

Sunflower and Sunn Hemp Potential as Summer Cover Crops in Southern Texas

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Above and Belowground Plant Traits
2.3. Statistical Analysis
3. Results
3.1. Growing Conditions
3.2. Aboveground Plant Traits
3.2.1. Emergence and Mortality
3.2.2. Aboveground Biomass
3.2.3. Plant Height
3.3. SPAD
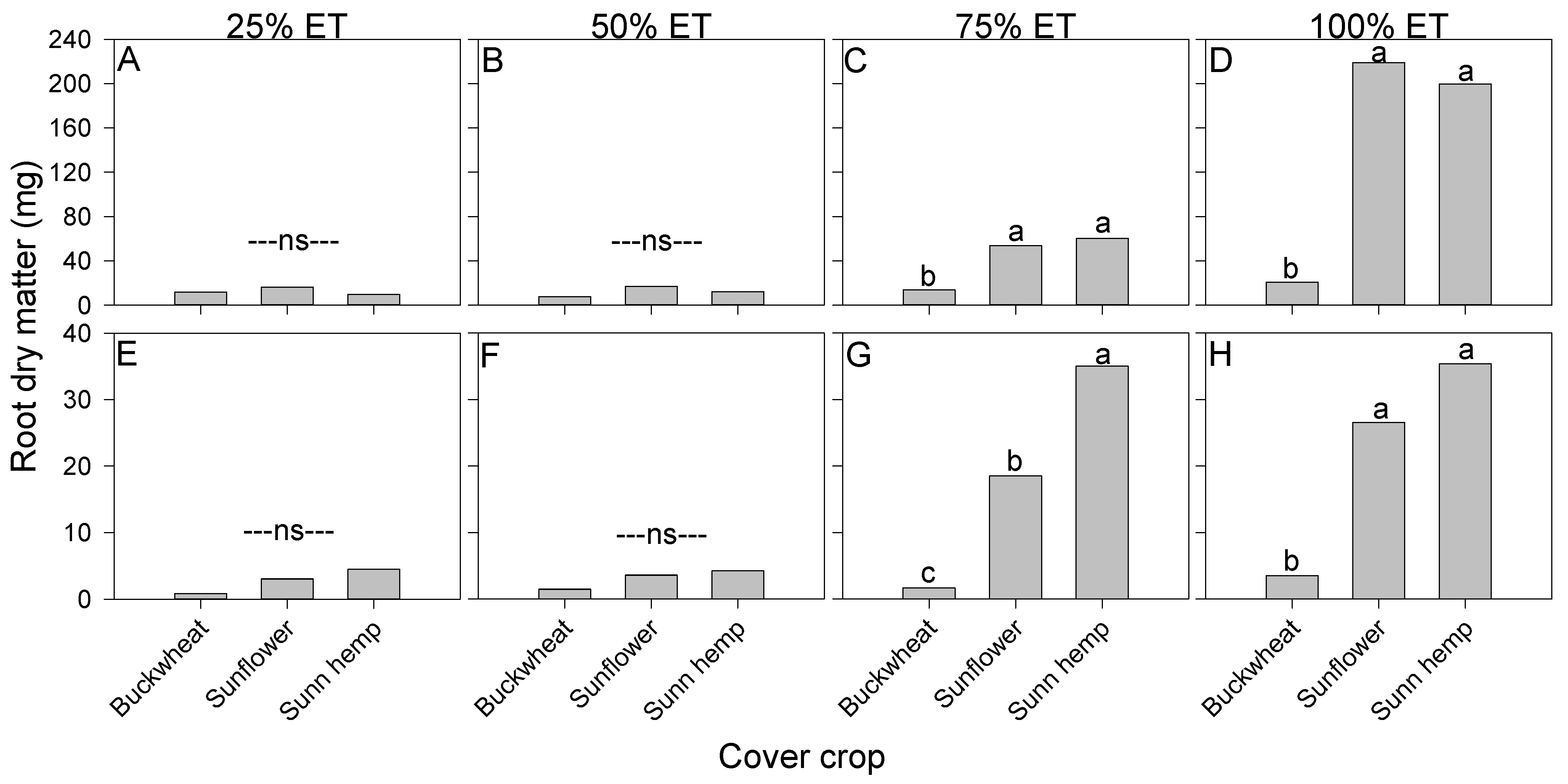
3.4. Root Traits
