Abstract
The flower buds of Zingiber striolatum Diels are considered a special vegetable in China, and they are rich in anthocyanins. However, the detailed composition types and the molecular mechanism of anthocyanin biosynthesis in Z. striolatum flower buds are still unclear. In this study, targeted metabolites were used to analyze and identify the anthocyanin types of Z. striolatum in three planting modes: monoculture (CK), intercropping with maize (ZP), and intercropping with soybean (SP). A total of 48 anthocyanins were identified with significant differential accumulation in Z. striolatum flower buds. Among them, cyanidin-3-O-glucoside was the main composition type of anthocyanins. Furthermore, the composition types of blue anthocyanin were identified in flower buds. A total of 15 structure genes were obtained from the transcriptome database of Z. striolatum flower buds. The qRT-PCR results revealed that the expression levels of ZsC4H-1, ZsC4H-2, ZsCHS-2, ZsCHI, ZsF3H, ZsF3′H, ZsDFR, ZsF3′5′H-3, and ZsANS genes were the highest in the ZP model. This study showed that the ZP model contributes to anthocyanin synthesis and accumulation of Z. striolatum flower buds among the three planting modes of Z. striolatum. These findings provide valuable information for research on the planting model and anthocyanin biosynthesis in Z. striolatum flower buds.
1. Introduction
Zingiber striolatum Diels, belonging to the Ginger family, is an herbaceous perennial root plant that has been cultivated nationwide in China in regions such as Guizhou, Yuannan, and Jiangsu [1,2]. The flower buds of Z. striolatum are considered a unique vegetable in China and are rich in anthocyanins [3]. The Z. striolatum roots are also used in traditional Chinese medicine for the treatment of constipation and diabetes and pain relief and detoxification [4,5,6,7].
Anthocyanins are important secondary metabolites that are widely distributed in many plants, such as tomato, onion, cabbage, purple carrot, blood orange, and red-flesh apple [8,9]. Anthocyanins protect plants from damage caused by high light irradiation, high temperature, low temperature, pathogens, and free radical scavengers produced in cells [10,11,12]. In higher plants, anthocyanins play crucial roles in flower color and attracting insects for pollination [13,14]. Anthocyanins are important nutrients and medicines for human health [15,16,17]. Furthermore, anthocyanins can be used as selection markers for breeding and biological research [18,19]. In the plant kingdom, anthocyanins are synthesized from phenylalanine through a biosynthetic pathway [20]. Anthocyanin biosynthesis can be divided into three main phases. The first step, which is the phenylpropanoid pathway, involves the conversion of phenylalanine to cinnamic acid to coumaric acid to 4-coumaroyl CoA, and these reactions are catalyzed by phenylalanine ammonia-lyase (PAL), cinnamate-4-hydroxylase (C4H), and 4-coumaroyl CoA ligase (4CL), respectively [20,21,22]. The second step, which is the flavonoid pathway, involves the conversion of 4-coumaroyl CoA to chalcone and naringein to dihydroflavonol, and these reactions are catalyzed by chalcone synthase (CHS), chalcone isomerase (CHI), and flavanone 3-hydroxylase (F3H), respectively [19,20,21,22,23]. The final step, which is the anthocyanin pathway, involves the conversion of dihydroflavonols to leucoanthocyanidins to anthocyanidins and the further modification of anthocyanidins; these reactions are catalyzed by dihydroflavonol reductase (DFR), anthocyanidin synthase (ANS, also called leucoanthocyanidin dioxygenase), and cyanidin 3-O-galactosyltransferase (3GT) [21,22,23]. The biosynthetic pathway for anthocyanins was first identified from Arabidospis thaliana leaf tissues in 2002 [24]. This biosynthesis pathway has been studied in numerous plant species, such as carrot [25], A. thaliana [26], sweet cherry [27], blood-flesh peach [28], and herbaceous peony [29].
The anthocyanins are divided into six categories in plants: petunidin, delphinidin, malvidin, peonidin, cyanidin, and pelargonidin [30]. The difference in anthocyanin type and content leads to the different color patterns in plants, such as fruit skins, fleshes, leaves, and so on [8,9]. In the flowers of Pericallis hybrida, the cultivars with pink flowers mainly accumulate pelargonidin, the cultivars with red flowers mainly contain cyanidin, and the cultivars with blue flowers mainly comprise delphinidin [13,31]. Chai et al. collected and analyzed the type and content of anthocyanins in 74 blueberry varieties and showed that peonidin-3-glucoside mainly had anthocyanin content in blueberry fruit, but the content of monomeric compounds was different among blueberry varieties [32]. At the same time, the anthocyanin synthesis was also affected by gene regulation, such as Viola tricolor VtF3′5′H and Iris tectorum ItDFR genes, which were overexpressed, and the expression of endogenous RcDFR gene was inhibited in R. chinensis, leading to the production of blue–purple varieties of R. chinensis [33]. Similarly, blue chrysanthemum varieties with high delphinidin content have been obtained by introducing the CmF3′5′H gene of Campanula medium and the CtUA3′5′GT gene of Clitoria ternatea into chrysanthemum [34].
Z. striolatum is a shady crop plant that likes to live in a cool environment. In a monoculture, direct sunlight can affect its growth and development. In recent years, intercropping with maize or soybeans has provided a cool environment for the growth of Z. striolatum, improving the yield of Z. striolatum flower buds and the content of anthocyanins in the Qixingguan District of Bijie City, China. To date, no insights have been made available on anthocyanin biosynthesis in Z. striolatum. In this study, we performed metabolite profiling and transcriptome analysis to explore the composition types, content, and molecular mechanism of anthocyanin biosynthesis in the flower buds of Z. striolatum among three cropping modes (monoculture (CK), intercropping with maize (ZP), and intercropping with soybean (SP)). The targeted metabolites were used to analyze and identify the anthocyanin types, and the expression levels of structural genes in anthocyanin biosynthesis in Z. striolatum were verified by the quantitative real-time polymerase chain reaction (qRT-PCR).
2. Materials and Methods
2.1. Plant Material and Sampling
Qixingguan District of Bijie City (27.622° N, 105.411° E, altitude 1320.600 m), the largest cultivated area of Z. striolatum in Guizhou Province was selected to adopt three planting modes, namely Z. striolatum monoculture (CK), intercropping with maize (ZP), and intercropping with soybean (SP), each planting pattern was repeated 3 times, 5 hectares each repeat. The spacing was 120 cm × 80 cm, with about 20,000~21,000 maize plants per hectare in ZP. The spacing was 120 cm × 20 cm, about 80,000~84,000 soybean plants per hectare in SP. Maize and soybeans were planted directly on 3 May 2023. Z. striolatum flower buds were harvested on 29 July 2023 (8:00 AM–9:00 AM) and sampled during the commodity period. Three biological replicates were set for each sample, and 20 flower buds were taken for each repetition, repeated 3 times. After harvesting, water was used to clean the dirt of Z. striolatum flower buds. Portions of the samples were vacuum freeze-dried for targeted metabolomics analysis. Some were immediately frozen in liquid nitrogen and then stored at −80 °C for RNA extraction and determination of total anthocyanins content.
2.2. Targeted Metabolite Profiling Analysis
The sample was freeze-dried and ground into powder (30 Hz, 1.5 min) using a ball mill (MM400, Retsch, Retsch-Allee, Germany) and stored at −80 °C. 0.05 mg powder was weighed and extracted with 0.5 mL methanol/water/hydrochloric acid (500:500:1, v/v/v). Then, the extract was vortexed for 5 min, subjected to ultrasound for 5 min, and centrifuged at 12,000 rpm at 4 °C for 3 min (5424R, Eppendorf, Hamburg, Germany). The residue was re-extracted by repeating the above-mentioned steps under the same conditions. The supernatants were collected and filtrated through a hydrophilic nylon membrane filter with a pore size of 0.22 μm before LC-MS/MS analysis (Q-Trap 6500+, SCIEX, Framingham, MA, USA).
