
S-Palmitoylation of Synaptic Proteins in Neuronal Plasticity in Normal and Pathological Brains

Abstract
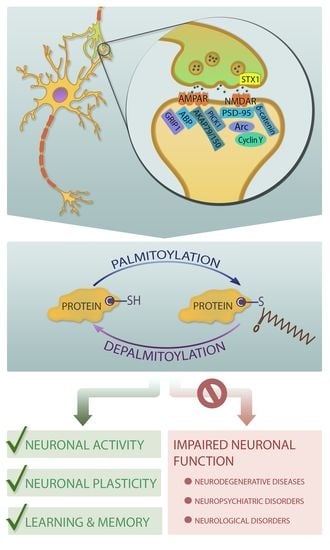
1. Introduction
1.1. Lipid Post-Translational Modifications
1.2. DHHC Family of Palmitoylation Enzymes
1.2.1. Enzymes Nomenclature and Structure
1.2.2. Localization
1.2.3. Substrates
1.3. Depalmitoylation Enzymes
1.4. Pharmacological Manipulation of Protein Palmitoylation
2. Palmitoylation—The Roles in the Function and Plasticity of Synapses
2.1. Synaptic Transmission and Synaptic Plasticity
2.2. Neuronal Activity and Synaptic Plasticity Affect Palmitoylation/Depalmitoylation Cycles
2.3. Impact of Protein Palmitoylation on Synaptic Transmission and Various Forms of Neuronal Plasticity
2.3.1. Pharmacological Modifications of Palmitoylation Machinery
2.3.2. Cysteine Mutants and Protein Engineering
2.3.3. Genetically Modified ZDHHCs and Synaptic Function
2.3.4. Depalmitoylation and Synaptic Function—Lessons from PPT1 Knockout Mice
3. Palmitoylation in Learning and Memory
4. Palmitoylation in Aging and Diseases Associated with Cognitive Decline
4.1. Palmitoylation and Aging
4.2. Neurodegenerative Diseases
4.2.1. Neuronal Ceroid Lipofuscinoses
4.2.2. Huntington’s Disease
4.2.3. Alzheimer’s Disease
4.3. Neuropsychiatric Disorders
4.3.1. Major Depressive Disorder
4.3.2. Schizophrenia
4.4. Neurological Disorders
Chromosome X-Linked Intellectual Disability
5. Conclusions and Final Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Kashina, A. Post-translational Modifications of the Protein Termini. Front. Cell Dev. Biol. 2021, 9, 719590. [Google Scholar] [CrossRef] [PubMed]
- Jensen, O.N. Modification-specific proteomics: Characterization of post-translational modifications by mass spectrometry. Curr. Opin. Chem. Biol. 2004, 8, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.D.; Overall, C.M.C. Proteolytic post-translational modification of proteins: Proteomic tools and methodology. Mol. Cell. Proteom. 2013, 12, 3532–3542. [Google Scholar] [CrossRef] [PubMed]
- Ramazi, S.; Zahiri, J. Post-translational modifications in proteins: Resources, tools and prediction methods. Database 2021, 2021, baab012. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T.; Garneau-Tsodikova, S.; Gatto, G.J. Protein posttranslational modifications: The chemistry of proteome diversifications. Angew. Chem. Int. Ed. 2005, 44, 7342–7372. [Google Scholar] [CrossRef]
- Thomas, G.M.; Hayashi, T. Smarter neuronal signaling complexes from existing components: How regulatory modifications were acquired during animal evolution: Evolution of palmitoylation-dependent regulation of AMPA-type ionotropic glutamate receptors. BioEssays 2013, 35, 929–939. [Google Scholar] [CrossRef]
- Schaffert, L.N.; Carter, W.G. Do post-translational modifications influence protein aggregation in neurodegenerative diseases: A systematic review. Brain Sciences 2020, 10, 232. [Google Scholar] [CrossRef]
- Kelley, A.R.; Bach, S.B.H.; Perry, G. Analysis of post-translational modifications in Alzheimer’s disease by mass spectrometry. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2040–2047. [Google Scholar] [CrossRef]
- Zaręba-Kozioł, M.; Figiel, I.; Bartkowiak-Kaczmarek, A.; Włodarczyk, J. Insights into protein S-palmitoylation in synaptic plasticity and neurological disorders: Potential and limitations of methods for detection and analysis. Front. Mol. Neurosci. 2018, 11, 175. [Google Scholar] [CrossRef]
- Chamberlain, L.H.; Shipston, M.J. The physiology of protein s-acylation. Physiol. Rev. 2015, 95, 341–376. [Google Scholar] [CrossRef]
- Resh, M.D. Fatty acylation of proteins: The long and the short of it. Prog. Lipid Res. 2016, 63, 120–131. [Google Scholar] [CrossRef]
- Narayana, V.K.; Tomatis, V.M.; Wang, T.; Kvaskoff, D.; Meunier, F.A. Profiling of Free Fatty Acids Using Stable Isotope Tagging Uncovers a Role for Saturated Fatty Acids in Neuroexocytosis. Chem. Biol. 2015, 22, 1552–1561. [Google Scholar] [CrossRef]
