 2024; 14(4):796. https://doi.org/10.3390/agronomy14040796
Chicago/Turabian StyleJi, Pengtao, Yujuan Peng, Xiangling Li, Rui Wang, Peijun Tao, and Yuechen Zhang. 2024. "Polyaspartic Acid Urea Increased Maize Yield by Enhancing Leaf N Turnover Efficiency and Soil Microbial Diversity" Agronomy 14, no. 4: 796. https://doi.org/10.3390/agronomy14040796
APA StyleJi, P., Peng, Y., Li, X., Wang, R., Tao, P., & Zhang, Y. (2024). Polyaspartic Acid Urea Increased Maize Yield by Enhancing Leaf N Turnover Efficiency and Soil Microbial Diversity. Agronomy, 14(4), 796. https://doi.org/10.3390/agronomy14040796