UPLC conditions: The sample extracts were analyzed using a UPLC-ESI-MS/MS system (UPLC, ExionLC™ AD, https://sciex.com.cn/ (accessed on15 February 2024); MS, Applied Biosystems 6500 Triple Quadrupole, https://sciex.com.cn/ (accessed on 15 February 2024)). The analytical conditions were as follows: UPLC: column, Waters ACQUITY BEH C18 (1.7 μm, 2.1 mm × 100−1 mm); solvent system A, water (0.1% formic acid, Sigma-Aldrich, Waltham, MA, USA): solvent system B, methanol (0.1% formic acid); gradient program, 95:5 v/v at 0 min, 50:50 v/v at 6 min, 5:95 v/v at 12 min, hold for 2 min, 95:5 v/v at 14 min, hold for 2 min; flow rate, 0.35 mL·min−1; temperature, 40 °C; injection volume, 2 μL.
The mass spectrometry conditions mainly include: The temperature of the electrospray ionization (ESI) source was 550 °C, the mass spectrum voltage in positive ion mode was 5500 V, and the Curtain Gas (CUR) was 35 psi. In Q-Trap 6500+, each ion pair is scanned and detected based on optimized declustering potential (DP) and collision energy (CE).
2.3. Qualitative and Quantitative Analysis of Metabolites
Based on the standard substance, the MWDB database (Metware Database) was constructed to qualitatively analyze the mass spectrometry data. Anthocyanin was analyzed using scheduled multiple reaction monitoring (MRM). First, the precursor ions of the target substance were screened by the four-stage bar, and the corresponding ions of other molecular weight substances were preliminarily excluded. The precursor ions were ionized by the collision chamber to form multiple fragment ions, and the fragment ions were triple-filtered to select the required characteristic fragment ions to eliminate the interference of non-target ions. After obtaining the mass spectrometry data of different samples, the chromatographic peaks of all targets were integrated and quantitatively analyzed using a standard curve. Data acquisitions were performed using Analyst 1.6.3 software (Sciex). Multiquant 3.0.3 software (Sciex) was used to quantify all metabolites.
2.4. Data Processing and Evaluation
Data preprocessing: The mass spectrum data were processed using the Analyst 1.6.3 software. MultiQuant 3.0.3 software was used to process the mass spectrum data, reference the retention time and peak type information of the standards, and the chromatographic peaks detected in different samples were corrected integrally to ensure the accuracy of qualitative and quantitative analysis.
Standard curve: To prepare a standard solution of 0.01, 0.05, 0.1, 0.5, 1.0, 5.0, 10, 50, 100, 500, 1000, 2000, and 5000 ng·mL−1 for anthocyanins analysis, the peak intensity data of the corresponding quantitative signals were obtained, and the standard curves of different substances were drawn with the reference concentration as the horizontal coordinate and the peak area as the vertical coordinate.
Calculation of anthocyanin content: The integral peak area of all the samples was put into the linear equation of the standard curve and then put into the calculation formula. Contents of metabolites in the sample (μg·g−1) = C × V × 1,000,000 m−1. C: Concentration value (ng·mL−1) of the area of the integral peak in the sample substituted for the standard curve; V: volume (mL) of the solution used at the time of extraction; m: the sample quality (g). The coefficient of variation was conducted using the Empirical Cumulative Distribution Function (ECDF, https://www.mathworks.com/help/stats/ecdf.html (accessed on 20 February 2024)).
2.5. Total Anthocyanin Content Determination
The total anthocyanin content of flower buds was determined in accordance with Xu et al. method [25]. The determination of total anthocyanin content in Z. striolatum flower buds among three planting modes was conducted as previously described. Fresh samples of Z. striolatum flower buds were ground into powder using liquid nitrogen. Not less than 2 g powder was weighed and placed into test tubes, added with 20 mL 0.1% methanol, and extracted for 8 h in dark conditions at room temperature. Three biological replicates were set for each sample. The extract was filtered using a 0.25 µm filter, and total anthocyanin quantities were reported in mg cyanidin 3-O-galactoside equivalents per g FW (mg·g−1 g, FW). Values were the means of three independent experiments.
2.6. Transcriptome Analysis and Genes Identification
According to the metabolic pathway for anthocyanins, anthocyanin-related gene sequences were obtained for the transcriptome database of Z. striolatum constructed by our group. The open reading frame and the amino acid sequence of these genes were obtained using BioXM2.7.1 software. Then, the amino acid sequences were compared with that of Zingiber officinale using the NCBI database (BioSample: SAMN15647981) to predict the correctness and integrity of the gene sequences. Finally, the gene sequence information of Z. striolatum involved in the anthocyanidin biosynthesis is listed in Table S1.
2.7. RNA Extraction and Quantitative RT-PCR (qRT-PCR)
The total RNA of flower bud samples (harvested between 8:00 AM and 9:00 AM, 29 July 2023) at three cropping patterns was isolated using TaKaRa MiniBEST Plant RNA Extraction Kit. One microgram of total RNA was used for reverse transcription into cDNA using the First Strand cDNA Synthesis Kit (Nuode Biotech., Beijing, China). qRT-PCR primers for these genes encoding the key enzymes associated with the anthocyanin biosynthesis are listed in Table S2. qRT-PCR was conducted using SYBR Premix Ex Tag (TaKaRa, San Jose, CA, USA) and cDNA as the template. Results were analyzed using the 2−ΔΔCT method with ZsActin gene expression level as an internal reference. The ZsActin gene is a gene that is stably expressed during flower development based on our study (Unpublished). Three biological and three technical replicates were set in this study.
2.8. Statistical Analysis
Significant analysis was calculated using ANOVA (Analysis of variance), and Pearson’s correlation was used for correlation analysis of the anthocyanin content and expression levels of anthocyanin-synthesis-related genes. Microsoft Excel 2007 and Origin 2021 software were used for data processing and mapping, respectively.
3. Results
3.1. Flower Buds of Z. striolatum in Three Planting Modes
Three main planting modes of Z. striolatum in Guizhou Province were considered, namely Z. striolatum monoculture (CK) (Figure 1(A1)), intercropping with maize (ZP) (Figure 1(B1)), and intercropping with soybean (SP) (Figure 1(C1)). In the three planting modes, the commodity organs and quality of Z. striolatum were significantly different. In CK, few flower buds were found in the roots (Figure 1(A2)), and the number of flower buds in a single plant was 7.87, and the commodity flower buds were small (single flower bud weight was 25.12 g, and the flower bud size was only 4.51 cm × 2.11 cm, Table 1), and had obvious chlorophyll accumulation at the top of Z. striolatum flower buds (Figure 1(A3)). The Z. striolatum roots produced abundant flower buds (Figure 1(B2)), the number of flower buds in single plant was 11.62, and the commodity flower buds were large (single flower bud weight was 35.08 g, and the flower bud size was only 4.51 cm × 2.69 cm, Table 1), and the anthocyanins was accumulation obvious (Figure 1(B3)) in ZP (0.26 mg·g−1, FW, Table 1). The number of root flower buds of SP was less than that of ZP and higher than that of CK (Figure 1(C2)), and the commodity flower buds were medium, but the anthocyanin was significantly lower than that of CK and ZP (Figure 1(C3), Table 1). The highest yield of flower buds in ZP was 1384.13 kg·667 m−2, the flower buds medium yield was 1179.22 kg/667 m2 in SP, the lowest flower buds was 1094.66 kg/667 m2 in the CK (Table 1).

