Abstract
Heat shock proteins (HSPs) are mainly known to play important roles in plants against high-temperature (HT) stress. Their main function is to act as molecular chaperones for other proteins. It has also been proven that HSPs have a protective effect during other environmental stresses including low temperature (LT). To the best of our knowledge, the expression and role of HSPs in plants that have been exposed to LT have not yet been sufficiently reviewed. The aims of this minireview were (1) to briefly describe the origin, classification, structure, localisation and functions of HSPs, (2) to present the current knowledge about the changes in the accumulation of HSPs in plants that have been exposed to LT, (3) to discuss some of the molecular changes that occur during LT action and that lead to the accumulation of HSPs in plants and (4) to discuss the potential role of HSPs in acquiring tolerance to cold and frost in plants including economically important crop species. Some directions of research on the role of HSPs in plants growing in LT conditions are proposed.
1. Introduction—Discovery, Classification and Structure of Heat Shock Proteins
Heat shock proteins (HSPs) are an important group of proteins that can be found in plants, fungi and animal cells [1,2]. They are known to play a protective role against many environmental stresses as well as in restoring cellular homeostasis [3,4]. The name of these proteins is associated with the fact that they were first discovered in Drosophila melanogaster that had been treated with high-temperature stress [5]. HSPs were first described in the work of Tissiéres et al. [6]. Later studies have shown that the expression of the hsp genes is not only induced by heat but also by other environmental stresses, among others, cold, UV light, water, salinity, osmotic and biotic stresses [2,3,4].
The main function of HSPs is to act as molecular chaperones and they are also responsible for the folding, assembly, translocation and degradation of proteins during normally occurring processes [2,3,7]. It is also known that HSPs play an important role in the stabilisation of proteins and membranes. Moreover, they can participate in protein refolding during stress [3]. It is also known that HSPs are taking part in plant growth and development under normal conditions, such as inflorescence induction or seeds and fruits set [8,9].
In plants, HSPs are divided into five main classes characterized by their activity as molecular chaperones, according to their molecular weight, i.e., HSP100, HSP90, HSP70, HSP60 (100 kDa, 90 kDa, 70 kDa and 60 kDa, respectively) and small heat shock proteins (sHSPs) (low-molecular-mass proteins; 12–40 kDa) [2,3,4,10,11]. In reference to Gupta et al. [12], classification for five HSP families (HSP100, HSP90, HSP70, HSP70 and sHSP) can be made according to their molecular weight, amino acid sequence homologies and functions.
Each HSP has some individual features, but generally, the structure of the HSPs in plants consists of an N- and a C-terminus, which are the nucleus-binding region -I and -II, and a middle one, which is characterised by an amino terminus followed by a carboxyl terminus [11]. Each of the HSP structure elements are playing an important role in understanding their functions. The role of those elements can be slightly different for different HSP types. In the case of HSP100, it can contain five or one nucleotide-binding domains (NBDs) and it depends on the HSP100 class (I or II, respectively). HSP100 structure also includes middle and C-terminal domains [7,13]. NBDs and middle domains are characterised with a higher degree of conservation in HSP100 structure and it can be related to the highest importance of these regions with respect to their biological activity. The N- and C-terminal domains can have species-specific function [13]. In the case of HSP90, its structure consists of three domains: an N-terminal with an ATP-binding region, a middle domain and a C-terminal domain. The N-terminal domain together with the middle domain catalyses the ATP hydrolysis and the C-terminal domain is responsible for the formation of the HSP90 dimer, which is crucial for chaperone function [14,15,16,17,18]. The structure of HSP70 also consists of three domains: a 44 kDa nucleotide-binding domain (NBD), a 15 kDa substrate binding domain (SBD) and a 10 kDa carboxyl-terminal domain (CTD). The CTD forms a lid-like structure that helps to bind a substrate to the SBD [19,20,21,22,23]. ATP hydrolysis is crucial for HSP70 chaperone function. Because of low levels of intrinsic ATPase, co-chaperones (J-domain proteins (JDPs) and nucleotide exchange factors (NEFs)) are needed [3,23]. It is worth noting that HSP70 has a very high degree of structural homology among many species [3,21]. In plants, HSP70 genes are encoded by a highly conserved multigene family. In Arabidopsis thaliana, HSP70 is encoded by at least 18 genes. Meanwhile, at least 12 genes encoding HSP70 have been detected in the spinach genome [3,20]. It is thought that the HSP70 family is the most highly conserved HSP. They are considered to have about 50% identical residues between the Escherichia coli homolog DnaK and the eukaryotic HSP70 [3,20,24]. According to Waters and Vierling [25], the structure of both plant and bacterial sHSP includes an α-crystallin domain (ACD) at their C-terminus that consists of approx. 90 amino acids that build up a dense, seven-stranded β-sandwich that forms the structural core of all sHSPs. Other important features of the sHSP structure are a dimeric substructure of the oligomer and dimer–dimer interactions that are crucial for stabilising oligomers [25,26,27,28,29,30,31]. Oligomer formation is a common feature of sHSPs [31]. Unlike other HSPs, sHSPs do not require ATP to bind substrate proteins and have a very high ability to bind denatured substrates [25]. sHSPs operating in specific organelles or cellular compartments possess N-terminal transit, leader or signal sequences necessary for transfer of the sHSP to the appropriate organelle. To date, little is known about the higher levels of structural variation among plant sHSPs [32].
In addition to a brief description of the discovery, classification and structure of HSPs presented above, the aim of the presented article is (1) to briefly describe HSPs localization and functions, (2) to present the current knowledge about the changes in the accumulation of HSPs in plants that have been exposed to low temperature (LT), (3) to discuss some of the molecular changes that occur during LT action and that lead to the accumulation of HSPs in plants and (4) to discuss the potential role of HSPs in acquiring tolerance to cold and frost in plants including economically important crop species. Moreover, some directions of further research on the role of HSPs in plants growing in LT conditions are proposed.
2. Heat Shock Proteins—Localisation and Function in Plants
HSPs can be found in cytoplasm as well as in other organelles. They were found in the nucleus of plants such as cereals including maize (Zea mays L.), wheat (Triticum aestivum L.), barley (Hordeum vulgare L.) and rice (Oryza sativa L.) [1,33,34]. Moreover, in plants such as Arabidopsis thaliana and soybean (Glycine max L.) [1,35], HSPs have been found in the nucleus. Other organelles in which HSPs occur are the mitochondria of plants such as cereals, including rice, maize, wheat and barley [34,36]. It is also known that HSPs are present in the mitochondria of plants such as tomatoes (Solanum lycopersicum L.), potatoes (Solanum tuberosum L.), tobacco (Nicotiana tabacum L.), Arabidopsis thaliana and soybean [1,35,37,38,39]. HSPs also occur in the chloroplasts of plants such as cereals including maize and wheat [1,28] as well as in creeping grass (Holcus mollis L.), tomatoes, Arabidopsis thaliana and soybean [1,35,40,41,42,43,44,45]. Finally, HSPs have also been found in the endoplasmic reticulum (ER) of plants such as pea (Pisum sativum L.), mulberry (Morus L.), Arabidopsis thaliana, soybean [1,35,39,44,46,47] and cereals including rice and maize [1].
The localisation of specific HSP classes and their main functions are as follows: HSP100 is located in the cytosol, mitochondria and chloroplasts [48,49] and the major function of HSP100 is that it is responsible for protein disaggregation and/or degradation, which is very important for maintaining cellular homeostasis [48,49]. HSP90 can be found in the cytosol, chloroplasts, mitochondria and ER [1,14,16,50]. It is responsible for managing protein folding and is crucial in the signal-transduction networks, cell-cycle control, protein degradation and protein trafficking [1,14,16,50]. HSP70 is one of the best known HSPs. It is located in the cytosol, mitochondria, chloroplasts and ER [19,20,21,22,23,51]. The main functions of HSP70 include preventing protein aggregation, and assisting with protein refolding, protein import and translocation, signal transduction and transcriptional activation [19,20,21,22,23,51]. HSP60 (so-called chaperonins) are present in the cytosol, mitochondria and chloroplasts [52,53]. The main functions of chaperonins are protein folding, refolding, aggregation and transferring them to the chloroplasts and mitochondria [53].