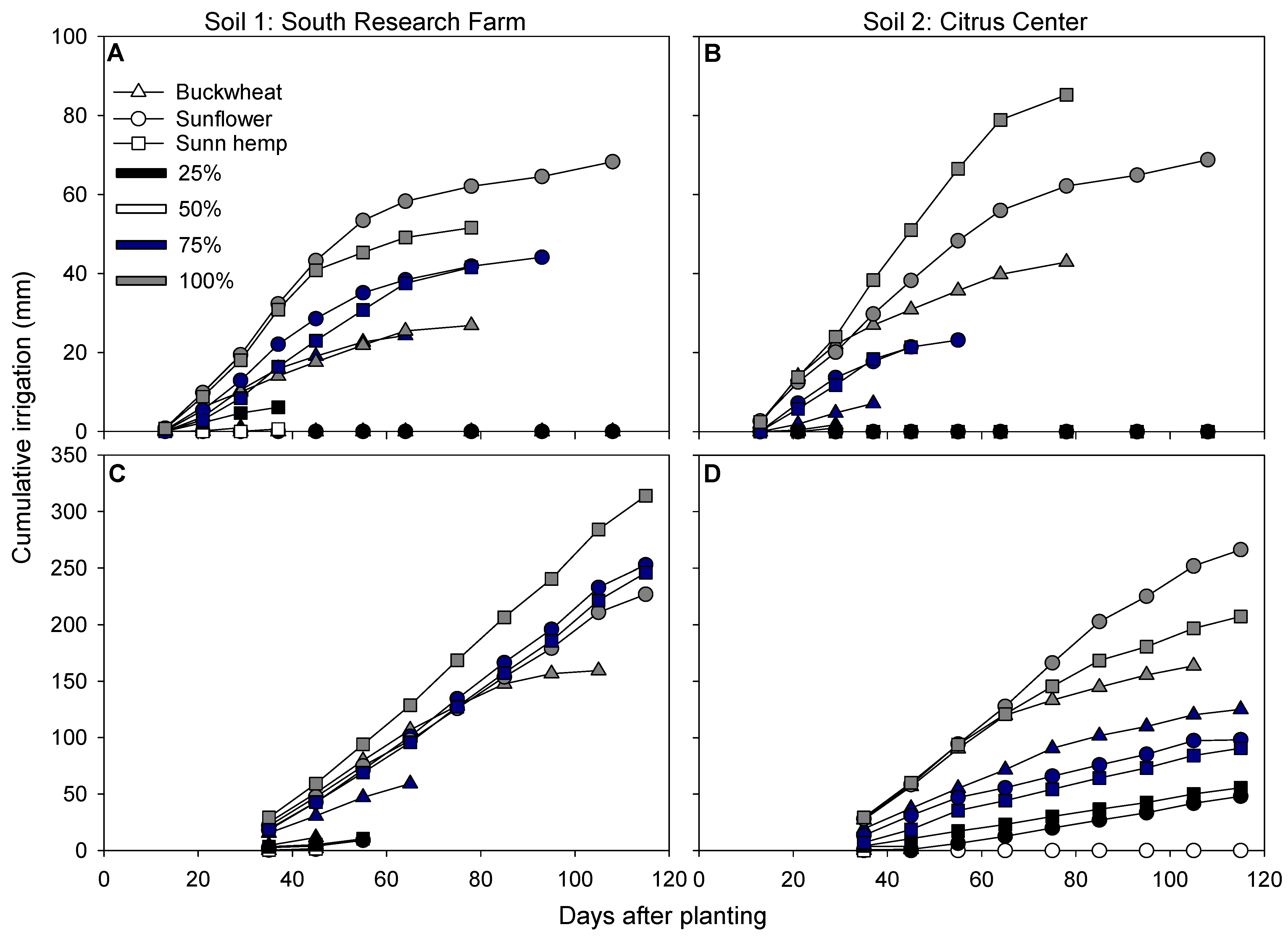
3.5. Water Dynamics
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soti, P.; Goolsby, J.A.; Racelis, A. Agricultural and environmental weeds of South Texas and their management. Subtrop. Agric. Environ. 2020, 71, 1–11. [Google Scholar]
- NASS. Crop Production 2023 Summary; U.S. Department of Agriculture, National Agricultural Statistics Service: Washington, DC, USA, 2024.