- Towler, D.; Glaser, L. Acylation of Cellular Proteins with Endogenously Synthesized Fatty Acids. Biochemistry 1986, 25, 878–884. [Google Scholar] [CrossRef]
- Muszbek, L.; Haramura, G.; Cluette-Brown, J.E.; Van Cott, E.M.; Laposata, M. The pool of fatty acids covalently bound to platelet proteins by thioester linkages can be altered by exogenously supplied fatty acids. Lipids 1999, 34, 331–337. [Google Scholar] [CrossRef]
- Liang, X.; Lu, Y.; Neubert, T.A.; Resh, M.D. Mass spectrometric analysis of GAP-43/neuromodulin reveals the presence of a variety of fatty acylated species. J. Biol. Chem. 2002, 277, 33032–33040. [Google Scholar] [CrossRef]
- Schmidt, M.F.G.; Schlesinger, M.J. Fatty acid binding to vesicular stomatitis virus glycoprotein: A new type of post-translational modification of the viral glycoprotein. Cell 1979, 17, 813–819. [Google Scholar] [CrossRef]
- Lobo, S.; Greentree, W.K.; Linder, M.E.; Deschenes, R.J. Identification of a Ras palmitoyltransferase in Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 41268–41273. [Google Scholar] [CrossRef]
- Roth, A.F.; Feng, Y.; Chen, L.; Davis, N.G. The yeast DHHC cysteine-rich domain protein Akr1p is a palmitoyl transferase. J. Cell Biol. 2002, 159, 23–28. [Google Scholar] [CrossRef]
- Mitchell, D.A.; Vasudevan, A.; Linder, M.E.; Deschenes, R.J. Protein palmitoylation by a family of DHHC protein S-acyltransferases. J. Lipid Res. 2006, 47, 1118–1127. [Google Scholar] [CrossRef]
- Matt, L.; Kim, K.; Chowdhury, D.; Hell, J.W. Role of palmitoylation of postsynaptic proteins in promoting synaptic plasticity. Front. Mol. Neurosci. 2019, 12, 8. [Google Scholar] [CrossRef]
- Main, A.; Fuller, W. Protein S-Palmitoylation: Advances and challenges in studying a therapeutically important lipid modification. FEBS J. 2022, 289, 861–882. [Google Scholar] [CrossRef] [PubMed]
- Greaves, J.; Munro, K.R.; Davidson, S.C.; Riviere, M.; Wojno, J.; Smith, T.K.; Tomkinson, N.C.O.; Chamberlain, L.H. Molecular basis of fatty acid selectivity in the zDHHC family of S-acyltransferases revealed by click chemistry. Proc. Natl. Acad. Sci. USA 2017, 114, E1365–E1374. [Google Scholar] [CrossRef] [PubMed]
- Rana, M.S.; Lee, C.J.; Banerjee, A. The molecular mechanism of DHHC protein acyltransferases. Biochem. Soc. Trans. 2018, 47, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.E.; Banerjee, A. In vitro reconstitution of substrate S-acylation by the zDHHC family of protein acyltransferases. Open Biol. 2022, 12, 210390. [Google Scholar] [CrossRef]
- Levy, A.D.; Devignot, V.; Fukata, Y.; Fukata, M.; Sobel, A.; Chauvin, S. Subcellular golgi localization of stathmin family proteins is promoted by a specific set of DHHC palmitoyl transferases. Mol. Biol. Cell 2011, 22, 1930–1942. [Google Scholar] [CrossRef]
- Brigidi, G.S.; Santyr, B.; Shimell, J.; Jovellar, B.; Bamji, S.X. Activity-regulated trafficking of the palmitoyl-acyl transferase DHHC5. Nat. Commun. 2015, 6, 8200. [Google Scholar] [CrossRef]
- Noritake, J.; Fukata, Y.; Iwanaga, T.; Hosomi, N.; Tsutsumi, R.; Matsuda, N.; Tani, H.; Iwanari, H.; Mochizuki, Y.; Kodama, T.; et al. Mobile DHHC palmitoylating enzyme mediates activity-sensitive synaptic targeting of PSD-95. J. Cell Biol. 2009, 186, 147–160. [Google Scholar] [CrossRef]
- Woolfrey, K.M.; Sanderson, J.L.; Dell’Acqua, M.L. The palmitoyl acyltransferase DHHC2 regulates recycling endosome exocytosis and synaptic potentiation through palmitoylation of AKAP79/150. J. Neurosci. 2015, 35, 442–456. [Google Scholar] [CrossRef]
- Thomas, G.M.; Hayashi, T.; Chiu, S.L.; Chen, C.M.; Huganir, R.L. Palmitoylation by DHHC5/8 Targets GRIP1 to Dendritic Endosomes to Regulate AMPA-R Trafficking. Neuron 2012, 73, 482–496. [Google Scholar] [CrossRef]
- Brigidi, G.S.; Sun, Y.; Beccano-Kelly, D.; Pitman, K.; Mobasser, M.; Borgland, S.L.; Milnerwood, A.J.; Bamji, S.X. Palmitoylation of δ-catenin by DHHC5 mediates activity-induced synapse plasticity. Nat. Neurosci. 2014, 17, 522–532. [Google Scholar] [CrossRef]