Finally, the class of sHSP is the most common HSP class in plants. These proteins are located in the cytosol, mitochondria, chloroplasts, peroxisomes, ER and membranes [26,27,28,49,54]. They are known to prevent protein aggregation and they help to stabilise non-native proteins [26,27,28,49,54].
3. The Role of Heat Shock Proteins in High-Temperature (HT) Stressed Plants
Because of the discovery of HSPs as a result of HT exposure, the vast majority of research concerning HSPs in plants has been conducted in this aspect [55]. The data that were obtained in the mentioned studies show that HT mainly results in an increase in HSPs accumulation in plants. Especially nowadays, when we are dealing with climate change, HT is a serious problem for plants. This is of great importance in agriculture as HT can lead to problems with the survival, yield and quality of plants [49,56]. An increased HSPs accumulation is one of the mechanisms by which plants avoid and minimise the damaging effects of this stress. Briefly, it has been studied in many plant species such as soybean in which Altschuler and Mascarenhas [57] observed an increased HSP accumulation after plants had been exposed to a temperature of 40 °C for up to 2.5 h, after which a decreased HSPs level was observed (between 2.5 and 6 h). Moisyadi and Harrington [58] observed a higher accumulation of high-molecular-mass HSP in cultured sugarcane cells (Saccharum officinarum L.), which was caused by HT (32 °C or 34 °C), which lasted for as little as ten minutes. In the case of sHSP (18 kDa), its increased level was observed after heat shock at 36 °C for 30 min. Dupuis and Dumas [59] found that maize plants that were subjected to 40 °C for 4 h were characterised by an increased accumulation of HSP101, HSP70 and HSP17.6. Similarly, DeRocher et al. [60] found an increased accumulation of HSP18.1 and HSP17.9 transcripts and HSP18.1 protein in pea (Pisum sativum L.) plants that had been exposed to HT 38 °C for 4 h. Additionally, Hopf et al. [61] found that after maize was exposed to 40/45 °C for 2 h, the HSP accumulation increased in the vegetative tissues. Studies on spinach (Spinacia oleracea L.) and tomato have shown that the HSP expression increased after 1 h of heat shock (37 °C) and that it decreased to the level before stress after 2 h [62]. Dhaubhadel et al. [63] studied oilseed rape plants that were exposed to 45 °C and they observed an increased accumulation of HSP101, HSP90 and HSP70 after 4 h of HT treatment, compared to the control plants. In the case of potato plants, exposure to an HT (35 °C or 40 °C) caused an increased accumulation of sHSP compared to the control plants [64]. Pavli et al. [65] found that in sorghum (Sorghum bicolor (L.) Moench) there was an increase in the accumulation of the hsp90 gene expression after plants were exposed to 47 °C for 30 or 60 min. However, the hsp90 expression levels differed among the studied genotypes. Recently, Sun et al. [40] studied creeping bentgrass (Agrostis stolonifera L.) plants (cv. Penn-A4) that had been exposed to an HT of 37 °C for 4 h and they observed an increased expression of HSP26.8a in both the leaves and roots.
4. Changes in the Heat Shock Proteins in Plants That Had Been Exposed to Low-Temperature Stress
HSPs not only accumulate in plants growing under HT stress, but their expression also increases under the influence of other abiotic stresses including low temperature (LT) stress [66,67]; however, this aspect has been investigated much less. Therefore, as mentioned, one of the aims of this minireview is to present the current knowledge about what is known to date about the expression and a potential role of HSPs in the process of plant acclimation to LT.
In the beginning, it is important to say that in general, plants can be divided into two main groups. The first group includes species for which cold is a stress factor (tomato, cucumber) and which are generally unable to survive temperatures below zero. The second group includes plants (i.e., spring and winter cereals, oilseed rape) able to survive frost particularly after a process known as cold acclimation (cold hardening). The metabolic changes that occur during cold acclimation are quite well known [56,68]. These processes include a change in the lipid composition and membrane structure (an increase in the content of unsaturated fatty acids in the membrane lipids) and changes in the photosynthesis efficiency [69], content of soluble sugars [70] and the accumulation of proteins such as late embryogenesis abundant proteins (LEA), anti-freezing proteins (AFPs) and cold-shock proteins (CSPs), which increases during cold acclimation [71]. As was mentioned earlier, cold acclimation also results in an accumulation of HSPs [11,47,67,72]. As mentioned, cold acclimation allows many economically important species to acquire better frost tolerance.
Importantly, during cold acclimation, genes performing various functions are activated, including genes responsible for the transduction of the signal caused by LT. One of the most important ones is the gene of the transcription factor crucial for cold acclimation, i.e., CBF (C-repeat Binding Factor). The cold-induced or constitutive expression of CBF1 reduces the level of bioactive gibberellins, resulting in the accumulation of DELLA proteins. As a result of the accumulation of DELLA, plant growth slows down, while an increase in the level of active gibberellins in spring is responsible for their growth [73,74]. CBF also induces the expression of COR genes (Cold-Responsive Genes), which are important in the processes of plant acclimation to LT [75,76]. Both CBF and COR transcription factors help provide plants with frost tolerance by, i.a., the acceleration of osmolyte synthesis or reconstruction of cell membranes [76,77]. The effect of LT, like other abiotic stresses, contributes to the induction of the expression of the so-called Heat Shock Factor (HSF) gene family, which contribute to the induction of HSPs gene expression [78]. More details are presented in Section 5.
Generally, LT stress is an especially serious problem in horticulture and agriculture [56,79]. That is why mechanisms of the plant response to LT are extensively studied. The accumulation of HSPs has also been studied in many plant species including horticultural plants and agricultural plants but also in model plants.
4.1. Changes in HSP in Model Plants
In the case of the main model plants, HSP accumulation was already studied in Arabidopsis thaliana that had been subjected to LT. A. thaliana belongs to a group of plants capable of surviving frost especially after cold acclimation [80]. Bae et al. [81] found that A. thaliana plants that had been subjected to LT (4 °C) for 6 h had an increased accumulation of HSP70 transcripts compared to the non-acclimated plants (grown at 22 °C). Kawamura and Uemura [82] studied the plasma membrane proteins that were present in A. thaliana plants that had been subjected to 2 °C for 1, 2, 3 and 7 days and they found that after 1 day of cold acclimation, the accumulation of chaperonin 20 decreased and that it increased after 2 days compared to the non-cold-acclimated plants. Kagale et al. [83] observed an increased accumulation of the HSP90 transcript in A. thaliana seedlings that had been exposed to 2 °C for three days. HSP90 occurred after 1 h of LT exposure and its highest level was observed after 12 h of LT. After one and three days of LT, a similar high amount of the HSP90 level was observed. Kaplan et al. [84] studied A. thaliana plants that had been exposed to 4 °C for up to 96 h and they observed an increased expression of HSP70 after 12 h and especially after 24 h of LT exposure compared to the control plants. In the work by Sewelam et al. [85], A. thaliana plants that had been exposed to 0 °C for 3 h were characterised with a 4-fold higher up-regulation of HSP17.4CI expression, in comparison to control plants.
