
Conversion of Natural Soil to Paddy Promotes Soil Organic Matter Degradation in Small-Particle Fractions: δ13C and Lipid Biomarker Evidence

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Soil Samples Collection and Pretreatment
2.2. Soil Particle-Size Fractionation
2.3. Extraction and Detection of Lipid Biomarkers in Soils
2.4. Stable Carbon Isotopic Analyses in All Soil Particle-Size Fractions
2.5. Calculation and Date Analysis
3. Results and Discussion
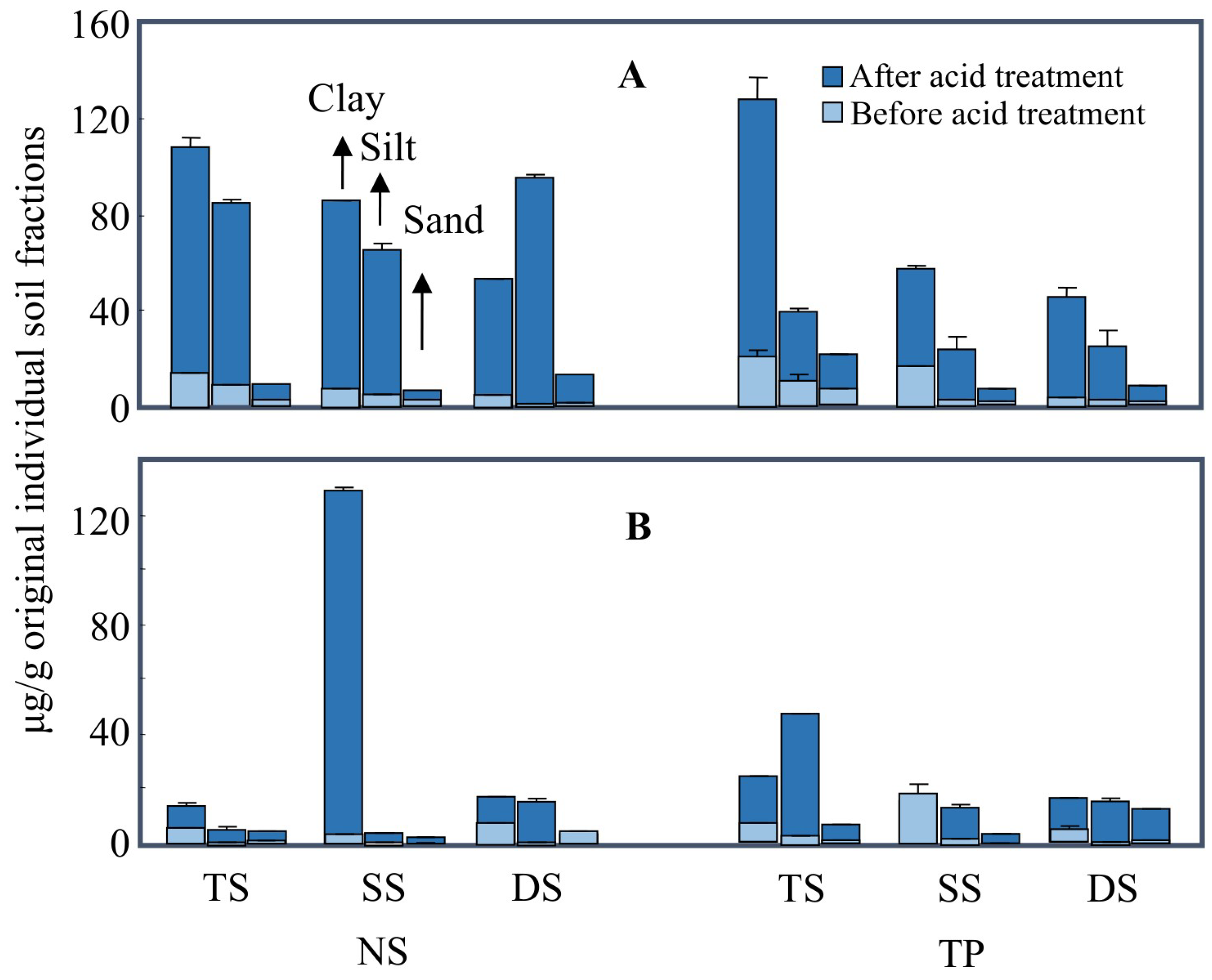
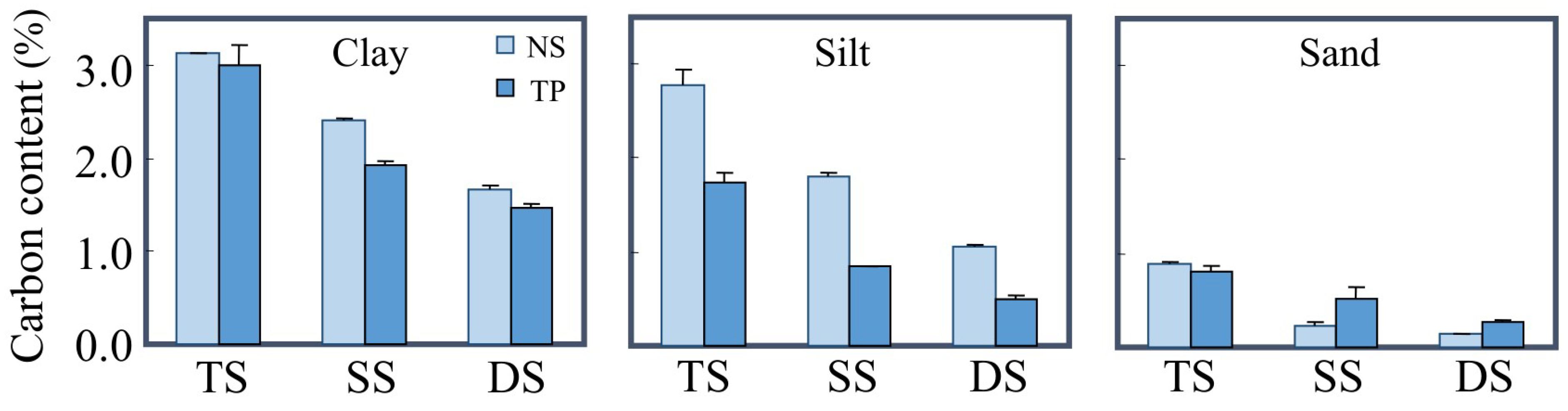
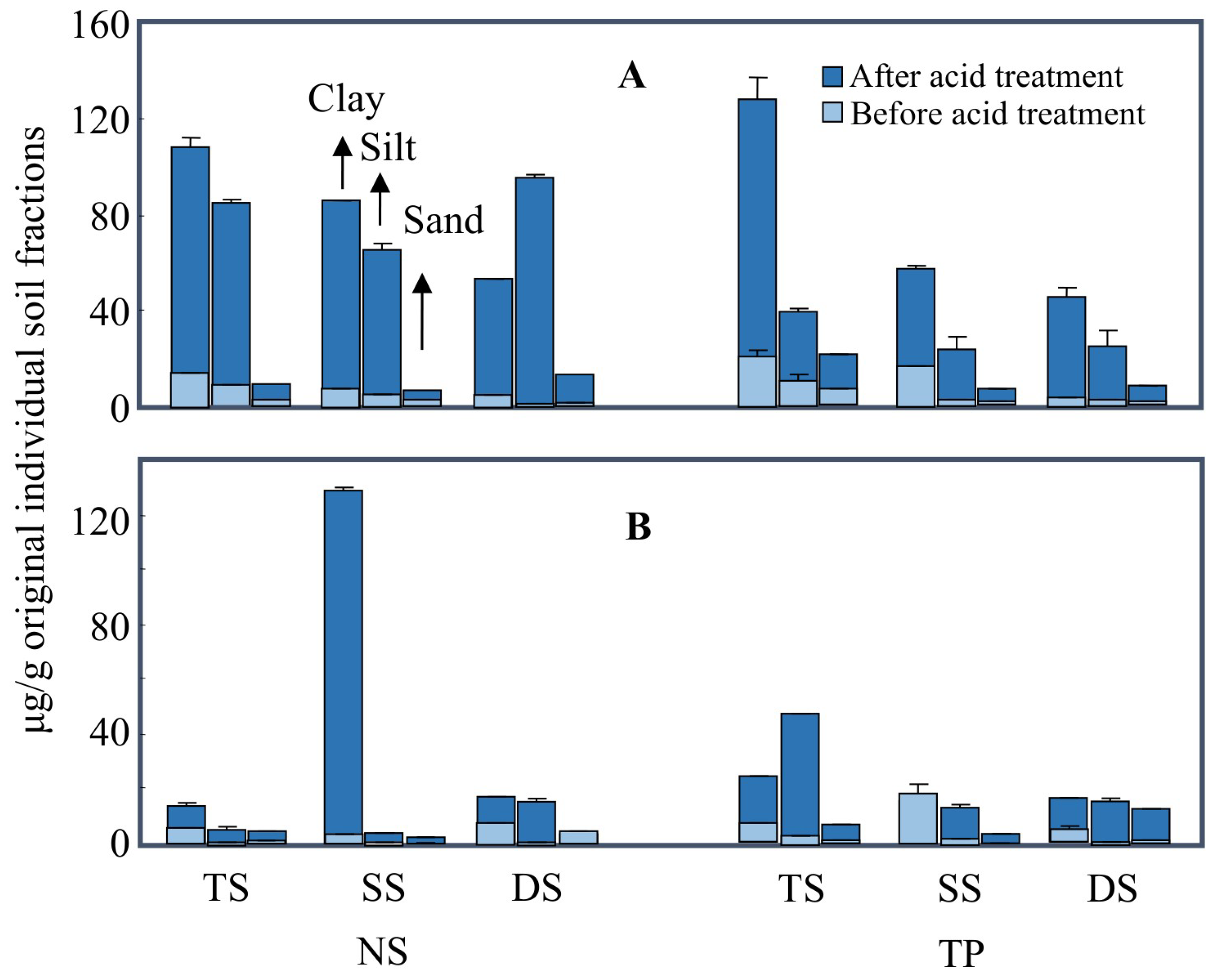
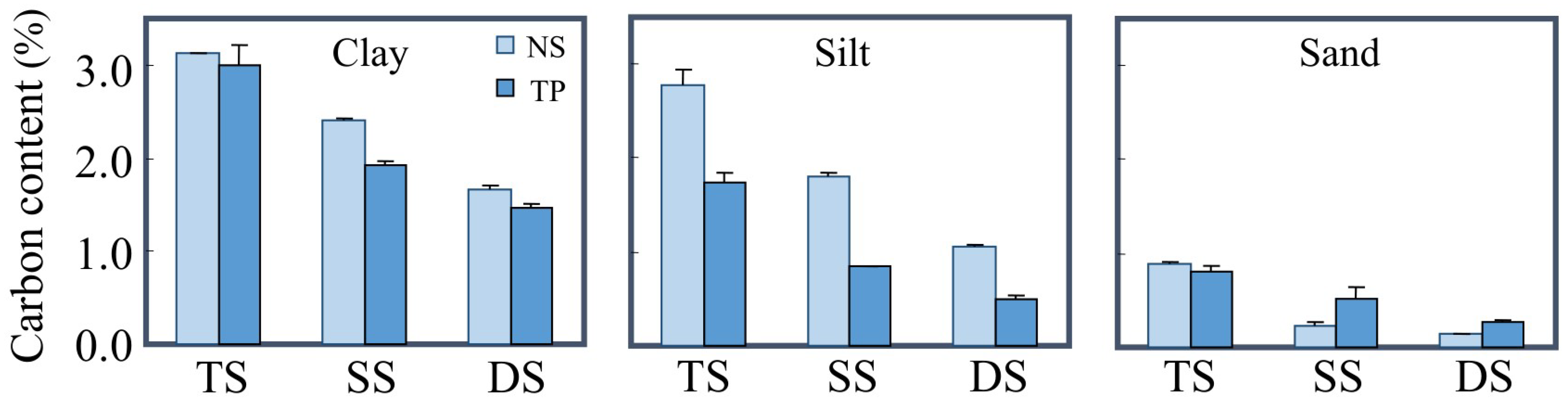
3.1. The Protection of Lipids by Mineral Particles
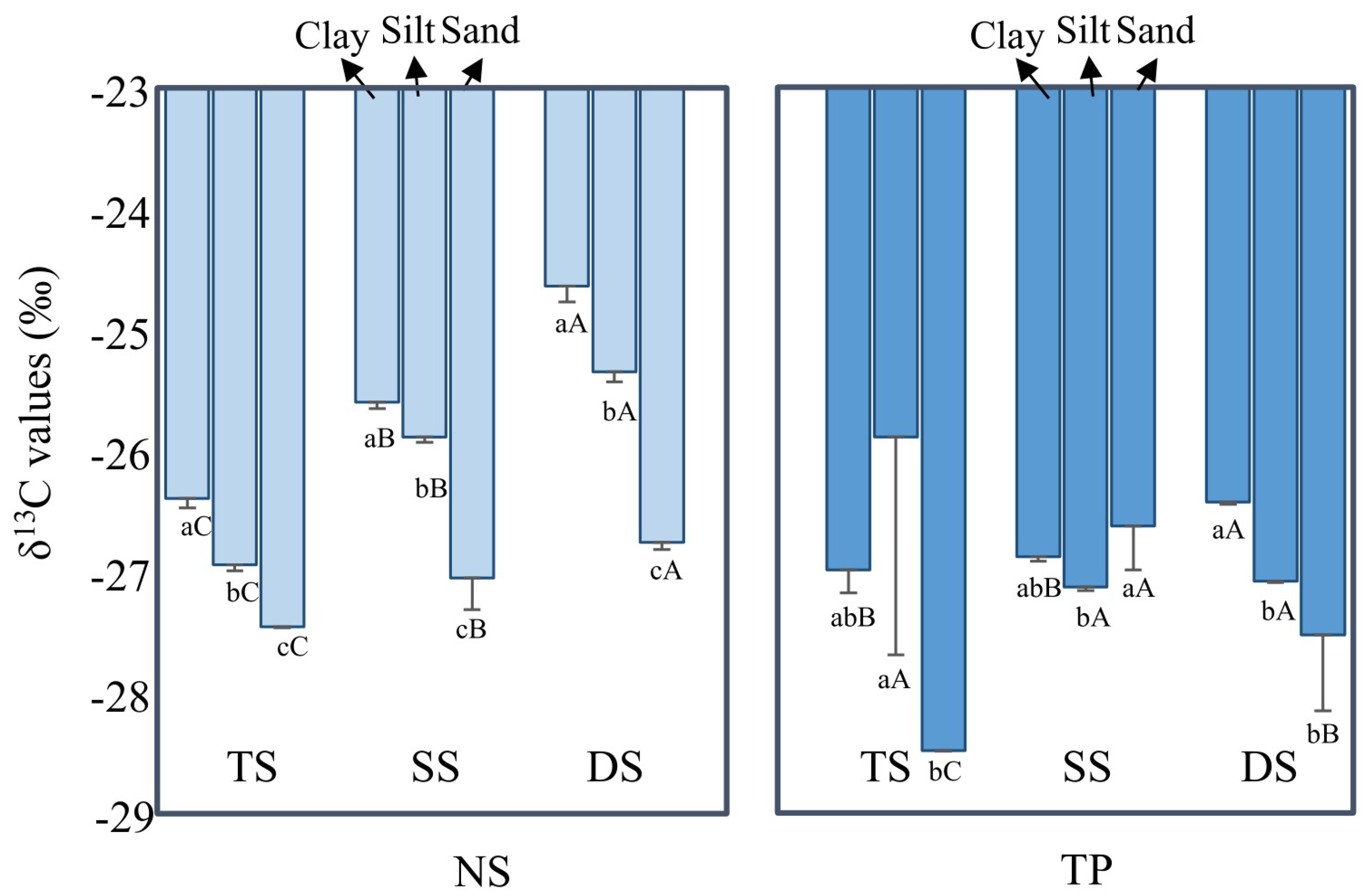
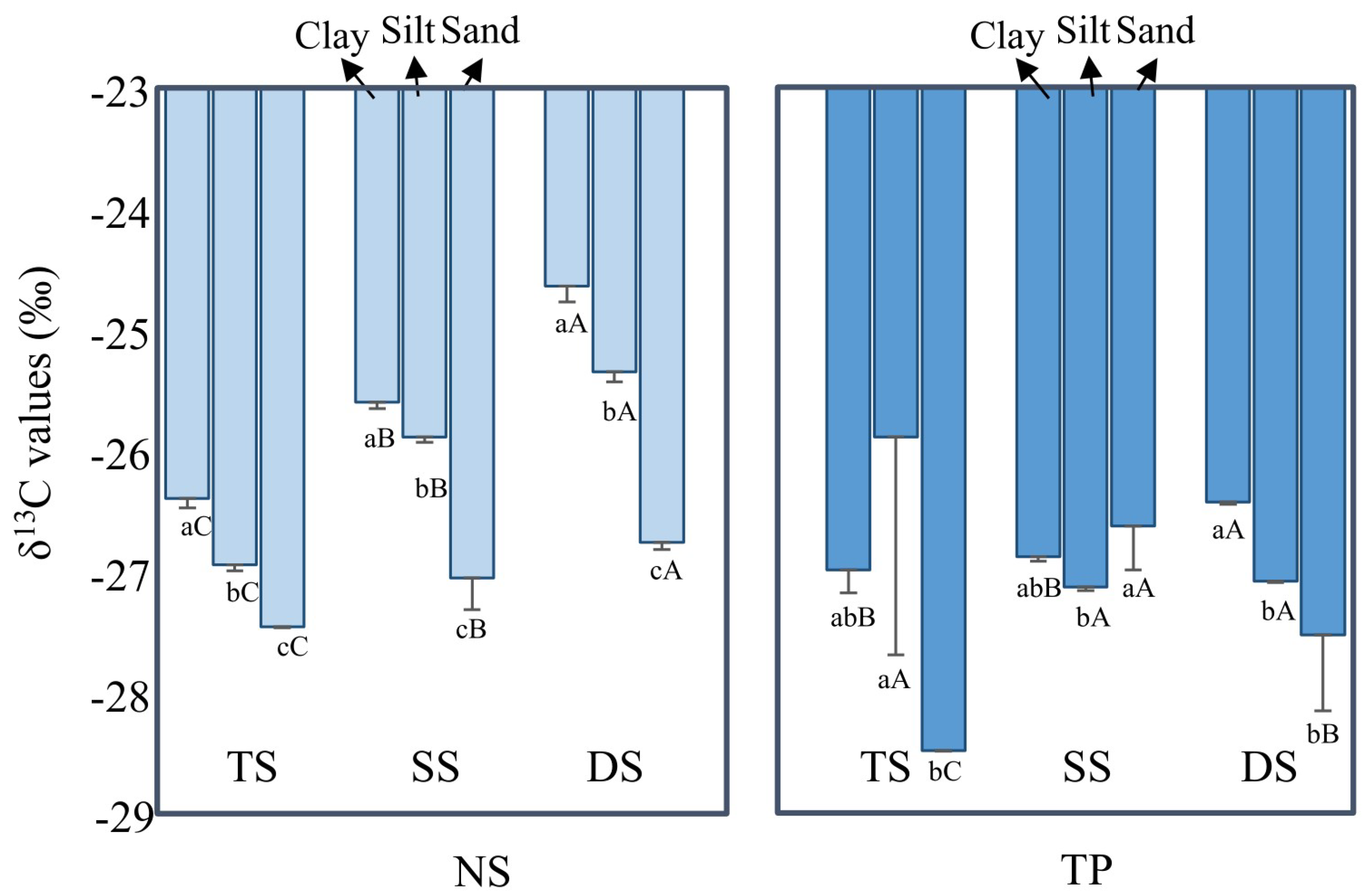
3.2. Fresh SOM Input after Cultivation, as Suggested by Bulk δ13C Values
3.2.1. δ13C Enrichment in Small Particles
3.2.2. δ13C Enrichment with Soil Depth
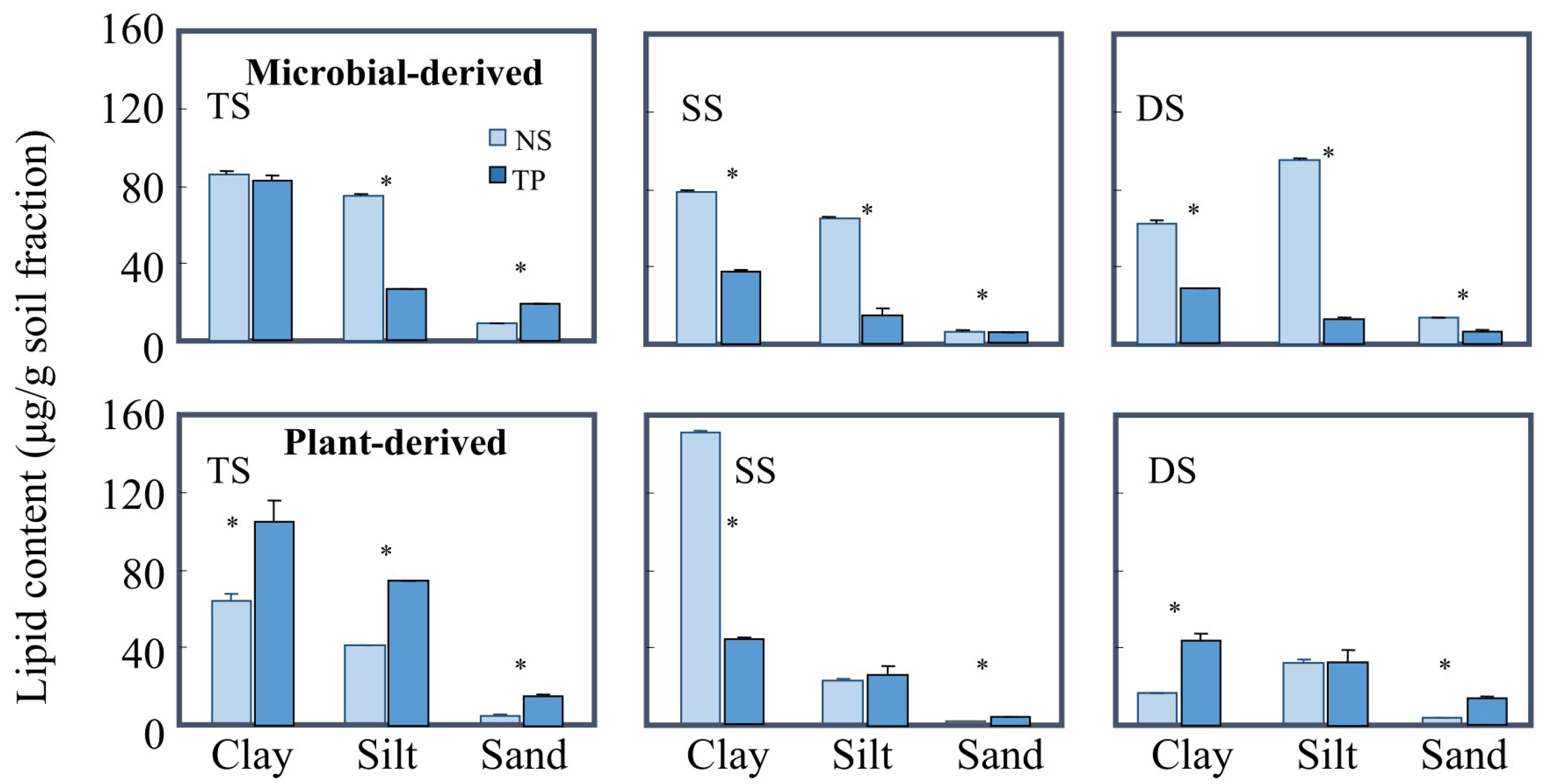
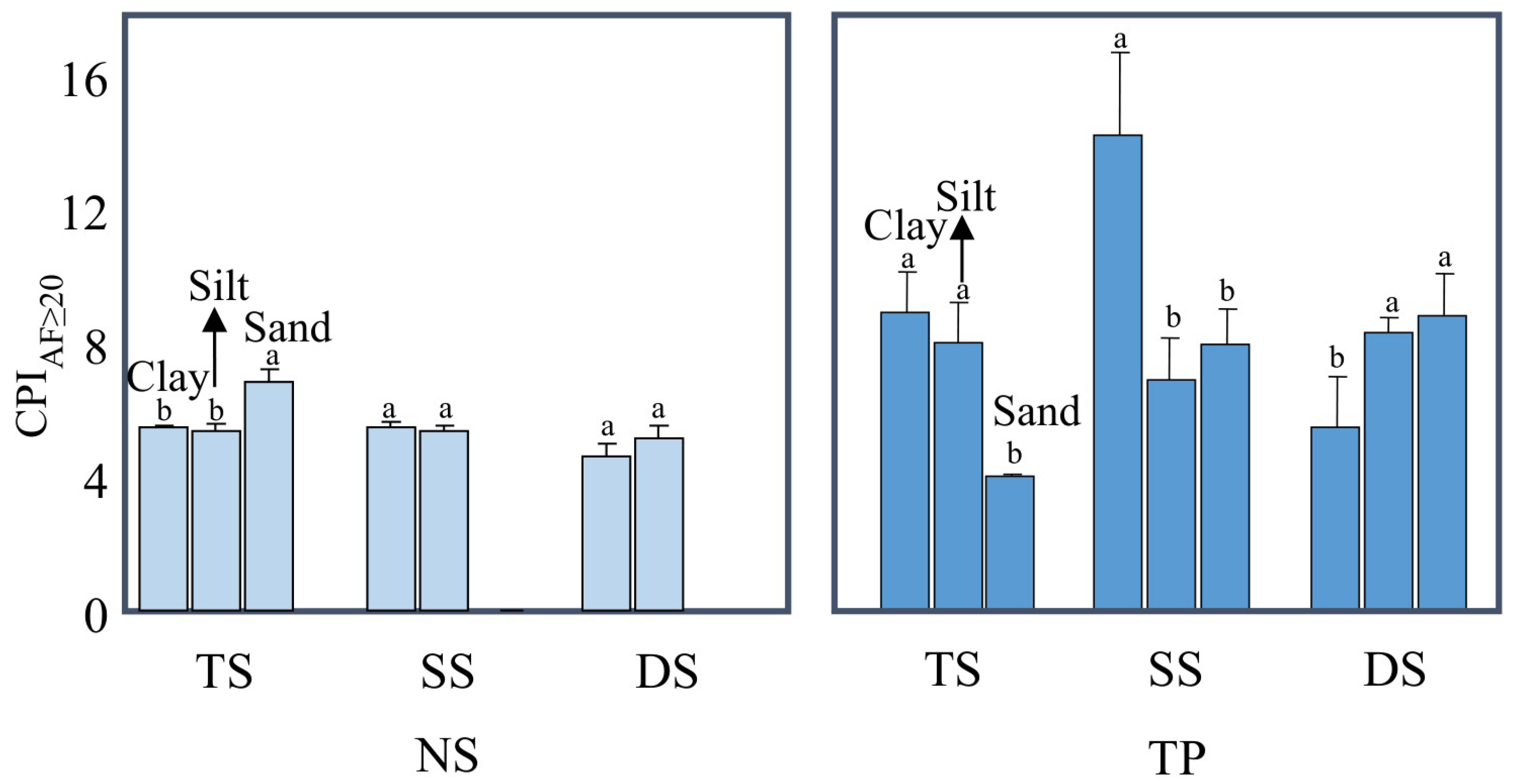
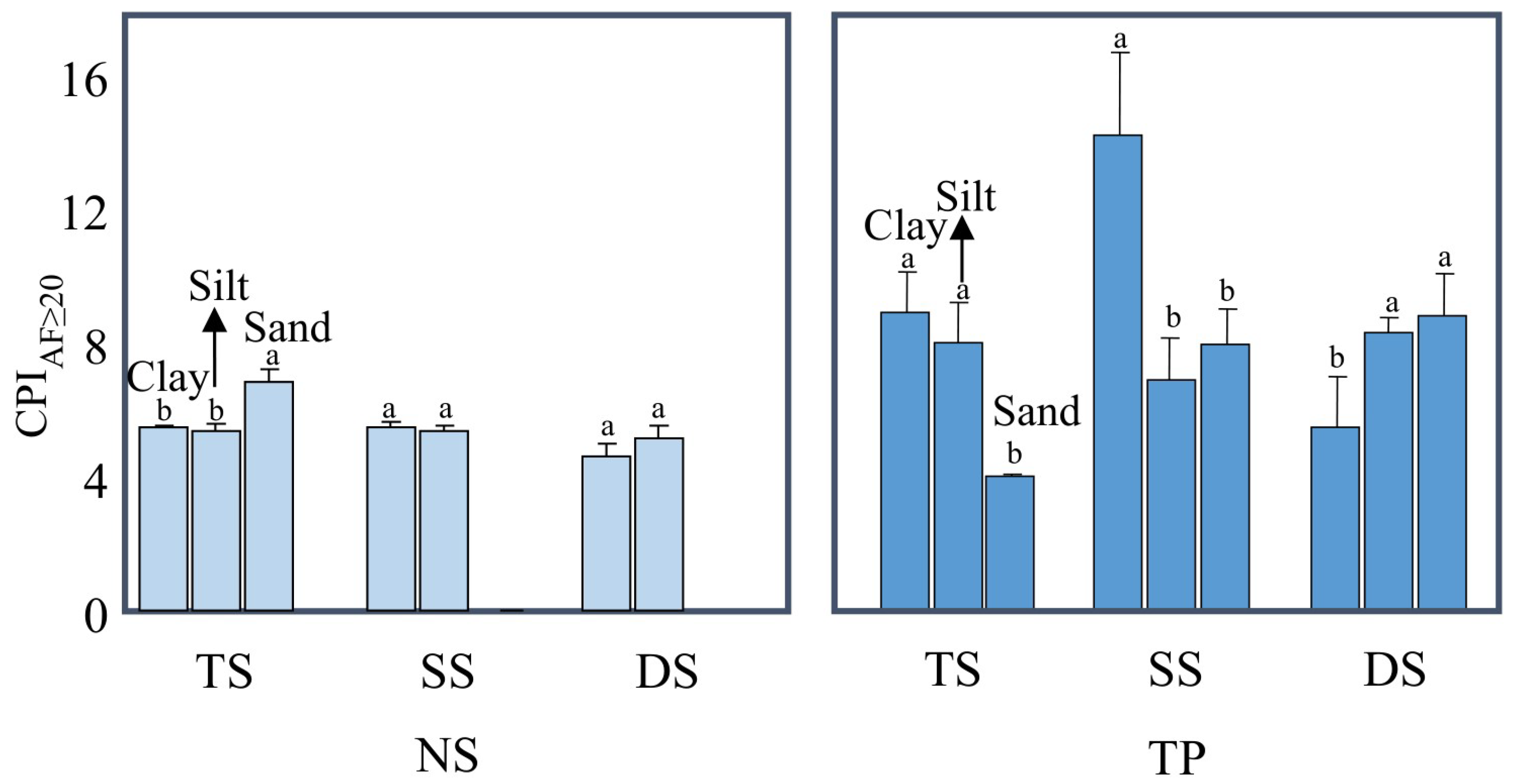
3.3. The Degradation of Mineral-Protected Lipids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Lorenz, K.; Lal, R.; Preston, C.M.; Nierop, K.G.J. Strengthening the soil organic carbon pool by increasing contributions from recalcitrant aliphatic bio(macro)molecules. Geoderma 2007, 142, 1–10. [Google Scholar] [CrossRef]