Figure 1.
The field, root flower buds, and commodity flower buds of Z. striolatum under three planting modes. (A1): CK; (B1): ZP; (C1): SP; (A2): Root flower buds of CK; (B2): Root flower buds of ZP; (C2): Root flower buds of SP; (A3): Commodity flower buds of CK; (B3): Commodity flower buds of ZP; (C3): Commodity flower buds of SP.

Table 1.
The information on Z. striolatum flower buds among three planting modes.
3.2. Types and Total Anthocyanin Content in Flower Buds among Three Planting Modes
To explore the types of anthocyanins in the Z. striolatum flower buds, 48 anthocyanins were identified from Z. striolatum flower buds by targeted metabolomics. They were divided into 6 categories (Table 2). Among them, there were 5 petunidin, 9 delphinidin, 6 malvidin, 7 peonidin, 17 cyanidin, and 4 pelargonidin. The metabolomics analysis results showed that the anthocyanins components in the Z. striolatum flower buds were complex and diverse. Furthermore, anthocyanin composition types of blue anthocyanins were also identified in flower buds, such as 9 delphinidin types, 5 petunidin types, and 6 malvidin types.
Table 2.
The anthocyanin types and content of Z. striolatum flower buds among three planting modes. Anthocyanin quantities were reported in μg cyanidin 3-O-galactoside equivalents per g DW (μg·g−1 g, DW).
The content of anthocyanin components was determined in three planting modes by targeted metabolomics. The results revealed that 48 anthocyanin components were detected in CK, 7 anthocyanin components (delphinidin-3-O-(6-O-malonyl-beta-D-glucoside),delphinidin-3-O-(6-O-malonyl)-glucoside-3′-glucoside, malvidin-3-O-rhamnoside, cyanidin-3-O-rhamnoside, cyanidin-rutinoside-rhamnoside, pelargonidin-3-O-(6″-O-acetyl)galactoside, pelargonidin-3-O-galactoside) were detected in ZP, and 12 anthocyanin components (delphinidin-3-O-(6″-O-tartaryl)glucoside, delphinidin-caffeoyl-rutinoside, malvidin-3-O-(6″-O-acetyl)galactoside, peonidin-3,5-O-diglucoside, peonidin-3-O-(6″O-acetyl-malonyl)glucoside, peonidin-3-O-arabinoside, peonidin-3-O-sambubioside-5-O-glucoside, peonidin-hydroxyben-malonyl-glucoside-xyloside, cyanidin-3,5-O-diglucoside, cyanidin-3-O-(6″-O-acetyl-2″-O-xylosyl)glucoside, cyanidin-3-O-(tartaryl)rhamnoside-5-O-glucoside, pelargonidin-3-O-sambubioside) were detected in SP.
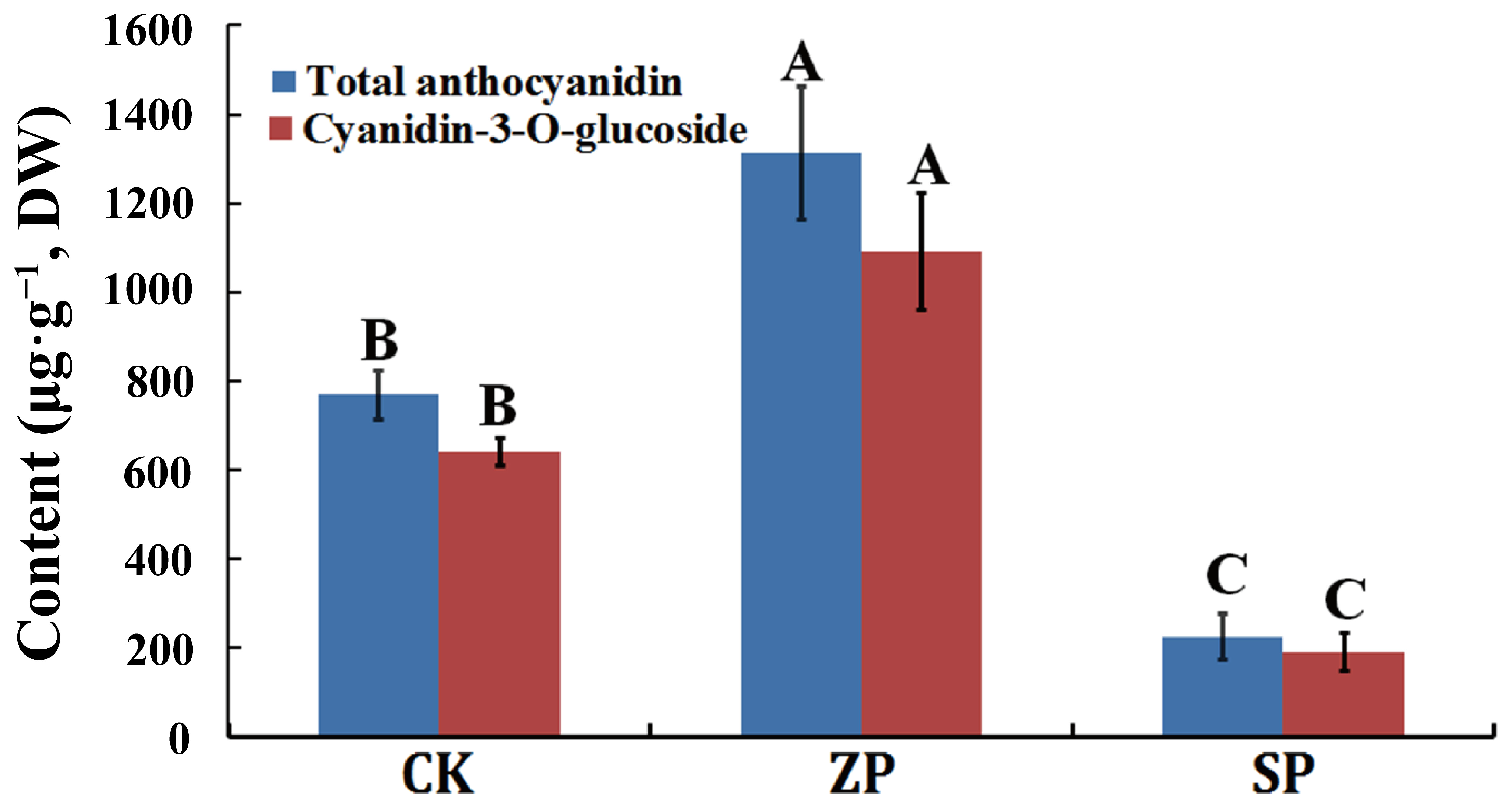
Among the three planting modes, the anthocyanin content was the highest (1312.9663 μg g−1, DW) in ZP, followed by CK (768.8993 μg g−1, DW), and the anthocyanin content was the lowest (224.8324 μg g−1, DW) in SP (Table 2). At the same time, cyanidin-3-O-glucoside content was the highest among all kinds of anthocyanin in the three planting modes. The content of total anthocyanin and cyanidin-3-O-glucoside were analyzed in the three planting modes, and the results indicated that the total anthocyanin content was consistent with the changing trend of cyanidin-3-O-glucoside in the three planting modes (Figure 2). In general, the planting mode of ZP was conducive to the accumulation of anthocyanin, and cyanidin-3-O-glucoside was the main anthocyanin component in the Z. striolatum flower buds.