4.2. Changes in HSP in Horticultural Plants
Horticultural plants such as tomato, cucumber and maize belong to a group of plants that are sensitive to cold [86,87,88,89]. A few authors studied the HSPs accumulation in such plants that had been exposed to LT stress. Anderson et al. [90] studied spinach plants (cv. Bloomsdale) that had been cold-acclimated at 5 °C for 1, 2, 3, 4 and 7 days and they found an up-regulation of HSP70 mRNA during cold acclimation compared to the control plants. Taylor et al. [91] studied pea plants that had been exposed to 4 °C for 36 h and they found an increased accumulation of HSP90, HSP70 and HSP22 compared to the control plants. Degand et al. [92] observed an increased HSP70 accumulation in the roots of chicory (Cichorium intybus L.) that had been exposed to a minimum temperature below 5 °C for several days compared to the non-cold-exposed plants. Moreover, Dumont et al. [93] studied pea plants [cv. Champagne (cold-tolerant) and cv. Terese (cold-sensitive)] that had been cold-acclimated at 10/2 °C (day/night) for 11 days and they found an increased accumulation of HSP70 in the pea stems of both of the studied cultivars compared to the non-acclimated plants. Research on three tomato genotypes by Kubienová et al. [94] showed a similar tendency, namely, the increase in HSP70 protein accumulation was observed in only one genotype (S. habrochaites) that had been exposed to 4 °C for 1 h, 4 h and 24 h compared to the non-stressed plants. Growth at 10 °C caused a slight decrease in the HSP70 protein accumulation in two genotypes (S. habrochaites and S. lycopersicum), while in the third (S. chmielewskii), no changes were observed compared to the control plants (grown at 20 °C) [94]. As for tomato, very interesting findings of practical importance were obtained by Sabehat et al. [95]. The authors observed that HT pretreatment of tomato fruits resulted in greater HSPs accumulation, which allowed them to be stored in LT for longer periods than normally possible. There are also studies that concern HSPs expression in cucumber fruits after storage at 4 °C [96]. Authors found that after 14 days of storage at 4 °C, cucumber fruits were characterised with a significantly higher relative expression level of HSP70 and HSP40.9 than control fruits [96].
4.3. Changes in HSP in Agricultural Plants
Winter plants—cereals and oilseed rape—acclimate to frost during the autumn months via a process of cold acclimation. Spring cultivars also have the ability to acclimate to the cold, which is especially useful during periods of temperature anomalies, e.g., frost in the early spring. Frost, especially with insufficient snow cover, can cause significant yield losses, thereby generating economic problems. That is why the mechanisms of cold acclimation and plant frost tolerance have been the subject of studies for many years. There have also been studies concerning the HSPs accumulation in LT-exposed agricultural plants. Cabané et al. [97] studied soybean plants (cv. Verdon and Maple Ardon) that had been cold-acclimated at 14/8 °C (d/n) for five days and they observed an increased accumulation of HSP70 compared to the control plants. Krishna et al. [98] studied HSP90 accumulation in spring oilseed rape (Brassica napus L., cv. Westar) plants that had been exposed to 5 °C for seven days. The authors found that the LT-exposed plants were characterised by a significantly higher accumulation of the HSP90 transcripts and proteins than the control plants (grown at 20 °C) [98]. Pareek et al. [99] observed an increased accumulation of HSP90 in the shoots of rice seedlings that had been cold-stressed at 5 ± 2 °C for 96 h compared to the control plants that had been grown at 28 °C. Cui et al. [100] found that rice seedlings that had been exposed to LT of 15, 10 and 5 °C (24 h for each exposure) were characterised by a greater than two-fold up-regulation of HSP70 compared to the control plants. The highest level of HSP70 was observed in the plants that had been exposed to 5 °C. Similar results were observed by Yan et al. [66] in rice plants that had been exposed to LT 6 °C for 24 h and they noticed an increased accumulation of HSP70 and HSP26 compared to the control plants. Moreover, Lee et al. [101] observed that after chilling stress of rice at 10 °C for 24 h and 72 h, there was an up-regulation of the HSP70 proteins in the rice roots especially after 72 h of chilling stress compared to the control plants that had been grown at 28 °C. Renaut et al. [102] studied two potato cultivars (PS3 and Desiree) that had been exposed to 4 °C for 21 days and they found an increase in the amount of the HSP70 protein in both cultivars compared to the control plants. The cold acclimation (6 °C, 12 weeks) of winter wheat (Triticum aestivum L.) caused an increase in the accumulation of HSP70 (after 3, 7 and 21 days of cold acclimation) and a decrease in HSP90 (after 21 and 84 days of cold acclimation) in a crude extract of the crowns [103]. Hlaváčková et al. [104] studied winter barley (cv. Luxor) and they found that LT exposure (3/2 °C (d/n) for three weeks) caused an increased accumulation of HSP70 protein in the crowns compared to the control plants (18/13 °C (d/n)).
Sadura et al. [105] studied the HSPs accumulation in barley that had been acclimated at 5 °C for 21 days. The authors found that while the cultivar Delisa accumulated significantly more HSP70 in the cell membrane fraction compared to the non-acclimated plants, the second studied cultivar, Bowman, was characterised by a notably lower accumulation of this protein in the cold-acclimated plants than in the non-acclimated plants. The accumulation of the HSP90 protein was lower for both of the studied cultivars that had been cold-acclimated than for the non-acclimated plants [105]. Similarly, in the work of Stachurska et al. [106], an increased accumulation of HSPs (HSP70 cytoplasmic, HSP70 chloroplastic and HSP90) was observed in three out of four of the tested cultivars of oilseed rape plants (cv. Bojan, Feliks and President), which had been cold-acclimated at 4 °C for three weeks compared to the control plants. In the case of the cultivar Rokas, the opposite effect was observed for the HSP70 cytoplasmic and HSP90 types, namely, the accumulation of these proteins was lower for the cold-acclimated plants than for the control plants [106]. The reviewed studies are summarised in Table 1.

Table 1.
The HSP accumulation in plants that had been exposed to a low temperature.
4.4. Are HSPs Required for Acquiring Frost Tolerance?
According to Thomashow [75], the HSP70 and HSP90 genes can be associated with the acquisition of frost tolerance by plants. Other authors investigated whether the changed HSPs content in cold-acclimated plants translated into a changed frost tolerance in these plants. In the work by Kawamura and Uemura [82], an increased accumulation of chaperonin 20 in the plasma membrane of A. thaliana plants that had been cold-acclimated at 2 °C corresponded to increased frost tolerance in the plants. Vítámvás et al. [103] found that the highest level of winter wheat tolerance to frost was observed after 21 days of cold acclimation at 6 °C and that at the same time, the HSP70 accumulation was higher compared to the control plants. However, the HSP90 content decreased after 21 and 84 days of cold exposure. The results that were obtained by Sadura et al. [105], which indicated an increased accumulation of HSP70 in the barley cultivar Delisa and its decreased accumulation in the cultivar Bowman, did not correspond with values of frost tolerance that were obtained for these plants. Lower values of frost tolerance were obtained for cultivar Delisa than for Bowman [107]. Stachurska et al. [106] studied the HSPs accumulation in four cultivars of oilseed rape (Bojan, President, Rokas and Feliks) that had been cold-acclimated at 4 °C for three weeks and they found that for three of the four cultivars, an increased HSP90 and HSP70 accumulation was accompanied by an increase in their frost tolerance. The exception was cultivar Rokas, which was characterised by an increased frost tolerance and a decrease in the accumulation of HSPs, although interestingly in non-acclimated plants of this cultivar, relatively high basal accumulations of HSP90 and cytoplasmic HSP70 were noted, in comparison to other cultivars.