- Dungait, J.A.J.; Hopkins, D.W.; Gregory, A.S.; Whitmore, A.P. Soil organic matter turnover is governed by accessibility not recalcitrance. Global Chang. Biol. 2012, 18, 1781–1796. [Google Scholar] [CrossRef]
- Angst, G.; Mueller, K.E.; Kogel-Knabner, I.; Freeman, K.H.; Mueller, C.W. Aggregation controls the stability of lignin and lipids in clay-sized particulate and mineral associated organic matter. Biogeochemistry 2017, 132, 307–324. [Google Scholar] [CrossRef]
- Balesdent, J.; Chenu, C.; Balabane, M. Relationship of soil organic matter dynamics to physical protection and tillage. Soil Till. Res. 2000, 53, 215–230. [Google Scholar] [CrossRef]
- Christensen, B.T. Physical fractionation of soil and structural and functional complexity in organic matter turnover. Eur. J. Soil Sci. 2010, 52, 345–353. [Google Scholar] [CrossRef]
- Chen, S.L.; Hong, H.L.; Huang, X.Y.; Fang, Q.; Yin, K.; Wang, C.W.; Zhang, Y.M.; Cheng, L.L.; Algeo, T.J. The role of organo-clay associations in limiting organic matter decay: Insights from the Dajiuhu peat soil, central China. Geoderma 2018, 320, 149–160. [Google Scholar] [CrossRef]
- Clemente, J.S.; Simpson, A.J.; Simpson, M.J. Association of specific organic matter compounds in size fractions of soils under different environmental controls. Org. Geochem. 2012, 42, 1169–1180. [Google Scholar] [CrossRef]
