


CsNWD Encoding VPS62 Emerges as a Candidate Gene Conferring the Glabrous Phenotype in Cucumber


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Conditions for Growth, and the Identification of Phenotypes
2.2. Genetic Analysis
2.3. Transpiration Rate, Net Photosynthetic Rate, and Chlorophyll Content Determination
2.4. Antioxidant Enzyme Activity Determination
2.5. Soluble Sugars, Malondialdehyde (MDA), and Electrical Conductivity Analysis
2.6. Scanning Electron Microscopy Observation
2.7. BSA-Seq Analysis of nwd Locus
2.8. RNA Extraction and qRT-PCR
2.9. RNA-Seq Analysis
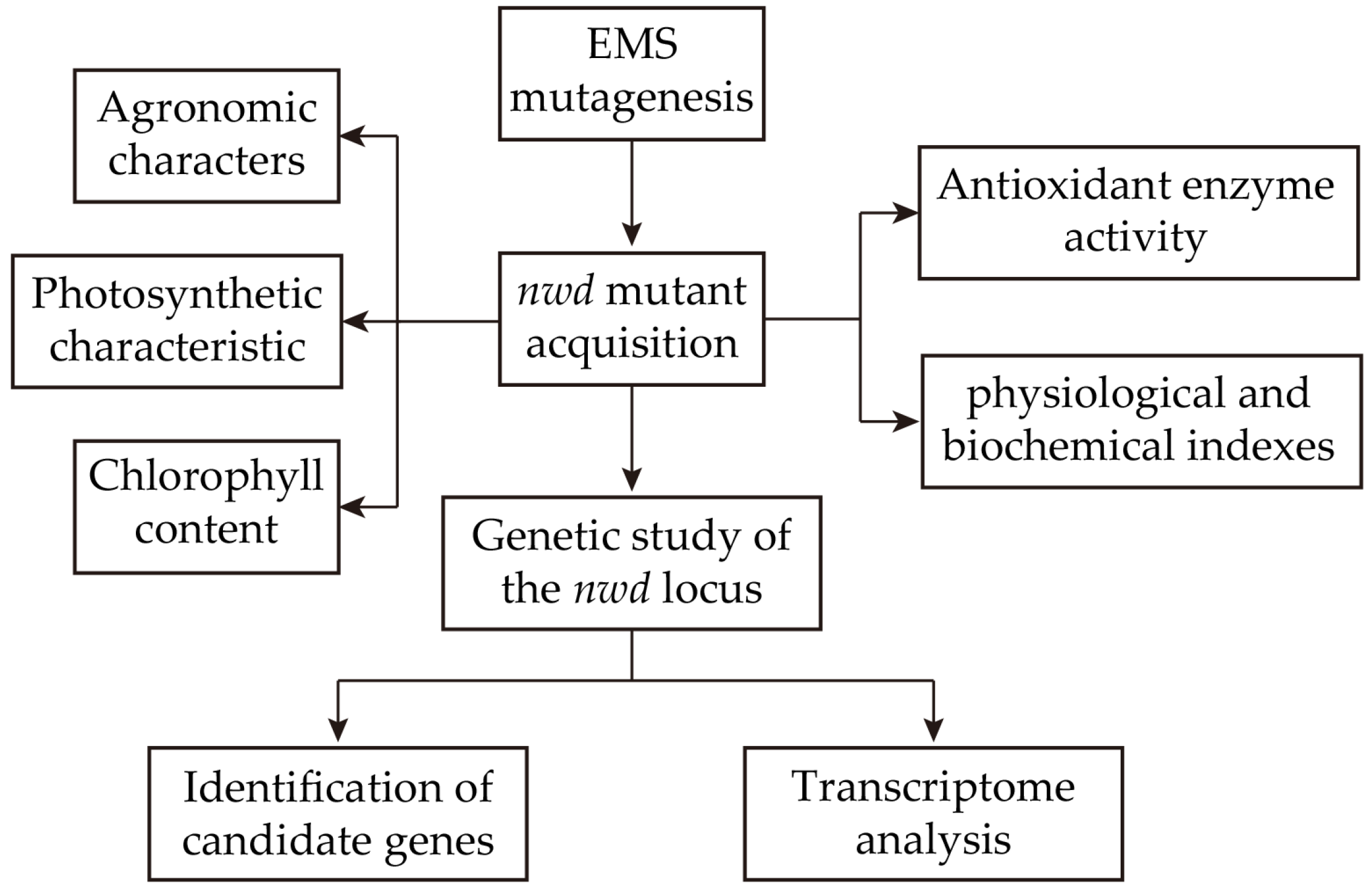
2.10. Statistical Analysis and Flowchart
3. Results
3.1. The EMS Mutant nwd Produced a Glabrous Phenotype
3.2. Inheritance of the nwd Locus
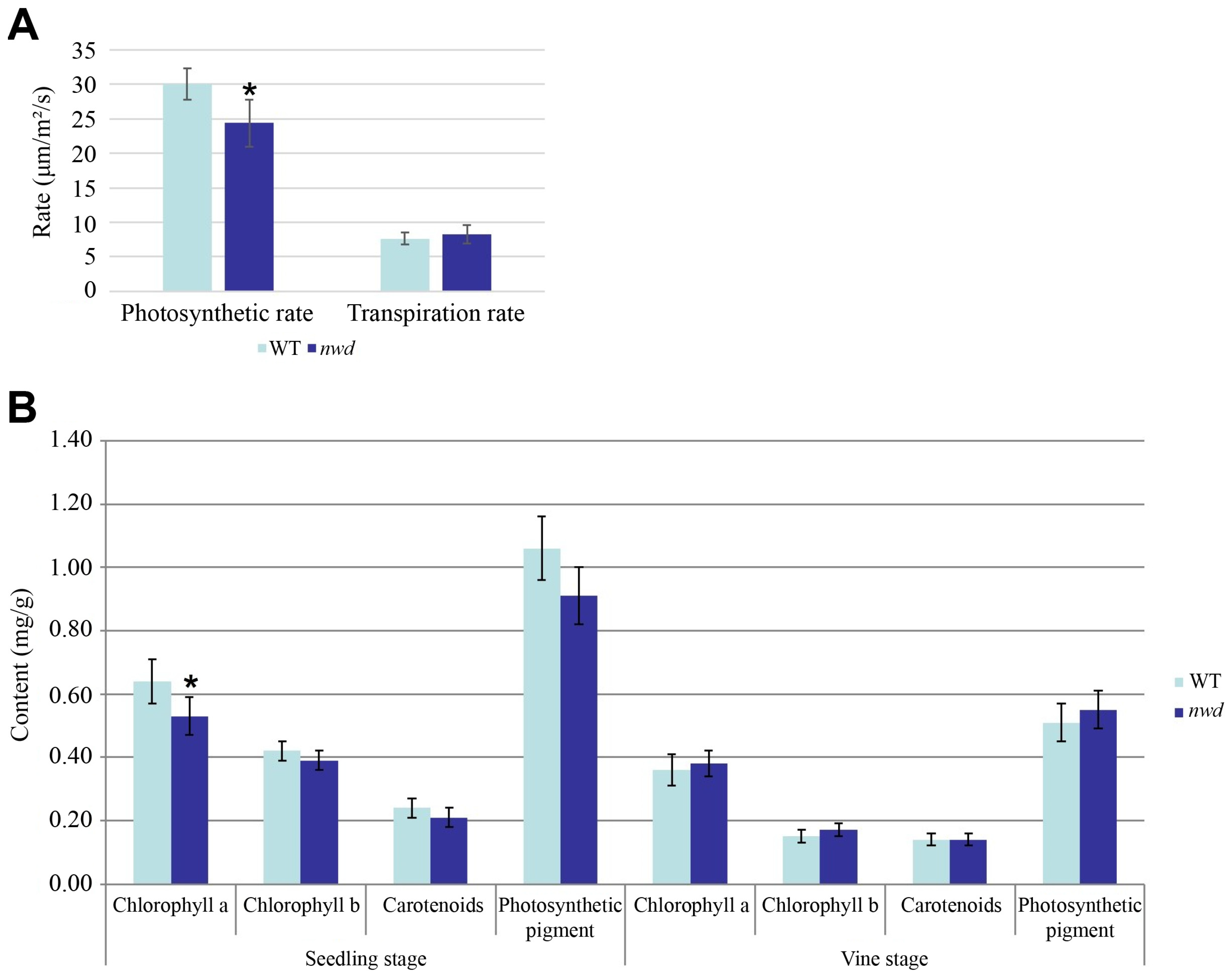
3.3. Determination of Major Agronomic Traits, Photosynthetic Characteristics, and Chlorophyll Content of nwd
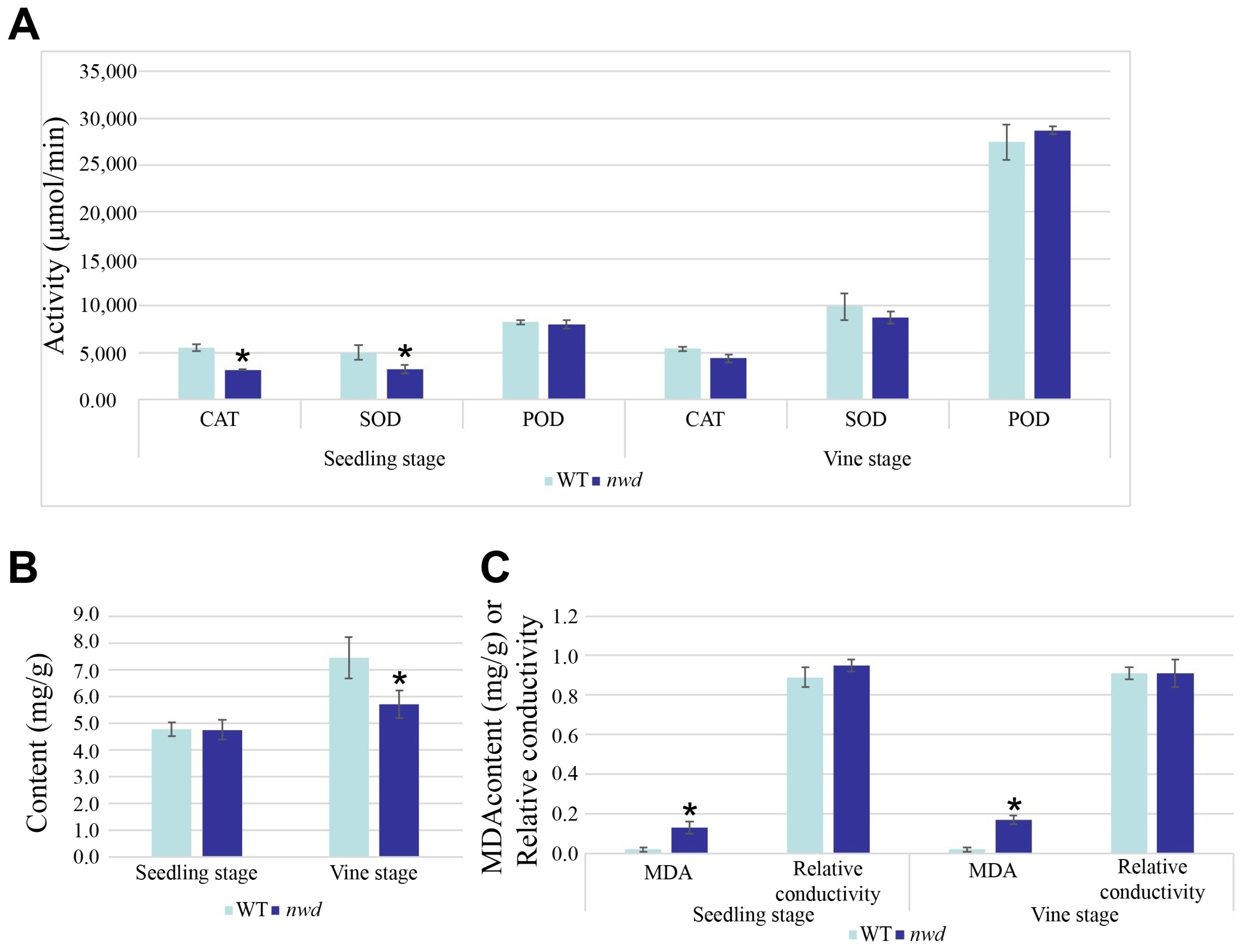
3.4. Changes in Antioxidant Enzyme Activity, Soluble Protein Content, and Malondialdehyde Content in nwd
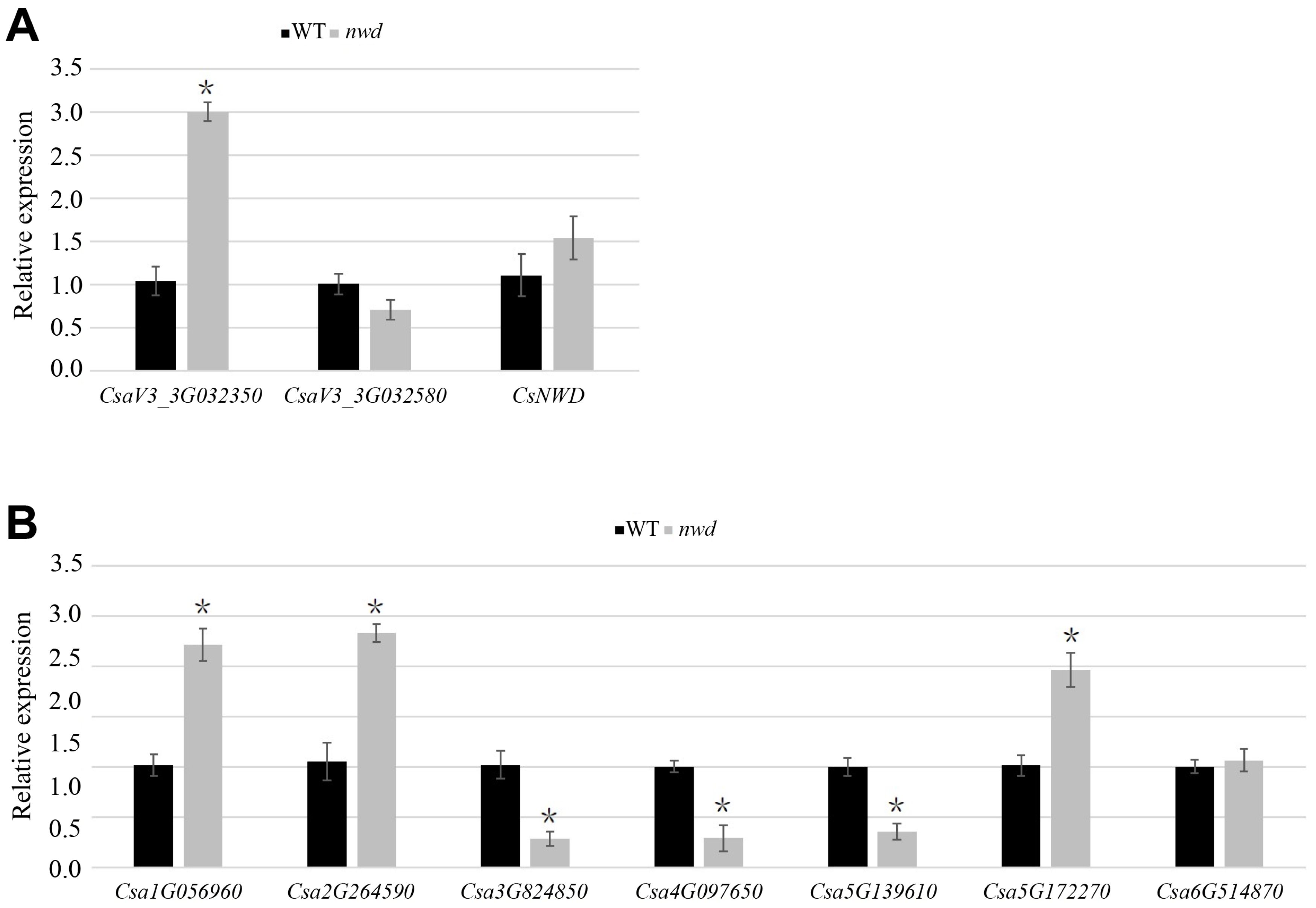
3.5. Identification of Candidate Genes for nwd Mutant
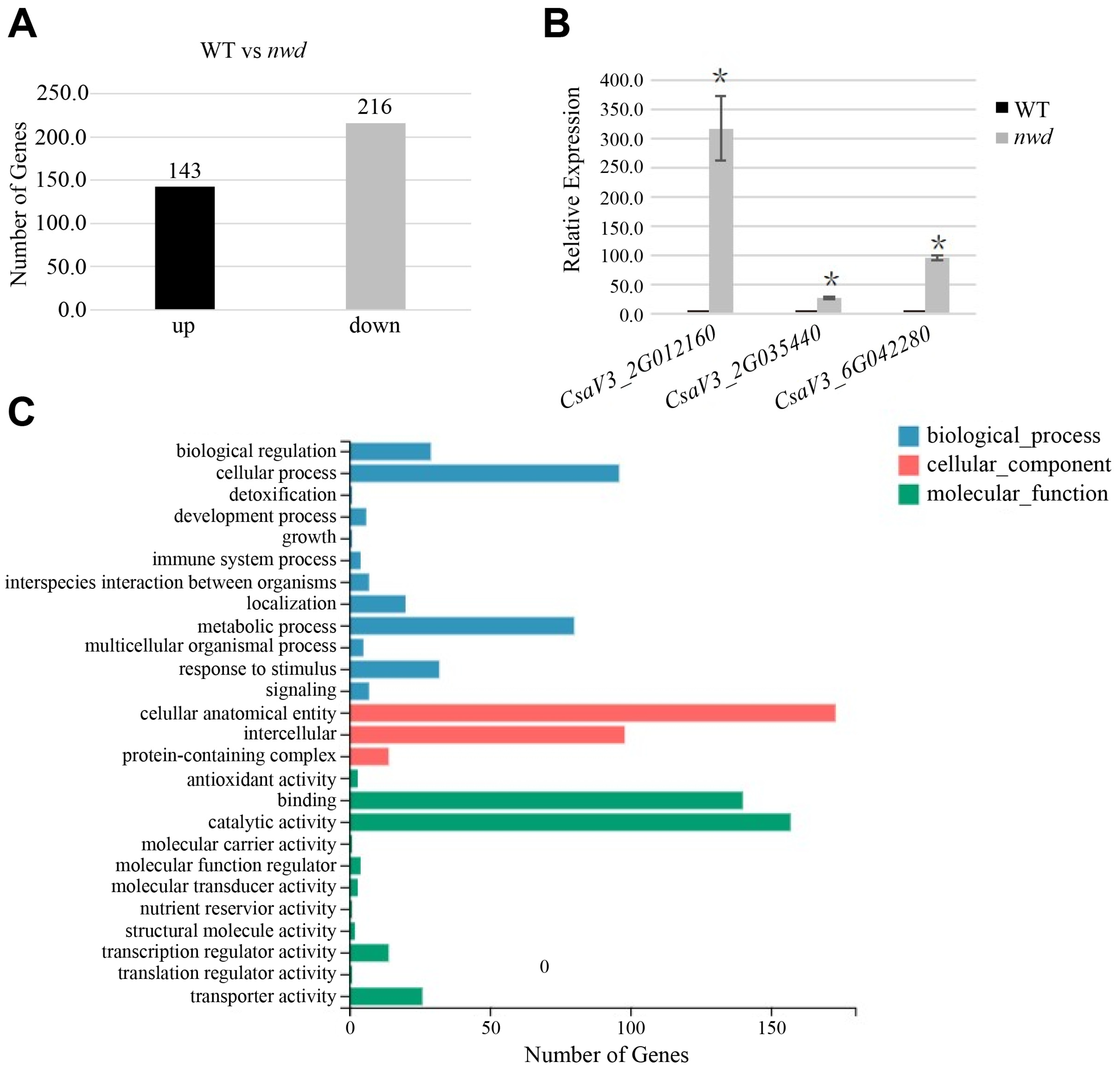
3.6. Quantitative Expression of Trichome-Related Genes in nwd
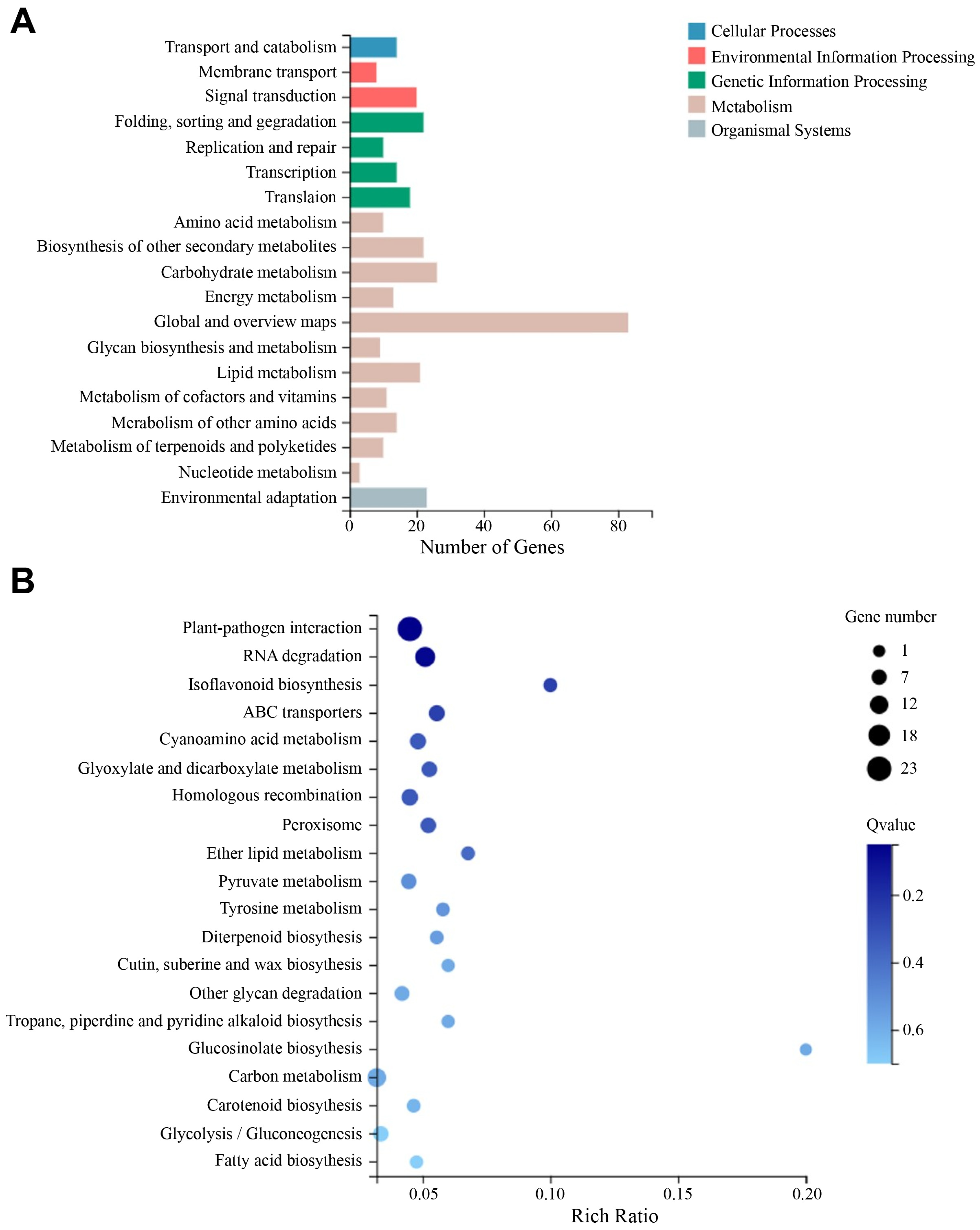
3.7. Comparative Transcriptome Analysis between nwd Mutant and Wild-Type