Therefore, it can be stated that the increased accumulation of HSPs in plants that have been exposed to LT is often accompanied by their increased frost tolerance, although a detailed regulatory mechanism of this phenomenon requires further research. In this context, particularly interesting studies were completed by Hlaváčková et al. [104]. The authors studied winter barley (cultivar Luxor) that had been exposed to a frost temperature (−3 °C) after three weeks of cold acclimation (3/2 °C (d/n)) and they observed a seven-fold increase in the HSP70 level in frost-treated plants compared to plants that had been exposed to cold. As shown, for example, by Sadura et al. [105], in some cases, i.e., in cereals, the accumulation of HSPs transcripts during cold acclimation is high. However, this is not accompanied by a high HSP protein content (Figure 1—Bowman plants and their NILs). Therefore, the question arises whether the increased transcript abundance is not a ‘preliminary’ mechanism during cold acclimation and whether the increase in the protein level does not occur/enhance as a result of stronger stress like frost in cereals?
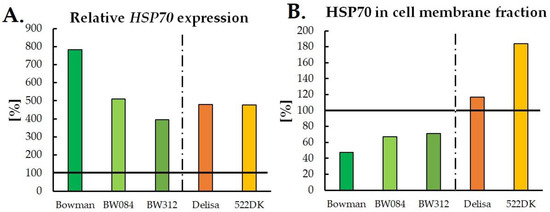
Figure 1.
Relative percentage changes in the HSP70 transcript accumulation (A) and HSP70 protein accumulation in the cell membrane fraction (B) in the barley plants that had been acclimated at 5 °C relative to the plants that had been grown at 20 °C. The results that were obtained for the plants that had been grown at 20 °C were considered to be 100% and are indicated by the horizontal black line. The original data of HSP70 expression are available in Sadura et al. [106]. Bowman—a reference cultivar for two NILs: BW084—plants with disturbances in the early stage of the BR biosynthetic pathway and BW312—plants with a BRI1 receptor defect. Delisa—the reference cultivar for the 522DK mutant with disturbances in the late stage of the BR biosynthesis pathway.
5. Possible Molecular Mechanisms of HSPs Action in Plants under LT Conditions
As mentioned above, a better understanding of the role of HSPs in LT-exposed plants requires a deeper study of the molecular mechanisms that are involved in the actions of HSPs. According to Horváth et al. [54], HSPs are responsible for thermal sensing via the membranes. sHSPs play especially important roles in membranes as under stressful conditions, they become membrane-associated, and they can modify the lipid order and phase state of membranes.
According to Usman et al. [22], after plants have been exposed to stressful conditions (such as HT or LT), specific and highly regulated signalling cascades are initiated, and the outer membrane proteins that are present in the cytosol send signals to the inner membrane proteins, which causes the up-regulation of the heat shock transcription factors (HSFs). As a result, the synthesis of HSPs is triggered [8,22,54,108]. The promoter regions of the HSF bind to the so-called Heat Shock Elements (HSEs), after which there is an increased amount of HSPs. There are three types of HSFs: HSFA, B and C [8,109]. The studies of Kotak et al. [49] and Scharf et al. [109] indicated that the increased expression of HSPs and other stress-responsive genes in conjunction with HSFs is an important factor in modifying the physiological and biochemical processes that are associated with the development of a tolerance to stresses.
The general model of the molecular mechanism of HSPs biosynthesis in plants exposed to various stresses, which can also be applied to LT stress, is presented in Figure 2.
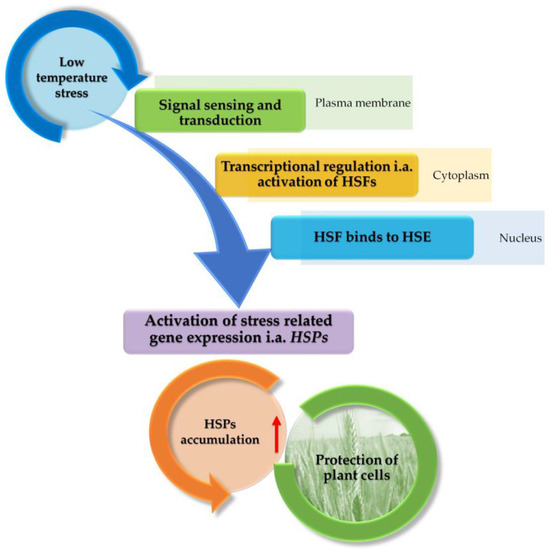
Figure 2.
The proposed model of the molecular mechanism of HSPs biosynthesis in plants exposed to LT stress (based on [8,22,54,108]). Briefly, LT is a signal that is received in the cell membrane and transmitted to the cytoplasm, where heat shock transcription factors (HSFs) are activated. In the nucleus, HSFs are associated with HSEs (Heat Shock Elements). HSPs gene expression is then activated, which may contribute to HSPs accumulation.
It is known that after plants have been exposed to HT stress, the gene expression of HSPs is initiated very quickly, in as little as a couple of seconds of the HT, and the maximal amount of HSPs can be observed after 2 h of HT stress [77]. A similar effect can be observed in plants that are exposed to LT. Increased HSPs transcript accumulation was observed after 1 h of LT stress [94]. To the best of our knowledge, changes in transcript accumulation after less than 1 h have not been studied. The biosynthesis of HSP proteins is under both transcriptional and post-transcriptional control [63,110], and therefore the levels of the HSP transcripts and proteins can differ during the exposure of plants to temperature stress. This was observed, for example, in the work on barley by Sadura et al. [105]. The authors observed that despite a higher HSP90 transcript level after cold acclimation (4 °C) relative to the control (20 °C), the HSP90 protein level in membranes was consistently lower in both of the studied cultivars, especially in the barley Bowman cultivar. Moreover, the several-times higher level of the HSP70 transcript in both cultivars (Bowman and Delisa) in cold conditions compared to the control at 20 °C did not have such a clear impact on the accumulation of HSP70 in the membranes and cytoplasm (Figure 1). In addition, in the work of Kubienová et al. [94] on tomato plants (Solanum lycopersicum L., cultivar Amateur), which had been cold-exposed at 4 °C, the HSP70 transcript accumulation was higher compared to the control plants. After the 1st h of exposure, there was an eleven-fold increase in the HSP70 gene expression and after the 4th and 24th h, a further increase was observed. However, low levels of the HSP70 proteins were observed for this cultivar. Other studied cultivars have also differed in the levels of HSP70 gene expression and HSP70 protein accumulation [94]. Therefore, the mechanism of the post-transcriptional regulation of HSPs biosynthesis may play an important role in LT-exposed plants and requires deeper studies.
6. Hormonal Control of HSPs Biosynthesis in LT-Exposed Plants
Some studies indicate that the phytohormone level, e.g., brassinosteroids (BR), might have an impact on the accumulation of HSPs in LT-exposed plants. Kagale et al. [83] studied A. thaliana plants that had been exposed to 2 °C for up to three days and then treated with 1 µM 24-epibrassinolide (EBR). However, the authors did not observe any differences in the HSP90 transcript accumulation between the EBR-treated and the untreated plants. On the other hand, Sadura et al. [105] studied the accumulation of HSPs in cold-acclimated barley mutants with a BR deficiency (BW084, 522DK) and impaired BR-signalling (BW312) (5 °C, 21 days) and the authors observed differences in HSPs accumulation in studied plants. Mutants with a BR deficit (with few exceptions) and a BRI1 receptor defect were most often characterised by lower levels of the accumulation of the transcripts of all of the HSPs compared to their wild types (cv. Bowman, Delisa). This could indicate a role for BR as positive regulators of HSPs transcription in barley. As for HSP protein accumulation, the BRI1 receptor defect was associated with a decrease in the accumulation of the HSP70 and HSP90 proteins after cold acclimation. This could also indicate the involvement of BR in the regulation of the biosynthesis of the HSP proteins via the BRI1 receptor and the associated signal transduction pathway. However, a BR deficit in plant tissues was not always associated with changes in the HSPs biosynthesis [105]. The relative percentage changes in the HSP70 expression in the barley BR mutants and their wild types that had been acclimated at 5 °C relative to the control plants that had been grown at 20 °C are presented in Figure 1.