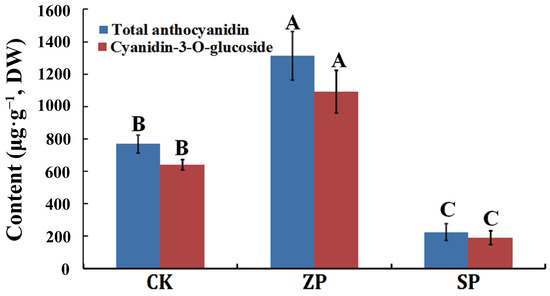
Figure 2.
The content of total anthocyanin and cyanidin-3-O-glucoside. The capitalized A, B, and C indicated a significant difference (p < 0.01) in this figure.
3.3. Transcriptome Analysis and Gene Identification
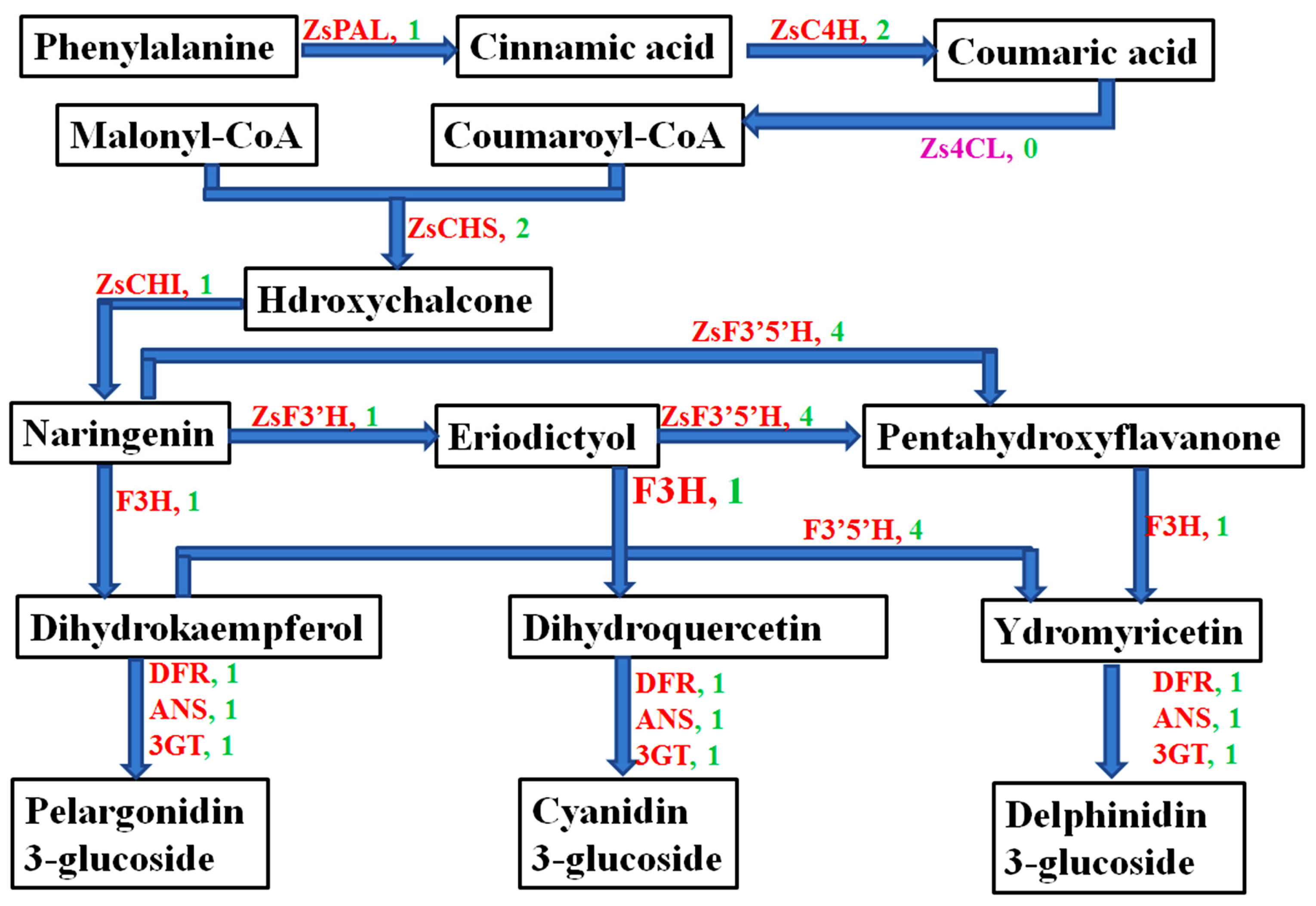
We obtained 15 anthocyanin-related structure genes from the transcriptome database of Z. striolatum, which were 1 ZsPAL, 2 ZsC4H, 2 ZsCHS, 1 ZsCHI, 4 ZsF3′5′H, 1 ZsF3′H, 1 ZsF3′H, 1 ZsDFR, 1 ZsANS, and 1 Zs3GT (Figure 3). The retrieved genes were deduced into corresponding amino acid sequences, and the amino acid sequences were compared with those of the Z. officinale plant using the NCBI online website. The results showed that the obtained anthocyanin-related gene sequences were complete, the consistency of gene sequences with Z. officinale plant was high, and the consistency level was between 88% and 99% (Table 3).
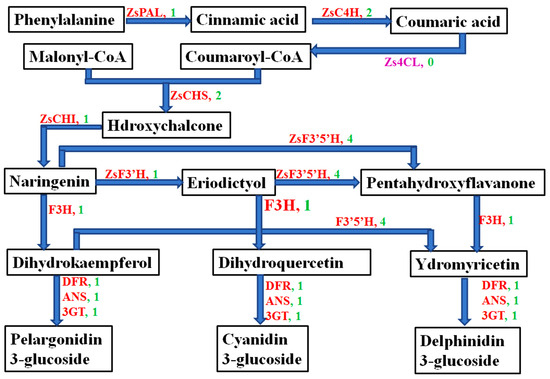
Figure 3.
Anthocyanin metabolic pathway and anthocyanin-related structure gene number of Z. striolatum. PAL: Phenylalanine ammonia-lyase; C4H: Cinnamate 4-hydroxylase; 4CL: 4-coumarate-CoA ligase; CHS: Chalcone synthesis; CHI: Chalcone isomerase; F3H: Flavanone-3 hydroxylase; F3′H: Flavonoid 3′-hydroxylase; F3′5′H: Flavonoid-3′,5′-hydroxylase; DFR: Dihydroflavonol 4-reductase; ANS: Anthocyanidin synthase; 3GT: UDP-flavonoid 3-O-glucosyItransferase. The numbers of genes are marked in green.
Table 3.
The information on anthocyanin biosynthesis genes in Z. striolatum.