- Fukata, Y.; Dimitrov, A.; Boncompain, G.; Vielemeyer, O.; Perez, F.; Fukata, M. Local palmitoylation cycles define activity-regulated postsynaptic subdomains. J. Cell Biol. 2013, 202, 145–161. [Google Scholar] [CrossRef]
- Woodley, K.T.; Collins, M.O. Regulation and function of the palmitoyl-acyltransferase ZDHHC5. FEBS J. 2021, 288, 6623–6634. [Google Scholar] [CrossRef]
- Mukai, J.; Liu, H.; Burt, R.; Swor, D.E.; Lai, W.-S.; Karayiorgou, M.; Gogos, J.A. Evidence that the gene encoding ZDHHC8 contributes to the risk of schizophrenia. Nat. Genet. 2004, 36, 725–731. [Google Scholar] [CrossRef]
- Saitoh, F.; Tian, Q.B.; Okano, A.; Sakagami, H.; Kondo, H.; Suzuki, T. NIDD, a Novel DHHC-containing Protein, Targets Neuronal Nitric-oxide Synthase (nNOS) to the Synaptic Membrane through a PDZ-dependent Interaction and Regulates nNOS Activity. J. Biol. Chem. 2004, 279, 29461–29468. [Google Scholar] [CrossRef]
- Shah, B.S.; Shimell, J.J.; Bamji, S.X. Regulation of dendrite morphology and excitatory synapse formation by zDHHC15. J. Cell Sci. 2019, 132, jcs230052. [Google Scholar] [CrossRef]
- Shimell, J.J.; Shah, B.S.; Cain, S.M.; Thouta, S.; Kuhlmann, N.; Tatarnikov, I.; Jovellar, D.B.; Brigidi, S.; Kass, J.; Milnerwood, A.J. The X-Linked Intellectual Disability Gene Zdhhc9 Is Essential for Dendrite Outgrowth and Inhibitory Synapse Formation. Cell Rep. 2019, 29, 2422–2437.e8. [Google Scholar] [CrossRef]
- Dejanovic, B.; Semtner, M.; Ebert, S.; Lamkemeyer, T.; Neuser, F.; Luscher, B.; Meier, J.; Schwarz, G. Palmitoylation of Gephyrin Controls Receptor Clustering and Plasticity of GABAergic Synapses. PLoS Biol. 2014, 12, e1001908. [Google Scholar] [CrossRef]
- Ohyama, T.; Verstreken, P.; Ly, C.V.; Rosenmund, T.; Rajan, A.; Tien, A.-C.; Haueter, C.; Schulze, K.L.; Bellen, H.J. Huntingtin-interacting protein 14, a palmitoyl transferase required for exocytosis and targeting of CSP to synaptic vesicles. J. Cell Biol. 2007, 179, 1481–1496. [Google Scholar] [CrossRef]
- Stowers, R.S.; Isacoff, E. Drosophila Huntingtin-Interacting Protein 14 Is a Presynaptic Protein Required for Photoreceptor Synaptic Transmission and Expression of the Palmitoylated Proteins Synaptosome-Associated Protein 25 and Cysteine String Protein. J. Neurosci. 2007, 27, 12874–12883. [Google Scholar] [CrossRef]
- Sanders, S.S.; Hernandez, L.M.; Soh, H.; Karnam, S.; Walikonis, R.S.; Tzingounis, A.V.; Thomas, G.M. The palmitoyl acyltransferase ZDHHC14 controls Kv1-family potassium channel clustering at the axon initial segment. eLife 2020, 9, e56058. [Google Scholar] [CrossRef]
- Nasseri, G.G.; Matin, N.; Wild, A.R.; Tosefsky, K.; Flibotte, S.; Stacey, R.G.; Hollman, R.B.; Foster, L.J.; Bamji, S.X. Synaptic activity–dependent changes in the hippocampal palmitoylome. Sci. Signal. 2022, 15, eadd2519. [Google Scholar] [CrossRef] [PubMed]
- Greaves, J.; Gorleku, O.A.; Salaun, C.; Chamberlain, L.H. Palmitoylation of the SNAP25 Protein Family: Specificity and Regulation by DHHC Palmitoyl Transferases. J. Biol. Chem. 2010, 285, 24629–24638. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Wan, J.; Arstikaitis, P.; Takahashi, H.; Huang, K.; Bailey, A.O.; Thompson, J.X.; Roth, A.F.; Drisdel, R.C.; Mastro, R.; et al. Neural palmitoyl-proteomics reveals dynamic synaptic palmitoylation. Nature 2008, 456, 904–909. [Google Scholar] [CrossRef] [PubMed]
- Jennings, B.C.; Linder, M.E. DHHC protein S-acyltransferases use similar ping-pong kinetic mechanisms but display different Acyl-CoA specificities. J. Biol. Chem. 2012, 287, 7236–7245. [Google Scholar] [CrossRef]
- Fukata, Y.; Iwanaga, T.; Fukata, M. Systematic screening for palmitoyl transferase activity of the DHHC protein family in mammalian cells. Methods 2006, 40, 177–182. [Google Scholar] [CrossRef]
- Fukata, M.; Fukata, Y.; Adesnik, H.; Nicoll, R.A.; Bredt, D.S. Identification of PSD-95 Palmitoylating Enzymes. Neuron 2004, 44, 987–996. [Google Scholar] [CrossRef]