Finally, Stachurska et al. [106] found that an increased accumulation of HSPs in cold-acclimated (4 °C, three weeks) oilseed rape plants was accompanied by an increased accumulation of abscisic acid (ABA) and a decreased accumulation of growth-promoting hormones. However, a direct connection between HSPs biosynthesis and phytohormones’ level was not proven in this work.
Hormonal mechanisms of HSPs biosynthesis regulation still require further research.
7. Conclusions and Further Perspectives
Despite the fact that the discovery of HSPs was as cell protective factors at HT, these proteins may also play important roles in counteracting the negative effects of LT stress in plants. The results that are discussed in this minireview indicate that the accumulation of HSPs occurs in plants that have been exposed to LT and that this often depends on the plant species, the genotype, the duration of LT exposure and the plants sensitivity to LT. A high accumulation of transcripts is not always reflected in the final content of the corresponding proteins. Therefore, further research is necessary, for example, on the mechanisms of the post-transcriptional regulation of HSPs biosynthesis. Further research on the hormonal regulation of HSPs proteins in plants exposed to LT seems to be equally important. According to some studies, a higher content of HSPs may translate into greater frost tolerance in plants. This is an issue worth further investigation as it would allow to determine whether the modification of the HSPs content in plants would be useful for breeders to breed more frost-tolerant plants. The same could also concern cold-sensitive species and possibilities of obtaining more cold-tolerant plants. An increase in the HSPs content may also have other practical significance, namely in cold storage of fruits. In the case of tomato fruit, for example, it has been proven that higher HSP content can extend the duration of its cold storage.
Author Contributions
Conceptualisation, I.S. and A.J.; data curation, I.S.; writing—original draft preparation, I.S.; writing—review and editing, A.J. and I.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Xu, Z.S.; Li, Z.Y.; Chen, Y.; Chen, M.; Li, L.C.; Ma, Y.Z. Heat Shock Protein 90 in Plants: Molecular Mechanisms and Roles in Stress Responses. Int. J. Mol. Sci. 2012, 13, 15706–15723. [Google Scholar] [CrossRef] [PubMed]
- Park, C.J.; Seo, Y.S. Heat Shock Proteins: A Review of the Molecular Chaperones for Plant Immunity. Plant Pathol. J. 2015, 31, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of Plant Heat-Shock Proteins and Molecular Chaperones in the Abiotic Stress Response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Al-Whaibi, M.H. Plant Heat-Shock Proteins: A Mini Review. J. King Saud Univ.-Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- Ritossa, F. A New Puffing Pattern Induced by Temperature Shock and DNP in Drosophila. Experientia 1962, 18, 571–573. [Google Scholar] [CrossRef]
- Tissiéres, A.; Mitchell, H.K.; Tracy, U.M. Protein Synthesis in Salivary Glands of Drosophila Melanogaster: Relation to Chromosome Puffs. J. Mol. Biol. 1974, 84, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.S.; Chakradhar, T.; Reddy, R.A.; Nitnavare, R.B.; Mahanty, S.; Reddy, M.K. Role of Heat Shock Proteins in Improving Heat Stress Tolerance in Crop Plants. In Heat Shock Proteins and Plants; Asea, A., Kaur, P., Calderwood, S., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 283–307. [Google Scholar]
- ul Haq, S.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.X.; Zhang, H.X.; Wei, A.M.; Gong, Z.H. Heat Shock Proteins: Dynamic Biomolecules to Counter Plant Biotic and Abiotic Stresses. Int. J. Mol. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef] [PubMed]
- Majee, A.; Kumari, D.; Sane, V.A.; Singh, R.K. Novel Roles of HSFs and HSPs, Other than Relating to Heat Stress, in Temperature-Mediated Flowering. Ann. Bot. 2023, 132, 1103–1106. [Google Scholar] [CrossRef] [PubMed]
- Yurina, N.P. Heat Shock Proteins in Plant Protection from Oxidative Stress. Mol. Biol. 2023, 57, 951–964. [Google Scholar] [CrossRef]
- Khan, S.; Jabeen, R.; Deeba, F.; Waheed, U.; Khanum, P.; Iqbal, N. Heat Shock Proteins: Classification, Functions and Expressions in Plants during Environmental Stresses. J. Bioresour. Manag. 2021, 8, 85–97. [Google Scholar] [CrossRef]
- Gupta, S.C.; Sharma, A.; Mishra, M.; Mishra, R.K.; Chowdhuri, D.K. Heat Shock Proteins in Toxicology: How Close and How Far? Life Sci. 2010, 86, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, M.; Katiyar-Agarwal, S.; Grover, A. Plant Hsp100 Proteins: Structure, Function and Regulation. Plant Sci. 2002, 163, 397–405. [Google Scholar] [CrossRef]
- Pearl, L.H.; Prodromou, C. Structure and Mechanism of the Hsp90 Molecular Chaperone Machinery. Annu. Rev. Biochem. 2006, 75, 271–294. [Google Scholar] [CrossRef] [PubMed]
- Wayne, N.; Mishra, P.; Bolon, D.N. Hsp90 and Client Protein Maturation. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2011; Volume 787, pp. 33–44. ISBN 9781617792946. [Google Scholar]
- Tichá, T.; Samakovli, D.; Kuchařová, A.; Vavrdová, T.; Šamaj, J. Multifaceted Roles of Heat Shock Protein 90 Molecular Chaperones in Plant Development. J. Exp. Bot. 2020, 71, 3966–3985. [Google Scholar] [CrossRef] [PubMed]
- Biebl, M.M.; Buchner, J. Structure, Function, and Regulation of the Hsp90 Machinery. Cold Spring Harb. Perspect. Biol. 2019, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Kadota, Y.; Shirasu, K. The HSP90 Complex of Plants. Biochim. Biophys. Acta-Mol. Cell Res. 2012, 1823, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Miernyk, J.A. The 70 KDa Stress-Related Proteins as Molecular Chaperones. Trends Plant Sci. 1997, 2, 180–187. [Google Scholar]
- Sung, D.-Y.; Kaplan, F.; Guy, C.L. Plant Hsp70 Molecular Chaperones: Protein Structure, Gene Family, Expression and Function. Physiol. Plant. 2001, 113, 443–451. [Google Scholar] [CrossRef]
- Sharma, D.; Masison, D. Hsp70 Structure, Function, Regulation and Influence on Yeast Prions. Protein Pept. Lett. 2009, 16, 571–581. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Martini, M.Y.; Yusuff, O.A.; Ismail, M.R.; Miah, G. Molecular Analysis of Hsp70 Mechanisms in Plants and Their Function in Response to Stress. Biotechnol. Genet. Eng. Rev. 2017, 33, 26–39. [Google Scholar] [CrossRef]
- Berka, M.; Kopecká, R.; Berková, V.; Brzobohatý, B.; Černý, M. Regulation of Heat Shock Proteins 70 and Their Role in Plant Immunity. J. Exp. Bot. 2022, 73, 1894–1909. [Google Scholar] [CrossRef] [PubMed]
- Ray, D.; Ghosh, A.; Mustafi, S.B.; Raha, S. Plant Stress Response: HSP70 in the Spotlight. In Heat Shock Proteins and Plants; Asea, A., Kaur, P., Calderwood, S., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 10, pp. 283–306. ISBN 978-3-319-46339-1. [Google Scholar]