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dong, Y.; Li, S.; Wu, H.; Gao, Y.; Feng, Z.; Zhao, X.; Shan, L.; Zhang, Z.; Ren, H.; Liu, X. Advances in understanding epigenetic regulation of plant trichome development: A comprehensive review. Hortic. Res. 2023, 10, uhad145. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Li, Y.; Yang, Z.; Wang, C.; Zhang, Y.; Wang, B. Molecular Mechanisms of Plant Trichome Development. Front. Plant Sci. 2022, 13, 910228. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Hulskamp, M.; Schnittger, A.; Folkers, U. Pattern formation and cell differentiation: Trichomes in Arabidopsis as a genetic model system. Int. Rev. Cytol. 1999, 186, 147–178. [Google Scholar] [PubMed]
- Hulskamp, M. Plant trichomes: A model for cell differentiation. Nat. Rev. Mol. Cell Biol. 2004, 5, 471–480. [Google Scholar] [CrossRef]
- Pan, Y.; Bo, K.; Cheng, Z.; Weng, Y. The loss-of-function GLABROUS 3 mutation in cucumber is due to LTR-retrotransposon insertion in a class IV HD-ZIP transcription factor gene CsGL3 that is epistatic over CsGL1. BMC Plant Biol. 2015, 15, 302. [Google Scholar] [CrossRef]
- Xue, S.; Dong, M.; Liu, X.; Xu, S.; Pang, J.; Zhang, W.; Weng, Y.; Ren, H. Classification of fruit trichomes in cucumber and effects of plant hormones on type II fruit trichome development. Planta 2019, 249, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, M.; Jiang, L.; Liu, X.; Zhao, J.; Yan, S.; Yang, S.; Ren, H.; Liu, R.; Zhang, X. Transcriptome profiling reveals roles of meristem regulators and polarity genes during fruit trichome development in cucumber (Cucumis sativus L.). J. Exp. Bot. 2014, 65, 4943–4958. [Google Scholar] [CrossRef]