- Fukata, Y.; Fukata, M. Protein palmitoylation in neuronal development and synaptic plasticity. Nat. Rev. Neurosci. 2010, 11, 161–175. [Google Scholar] [CrossRef]
- Jiang, J.; Suppiramaniam, V.; Wooten, M.W. Posttranslational Modifications and Receptor-Associated Proteins in AMPA Receptor Trafficking and Synaptic Plasticity. Neurosignals 2006, 15, 266–282. [Google Scholar] [CrossRef]
- Koster, K.P. AMPAR Palmitoylation Tunes Synaptic Strength: Implications for Synaptic Plasticity and Disease. J. Neurosci. 2019, 39, 5040–5043. [Google Scholar] [CrossRef]
- Lu, W.; Roche, K.W. Posttranslational regulation of AMPA receptor trafficking and function. Curr. Opin. Neurobiol. 2012, 22, 470–479. [Google Scholar] [CrossRef]
- Sohn, H.; Park, M. Palmitoylation-mediated synaptic regulation of AMPA receptor trafficking and function. Arch. Pharmacal Res. 2019, 42, 426–435. [Google Scholar] [CrossRef]
- Gladding, C.M.; Raymond, L.A. Mechanisms underlying NMDA receptor synaptic/extrasynaptic distribution and function. Mol. Cell. Neurosci. 2011, 48, 308–320. [Google Scholar] [CrossRef]
- Lussier, M.; Sanz-Clemente, A.; Roche, K.W. Dynamic Regulation of N-Methyl-d-aspartate (NMDA) and α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic Acid (AMPA) Receptors by Posttranslational Modifications. J. Biol. Chem. 2015, 290, 28596–28603. [Google Scholar] [CrossRef]
- Qiu, S.; Li, X.-Y.; Zhuo, M. Post-translational modification of NMDA receptor GluN2B subunit and its roles in chronic pain and memory. Semin. Cell Dev. Biol. 2011, 22, 521–529. [Google Scholar] [CrossRef]
- Warnet, X.L.; Bakke Krog, H.; Sevillano-Quispe, O.G.; Poulsen, H.; Kjaergaard, M. The C-terminal domains of the NMDA receptor: How intrinsically disordered tails affect signalling, plasticity and disease. Eur. J. Neurosci. 2021, 54, 6713–6739. [Google Scholar] [CrossRef]
- Shipston, M.J. Europe PMC Funders Group Regulation of large conductance calcium- and voltage-activated potassium (BK) channels by S-palmitoylation. Biochem. Soc. Trans. 2014, 41, 67–71. [Google Scholar] [CrossRef]
- Rastedt, D.E.; Vaughan, R.A.; Foster, J.D. Palmitoylation mechanisms in dopamine transporter regulation. J. Chem. Neuroanat. 2017, 83–84, 3–9. [Google Scholar] [CrossRef]
- Prescott, G.R.; Gorleku, O.A.; Greaves, J.; Chamberlain, L.H. Palmitoylation of the synaptic vesicle fusion machinery. J. Neurochem. 2009, 110, 1135–1149. [Google Scholar] [CrossRef]
- Cassinelli, S.; Viñola-Renart, C.; Benavente-Garcia, A.; Navarro-Pérez, M.; Capera, J.; Felipe, A. Palmitoylation of Voltage-Gated Ion Channels. Int. J. Mol. Sci. 2022, 23, 9357. [Google Scholar] [CrossRef]
- Montersino, A.; Thomas, G.M. Slippery signaling: Palmitoylation-dependent control of neuronal kinase localization and activity. Mol. Membr. Biol. 2015, 32, 179–188. [Google Scholar] [CrossRef]
- Patwardhan, A.; Cheng, N.; Trejo, J. Post-Translational Modifications of G Protein–Coupled Receptors Control Cellular Signaling Dynamics in Space and Time. Pharmacol. Rev. 2021, 73, 120–151. [Google Scholar] [CrossRef]
- Martin, B.R.; Wang, C.; Adibekian, A.; Tully, S.E.; Cravatt, B.F. Global profiling of dynamic protein palmitoylation. Nat. Methods 2012, 9, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Sanders, S.S.; Martin, D.D.O.; Butland, S.L.; Lavallée-Adam, M.; Calzolari, D.; Kay, C.; Yates, J.R.; Hayden, M.R. Curation of the Mammalian Palmitoylome Indicates a Pivotal Role for Palmitoylation in Diseases and Disorders of the Nervous System and Cancers. PLoS Comput. Biol. 2015, 11, e1004405. [Google Scholar] [CrossRef] [PubMed]
- Zareba-Koziol, M.; Bartkowiak-Kaczmarek, A.; Figiel, I.; Krzystyniak, A.; Wojtowicz, T.; Bijata, M.; Wlodarczyk, J. Stress-induced Changes in the S-palmitoylation and S-nitrosylation of Synaptic Proteins. Mol. Cell. Proteom. 2019, 18, 1916–1938. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.S.; Halder, A.K.; Zaręba-Kozioł, M.; Bartkowiak-Kaczmarek, A.; Dutta, A.; Chatterjee, P.; Nasipuri, M.; Wójtowicz, T.; Wlodarczyk, J.; Basu, S. RFCM-PALM: In-Silico Prediction of S-Palmitoylation Sites in the Synaptic Proteins for Male/Female Mouse Data. Int. J. Mol. Sci. 2021, 22, 9901. [Google Scholar] [CrossRef]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. CSS-Palm 2.0: An updated software for palmitoylation sites prediction. Protein Eng. Des. Sel. 2008, 21, 639–644. [Google Scholar] [CrossRef]