- Waters, E.R.; Vierling, E. Plant Small Heat Shock Proteins – Evolutionary and Functional Diversity. New Phytol. 2020, 227, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Scharf, K.D.; Siddique, M.; Vierling, E. The Expanding Family of Arabidopsis Thaliana Small Heat Stress Proteins and a New Family of Proteins Containing α-Crystallin Domains (Acd Proteins). Cell Stress Chaperones 2001, 6, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Van Montagu, M.; Verbruggen, N. Small Heat Shock Proteins and Stress Tolerance in Plants. Biochim. Biophys. Acta 2002, 1577, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, H.; Khurana, N.; Nijhavan, A.; Khurana, J.P.; Khurana, P. The Wheat Chloroplastic Small Heat Shock Protein (SHSP26) Is Involved in Seed Maturation and Germination and Imparts Tolerance to Heat Stress. Plant Cell Environ. 2012, 35, 1912–1931. [Google Scholar] [CrossRef] [PubMed]
- Carra, S.; Alberti, S.; Arrigo, P.A.; Benesch, J.L.; Benjamin, I.J.; Boelens, W.; Bartelt-Kirbach, B.; Brundel, B.J.J.M.; Buchner, J.; Bukau, B.; et al. The Growing World of Small Heat Shock Proteins: From Structure to Functions. Cell Stress Chaperones 2017, 22, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Guihur, A.; Rebeaud, M.E.; Goloubinoff, P. How Do Plants Feel the Heat and Survive? Trends Biochem. Sci. 2022, 47, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Leroux, M.R.; Melki, R.; Gordon, B.; Batelier, G.; Candido, E.P.M. Structure-Function Studies on Small Heat Shock Protein Oligomeric Assembly and Interaction with Unfolded Polypeptides. J. Biol. Chem. 1997, 272, 24646–24666. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.R. The Evolution, Function, Structure, and Expression of the Plant SHSPs. J. Exp. Bot. 2013, 64, 391–403. [Google Scholar] [CrossRef]
- Nieto-Sotelo, J.; Martínez, L.M.; Ponce, G.; Cassab, G.I.; Alagón, A.; Meeley, R.B.; Ribaut, J.M.; Yang, R. Maize HSP101 Plays Important Roles in Both Induced and Basal Thermotolerance and Primary Root Growth. Plant Cell 2002, 14, 1621–1633. [Google Scholar] [CrossRef]
- Pandey, B.; Kaur, A.; Gupta, O.P.; Sharma, I.; Sharma, P. Identification of HSP20 Gene Family in Wheat and Barley and Their Differential Expression Profiling Under Heat Stress. Appl. Biochem. Biotechnol. 2014, 175, 2427–2446. [Google Scholar] [CrossRef] [PubMed]
- Chong, L.P.; Wang, Y.; Gad, N.; Anderson, N.; Shah, B.; Zhao, R. A Highly Charged Region in the Middle Domain of Plant Endoplasmic Reticulum (ER)-Localized Heat-Shock Protein 90 Is Required for Resistance to Tunicamycin or High Calcium-Induced ER Stresses. J. Exp. Bot. 2015, 66, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Korotaeva, N.E.; Antipina, A.I.; Grabelnykh, O.I.; Varakina, N.N.; Borovskii, G.B.; Voinikov, V.K. Mitochondrial Low-Molecular-Weight Heat-Shock Proteins and the Tolerance of Cereal Mitochondria to Hyperthermia. Russ. J. Plant Physiol. 2001, 48, 798–803. [Google Scholar] [CrossRef]
- Neumann, D.; Emmermann, M.; Thierfelder, J.-M.; zur Nieden, U.; Clericus, M.; Braun, H.-P.; Nover, L.; Schmitz, U.K. HSP68—A DnaK-like Heat-Stress Protein of Plant Mitochondria. Planta 1993, 190, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Sanmiya, K.; Suzuki, K.; Egawa, Y.; Shono, M. Mitochondrial Small Heat-Shock Protein Enhances Thermotolerance in Tobacco Plants. FEBS Lett. 2004, 557, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Krishna, P.; Gloor, G. The Hsp90 Family of Proteins in Arabidopsis Thaliana; Cell Stress Society International: Storrs, CT, USA, 2001; Volume 6. [Google Scholar]
- Sun, X.; Zhu, J.; Li, X.; Li, Z.; Han, L.; Luo, H. AsHSP26.8a, a Creeping Bentgrass Small Heat Shock Protein Integrates Different Signaling Pathways to Modulate Plant Abiotic Stress Response. BMC Plant Biol. 2020, 20, 184. [Google Scholar] [CrossRef] [PubMed]
- Heckathorn, S.A.; Downs, C.A.; Sharkey, T.D.; Coleman, J.S. The Small, Methionine-Rich Chloroplast Heat-Shock Protein Protects Photosystem II Electron Transport during Heat Stress. Plant Physiol. 1998, 116, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Neta-Sharir, I.; Isaacson, T.; Lurie, S.; Weiss, D. Dual Role for Tomato Heat Shock Protein 21: Protecting Photosystem II from Oxidative Stress and Promoting Color Changes during Fruit Maturation. Plant Cell 2005, 17, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Sun, Y.; Sun, A.Q.; Yi, S.Y.; Qin, J.; Li, M.H.; Liu, J. The Involvement of Chloroplast HSP100/ClpB in the Acquired Thermotolerance in Tomato. Plant Mol. Biol. 2006, 62, 385–395. [Google Scholar] [CrossRef]
- Song, H.; Zhao, R.; Fan, P.; Wang, X.; Chen, X.; Li, Y. Overexpression of AtHsp90.2, AtHsp90.5 and AtHsp90.7 in Arabidopsis Thaliana Enhances Plant Sensitivity to Salt and Drought Stresses. Planta 2009, 229, 955–964. [Google Scholar] [CrossRef]
- Bernfur, K.; Rutsdottir, G.; Emanuelsson, C. The Chloroplast-Localized Small Heat Shock Protein Hsp21 Associates with the Thylakoid Membranes in Heat-Stressed Plants. Protein Sci. 2017, 26, 1773–1784. [Google Scholar] [CrossRef] [PubMed]
- Helm, K.W.; Lafayette, P.R.; Nagao, R.T.; Key, J.L.; Vierling, E. Localization of Small Heat Shock Proteins to the Higher Plant Endomembrane System. Mol. Cell. Biol. 1993, 13, 238–247. [Google Scholar] [PubMed]
- Ukaji, N.; Kuwabara, C.; Takezawa, D.; Arakawa, K.; Yoshida, S.; Fujikawa, S. Accumulation of Small Heat-Shock Protein Homologs in the Endoplasmic Reticulum of Cortical Parenchyma Cells in Mulberry in Association with Seasonal Cold Acclimation 1. Plant Physiol. 1999, 120, 481–489. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bösl, B.; Grimminger, V.; Walter, S. The Molecular Chaperone Hsp104-A Molecular Machine for Protein Disaggregation. J. Struct. Biol. 2006, 156, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. Complexity of the Heat Stress Response in Plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Bao, F.; Huang, X.; Zhu, C.; Zhang, X.; Li, X.; Yang, S. Arabidopsis HSP90 Protein Modulates RPP4-Mediated Temperature-Dependent Cell Death and Defense Responses. New Phytol. 2014, 202, 1320–1334. [Google Scholar] [CrossRef] [PubMed]
- Guy, C.L.; Li, Q.-B. The Organization and Evolution of the Spinach Stress 70 Molecular Chaperone Gene Family. Plant Cell 1998, 10, 539–556. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ranson, N.A.; White, H.E.; Saibil, H.R. Chaperonins. Biochem J. 1998, 333, 233–242. [Google Scholar] [CrossRef]
- Nagaraju, M.; Kumar, A.; Jalaja, N.; Rao, D.M.; Kishor, P.B.K. Functional Exploration of Chaperonin (HSP60/10) Family Genes and Their Abiotic Stress-Induced Expression Patterns in Sorghum Bicolor. Curr. Genomics 2021, 22, 137–152. [Google Scholar] [CrossRef]