- Schuurink, R.; Tissier, A. Glandular trichomes: Micro-organs with model status? New Phytol. 2020, 225, 2251–2266. [Google Scholar] [CrossRef] [PubMed]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P. Protective and defensive roles of non-glandular trichomes against multiple stresses: Structure–function coordination. J. For. Res. 2019, 31, 1–12. [Google Scholar] [CrossRef]
- Weng, Y. Cucumis sativus Chromosome Evolution, Domestication, and Genetic Diversity: Implications for Cucumber Breeding. Plant Breed. Rev. 2021, 44, 79–111. [Google Scholar]
- Li, Q.; Cao, C.; Zhang, C.; Zheng, S.; Wang, Z.; Wang, L.; Ren, Z. The identification of Cucumis sativus Glabrous 1 (CsGL1) required for the formation of trichomes uncovers a novel function for the homeodomain-leucine zipper I gene. J. Exp. Bot. 2015, 66, 2515–2526. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.L.; Pan, J.S.; Guan, Y.; Zhang, W.W.; Bie, B.B.; Wang, Y.L.; He, H.L.; Lian, H.L.; Cai, R. Micro-trichome as a class I homeodomain-leucine zipper gene regulates multicellular trichome development in Cucumis sativus. J. Integr. Plant Biol. 2015, 57, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Zhang, L.; Chen, G.; Wen, H.; Chen, Y.; Du, H.; Zhao, J.; He, H.; Lian, H.; Chen, H.; et al. Study of micro-trichome (mict) reveals novel connections between transcriptional regulation of multicellular trichome development and specific metabolism in cucumber. Hortic. Res. 2021, 8, 21. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Nie, J.-T.; Chen, H.-M.; Guo, C.-L.; Pan, J.; He, H.-L.; Pan, J.-S.; Cai, R. Identification and mapping of Tril, a homeodomain-leucine zipper gene involved in multicellular trichome initiation in Cucumis sativus. Theor. Appl. Genet. 2016, 129, 305–316. [Google Scholar] [CrossRef]
- Du, H.; Wang, G.; Pan, J.; Chen, Y.; Xiao, T.; Zhang, L.; Zhang, K.; Wen, H.; Xiong, L.; Yu, Y.; et al. The HD-ZIP IV transcription factor Tril regulates fruit spine density through gene dosage effects in cucumber. J. Exp. Bot. 2020, 71, 6297–6310. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.-Y.; Miao, H.; Ding, L.-H.; Wehner, T.C.; Liu, P.-N.; Wang, Y.; Zhang, S.-P.; Gu, X.-F. A New Glabrous Gene (csgl3) Identified in Trichome Development in Cucumber (Cucumis sativus L.). PLoS ONE 2016, 11, e0148422. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Cai, Y.; Liu, X.; Dong, M.; Zhang, Y.; Chen, S.; Zhang, W.; Li, Y.; Tang, M.; Zhai, X.; et al. A CsMYB6-CsTRY module regulates fruit trichome initiation in cucumber. J. Exp. Bot. 2018, 69, 1887–1902. [Google Scholar] [CrossRef]
- Yang, Z.; Song, M.; Cheng, F.; Zhang, M.; Davoudi, M.; Chen, J.; Lou, Q. A SNP Mutation in Homeodomain-DDT (HD-DDT) Transcription Factor Results in Multiple Trichomes (mt) in Cucumber (Cucumis sativus L.). Genes 2021, 12, 1478. [Google Scholar] [CrossRef]