- Simpson, C.R.; Gonzalez III, J.; Enciso, J.; Nelson, S.D.; Sétamou, M. Root distribution and seasonal fluctuations under different grove floor management systems. Sci. Hortic. 2020, 272, 109364. [Google Scholar] [CrossRef]
- Simpson, C.R.; Sétamou, M.; Nelson, S.D. Evaluation of citrus grove floor management strategies for water use efficiency and conservation. Sci. Hortic. 2023, 309, 111680. [Google Scholar] [CrossRef]
- De Leon, D.A.; Nelson, S.D.; Sétamou, M.; Bhandari, A.; Zamora, E.; Zaragoza, A.; Donato, M.C.; Reyes-Cabrera, J. Enhanced nutrition programs to rehabilitate freeze-stressed citrus. HortScience 2024, 59, 1763–1771. [Google Scholar] [CrossRef]
- Allen, D.E.; Singh, B.P.; Dalal, R.C. Soil health indicators under climate change: A review of current knowledge. Soil Health Clim. Change 2011, 29, 25–45. [Google Scholar]
- Hallman, L.M.; Fox, J.-P.; Rossi, L. Use of cover crops in Florida’s citrus industry: History and resurgence, current practices, challenges, and opportunities. HortTechnology 2024, 34, 539–554. [Google Scholar] [CrossRef]
- Altieri, M.A. Agroecology: The Science of Sustainable Agriculture, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Blanco-Canqui, H.; Shaver, T.M.; Lindquist, J.L.; Shapiro, C.A.; Elmore, R.W.; Francis, C.A.; Hergert, G.W. Cover crops and ecosystem services: Insights from studies in temperate soils. Agron. J. 2015, 107, 2449–2474. [Google Scholar] [CrossRef]
- Farooq, S.; Rehman, R.U.; Pirzadah, T.B.; Malik, B.; Dar, F.A.; Tahir, I. Cultivation, agronomic practices, and growth performance of buckwheat. In Molecular Breeding and Nutritional Aspects of Buckwheat; Academic Press: Cambridge, MA, USA, 2016; pp. 299–319. [Google Scholar]
- Schelfhout, S.; De Schrijver, A.; Verheyen, K.; Beelde, R.D.; Haesaert, G.; Mertens, J. Phosphorus mining efficiency declines with decreasing soil P concentration and varies across crop species. Int. J. Phytoremediat. 2018, 20, 939–946. [Google Scholar] [CrossRef]
- Flagella, Z.; Rotunno, T.; Tarantino, E.; Di Caterina, R.; De Caro, A. Changes in seed yield and oil fatty acid composition of high oleic sunflower (Helianthus annuus L:) hybrids in relation to the sowing date and the water regime. Eur. J. Agron. 2002, 17, 221–230. [Google Scholar] [CrossRef]
- Kane, N.C.; Rieseberg, L.H. Selective sweeps reveal candidate genes for adaptation to drought and salt tolerance in common sunflower, Helianthus annuus. Genetics 2007, 175, 1823–1834. [Google Scholar] [CrossRef]
- Ashworth, A.J.; West, C.P.; Allen, F.L.; Keyser, P.D.; Weiss, S.A.; Tyler, D.D.; Taylor, A.M.; Warwick, K.L.; Beamer, K.P. Biologically fixed nitrogen in legume intercropped systems: Comparison of nitrogen-difference and nitrogen-15 enrichment techniques. Agron. J. 2015, 107, 2419–2430. [Google Scholar] [CrossRef]
- Cook, C.G.; White, G.A. Crotalaria juncea: A potential multipurpose fiber crop. In Progress in New Crops; Janick, J., Ed.; ASHS: Alexandria, VA, USA, 1996; pp. 389–394. [Google Scholar]