- Horváth, I.; Glatz, A.; Nakamoto, H.; Mishkind, M.L.; Munnik, T.; Saidi, Y.; Goloubinoff, P.; Harwood, J.L.; Vigh, L. Heat Shock Response in Photosynthetic Organisms: Membrane and Lipid Connections. Prog. Lipid Res. 2012, 51, 208–220. [Google Scholar] [CrossRef]
- Nover, L.; Scharf, K.D. Heat Stress Proteins and Transcription Factors. Cell. Mol. Life Sci. 1997, 53, 80–103. [Google Scholar] [CrossRef] [PubMed]
- Sadura, I.; Janeczko, A. Brassinosteroids and the Tolerance of Cereals to Low and High Temperature Stress: Photosynthesis and the Physicochemical Properties of Cell Membranes. Int. J. Mol. Sci. 2022, 23, 342. [Google Scholar] [CrossRef] [PubMed]
- Altschuler, M.; Mascarenhas, J.P. Heat Shock Proteins and Effects of Heat Shock in Plants. Plant Mol. Biol. 1982, 1, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Moisyadi, S.; Harrington, H.M. Characterization of the Heat Shock Response in Cultured Sugarcane Cells: I. Physiology of the Heat Shock Response and Heat Shock Protein Synthesis. Plant Physiol. 1989, 90, 1156–1162. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, I.; Dumas, C. Influence of Temperature Stress on in Vitro Fertilization and Heat Shock Protein Synthesis in Maize (Zea mays L.) Reproductive Tissues. Plant Physiol. 1990, 94, 665–670. [Google Scholar] [CrossRef] [PubMed]
- DeRocher, A.E.; Helm, K.W.; Lauzon, L.M.; Vierling, E. Expression of a Conserved Family of Cytoplasmic Low Molecular Weight Heat Shock Proteins during Heat Stress and Recovery. Plant Physiol. 1991, 96, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Hopf, N.; Plesofsky-Vig, N.; Brambl, R. The Heat Shock Response of Pollen and Other Tissues of Maize. Plant Mol. Biol. 1992, 19, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.B.; Haskell, D.W.; Guy, C.L. Coordinate and Non-Coordinate Expression of the Stress 70 Family and Other Molecular Chaperones at High and Low Temperature in Spinach and Tomato. Plant Mol. Biol. 1999, 39, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Dhaubhadel, S.; Chaudhary, S.; Dobinson, K.F.; Krishna, P. Treatment with 24-Epibrassinolide, a Brassinosteroid, Increases the Basic Thermotolerance of Brassica Napus and Tomato Seedlings. Plant Mol. Biol. 1999, 40, 333–342. [Google Scholar] [CrossRef]
- Ahn, Y.J.; Claussen, K.; Zimmerman, J.L. Genotypic Differences in the Heat-Shock Response and Thermotolerance in Four Potato Cultivars. Plant Sci. 2004, 166, 901–911. [Google Scholar] [CrossRef]
- Pavli, O.I.; Ghikas, D.V.; Katsiotis, A.; Skaracis, G.N. Differential Expression of Heat Shock Protein Genes in Sorghum (Sorghum bicolor L.) Genotypes under Heat Stress. Aust. J. Crop Sci. 2011, 5, 511–515. [Google Scholar]
- Yan, S.P.; Zhang, Q.Y.; Tang, Z.C.; Su, W.A.; Sun, W.N. Comparative Proteomic Analysis Provides New Insights into Chilling Stress Responses in Rice. Mol. Cell. Proteom. 2006, 5, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Heidarvand, L.; Maali Amiri, R. What Happens in Plant Molecular Responses to Cold Stress? Acta Physiol. Plant 2010, 32, 419–431. [Google Scholar] [CrossRef]
- Ambroise, V.; Legay, S.; Guerriero, G.; Hausman, J.F.; Cuypers, A.; Sergeant, K. The Roots of Plant Frost Hardiness and Tolerance. Plant Cell Physiol. 2020, 61, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and Drought Stresses in Crop Plants: Implications, Cross Talk, and Potential Management Opportunities. Front. Plant Sci. 2018, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature Response of Photosynthesis in C3, C4, and CAM Plants: Temperature Acclimation and Temperature Adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Shi, Y.; Yang, S. Advances and Challenges in Uncovering Cold Tolerance Regulatory Mechanisms in Plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef] [PubMed]
- Timperio, A.M.; Egidi, M.G.; Zolla, L. Proteomics Applied on Plant Abiotic Stresses: Role of Heat Shock Proteins (HSP). J. Proteomics 2008, 71, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Gong, F.; Cheminant, S.; Alioua, M.; Hedden, P.; Genschika, P. The Cold-Inducible CBF1 Factor-Dependent Signaling Pathway Modulates the Accumulation of the Growth-Repressing DELLA Proteins via Its Effect on Gibberellin Metabolism. Plant Cell 2008, 20, 2117–2129. [Google Scholar] [CrossRef]
- Zhou, M.; Chen, H.; Wei, D.; Ma, H.; Lin, J. Arabidopsis CBF3 and DELLAs Positively Regulate Each Other in Response to Low Temperature. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Thomashow, M.F. Role of Cold-Responsive Genes in Plant Freezing Tolerance. Plant Physiol. 1998, 118, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K. Cold Stress Tolerance Mechanisms in Plants. A Review. Agron. Sustain. Dev. 2010, 30, 515–527. [Google Scholar] [CrossRef]
- Nievola, C.C.; Carvalho, C.P.; Carvalho, V.; Rodrigues, E. Rapid Responses of Plants to Temperature Changes. Temperature 2017, 4, 371–405. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shi, X.; Chen, S.; Ma, C.; Xu, S. Evolutionary Origin, Gradual Accumulation and Functional Divergence of Heat Shock Factor Gene Family with Plant Evolution. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Koyro, H.-W.; Ahmad, P.; Geissler, N. Abiotic Stress Responses in Plants: An Overview. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M., Eds.; Springer: New York, NY, USA, 2012; pp. 1–28. ISBN 978-1-4614-0815-4. [Google Scholar]
- Eremina, M.; Unterholzner, S.J.; Rathnayake, A.I.; Castellanos, M.; Khan, M.; Kugler, K.G.; May, S.T.; Mayer, K.F.X.; Rozhon, W.; Poppenberger, B. Brassinosteroids Participate in the Control of Basal and Acquired Freezing Tolerance of Plants. Proc. Natl. Acad. Sci. 2016, 113, 5982–5991. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.S.; Cho, E.J.; Choi, E.Y.; Park, O.K. Analysis of the Arabidopsis Nuclear Proteome and Its Response to Cold Stress. Plant J. 2003, 36, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, Y.; Uemura, M. Mass Spectrometric Approach for Identifying Putative Plasma Membrane Proteins of Arabidopsis Leaves Associated with Cold Acclimation. Plant J. 2003, 36, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Kagale, S.; Divi, U.K.; Krochko, J.E.; Keller, W.A.; Krishna, P. Brassinosteroid Confers Tolerance in Arabidopsis Thaliana and Brassica Napus to a Range of Abiotic Stresses. Planta 2007, 225, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.; Sung, D.-Y.; Haskell, D.; Riad, G.S.; Popp, M.; Amaya, M.; LaBoon, A.; Kawamura, Y.; Tominaga, Y.; Kopka, J.; et al. Could Ethanolic Fermentation During Cold Shock Be a Novel Plant Cold Stress Coping Strategy? In Plant Cold Hardiness: From the Laboratory to the Field; Gusta, L.V., Wisniewski, M.E., Tanino, K.K., Eds.; CAB International: Wallingford, UK, 2009; pp. 80–90. [Google Scholar]