- Che, G.; Zhang, X. Molecular basis of cucumber fruit domestication. Curr. Opin. Plant Biol. 2019, 47, 38–46. [Google Scholar] [CrossRef]
- Guo, P.; Chang, H.; Li, Q.; Wang, L.; Ren, Z.; Ren, H.; Chen, C. Transcriptome profiling reveals genes involved in spine development during CsTTG1-regulated pathway in cucumber (Cucumis sativus L.). Plant Sci. 2020, 291, 110354. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.S.; Giardina, T. Sucrose and invertases, a part of the plant defense response to the biotic stresses. Front. Plant Sci. 2014, 5, 293. [Google Scholar] [CrossRef] [PubMed]
- Ulrike Oster, R.T.; Tanaka, A.; Rüdiger, W. Cloning and functional expression of the gene encoding the key enzyme for chlorophyll b biosynthesis (CAO) from Arabidopsis thaliana. Plant J. 2000, 21, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Suda, I.; Furuta, S.; Nishiba, Y. Fluorometric Determination of a 1,3-Diethyl-2-thiobarbituric Acid–Malondialdehyde Adduct as an Index of Lipid Peroxidation in Plant Materials. Biosci. Biotechnol. Biochem. 2014, 58, 14–17. [Google Scholar] [CrossRef]
- Waters, B.M.; Chu, H.H.; Didonato, R.J.; Roberts, L.A.; Eisley, R.B.; Lahner, B.; Salt, D.E.; Walker, E.L. Mutations in Arabidopsis yellow stripe-like1 and yellow stripe-like3 reveal their roles in metal ion homeostasis and loading of metal ions in seeds. Plant Physiol. 2006, 141, 1446–1458. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, X.; He, B.; Diao, L.; Sheng, S.; Wang, J.; Guo, X.; Su, N.; Wang, L.; Jiang, L.; et al. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol. 2007, 145, 29–40. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Oppenheimer, D.G.; Herman, P.L.; Sivakumaran, S.; Esch, J.; Marks, M. A myb gene required for leaf trichome differentiation in Arabidopsis is expressed in stipules. Cell 1991, 67, 483–493. [Google Scholar] [CrossRef]
- Walker, A.R.; Davison, P.A.; Bolognesi-Winfield, A.C.; James, C.M.; Srinivasan, N.; Blundell, T.L.; Esch, J.J.; Marks, M.D.; Gray, J.C. The Transparent Testa GLABRA1 Locus, which Regulates Trichome Differentiation and Anthocyanin Biosynthesis in Arabidopsis, Encodes a WD40 Repeat Protein. Plant Cell 1999, 11, 1337–1349. [Google Scholar] [CrossRef]
- Payne, C.T.; Zhang, F.; Lloyd, A.M. GL3 encodes a bHLH protein that regulates trichome development in Arabidopsis through interaction with GL1 and TTG1. Genetics 2000, 156, 1349–1362. [Google Scholar] [CrossRef]
- Schena, M.; Davis, R.W. HD-Zip proteins: Members of an Arabidopsis homeodomain protein superfamily. Proc. Natl. Acad. Sci. USA 1992, 89, 3894–3898. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, J.-W.; Yu, N.; Li, C.-H.; Luo, B.; Gou, J.-Y.; Wang, L.-J.; Chen, X.-Y. Control of Plant Trichome Development by a Cotton Fiber MYB Gene. Plant Cell 2004, 16, 2323–2334. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.Y.; Li, Q.J.; Shan, C.M.; Wang, S.