- Creamer, N.G.; Baldwin, K.R. An evaluation of summer cover crops for use in vegetable production systems in North Carolina. HortScience 2000, 35, 600–603. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates Incorporated: Sunderland, MA, USA, 2006. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- SAS. The SAS System for Windows. Release 9.4; SAS Institute. Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Ågren, G.I.; Franklin, O. Root:shoot ratios, optimization and nitrogen productivity. Ann. Bot. 2003, 92, 795–800. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, T.; Wei, W.; Li, Z.; Li, G.; Shen, L.; Wang, X.; Zhu, Y.; Li, L.; Zhang, W. In arid regions, the different intercropping systems reduce interspecific competition to improve system productivity. Plant Soil 2024, 506, 137–155. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Klassen, W.; Hanlon, E.A., Jr. Sunn Hemp—A Promising Cover Crop in Florida. Florida Cooperative Extension’s Electronic Data Information Source (EDIS); University of Florida: Gainesville, FL, USA, 2022. [Google Scholar]
- Schomberg, H.H.; Martini, N.L.; Diaz-Perez, J.C.; Phatak, S.C.; Balkcom, K.S.; Bhardwaj, H.L. Potential for using sunn hemp as a source of biomass and nitrogen for the Piedmont and Coastal Plain regions of the southeastern USA. Agron. J. 2007, 99, 1448–1457. [Google Scholar] [CrossRef]
- Bhardwaj, H.L.; Webber III, C.L.; Sakamoto, G.S. Cultivation of kenaf and sunn hemp in the mid-Atlantic United States. Ind. Crops Prod. 2005, 22, 151–155. [Google Scholar] [CrossRef]
- Halvorson, A.D.; Black, A.L.; Krupinsky, J.M.; Merrill, S.D.; Tanaka, D.L. Sunflower response to tillage and nitrogen fertilization under intensive cropping in a wheat rotation. Agron. J. 1999, 91, 637–642. [Google Scholar] [CrossRef]
- Martinez-Goni, X.S.; Miranda-Apodaca, J.; Perez-Lopez, U. Could buckwheat and spelt be alternatives to wheat under future environmental conditions? Study of their physiological response to drought. Agric. Water Manag. 2023, 278, 108176. [Google Scholar] [CrossRef]
- Aubert, L.; Knorádová, D.; Barris, S.; Quinet, M. Different drought resistance mechanisms between two buckwheat species Fagopyrum esculentum and Fagopyrum tataricum. Physiol. Plant 2020, 172, 577–586. [Google Scholar] [CrossRef]
- Hunt, H.V.; Shang, X.; Jones, M.K. Buckwheat: A crop from outside the major Chinese domestication centers? A review of the archaeobotanical, palynological and genetic evidence. Veg. Hist. Archaeobot. 2018, 27, 493–506. [Google Scholar] [CrossRef]
- Sherwood, S.C. Adapting to the challenges of warming. Science 2020, 370, 782–783. [Google Scholar] [CrossRef]
- Ghebreyesus, D.; Sharif, H.O. Time series analysis of monthly and annual precipitation in the state of Texas using high-resolution radar products. Water 2021, 13, 982. [Google Scholar] [CrossRef]
- Liu, L.; Hong, Y.; Hocker, J.E.; Shafer, M.A.; Carter, L.M.; Gourley, J.J.; Bednarczyk, C.N.; Yong, B.; Adhikari, P. Analyzing projected changes and trends of temperature and precipitation in the southern USA from 16 downscaled global climate models. Theor. Appl. Climatol. 2012, 109, 345–360. [Google Scholar] [CrossRef]
- Meehl, G.A.; Tebaldi, C. More intense, more frequent and longer lasting heat waves in the 21st century. Science 2004, 305, 994–997. [Google Scholar] [CrossRef]
- Witze, A. Extreme heatwaves: Surprising lessons from the record warmth. Nature 2022, 608, 464–465. [Google Scholar] [CrossRef]