- Xue, Y.; Chen, H.; Jin, C.; Sun, Z.; Yao, X. NBA-Palm: Prediction of palmitoylation site implemented in Naïve Bayes algorithm. BMC Bioinform. 2006, 7, 458. [Google Scholar] [CrossRef]
- Wild, A.R.; Hogg, P.W.; Flibotte, S.; Nasseri, G.G.; Hollman, R.B.; Abazari, D.; Haas, K.; Bamji, S.X. Exploring the expression patterns of palmitoylating and de-palmitoylating enzymes in the mouse brain using the curated RNA-seq database BrainPalmSeq. eLife 2022, 11, e75804. [Google Scholar] [CrossRef]
- Camp, L.A.; Hofmann, S.L. Purification and properties of a palmitoyl-protein thioesterase that cleaves palmitate from H-Ras. J. Biol. Chem. 1993, 268, 22566–22574. [Google Scholar] [CrossRef]
- Gorenberg, E.L.; Tieze, S.M.; Yücel, B.; Zhao, H.R.; Chou, V.; Wirak, G.S.; Tomita, S.; Lam, T.T.; Chandra, S.S. Identification of substrates of palmitoyl protein thioesterase 1 highlights roles of depalmitoylation in disulfide bond formation and synaptic function. PLoS Biol. 2022, 20, e3001590. [Google Scholar] [CrossRef]
- Lehtovirta, M.; Kyttala, A.; Eskelinen, E.L.; Hess, M.; Heinonen, O.; Jalanko, A. Palmitoyl protein thioesterase (PPT) localizes into synaptosomes and synaptic vesicles in neurons: Implications for infantile neuronal ceroid lipofuscionosis (INCL). Hum. Mol. Genet. 2001, 10, 69–75. [Google Scholar] [CrossRef]
- Soyombo, A.A.; Hofmann, S.L. Molecular Cloning and Expression of Palmitoyl-protein Thioesterase 2 (PPT2), a Homolog of Lysosomal Palmitoyl-protein Thioesterase with a Distinct Substrate Specificity. J. Biol. Chem. 1997, 272, 27456–27463. [Google Scholar] [CrossRef]
- Duncan, J.A.; Gilman, A.G. A Cytoplasmic Acyl-Protein Thioesterase That Removes Palmitate from G Protein α Subunits and p21RAS. J. Biol. Chem. 1998, 273, 15830–15837. [Google Scholar] [CrossRef]
- Won, S.J.; Cheung See Kit, M.C.S.; Martin, B.R. Protein depalmitoylases. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 83–98. [Google Scholar] [CrossRef]
- De, I.; Sadhukhan, S. Emerging Roles of DHHC-mediated Protein S-palmitoylation in Physiological and Pathophysiological Context. Eur. J. Cell Biol. 2018, 97, 319–338. [Google Scholar] [CrossRef]
- Lin, D.T.; Conibear, E. Enzymatic protein depalmitoylation by acyl protein thioesterases. Biochem. Soc. Trans. 2015, 43, 193–198. [Google Scholar] [CrossRef]
- Lin, D.T.S.; Conibear, E. Author response: ABHD17 proteins are novel protein depalmitoylases that regulate N-Ras palmitate turnover and subcellular localization. eLife 2015, 4, e11306. [Google Scholar] [CrossRef]
- Cao, Y.; Qiu, T.; Kathayat, R.S.; Azizi, S.-A.; Thorne, A.; Ahn, D.; Fukata, Y.; Fukata, M.; Rice, P.A.; Dickinson, B.C. ABHD10 is an S-depalmitoylase affecting redox homeostasis through peroxiredoxin-5. Nat. Chem. Biol. 2019, 15, 1232–1240. [Google Scholar] [CrossRef]
- Long, J.Z.; Cravatt, B.F. The Metabolic Serine Hydrolases and Their Functions in Mammalian Physiology and Disease. Chem. Rev. 2011, 111, 6022–6063. [Google Scholar] [CrossRef]
- Hu, L.; Tao, Z.; Wu, X. Insights into auto-: S -fatty acylation: Targets, druggability, and inhibitors. RSC Chem. Biol. 2021, 2, 1667–1679. [Google Scholar] [CrossRef]
- Hu, X.; Zhu, X.; Yu, W.; Zhang, Y.; Yang, K.; Liu, Z.; Qiao, X.; Song, Y. A mini review of small-molecule inhibitors targeting palmitoyltransferases. Eur. J. Med. Chem. Rep. 2022, 5, 100041. [Google Scholar] [CrossRef]
- Webb, Y.; Hermida-Matsumoto, L.; Resh, M.D. Inhibition of Protein Palmitoylation, Raft Localization, and T Cell Signaling by 2-Bromopalmitate and Polyunsaturated Fatty Acids. Available online: http://www.jbc.org/ (accessed on 21 December 2020).