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. The HD-Zip IV gene GaHOX1 from cotton is a functional homologue of the Arabidopsis GLABRA2. Physiol. Plant. 2008, 134, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Payne, T.; Clement, J.; Arnold, D.; Lloyd, A. Heterologous myb genes distinct from GL1 enhance trichome production when overexpressed in Nicotiana tabacum. Development 1999, 126, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Bassham, D.C.; Raikhel, N.V. Unique features of the plant vacuolar sorting machinery. Curr. Opin. Cell Biol. 2000, 12, 491–595. [Google Scholar] [CrossRef]
- Robinson, D.G.; Herranz, M.-C.; Bubeck, J.; Pepperkok, R.; Ritzenthaler, C. Membrane Dynamics in the Early Secretory Pathway. Crit. Rev. Plant Sci. 2007, 26, 199–225. [Google Scholar] [CrossRef]
- Scheuring, D.; Viotti, C.; Kruger, F.; Kunzl, F.; Sturm, S.; Bubeck, J.; Hillmer, S.; Frigerio, L.; Robinson, D.G.; Pimpl, P.; et al. Multivesicular Bodies Mature from the Trans-Golgi Network/Early Endosome in Arabidopsis. Plant Cell 2011, 23, 3463–3481. [Google Scholar] [CrossRef]
- Gao, C.; Zhuang, X.; Shen, J.; Jiang, L. Plant ESCRT Complexes: Moving Beyond Endosomal Sorting. Trends Plant Sci. 2017, 22, 986–998. [Google Scholar] [CrossRef]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. Transparent Testa GLABRA2, a Trichome and Seed Coat Development Gene of Arabidopsis, Encodes a WRKY Transcription Factor. Plant Cell 2002, 14, 1359–1375. [Google Scholar] [CrossRef]