- Liu, T.Z.; Zhao, Z.Q.; Lang, Y.C.; Ding, H. Profile distribution and accumulation characteristics of organic carbon in a karst hillslope based on particle-size fractionation and stable isotope analysis. J. Environ. Biol. 2015, 36, 721–726. [Google Scholar] [PubMed]
- Angst, G.; Nierop, K.G.J.; Angst, S.; Frouz, J. Abundance of lipids in differently sized aggregates depends on their chemical composition. Biogeochemistry 2018, 140, 111–125. [Google Scholar] [CrossRef]
- Tang, Z.; Nan, Z.B. The potential of cropland soil carbon sequestration in the Loess Plateau, China. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 889–902. [Google Scholar] [CrossRef]
- Di, J.Y.; Feng, W.T.; Zhang, W.J.; Cai, A.D.; Xu, M.G. Soil organic carbon saturation deficit under primary agricultural managements across major croplands in China. Ecosyst. Health Sust. 2017, 3, 1364047. [Google Scholar] [CrossRef]
- Chen, S.; Arrouays, D.; Angers, D.A.; Martin, M.P.; Walter, C. Soil carbon stocks under different land uses and the applicability of the soil carbon saturation concept. Soil Till. Res. 2019, 188, 53–58. [Google Scholar] [CrossRef]
- Shang, C.; Tiessen, H. Carbon turnover and carbon-13 natural abundance in organo-mineral fractions of a tropical dry forest soil under cultivation. Soil Sci. Soc. Am. J. 2000, 64, 2149–2155. [Google Scholar] [CrossRef]