3.4. Expression Level of Anthocyanin-Related Genes in Three Planting Modes of Z. striolatum
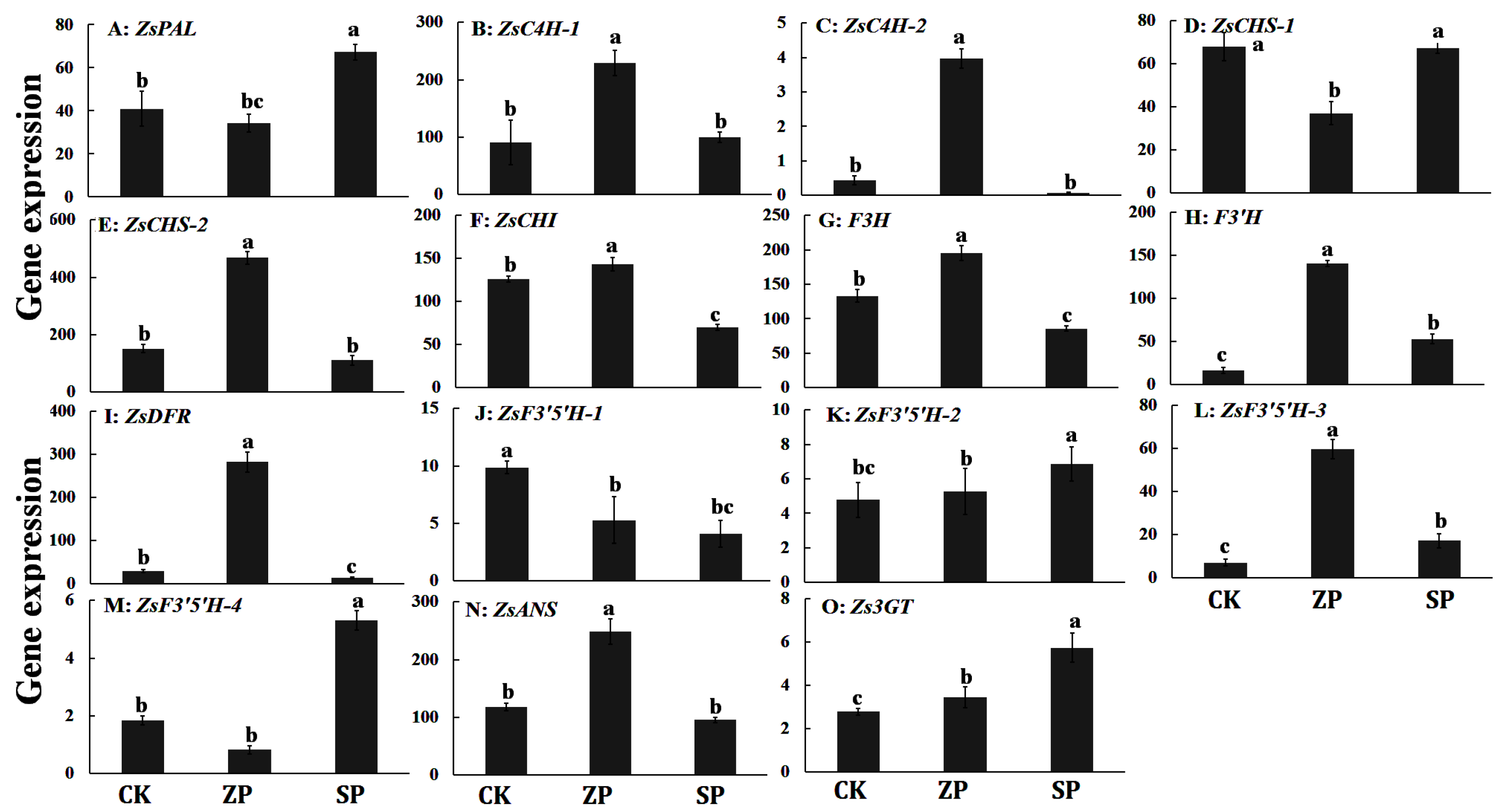
We detected the gene expression levels of anthocyanin-related genes under three planting modes of Z. striolatum by qPCR. The results indicated that the expression level of anthocyanin-related genes was significantly different in three planting modes of Z. striolatum. The anthocyanin-related genes were expressed in three planting modes. The expression levels of ZsC4H-2 and ZsDFR genes were nearly 0 in SP, and the expression levels were the highest in ZP. The expression levels of ZsPAL, ZsF3′5′H-2, ZsF3′5′H-4, and Zs3GT genes in SP were significantly higher than those in ZP and CK. The expression levels of ZsC4H-1, ZsC4H-2, ZsCHS-2, ZsCHI, ZsF3H, ZsF3′H, ZsDFR, ZsF3′5′H-3, and ZsANS genes in ZP were significantly higher than those in the two other planting modes of Z. striolatum. Only the expression level of the ZsF3′5′H-3 gene was significantly higher than that in ZP and SP. The expression level of the ZsCHS-1 gene in CK and SP was basically the same, and the expression levels of both genes were significantly higher than that in ZP (Figure 4).
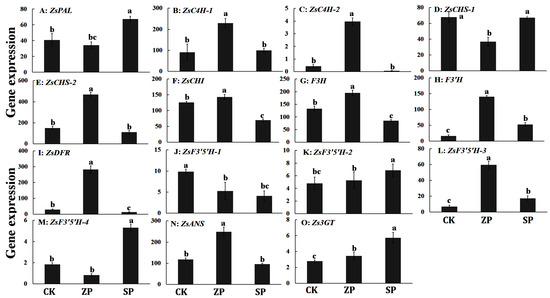
Figure 4.
The expression level of anthocyanin-related genes of Z. striolatum in three planting modes. Different lowercase letters indicate the significance of the difference (p < 0.05).
3.5. Correlation Analysis Anthocyanin of Content and Expression Levels of Anthocyanin-Synthesis-Related Genes
The relationship between anthocyanin content and the expression level of anthocyanin-synthesis-related genes was further defined through correlation analysis. The correlation results (Table S3) showed that the total anthocyanin content was significantly positively correlated with the expression levels of ZsC4H-1, ZsC4H-2, ZsCHS-2, ZsCHI, ZsF3H, ZsDFR, ZsF3′H, ZsF3′5′H-3, and ZsANS genes. Meanwhile, it was significantly negatively correlated with the expression levels of ZsPAL, ZsCHS-1, and ZsF3′5′H-4 genes. The total anthocyanin content was positively correlated with the expression level of the ZsF3′5′H-1 gene, while it was negatively correlated with the expression level of the ZsF3′5′H-2 gene. Petunidin-3-O-(6-O-p-coumaroyl)-glucoside, delphinidin-3-O-(6-O-malonyl-beta-D-glucoside), delphinidin-3-O-(6-O-malonyl)-glucoside-3′-glucoside, malvidin-3-O-(6″-O-acetyl) glucoside, malvidin-3-O-rhamnoside, cyanidin-3-O-rhamnoside, cyanidin-rutinoside-rhamnoside, pelargonidin-3-O-(6″-O-acetyl) galactoside, and pelargonidin-3-O-galactoside showed an opposite relationship to the correlation between total anthocyanin content and the expression levels of anthocyanin structure genes, while the remaining anthocyanin monomer remained consistent.
4. Discussion
We selected the purple Z. striolatum with the largest cultivation area in Guizhou Province as the experimental material to explore the best planting method for improving the yield and quality of Z. striolatum. Moreover, three planting methods—CK, ZP, and SP—were adopted. The results showed significant differences in yield and anthocyanin accumulation of Z. striolatum among the three planting patterns. Analysis of the three aspects of flower bud germination, commercial flower bud size, and anthocyanin accumulation of Z. striolatum indicated that the germination rate of Z. striolatum flower buds in the root was high, the commodity flower buds was large, and the anthocyanin accumulation was obvious, which improved the yield and anthocyanin accumulation of Z. striolatum. This phenomenon was directly related to light. On the one hand, the planting mode of SP can increase canopy photosynthesis [35,36], and the Z. striolatum flower buds prefer to live in shady and cool environments [37]. On the other hand, anthocyanins can accelerate the degradation of anthocyanins in light [38,39]. In this study, maize could better shade for the Z. striolatum growth and increase its flower bud yield and anthocyanins content, indicating that ZP can provide a cool and shaded light environment for the growth of Z. striolatum.