- Davda, D.; El Azzouny, M.A.; Tom, C.T.; Hernandez, J.L.; Majmudar, J.D.; Kennedy, R.T.; Martin, B.R. Profiling Targets of the Irreversible Palmitoylation Inhibitor 2-Bromopalmitate. ACS Chem. Biol. 2013, 8, 1912–1917. [Google Scholar] [CrossRef] [PubMed]
- Pedro, M.D.P.; Vilcaes, A.A.; Tomatis, V.; Oliveira, R.G.; Gomez, G.; Daniotti, J.L. 2-Bromopalmitate Reduces Protein Deacylation by Inhibition of Acyl-Protein Thioesterase Enzymatic Activities. PLoS ONE 2013, 8, e75232. [Google Scholar] [CrossRef] [PubMed]
- Vartak, N.; Papke, B.; Grecco, H.E.; Rossmannek, L.; Waldmann, H.; Hedberg, C.; Bastiaens, P.I. The Autodepalmitoylating Activity of APT Maintains the Spatial Organization of Palmitoylated Membrane Proteins. Biophys. J. 2014, 106, 93–105. [Google Scholar] [CrossRef] [PubMed]
- DeJesus, G.; Bizzozero, O.A. Effect of 2-Fluoropalmitate, Cerulenin and Tunicamycin on the Palmitoylation and Intracellular Translocation of Myelin Proteolipid Protein. Neurochem. Res. 2002, 27, 1669–1675. [Google Scholar] [CrossRef]
- Azizi, S.-A.; Lan, T.; Delalande, C.; Kathayat, R.S.; Mejia, F.B.; Qin, A.; Brookes, N.; Sandoval, P.J.; Dickinson, B.C. Development of an Acrylamide-Based Inhibitor of Protein S-Acylation. ACS Chem. Biol. 2021, 16, 1546–1556. [Google Scholar] [CrossRef]
- Ducker, C.E.; Griffel, L.K.; Smith, R.A.; Keller, S.N.; Zhuang, Y.; Xia, Z.; Diller, J.D.; Smith, C.D. Discovery and characterization of inhibitors of human palmitoyl acyltransferases. Mol. Cancer Ther. 2006, 5, 1647–1659. [Google Scholar] [CrossRef]
- Jennings, B.C.; Nadolski, M.J.; Ling, Y.; Baker, M.B.; Harrison, M.L.; Deschenes, R.J.; Linder, M.E. 2-Bromopalmitate and 2-(2-hydroxy-5-nitro-benzylidene)-benzo[b]thiophen-3-one inhibit DHHC-mediated palmitoylation in vitro. J. Lipid Res. 2009, 50, 233–242. [Google Scholar] [CrossRef]
- Coleman, D.T.; Soung, Y.H.; Surh, Y.-J.; Cardelli, J.A.; Chung, J. Curcumin Prevents Palmitoylation of Integrin β4 in Breast Cancer Cells. PLoS ONE 2015, 10, e0125399. [Google Scholar] [CrossRef]
- Qiu, N.; Abegg, D.; Guidi, M.; Gilmore, K.; Seeberger, P.H.; Adibekian, A. Artemisinin inhibits NRas palmitoylation by targeting the protein acyltransferase ZDHHC6. Cell Chem. Biol. 2021, 29, 530–537.e7. [Google Scholar] [CrossRef]
- Dekker, F.J.; Rocks, O.; Vartak, N.; Menninger, S.; Hedberg, C.; Balamurugan, R.; Wetzel, S.; Renner, S.; Gerauer, M.; Schölermann, B.; et al. Small-molecule inhibition of APT1 affects Ras localization and signaling. Nat. Chem. Biol. 2010, 6, 449–456. [Google Scholar] [CrossRef]
- Hedberg, C.; Dekker, F.J.; Rusch, D.M.; Renner, S.; Wetzel, S.; Vartak, N.; Gerding-Reimers, C.; Bon, R.S.; Bastiaens, P.I.H.; Waldmann, H. Development of Highly Potent Inhibitors of the Ras-Targeting Human Acyl Protein Thioesterases Based on Substrate Similarity Design. Angew. Chem. Int. Ed. 2011, 50, 9832–9837. [Google Scholar] [CrossRef]
- Kemp, L.E.; Rusch, M.; Adibekian, A.; Bullen, H.E.; Graindorge, A.; Freymond, C.; Rottmann, M.; Braun-Breton, C.; Baumeister, S.; Porfetye, A.T.; et al. Characterization of a Serine Hydrolase Targeted by Acyl-protein Thioesterase Inhibitors in Toxoplasma gondii. J. Biol. Chem. 2013, 288, 27002–27018. [Google Scholar] [CrossRef]
- Adibekian, A.; Martin, B.R.; Speers, A.E.; Brown, S.J.; Spicer, T.; Fernandes-Vega, V.; Ferguson, J.; Cravatt, B.F.; Hodder, P.; Rosen, H. Optimization and characterization of a triazole urea dual inhibitor for lysophospholipase 1 (LYPLA1) and lysophospholipase 2 (LYPLA2). Probe Rep. NIH Mol. Libr. Progr. 2010, 1, 1–42. [Google Scholar]
- Sarkar, C.; Chandra, G.; Peng, S.; Zhang, Z.; Liu, A.; Murkherjee, A.B. Neuroprotection and lifespan extension in Ppt1-/- mice by NtBuHA: Therapeutic implications for INCL. Nat. Neurosci. 2013, 16, 1608–1617. [Google Scholar] [CrossRef]