- Wei, X.; Shao, M.; Gale, W.J.; Zhang, X.; Li, L. Dynamics of aggregate-associated organic carbon following conversion of forest to cropland. Soil Biol. Biochem. 2013, 57, 876–883. [Google Scholar] [CrossRef]
- Churchman, G.J.; Foster, R.C.; D’Acqui, L.P.; Janik, L.J.; Skjemstad, J.O.; Merry, R.H.; Weissmann, D.A. Effect of land-use history on the potential for carbon sequestration in an Alfisol. Soil Till. Res. 2010, 109, 23–35. [Google Scholar] [CrossRef]
- Keiluweit, M.; Bougoure, J.J.; Nico, P.S.; Pett-Ridge, J.; Weber, P.K.; Kleber, M. Mineral protection of soil carbon counteracted by root exudates. Nat. Clim. Chang. 2015, 5, 588–595. [Google Scholar] [CrossRef]
- Wiesenberg, G.L.B.; Schwark, L.; Schmidt, M.W.I. Extractable lipid contents and colour in particle-size separates and bulk arable soils. Eur. J. Soil Sci. 2006, 57, 634–643. [Google Scholar]
- Nierop, K.G.J.; Naafs, D.F.W.; van Bergen, P.F. Origin, occurrence and fate of extractable lipids in Dutch coastal dune soils along a pH gradient. Org. Geochem. 2005, 36, 555–566. [Google Scholar] [CrossRef]
- Wiesenberg, G.L.B.; Dorodnikov, M.; Kuzyakov, Y. Source determination of lipids in bulk soil and soil density fractions after four years of wheat cropping. Geoderma 2010, 156, 267–277. [Google Scholar] [CrossRef]