- Nelson, G.C.; Valin, H.; Sands, R.D.; Havlík, P.; Ahammad, H.; Deryng, D.; Willenbockel, D. Climate change effects on agriculture: Economic responses to biophysical shocks. Proc. Natl. Acad. Sci. USA 2014, 111, 3274–3279. [Google Scholar] [CrossRef]
- Kumar, S.V.; Wigge, P.A. H2A.Z-containing nucleosomes mediate the thermosensory response in Arabidopsis. Cell 2010, 140, 136–147. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, A.; Kahlon, C.S.; Brar, A.S.; Grover, K.K.; Dia, M.; Steiner, R.L. The role of cover crops towards sustainable soil health and agriculture—A review paper. Am. J. Plant Sci. 2018, 9, 1935–1951. [Google Scholar] [CrossRef]
- Brewer, M.; Kanissery, R.G.; Strauss, S.L.; Kadyampakeni, D.M. Impact of cover cropping on temporal nutrient distribution and availability in the soil. Horticulturae 2023, 9, 1160. [Google Scholar] [CrossRef]
- Alonso-Ayuso, M.; Gabriel, J.L.; Quemada, M. The kill date as a management tool for cover cropping success. PLoS ONE 2014, 9, e109587. [Google Scholar] [CrossRef]
- Ramirez, D.A.; Yactayo, W.; Gutierrez, R.; Mares, V.; Mendiburu, F.; Posadas, A.; Quiroz, R. Chlorophyll concentration in leaves is an indicator of potato tuber yield in water-shortage conditions. Sci. Hortic. 2014, 168, 202–209. [Google Scholar] [CrossRef]
- Gavili, E.; Moosavi, A.A.; Haghighi, A.A.K. Does biochar mitigate the adverse effects of drought on the agronomic traits and yield components of soybean? Ind. Crop Prod. 2019, 128, 445–454. [Google Scholar] [CrossRef]
- Mansouri-Far, C.; Modarres Sanavy, S.A.M.; Saberali, S.F. Maize yield response to deficit irrigation during low-sensitive growth stages and nitrogen rate under semi-arid climatic conditions. Agric. Water Manag. 2010, 97, 12–22. [Google Scholar] [CrossRef]
- Pandey, R.K.; Maranville, J.W.; Chetima, M.M. Deficit irrigation and nitrogen effects on maize in a Sahelian environment: II. Shoot growth, nitrogen uptake and water extraction. Agric. Water Manag. 2000, 46, 15–27. [Google Scholar] [CrossRef]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; van Ruiven, J.; et al. High plant diversity is needed to maintain ecosystem services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef]
- Wang, Z.; Dong, B.; Stomph, T.J.; Evers, J.B.; van der Putten, P.E.L.; Ma, H.; Missale, R.; van der Werf, W. Temporal complementarity drives species combinability in strip intercropping in the Netherlands. Field Crops Res. 2023, 291, 108757. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, W.P.; Xu, H.S.; Yu, R.P.; Su, Y.; Surigaoge, S.; Wang, P.X.; Yang, X.; Lambers, H.; Li, L. Trade-offs and synergies of plant traits co-drive efficient nitrogen use in intercropping systems. Field Crops Res. 2023, 302, 109093. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | SRF a | Citrus Center b |
|---|---|---|
| pH | 7.9 | 8.0 |
| P c | 34 | 76 |
| K | 300 | 541 |
| Ca | 6200 | 5630 |
| Mg | 518 | 654 |
| S | 46 | 55 |
| B | 1.9 | 3.0 |
| Zn | 1.2 | 4.6 |
| Mn | 67 | 115 |
| Fe | 21 | 35 |
| Cu | 3.3 | 14.5 |
| Na | 247 | 265 |
| Cl | 48 | 25 |
| SOM d (%) | 1.1 | 1.7 |
| Sand (%) | 57.5 | 42.5 |
| Silt (%) | 29.8 | 46.8 |
| Clay (%) | 12.7 | 10.7 |
| Dry Matter Accumulation | Root Traits b | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Main Effect | d.f. | Emergence | Plant Height | SPAD a | Aboveground | Belowground | Length | Surface Area | Diameter | R–S c |