- Ji, B.; Skup, M. Roles of palmitoylation in structural long-term synaptic plasticity. Mol. Brain 2021, 14, 8. [Google Scholar] [CrossRef]
- Albanesi, J.P.; Barylko, B.; DeMartino, G.N.; Jameson, D.M. Palmitoylated Proteins in Dendritic Spine Remodeling. Front. Synaptic Neurosci. 2020, 12, 22. [Google Scholar] [CrossRef]
- Globa, A.K.; Bamji, S.X. Protein palmitoylation in the development and plasticity of neuronal connections. Curr. Opin. Neurobiol. 2017, 45, 210–220. [Google Scholar] [CrossRef]
- Petroff, O.A.C. GABA and Glutamate in the Human Brain. Neuroscientist 2002, 8, 562–573. [Google Scholar] [CrossRef]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate Receptor Ion Channels: Structure, Regulation, and Function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef]
- Ghit, A.; Assal, D.; Al-shami, A.S.; Hussein, D.E.E. GABA A receptors: Structure, function, pharmacology, and related disorders. J. Genet. Eng. Biotechnol. 2021, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Bean, B.P. The action potential in mammalian central neurons. Nat. Rev. Neurosci. 2007, 8, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Malenka, R.C.; Bear, M.F. LTP and LTD: An embarrassment of riches. Neuron 2004, 44, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Citri, A.; Malenka, R.C. Synaptic Plasticity: Multiple Forms, Functions, and Mechanisms. Neuropsychopharmacology 2008, 33, 18–41. [Google Scholar] [CrossRef]
- Regehr, W.G. Short-Term Presynaptic Plasticity. Cold Spring Harb. Perspect. Biol. 2012, 4, a005702. [Google Scholar] [CrossRef]
- Kullmann, D.M.; Lamsa, K.P. Long-term synaptic plasticity in hippocampal interneurons. Nat. Rev. Neurosci. 2007, 8, 687–699. [Google Scholar] [CrossRef]
- Nicoll, R.A.; Schmitz, D. Synaptic plasticity at hippocampal mossy fibre synapses. Nat. Rev. Neurosci. 2005, 6, 863–876. [Google Scholar] [CrossRef]
- Bliss, T.V.P.; Lømo, T. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. 1973, 232, 331–356. [Google Scholar] [CrossRef]
- Bliss, T.V.P.; Collingridge, G.L.; Morris, R.G.M.; Reymann, K.G. Long-term potentiation in the hippocampus: Discovery, mechanisms and function. Neuroforum 2018, 24, A103–A120. [Google Scholar] [CrossRef]
- Lisman, J.; Buzsáki, G.; Eichenbaum, H.; Nadel, L.; Ranganath, C.; Redish, A.D. Viewpoints: How the hippocampus contributes to memory, navigation and cognition. Nat. Neurosci. 2017, 20, 1434–1447. [Google Scholar] [CrossRef]
- Voss, J.L.; Bridge, D.J.; Cohen, N.J.; Walker, J.A. A Closer Look at the Hippocampus and Memory. Trends Cogn. Sci. 2017, 21, 577–588. [Google Scholar] [CrossRef]
- Jarrard, L.E. On the role of the hippocampus in learning and memory in the rat. Behav. Neural Biol. 1993, 60, 9–26. [Google Scholar] [CrossRef]
- Anand, K.S.; Dhikav, V. Hippocampus in health and disease: An overview. Ann. Indian Acad. Neurol. 2012, 15, 239–246. [Google Scholar]
- Magee, J.C.; Grienberger, C. Synaptic Plasticity Forms and Functions. Annu. Rev. Neurosci. 2020, 43, 95–117. [Google Scholar] [CrossRef]
- Malenka, R.C. Synaptic plasticity in the hippocampus: LTP and LTD. Cell 1994, 78, 535–538. [Google Scholar] [CrossRef]
- Adams, J.P.; Dudek, S.M. Late-phase long-term potentiation: Getting to the nucleus. Nat. Rev. Neurosci. 2005, 6, 737–743. [Google Scholar] [CrossRef]
- Bin Ibrahim, M.Z.; Benoy, A.; Sajikumar, S. Long-term plasticity in the hippocampus: Maintaining within and ‘tagging’ between synapses. FEBS J. 2022, 289, 2176–2201. [Google Scholar] [CrossRef]
- Turrigiano, G.G.; Nelson, S.B. Homeostatic plasticity in the developing nervous system. Nat. Rev. Neurosci. 2004, 5, 97–107. [Google Scholar] [CrossRef]
- Tien, N.-W.; Kerschensteiner, D. Homeostatic plasticity in neural development. Neural Dev. 2018, 13, 9. [Google Scholar] [CrossRef]