The composition and content of anthocyanin are the main factors affecting the coloration of Z. striolatum flower buds. In this study, 48 anthocyanins were identified in the Z. striolatum flower buds by the combination of targeted metabolomics for the first time. A total of 17 cyanidins were detected in the Z. striolatum flower buds, among which the cyanidin-3-O-glucoside content was the highest, which may be the main pigment type in Z. striolatum flower buds. Cyanidin was the main pigment that leads to purple color in many horticultural crops [40,41]. Notably, 7 peonidins were detected in CK and ZP, while only 2 peonidins were detected in SP. The result indicated that the planting mode of SP was detrimental to the synthesis and accumulation of peonidin, which may be related to the nitrogen fixation of leguminous crops. Nitrogen is an important regulator of anthocyanin synthesis and accumulation in plants. In general, plants can promote anthocyanin synthesis under low nitrogen conditions, while they inhibit anthocyanin synthesis and accumulation under high nitrogen conditions [42,43,44].
The biosynthesis pathway for anthocyanins has been studied in various model crops, such as Arabidopsis thaliana, and was highly conserved in many plants [45]. ZsCHS, ZsCHI, ZsF3H, ZsF3′H, ZsF3′5′H, ZsDFR, ZsANS, and Zs3GT genes were the key genes affecting the coloration of Z. striolatum flower buds. These genes were significantly upregulated in Z. striolatum flower buds, which promote anthocyanin biosynthesis. F3H was an important branch point in the anthocyanin biosynthesis pathway, and the ZsF3H gene was significantly related to the total anthocyanin content of Z. striolatum flower buds. Thus, it may be one of the key genes leading to the anthocyanin difference of Z. striolatum flower buds [46]. DFR, ANS, and 3GT genes were downstream genes of the biosynthesis pathway for anthocyanins. Flavonols were catalyzed to colorless anthocyanin by dihydroflavonol-4-reductase, which was further catalyzed to form the colored anthocyanins by anthocyanidin synthase [47,48]. The expression levels of ZsDFR and ZsANS genes in ZP were significantly higher than those in CK and SP, which was consistent with the anthocyanins content in the three planting modes. Colored anthocyanins were catalyzed to stabilize anthocyanins using UDP-flavonoid 3-O-glucosyItransferase [49]. The results revealed that the expression level of the Zs3GT gene was significantly higher in ZP than in the two other planting modes, which indicated that the anthocyanin of Z. striolatum flower buds was more stable and difficult to degrade. Studies have found that F3′H and F3′5′H genes are key genes in the biosynthesis pathways of red and blue anthocyanins, respectively [50]. Blue anthocyanins were also detected in Z. striolatum flower buds in the present study. Therefore, Z. striolatum may contain abundant modified gene resources for breeding red and blue Z. striolatum flower buds.
In this study, we identified 15 structural genes from the transcriptome database of Z. striolatum according to the biosynthesis pathway for anthocyanins. Intriguingly, the Zs4CL gene was not aligned. We also found similar situations in Dendrobium [51], red okra [52], and Cymbidium orchid [53]. We speculate that the biosynthesis pathway for anthocyanins in Z. striolatum flower buds may be the same as most crops and affected by light-induced [54,55,56]. Therefore, the biosynthesis pathway for anthocyanins in Z. striolatum flower buds still needs to be further improved.
5. Conclusions
We adopted three planting modes of Z. striolatum, including Z. striolatum monoculture (CK), intercropping with maize (ZP), and intercropping with soybean (SP). The results showed that the commodity flower buds were large, and the anthocyanin content was high in the planting mode of ZP, which could effectively improve the yield and quality of Z. striolatum flower buds, the content of cyanidin-3-O-glucoside was the highest, and the main type of anthocyanins in Z. striolatum flower buds. These findings provide valuable information for the research of the planting model of Z. striolatum flower buds.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14071414/s1, Table S1. The sequence information of genes involved in the anthocyanidin biosynthesis; Table S2. Primers used in this study; Table S3. Correlation of anthocyanin contents and anthocyanin-related genes relative expression levels in three planting modes.
Author Contributions
D.Z., T.W. and G.T. conceived and designed the experiment; D.Z. analyzed the data; D.Z., T.W., Q.Z. and G.T. contributed reagents/materials/analysis tools; D.Z. performed the experiments; D.Z. and Q.Z. wrote the paper: D.Z. and G.T. revised the paper. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by the Qiankehe Service Enterprise [2022] 005, Qianke Hezhong Land Diversion [2023] 033), Construction of Modern Agricultural Industry Technology System in Guizhou Province Vegetable System Project (GZCYTX2024-0101), and Guizhou Province Plateau Characteristic Vegetable Industry Technology System (GZMARS).
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Huang, Z.B.; Xie, L.N.; Wang, H.W.; Zhong, J.B.; Li, Y.C.; Liu, J.L.; Ou, Z.R.; Liang, X.; Li, Y.S.; Huang, H.Y.; et al. Geographic distribution and impacts of climate change on the suitable habitats of Zingiber species in China. Ind. Crops Prod. 2019, 138, e111429. [Google Scholar] [CrossRef]
- Tian, S.M.; Jiang, D.Z.; Wan, Y.P.; Wang, X.; Liao, Q.H.; Li, Q.; Li, H.L.; Liao, L.Z. The complete chloroplast genome of Zingiber striolatum Diels (Zingiberaceae). Mitochondrial DNA Part B Resour. 2023, 8, 48–51. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.C.; He, Y.X.; He, H.Y.; Chen, X.Q.; Liu, T.F.; Zhu, B. Wild vs. cultivated Zingiber striolatum Diels: Nutritional and biological activity differences. Plants 2023, 12, e2180. [Google Scholar] [CrossRef]
- Huang, Z.B.; Xie, L.N.; Xu, Y.Y.; Zhao, K.; Li, X.T.; Zhong, J.B.; Lu, Y.J.; Xu, X.T.; Goodin, S.; Zhang, K.; et al. Essential oils from Zingiber striolatum Diels attenuate inflammatory response and oxidative stress through regulation of MAPK and NF-κB signaling pathways. Antioxidants 2021, 10, e2019. [Google Scholar] [CrossRef]
- Chen, T.; Cai, J.; Ni, J.; Yang, F. An UPLC-MS/MS application to investigate chemical compositions in the ethanol extract with hypoglycemic activity from Zingiber striolatum Diels. J. Chin. Pharm. Sci. 2016, 25, 116–121. [Google Scholar] [CrossRef]