- Sewelam, N.; Kazan, K.; Hüdig, M.; Maurino, V.G.; Schenk, P.M. The ATHSP17.4c1 Gene Expression Is Mediated by Diverse Signals That Link Biotic and Abiotic Stress Factors with Ros and Can Be a Useful Molecular Marker for Oxidative Stress. Int. J. Mol. Sci. 2019, 20, 3201. [Google Scholar] [CrossRef]
- Barrero-Gil, J.; Huertas, R.; Rambla, J.L.; Granell, A.; Salinas, J. Tomato Plants Increase Their Tolerance to Low Temperature in a Chilling Acclimation Process Entailing Comprehensive Transcriptional and Metabolic Adjustments. Plant Cell Environ. 2016, 39, 2303–2318. [Google Scholar] [CrossRef]
- Qi, C.; Dong, D.; Li, Y.; Wang, X.; Guo, L.; Liu, L.; Dong, X.; Li, X.; Yuan, X.; Ren, S.; et al. Heat Shock-induced Cold Acclimation in Cucumber through CsHSFA1d-activated JA Biosynthesis and Signaling. Plant J. 2022, 111, 85–102. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Tan, R.; Zhao, J. Chilling Tolerance in Maize: Insights into Advances—Toward Physio-Biochemical Responses’ and QTL/Genes’ Identification. Plants 2022, 11, 2082. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Muhammad, I.; Lan, H.; Xia, C. Recent Advances in the Analysis of Cold Tolerance in Maize. Front. Plant Sci. 2022, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.V.; Li, Q.-B.; Haskell, D.W.; Guy, C.L. Structural Organization of the Spinach Endoplasmic Reticulum - Luminal 70- Kilodalton Heat - Shock Cognate Gene and Expression of 70-Kilodalton Heat-Shock Genes during Cold Acclimation. Plant Physiol. 1994, 104, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.L.; Heazlewood, J.L.; Day, D.A.; Millar, A.H. Differential Impact of Environmental Stresses of the Pea Mitochondrial Proteome. Mol. Cell. Proteom. 2005, 4, 1122–1133. [Google Scholar] [CrossRef] [PubMed]
- Degand, H.; Faber, A.M.; Dauchot, N.; Mingeot, D.; Watillon, B.; Van Cutsem, P.; Morsomme, P.; Boutry, M. Proteomic Analysis of Chicory Root Identifies Proteins Typically Involved in Cold Acclimation. Proteomics 2009, 9, 2903–2907. [Google Scholar] [CrossRef] [PubMed]
- Dumont, E.; Bahrman, N.; Goulas, E.; Valot, B.; Sellier, H.; Hilbert, J.L.; Vuylsteker, C.; Lejeune-Hénaut, I.; Delbreil, B. A Proteomic Approach to Decipher Chilling Response from Cold Acclimation in Pea (Pisum sativum L.). Plant Sci. 2011, 180, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Kubienová, L.; Sedlářová, M.; Vítečková-Wünschová, A.; Piterková, J.; Luhová, L.; Mieslerová, B.; Lebeda, A.; Navrátil, M.; Petřivalský, M. Effect of Extreme Temperatures on Powdery Mildew Development and Hsp70 Induction in Tomato and Wild Solanum spp. Plant Prot. Sci. 2013, 49, 41–54. [Google Scholar] [CrossRef]
- Sabehat, A.; Weiss, D.; Lurie, S. The Correlation between Heat-Shock Protein Accumulation and Persistence and Chilling Tolerance in Tomato Fruit. Plant Physiol. 1996, 110, 531–537. [Google Scholar] [CrossRef]
- Ru, L.; Jiang, L.; Wills, R.B.H.; Golding, J.B.; Huo, Y.; Yang, H.; Li, Y. Chitosan Oligosaccharides Induced Chilling Resistance in Cucumber Fruit and Associated Stimulation of Antioxidant and HSP Gene Expression. Sci. Hortic. 2020, 264, 109187. [Google Scholar] [CrossRef]
- Cabané, M.; Calvet, P.; Vincens, P.; Boudet, A.M. Characterization of Chilling-Acclimation-Related Proteins in Soybean and Identification of One as a Member of the Heat Shock Protein (HSP 70) Family. Planta 1993, 190, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Krishna, P.; Sacco, M.; Cherutti, J.F.; Hill, S. Cold-Induced Accumulation of Hsp90 Transcripts in Brassica Napus. Plant Physiol. 1995, 107, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Pareek, A.; Singla, S.L.; Grover, A. Immunological Evidence for Accumulation of Two High-Molecular-Weight (104 and 90 KDa) HSPs in Response to Different Stresses in Rice and in Response to High Temperature Stress in Diverse Plant Genera. Plant Mol. Biol. 1995, 29, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Huang, F.; Wang, J.; Ma, X.; Cheng, Y.; Liu, J. A Proteomic Analysis of Cold Stress Responses in Rice Seedlings. Proteomics 2005, 5, 3162–3172. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-G.; Ahsan, N.; Lee, S.-H.; Lee, J.J.; Bahk, J.D.; Kang, K.Y.; Lee, B.H. Chilling Stress-Induced Proteomic Changes in Rice Roots. J. Plant Physiol. 2009, 166, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Renaut, J.; Planchon, S.; Oufir, M.; Hausman, J.-F.; Hoffman, L.; Evers, D. Identification of Proteins from Potato Leaves Submitted to Chilling Temperature. In Plant Cold Hardiness: From the Laboratory to the Field; Gusta, L.V., Wisniewski, M.E., Tanino, K.K., Eds.; CAB International: Wallingford, UK, 2009; pp. 279–292. [Google Scholar]
- Vítámvás, P.; Prášil, I.T.; Kosová, K.; Planchon, S.; Renaut, J. Analysis of Proteome and Frost Tolerance in Chromosome 5A and 5B Reciprocal Substitution Lines between Two Winter Wheats during Long-Term Cold Acclimation. Proteomics 2012, 12, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Hlaváčková, I.; Vítámvás, P.; Šantrůček, J.; Kosová, K.; Zelenková, S.; Prášil, I.; Ovesná, J.; Hynek, R.; Kodíček, M. Proteins Involved in Distinct Phases of Cold Hardening Process in Frost Resistant Winter Barley (Hordeum vulgare L.) Cv Luxor. Int. J. Mol. Sci. 2013, 14, 8000–8024. [Google Scholar] [CrossRef] [PubMed]
- Sadura, I.; Libik-Konieczny, M.; Jurczyk, B.; Gruszka, D.; Janeczko, A. HSP Transcript and Protein Accumulation in Brassinosteroid Barley Mutants Acclimated to Low and High Temperatures. Int. J. Mol. Sci. 2020, 21, 1889. [Google Scholar] [CrossRef] [PubMed]
- Stachurska, J.; Sadura, I.; Rys, M.; Dziurka, M.; Janeczko, A. Insight into Hormonal Homeostasis and the Accumulation of Selected Heat Shock Proteins in Cold Acclimated and Deacclimated Winter Oilseed Rape (Brassica napus L.). Agriculture 2023, 13, 641. [Google Scholar] [CrossRef]
- Sadura, I.; Pociecha, E.; Dziurka, M.; Oklestkova, J.; Novak, O.; Gruszka, D.; Janeczko, A. Mutations in the HvDWARF, HvCPD and HvBRI1 Genes-Involved in Brassinosteroid Biosynthesis/Signalling: Altered Photosynthetic Efficiency, Hormonal Homeostasis and Tolerance to High/Low Temperatures in Barley. J. Plant Growth Regul. 2019, 38, 1062–1081. [Google Scholar] [CrossRef]
- Bourgine, B.; Guihur, A. Heat Shock Signaling in Land Plants: From Plasma Membrane Sensing to the Transcription of Small Heat Shock Proteins. Front. Plant Sci. 2021, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The Plant Heat Stress Transcription Factor (Hsf) Family: Structure, Function and Evolution. Biochim. Biophys. Acta-Gene Regul. Mech. 2012, 1819, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Dhaubhadel, S.; Browning, K.S.; Gallie, D.R.; Krishna, P. Brassinosteroid Functions to Protect the Translational Machinery and Heat-Shock Protein Synthesis Following Thermal Stress. Plant J. 2002, 29, 681–691. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).