- Medeiros, P.M.; Simoneit, B.R.T. Gas chromatography coupled to mass spectrometry for analyses of organic compounds and biomarkers as tracers for geological, environmental, and forensic research. J. Sep. Sci. 2007, 30, 1516–1536. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.H.; Simpson, M.J. Enhanced extractability of cutin- and suberin-derived organic matter with demineralization implies physical protection over chemical recalcitrance in soil. Org. Geochem. 2016, 97, 111–121. [Google Scholar] [CrossRef]
- Li, F.F.; Zhang, P.C.; Wu, D.P.; Xu, Y.; Chen, F.Y.; Chang, Z.F.; Chu, G.; Wang, L.; Pan, B.; Xing, B.S. Acid pretreatment increased lipid biomarker extractability: A case study to reveal soil organic matter input from rubber trees after long-term cultivation. Eur. J. Soil Sci. 2018, 69, 315–324. [Google Scholar] [CrossRef]
- Brodowski, S.; Amelung, W.; Haumaier, L.; Abetz, C.; Zech, W. Morphological and chemical properties of black carbon in physical soil fractions as revealed by scanning electron microscopy and energy-dispersive X-ray spectroscopy. Geoderma 2005, 128, 116–129. [Google Scholar] [CrossRef]
- Chang, Z.F.; Tian, L.P.; Li, F.F.; Wu, M.; Steinberg, C.E.W.; Pan, B.; Xing, B.S. Organo-mineral complexes protect condensed organic matter as revealed by benzene-polycarboxylic acids. Environ. Pollut. 2020, 260, 113977. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, C.; Hu, W.; Wang, L.; Li, Z.; Pan, J.; Chen, F. Stable isotope fractionation provides information on carbon dynamics in soil aggregates subjected to different long-term fertilization practices. Soil Till. Res. 2018, 177, 54–60. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Buchmann, N.; Flanagan, L.B. Carbon isotope ratios in belowground carbon cycle processes. Ecol. Appl. 2000, 10, 412–422. [Google Scholar] [CrossRef]
- Werth, M.; Kuzyakov, Y. C-13 fractionation at the root-microorganisms-soil interface: A review and outlook for partitioning studies. Soil Biol. Biochem. 2010, 42, 1372–1384. [Google Scholar] [CrossRef]
- Totsche, K.U.; Amelung, W.; Gerzabek, M.H.; Guggenberger, G.; Klumpp, E.; Knief, C.; Lehndorff, E.; Mikutta, R.; Peth, S.; Prechtel, A.; et al. Microaggregates in soils. J. Plant Nutr. Soil Sci. 2018, 181, 104–136. [Google Scholar] [CrossRef]
- Baldock, J.A.; Skjemstad, J.O. Role of the soil matrix and minerals in protecting natural organic materials against biological attack. Org. Geochem. 2000, 31, 697–710. [Google Scholar] [CrossRef]
- Coward, E.K.; Ohno, T.; Sparks, D.L. Direct evidence for temporal molecular fractionation of dissolved organic matter at the iron oxyhydroxide interface. Environ. Sci. Technol. 2019, 53, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Yang, Y. Selective stabilization of aliphatic organic carbon by iron oxide. Sci. Rep. 2015, 5, 11214. [Google Scholar] [CrossRef] [PubMed]