- Kim, H.W.; Murakami, A.; Abe, M.; Ozawa, Y.; Morimitsu, Y.; Williams, M.V.; Ohigashi, H. Suppressive effects of mioga ginger and ginger constituents on reactive oxygen and nitrogen species generation, and the expression of inducible pro-inflammatory genes in macrophages. Antioxid. Redox Signal. 2005, 7, 1621–1629. [Google Scholar] [CrossRef]
- Tian, M.; Liu, T.; Wu, X.; Hong, Y.; Zhou, Y. Chemical composition, antioxidant, antimicrobial and anticancer activities of the essential oil from the rhizomes of Zingiber striolatum Diels. Nat. Prod. Res. 2020, 34, 2621–2625. [Google Scholar] [CrossRef]
- Chaves-Silva, S.; dos Santos, A.L.; Chalfun-Junior, A.; Zhao, J.; Peres, L.E.P.; Benedito, V.A. Understanding the genetic regulation of anthocyanin biosynthesis in plants—Tools for breeding purple varieties of fruits and vegetables. Phytochem. 2018, 153, 11–27. [Google Scholar] [CrossRef]
- Sharma, H.; Sharma, P.; Kumar, A.; Chawla, N.; Dhatt, A.S. Multifaceted regulation of anthocyanin biosynthesis in plants: A comprehensive review. J. Plant Growth Regul. 2024, 12, 1–15. [Google Scholar] [CrossRef]
- Ren, Y.; Huang, Z.Q.; Jiang, H.; Wang, Z.; Wu, F.S.; Xiong, Y.F.; Yao, J.L. A heat stress responsive NAC transcription factor heterodimer plays key roles in rice grain filling. J. Exp. Bot. 2021, 72, 2947–2964. [Google Scholar] [CrossRef]
- Cui, L.G.; Shan, J.X.; Shi, M.; Gao, J.P.; Lin, H.X. The miR156-SPL9-DFR pathway coordinates the relationship between development and abiotic stress tolerance in plants. Plant J. 2014, 80, 1108–1117. [Google Scholar] [CrossRef]
- Hatier, J.H.B.; Clearwater, M.J.; Gould, K.S. The functional significance of black-pigmented leaves: Photosynthesis, photoprotection and productivity in Ophiopogon planiscapus ‘Nigrescens’. PLoS ONE 2013, 8, e67850. [Google Scholar] [CrossRef]
- Qi, F.T.; Liu, Y.T.; Luo, Y.L.; Cui, Y.M.; Lu, C.F.; Li, H.; Huang, H.; Dai, S.L. Functional analysis of the ScAG and SCAGL11 MADS-box transcription factors for anthocyanin biosynthesis and bicolour pattern formation in Senecio cruentus ray florets. Hortic. Res. 2022, 9, uhac071. [Google Scholar] [CrossRef]
- Baumann, K.; Perez-Rodriguez, M.; Bradley, D.; Venail, J.; Bailey, P.; Jin, H.; Koes, R.; Roberts, K.; Martin, C. Control of cell and petal morphogenesis by R2R3 MYB transcription factors. Development 2007, 134, 1691–1701. [Google Scholar] [CrossRef]
- Shen, H.; Han, J.; Liu, C.L.; Cao, F.; Huang, Y.J. Grape seed proanthocyanidins exert a radioprotective effect on the testes and intestines through antioxidant effects and inhibition of MAPK signal pathways. Front. Med. 2022, 8, e836528. [Google Scholar] [CrossRef]
- Zhou, P.Y.; Zhang, L.M.; Li, W.; Zhang, S.T.; Luo, L.X.; Wang, J.; Sun, B.S. In vitro evaluation of the anti-digestion and antioxidant effects of grape seed procyanidins according to their degrees of polymerization. J. Funct. Foods 2018, 49, 85–95. [Google Scholar] [CrossRef]
- Le, D.H.; Nishimura, K.; Takenaka, Y.; Mizushina, Y.; Tanahashi, T. Polyprenylated benzoylphloroglucinols with DNA polymerase inhibitory activity from the fruits of Garcinia schomburgkiana. J. Nat. Prod. 2016, 79, 1798–1807. [Google Scholar] [CrossRef]
- Yamagishi, M. High temperature enhances anthocyanin coloration in Asiatic hybrid lily flowers via upregulation of the MYB12 positive regulator. Hortic. Plant J. 2022, 8, 769–776. [Google Scholar] [CrossRef]
- Brugliera, F.; Tao, G.Q.; Tems, U.; Kalc, G.; Mouradova, E.; Price, K.; Stevenson, K.; Nakamura, N.; Stacey, I.; Katsumoto, Y.; et al. Violet/blue chrysanthemums: Metabolic engineering of the anthocyanin biosynthetic pathway results in novel petal colors. Plant Cell Physiol. 2013, 54, 1696–1710. [Google Scholar] [CrossRef]
- Feng, X.K.; Gao, G.; Yu, C.M.; Zhu, A.G.; Chen, J.K.; Chen, K.M.; Wang, X.F.; Abubakar, A.S.; Chen, P. Transcriptome and metabolome analysis reveals anthocyanin biosynthesis pathway associated with ramie (Boehmeria nivea (L.) Gaud. leaf color formation. BMC Genom. 2021, 22, e684. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Chu, G.H.; Hu, Z.L.; Gao, Q.; Cui, B.L.; Tian, S.B.; Wang, B.; Chen, G.P. Genetically engineered anthocyanin pathway for high health-promoting pigment production in eggplant. Mol. Breed. 2016, 36, 54. [Google Scholar] [CrossRef]
- Qi, X.W.; Shuai, Q.; Chen, H.; Fan, L.; Zeng, Q.W.; He, N.J. Cloning and expression analyses of the anthocyanin biosynthetic genes in mulberry plants. Mol. Genet. Genom. 2014, 289, 783–793. [Google Scholar] [CrossRef]
- Sunil, L.; Shetty, N.P. Biosynthesis and regulation of anthocyanin pathway genes. Appl. Microbiol. Biotechnol. 2022, 106, 1783–1798. [Google Scholar] [CrossRef]
- Bloor, S.J.; Abrahams, S. The structure of the major anthocyanin in Arabidopsis thaliana. Phytochemistry 2002, 59, 343–346. [Google Scholar] [CrossRef]
- Xu, Z.S.; Huang, Y.; Wang, F.; Song, X.; Wang, G.L.; Xiong, A.S. Transcript profiling of structural genes involved in cyanidin-based anthocyanin biosynthesis between purple and non-purple carrot (Daucus carota L.) cultivars reveals distinct patterns. BMC Plant Biol. 2014, 14, e262. [Google Scholar] [CrossRef]
- Liu, Y.J.; Li, M.; Li, T.T.; Chen, Y.J.; Zhang, L.J.; Zhao, G.F.; Zhuang, J.H.; Zhao, W.Y.; Gao, L.P.; Xia, T. Airborne fungus-induced biosynthesis of anthocyanins in Arabidopsis thaliana via jasmonic acid and salicylic acid signaling. Plant Sci. 2020, 300, e110635. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, C.Q.; Cui, Y.L.; Du, Q.S.; Tang, W.J.; Yang, W.L.; Kou, G.Q.; Tang, W.J.; Chen, H.X.; Gong, R.G. Potential regulatory genes of light induced anthocyanin accumulation in sweet cherry identified by combining transcriptome and metabolome analysis. Front. Plant Sci. 2023, 14, e1238624. [Google Scholar] [CrossRef]
- Jiao, Y.; Ma, R.J.; Shen, Z.J.; Yan, J.; Yu, M.L. Gene regulation of anthocyanin biosynthesis in two blood-flesh peach (Prunus persica (L.) Batsch) cultivars during fruit development. J. Zhejiang Univ. Sci. B 2014, 15, 809–819. [Google Scholar] [CrossRef]
- Wu, Y.Q.; Hao, Z.J.; Tang, Y.H.; Zhao, D.Q. Anthocyanin accumulation and differential expression of the biosynthetic genes result in a discrepancy in the red color of herbaceous peony (Paeonia lactiflora Pall.) flowers. Horticulturae 2022, 8, e349. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Jin, X.H.; Huang, H.; Wang, L.; Sun, Y.; Dai, S.L. Transcriptomics and metabolite analysis reveals the molecular mechanism of anthocyanin biosynthesis branch pathway in different Senecio cruentus cultivars. Front. Plant Sci. 2016, 7, e1307. [Google Scholar] [CrossRef] [PubMed]