| Cover crop (C) | 2 | <0.0001 | 0.0015 | <0.0001 | 0.0017 | 0.0046 | <0.0001 | <0.0001 | 0.0104 | 0.0002 |
| Irrigation level (I) | 3 | 0.0699 | <0.0001 | 0.0215 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0487 | 0.0023 |
| Soil type (S) | 1 | 0.1624 | 0.0037 | 0.0030 | 0.0059 | 0.8100 | 0.1499 | 0.1874 | 0.2413 | 0.7159 |
| Repetition (R) | 1 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.5109 | 0.0488 | 0.0004 | <0.0001 |
| C × I | 6 | 0.4550 | 0.0005 | 0.4842 | 0.0819 | 0.0112 | <0.0001 | <0.0001 | 0.6517 | 0.0101 |
| C × S | 2 | 0.07003 | 0.0129 | 0.3687 | 0.0739 | 0.8435 | 0.1187 | 0.0581 | 0.1832 | 0.6679 |
| I × S | 3 | 0.0940 | 0.1470 | 0.0092 | 0.0493 | 0.2583 | 0.0601 | 0.0721 | 0.6105 | 0.6852 |
| C × I × S | 6 | 0.8771 | 0.1665 | 0.0011 | 0.3751 | 0.5787 | 0.6854 | 0.4089 | 0.0062 | 0.5715 |
| Repetition | Main Effect | Root Length (cm) | Surface Area (cm2) | Root Diameter (mm) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BW | SF | SH | BW | SF | SH | BW | SF | SH | ||
| 1 | Irrigation | |||||||||
| 25 | 20.38 a a | 42.99 b | 19.04 c | 1.34 b | 3.76 b | 1.89 b | 0.21 a | 0.28 b | 0.32 a | |
| 50 | 19.16 a | 61.32 b | 31.21 c | 1.34 b | 4.54 b | 3.20 b | 0.27 a | 0.24 b | 0.34 a | |
| 75 | 27.85 a | 45.70 b | 83.19 b | 1.91 a | 3.98 b | 6.79 b | 0.23 a | 0.34 b | 0.31 a | |
| 100 | 35.84 a | 127.58 a | 209.09 a | 2.11 a | 15.49 a | 15.26 a | 0.21 a | 0.51 a | 0.25 a | |
| Soil type | ||||||||||
| SRF | 27.61 a | 70.02 a | 78.09 b | 1.80 a | 5.77 a | 6.11 a | 0.25 a | 0.29 a | 0.28 a | |
| Citrus Center | 24.00 a | 68.78 a | 93.18 a | 1.54 a | 8.12 a | 7.47 a | 0.21 a | 0.40 a | 0.33 a | |
| 2 | Irrigation | |||||||||
| 25 | 6.49 b | 28.99 c | 29.20 c | 0.69 c | 3.06 b | 3.11 c | 0.34 a | 0.37 a | 0.35 b | |
| 50 | 12.61 a | 24.95 c | 36.19 c | 1.21 b | 3.18 b | 3.79 c | 0.33 a | 0.40 a | 0.33 b | |
| 75 | 19.86 a | 59.51 b | 159.45 b | 1.92 b | 7.46 b | 17.84 b | 0.32 a | 0.39 a | 0.40 b | |
| 100 | 24.56 a | 94.42 a | 298.28 a | 2.49 a | 12.29 a | 30.32 a | 0.36 a | 0.42 a | 0.70 a | |
| Soil type | ||||||||||
| SRF | 14.46 a | 51.36 a | 185.10 a | 1.47 a | 6.82 a | 19.85 a | 0.35 a | 0.41 a | 0.57 a | |
| Citrus Center | 17.29 a | 52.57 a | 76.46 b | 1.69 a | 6.17 a | 7.69 b | 0.35 a | 0.37 a | 0.33 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alapati, D.K.; Ancona, V.; Sétamou, M.; Donato, C.; Nelson, S.D.; Reyes-Cabrera, J. Sunflower and Sunn Hemp Potential as Summer Cover Crops in Southern Texas. Agronomy 2025, 15, 986. https://doi.org/10.3390/agronomy15040986
Alapati DK, Ancona V, Sétamou M, Donato C, Nelson SD, Reyes-Cabrera J. Sunflower and Sunn Hemp Potential as Summer Cover Crops in Southern Texas. Agronomy. 2025; 15(4):986. https://doi.org/10.3390/agronomy15040986
Chicago/Turabian StyleAlapati, Dileep Kumar, Veronica Ancona, Mamoudou Sétamou, Consuelo Donato, Shad D. Nelson, and Joel Reyes-Cabrera. 2025. "Sunflower and Sunn Hemp Potential as Summer Cover Crops in Southern Texas" Agronomy 15, no. 4: 986. https://doi.org/10.3390/agronomy15040986
APA StyleAlapati, D. K., Ancona, V., Sétamou, M., Donato, C., Nelson, S. D., & Reyes-Cabrera, J. (2025). Sunflower and Sunn Hemp Potential as Summer Cover Crops in Southern Texas. Agronomy, 15(4), 986. https://doi.org/10.3390/agronomy15040986