- Kopittke, P.M.; Dalal, R.C.; Hoeschen, C.; Li, C.; Menzies, N.W.; Mueller, C.W. Soil organic matter is stabilized by organo-mineral associations through two key processes: The role of the carbon to nitrogen ratio. Geoderma 2020, 357, 113974. [Google Scholar] [CrossRef]
- Kruger, J.P.; Leifeld, J.; Alewell, C. Degradation changes stable carbon isotope depth profiles in palsa peatlands. Biogeosciences 2014, 11, 3369–3380. [Google Scholar] [CrossRef]
- Cheng, X.; Yang, Y.; Li, M.; Dou, X.; Zhang, Q. The impact of agricultural land use changes on soil organic carbon dynamics in the Danjiangkou Reservoir area of China. Plant Soil 2013, 366, 415–424. [Google Scholar] [CrossRef]
- Pautler, B.G.; Austin, J.; Otto, A.; Stewart, K.; Lamoureux, S.F.; Simpson, M.J. Biomarker assessment of organic matter sources and degradation in Canadian High Arctic littoral sediments. Biogeochemistry 2010, 100, 75–87. [Google Scholar] [CrossRef]
- Wang, X.; Huang, X.; Sachse, D.; Hu, Y.; Xue, J.; Meyers, P.A. Comparisons of lipid molecular and carbon isotopic compositions in two particle-size fractions from surface peat and their implications for lipid preservation. Environ. Earth Sci. 2016, 75, 1142. [Google Scholar] [CrossRef]
- Chen, C.; Hall, S.J.; Coward, E.; Thompson, A. Iron-mediated organic matter decomposition in humid soils can counteract protection. Nat. Commun. 2020, 11, 2255. [Google Scholar] [CrossRef] [PubMed]