- Chai, Z.; Herrera-Balandrano, D.D.; Yu, H.; Beta, T.; Zeng, Q.L.; Zhang, X.X.; Tian, L.L.; Niu, L.Y.; Huang, W.Y. A comparative analysis on the anthocyanin composition of 74 blueberry cultivars from China. J. Food Compos. Anal. 2021, 102, e104051. [Google Scholar] [CrossRef]
- Katsumoto, Y.; Fukuchi-mizutani, M.; Fukui, Y.; Brugliera, F.; Holton, T.; Karan, M.; Nakamura, N.; Yonekura-Sakakibara, K.; Togami, J.; Pigeaire, A.; et al. Engineering of the rose flavonoid biosynthetic pathway successfully generated blue-hued flowers accumulating delphinidin. Plant Cell Physiol. 2007, 48, e1589. [Google Scholar] [CrossRef]
- Noda, N. Recent advances in the research and development of blue flowers. Breed. Sci. 2018, 68, 79–87. [Google Scholar] [CrossRef]
- Wang, Q.S.; Sun, D.B.; Hao, H.; Zhao, X.J.; Hao, W.P.; Liu, Q. Photosynthetically active radiation determining yields for an intercrop of maize with cabbage. Eur. J. Agron. 2015, 69, 32–40. [Google Scholar] [CrossRef]
- Chi, B.J.; Liu, J.; Dai, J.L.; Li, Z.H.; Zhang, D.M.; Xu, S.Z.; Nie, J.J.; Wan, S.M.; Li, C.D.; Dong, H.Z. Alternate intercropping of cotton and peanut increases productivity by increasing canopy photosynthesis and nutrient uptake under the influence of rhizobacteria. Field Crop. Res. 2023, 302, e109059. [Google Scholar] [CrossRef]
- Deng, K.P.; Deng, R.J.; Fan, J.X.; Chen, E.F. Transcriptome analysis and development of simple sequence repeat (SSR) markers in Zingiber striolatum Diels. Physiol. Mol. Biol. Plants 2018, 24, 125–134. [Google Scholar] [CrossRef]
- Cai, D.B.; Li, X.S.; Chen, J.L.; Jiang, X.W.; Ma, X.Q.; Sun, J.X.; Tian, L.M.; Vidyarthi, S.K.; Xu, J.W.; Pan, Z.L.; et al. A comprehensive review on innovative and advanced stabilization approaches of anthocyanin by modifying structure and controling environmental factors. Food Chem. 2022, 366, e130611. [Google Scholar] [CrossRef]
- Jin, W.P.; Xiang, L.; Peng, D.F.; Liu, G.; He, J.R.; Cheng, S.Y.; Li, B.; Huang, Q.R. Study on the coupling progress of thermo-induced anthocyanins degradation and polysaccharides gelation. Food Hydrocoll. 2020, 105, e105822. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, e52. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.H.; Dai, M.Y.; Zheng, G.; Chang, P.J.; Xuan, L.J.; Liu, Z.G.; Wang, Y.L.; Cheng, S.Y.; Wang, Z.; Wang, H.; et al. Flavonold components and gene expression analysis reveal flower pigmentation difference between Magnolia biondi and its variety M. biondi var. purpurascens. Trees 2022, 36, 583–591. [Google Scholar] [CrossRef]
- Rubin, G.; Tohge, T.; Matsuda, F.; Saito, K.; Scheible, W.R. Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. Plant Cell 2009, 21, 3567–3584. [Google Scholar] [CrossRef]
- Zhou, L.L.; Shi, M.Z.; Xie, D.Y. Regulation of anthocyanin biosynthesis by nitrogen in TTG1-GL3/TT8-PAP1- programmed red cells of Arabidopsis thaliana. Planta 2012, 236, 825–837. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Yang, J.; Ma, C.L.; Zhang, Y.; Ge, T.; Qi, Z.; Kang, Y. Arabidopsis ROOT HAIR DEFECTIVE3 is involved in nitrogen starvation-induced anthocyanin accumulation. J. Integr. Plant Biol. 2015, 57, 708–721. [Google Scholar] [CrossRef]
- Grützner, R.; König, K.; Horn, C.; Engler, C.; Laub, A.; Vogt, T.; Marillonnet, S. A transient expression tool box for anthocyanin biosynthesis in Nicotiana benthamiana. Plant Biotechnol. J. 2023, 143, 1238–1250. [Google Scholar] [CrossRef]
- Song, B.; Xu, H.; Chen, L.Z.; Fan, X.X.; Jing, Z.G.; Chen, S.; Xu, Z.G. Study of the relationship between leaf color formation and anthocyanin metabolism among different purple pakchoi lines. Molecules 2020, 25, e4809. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, Y.T.; Wang, H.; Tian, Z.D.; Xin, S.Y.; Zhu, P.F. The dihydroflavonol 4-reductase BoDFR1 drives anthocyanin accumulation in pink-leaved ornamental kale. Theor. Appl. Genet. 2021, 134, 159–169. [Google Scholar] [CrossRef]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef]
- Matsuba, Y.; Sasaki, N.; Tera, M.; Okamura, M.; Abe, Y.; Okamoto, E.; Nakamura, H.; Funabashi, H.; Takatsu, M.; Saito, M.; et al. A novel glucosylation reaction on anthocyanin catalyzed by acyl-glucose-dependent glucosyltransferase in the petals of carnation and delphinium. Plant Cell 2010, 22, 3374–3389. [Google Scholar] [CrossRef]
- Huang, H.; Hu, K.; Han, K.T.; Xiang, Q.Y.; Dai, S.L. Flower colour modification of chrysanthemum by suppression of F3′H and overexpression of the exogenous Senecio cruentus F3′5′H gene. PLoS ONE 2013, 8, e74395. [Google Scholar] [CrossRef]
- Wu, M.Q.; Liao, Y.; Lu, S.J.; Yin, H.T.; Yu, W.G.; Li, C.H. Metabolomics analysis of anthocyanin in different flower colors of phalaenopsis-type Dendrobium (in Chinese with abstract). Chin. J. Trop. Crops 2023, 44, 2167–2178. [Google Scholar] [CrossRef]
- Wang, J.Q.; Zhang, Y.H.; Yi, W.Y.; Gong, Z.G.; Xu, H.B.; Li, Y.Y. Study on genes related to anthocyanin biosynthesis at the color turning stages in red okra fruit based on transcriptome sequencing. N. Hortic. 2022, 14, 9–16. [Google Scholar]
- Wang, L.; Albert, N.W.; Zhang, H.B.; Arathoon, S.; Boase, M.R.; Ngo, H.; Schwinn, K.E.; Davies, K.M.; Lewis, D.H. Temporal and spatial regulation of anthocyanin biosynthesis provide diverse flower colour intensities and patterning in Cymbidium orchid. Planta 2014, 240, 983–1002. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wu, W.S.; Sun, Y.; Shen, Y.Y.; Mao, L.Z.; Dai, Y.H.; Yang, B.Z.; Liu, Z.B. Integrated transcriptome and metabolome analysis reveals anthocyanin biosynthesis mechanisms in pepper (Capsicum annuum L.) leaves under continuous blue light irradiation. BMC Plant Biol. 2024, 24, e10. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Wang, X.; Liu, T.; Liu, H.; Peng, Y.; Tang, X.; Ye, X.; Sun, K.; Yue, Y.; Xu, D.; et al. Epigenetic regulation of high light-induced anthocyanin biosynthesis by histone demethylase IBM1 in Arabidopsis. New Phytol. 2024, 242, 2570–2585. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhong, X.L.; Cai, X.; Zhu, S.H.; Meng, P.H.; Zhang, J.; Tan, G.F. Comparative physiological analysis of lignification, anthocyanin metabolism and correlated gene expression in red Toona sinensis buds during cold storage. Agronomy 2023, 13, e119. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).