- Kazama, H.; Dan, H.; Imaseki, H.; Wasteneys, G.O. Transient Exposure to Ethylene Stimulates Cell Division and Alters the Fate and Polarity of Hypocotyl Epidermal Cells. Plant Physiol. 2004, 134, 1614–1623. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Total Plant | Trichomes | Glabrous | Theoretical Ratio |
|---|---|---|---|---|
| P1 (HB) | 11 | 11 | / | / |
| P2 (nwd) | 11 | / | 11 | / |
| F1 | 11 | 11 | / | / |
| F2 | 152 | 115 | 37 | 3:1 |
| BC1P1 (F1 × HB) | 106 | 106 | / | / |
| BC1P2 (F1 × nwd) | 106 | 55 | 51 | 1:1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Xu, M.; Zhou, Y.; Hu, Z.; Liu, S. CsNWD Encoding VPS62 Emerges as a Candidate Gene Conferring the Glabrous Phenotype in Cucumber. Agronomy 2024, 14, 2019. https://doi.org/10.3390/agronomy14092019
Zhou Y, Xu M, Zhou Y, Hu Z, Liu S. CsNWD Encoding VPS62 Emerges as a Candidate Gene Conferring the Glabrous Phenotype in Cucumber. Agronomy. 2024; 14(9):2019. https://doi.org/10.3390/agronomy14092019
Chicago/Turabian StyleZhou, Yuelong, Mingyuan Xu, Yong Zhou, Zhaoyang Hu, and Shiqiang Liu. 2024. "CsNWD Encoding VPS62 Emerges as a Candidate Gene Conferring the Glabrous Phenotype in Cucumber" Agronomy 14, no. 9: 2019. https://doi.org/10.3390/agronomy14092019
APA StyleZhou, Y., Xu, M., Zhou, Y., Hu, Z., & Liu, S. (2024). CsNWD Encoding VPS62 Emerges as a Candidate Gene Conferring the Glabrous Phenotype in Cucumber. Agronomy, 14(9), 2019. https://doi.org/10.3390/agronomy14092019