- Jilling, A.; Keiluweit, M.; Gutknecht, J.L.; Grandy, A.S. Priming mechanisms providing plants and microbes access to mineral-associated organic matter. Soil Biol. Biochem. 2021, 158, 108265. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Li, Y.; Zhang, Y.; Wu, B.; Zhou, D.; Peng, H.; Li, F.; Wu, M. Conversion of Natural Soil to Paddy Promotes Soil Organic Matter Degradation in Small-Particle Fractions: δ13C and Lipid Biomarker Evidence. Agronomy 2024, 14, 818. https://doi.org/10.3390/agronomy14040818
Li Y, Li Y, Zhang Y, Wu B, Zhou D, Peng H, Li F, Wu M. Conversion of Natural Soil to Paddy Promotes Soil Organic Matter Degradation in Small-Particle Fractions: δ13C and Lipid Biomarker Evidence. Agronomy. 2024; 14(4):818. https://doi.org/10.3390/agronomy14040818
Chicago/Turabian StyleLi, Yuxuan, Yan Li, Yu Zhang, Bingzhen Wu, Dandan Zhou, Hongbo Peng, Fangfang Li, and Min Wu. 2024. "Conversion of Natural Soil to Paddy Promotes Soil Organic Matter Degradation in Small-Particle Fractions: δ13C and Lipid Biomarker Evidence" Agronomy 14, no. 4: 818. https://doi.org/10.3390/agronomy14040818
APA StyleLi, Y., Li, Y., Zhang, Y., Wu, B., Zhou, D., Peng, H., Li, F., & Wu, M. (2024). Conversion of Natural Soil to Paddy Promotes Soil Organic Matter Degradation in Small-Particle Fractions: δ13C and Lipid Biomarker Evidence. Agronomy, 14(4), 818. https://doi.org/10.3390/agronomy14040818