- Barberis, A. Postsynaptic plasticity of GABAergic synapses. Neuropharmacology 2020, 169, 107643. [Google Scholar] [CrossRef]
- Castillo, P.E.; Chiu, C.Q.; Carroll, R.C. Long-term plasticity at inhibitory synapses. Curr. Opin. Neurobiol. 2011, 21, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Petrini, E.M.; Barberis, A. Diffusion dynamics of synaptic molecules during inhibitory postsynaptic plasticity. Front. Cell. Neurosci. 2014, 8, 300. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Q.; Barberis, A.; Higley, M.J. Preserving the balance: Diverse forms of long-term GABAergic synaptic plasticity. Nat. Rev. Neurosci. 2019, 20, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Capogna, M.; Castillo, P.E.; Maffei, A. The ins and outs of inhibitory synaptic plasticity: Neuron types, molecular mechanisms and functional roles. Eur. J. Neurosci. 2021, 54, 6882–6901. [Google Scholar] [CrossRef] [PubMed]
- Collingridge, G.L.; Peineau, S.; Howland, J.G.; Wang, Y.T. Long-term depression in the CNS. Nat. Rev. Neurosci. 2010, 11, 459–473. [Google Scholar] [CrossRef]
- Dringenberg, H.C. The history of long-term potentiation as a memory mechanism: Controversies, confirmation, and some lessons to remember. Hippocampus 2020, 30, 987–1012. [Google Scholar] [CrossRef]
- Nicoll, R.A. A Brief History of Long-Term Potentiation. Neuron 2017, 93, 281–290. [Google Scholar] [CrossRef]
- Sacchetti, B.; Lorenzini, C.A.; Baldi, E.; Bucherelli, C.; Roberto, M.; Tassoni, G.; Brunelli, M. Long-lasting hippocampal potentiation and contextual memory consolidation. Eur. J. Neurosci. 2001, 13, 2291–2298. [Google Scholar] [CrossRef]
- Whitlock, J.R.; Heynen, A.J.; Shuler, M.G.; Bear, M.F. Learning Induces Long-Term Potentiation in the Hippocampus. Science 2006, 313, 1093–1097. [Google Scholar] [CrossRef]
- Shimshek, D.R.; Jensen, V.; Celikel, T.; Geng, Y.; Schupp, B.; Bus, T.; Mack, V.; Marx, V.; Hvalby, Ø.; Seeburg, P.H.; et al. Forebrain-Specific Glutamate Receptor B Deletion Impairs Spatial Memory But Not Hippocampal Field Long-Term Potentiation. J. Neurosci. 2006, 26, 8428–8440. [Google Scholar] [CrossRef]
- Zamanillo, D.; Sprengel, R.; Hvalby, B.; Sakmann, B. Importance of AMPA Receptors for Hippocampal Synaptic Learning. Science 1999, 284, 1805–1811. [Google Scholar] [CrossRef]
- Häusser, M.; Mel, B. Dendrites: Bug or feature? Curr. Opin. Neurobiol. 2003, 13, 372–383. [Google Scholar] [CrossRef]
- Poirazi, P.; Mel, B.W. Impact of Active Dendrites and Structural Plasticity on the Memory Capacity of Neural Tissue. Neuron 2001, 29, 779–796. [Google Scholar] [CrossRef]
- Polsky, A.; Mel, B.W.; Schiller, J. Computational subunits in thin dendrites of pyramidal cells. Nat. Neurosci. 2004, 7, 621–627. [Google Scholar] [CrossRef]
- Zhang, W.; Linden, D.J. The other side of the engram: Experience-driven changes in neuronal intrinsic excitability. Nat. Rev. Neurosci. 2003, 4, 885–900. [Google Scholar] [CrossRef]
- Rocks, O.; Gerauer, M.; Vartak, N.; Koch, S.; Huang, Z.-P.; Pechlivanis, M.; Kuhlmann, J.; Brunsveld, L.; Chandra, A.; Ellinger, B.; et al. The Palmitoylation Machinery Is a Spatially Organizing System for Peripheral Membrane Proteins. Cell 2010, 141, 458–471. [Google Scholar] [CrossRef]
- Magee, A.I.; Gutierrez, L.; McKay, I.A.; Marshall, C.J.; Hall, A. Dynamic fatty acylation of p21N-ras. EMBO J. 1987, 6, 3353–3357. [Google Scholar] [CrossRef]
- Staufenbiel, M. Ankyrin-bound fatty acid turns over rapidly at the erythrocyte plasma membrane. Mol. Cell. Biol. 1987, 7, 2981–2984. [Google Scholar]
- Rocks, O.; Peyker, A.; Kahms, M.; Verveer, P.J.; Koerner, C.; Lumbierres, M.; Kuhlmann, J.; Waldmann, H.; Wittinghofer, A.; Bastiaens, P.I.H. An Acylation Cycle Regulates Localization and Activity of Palmitoylated Ras Isoforms. Science 2005, 307, 1746–1752. [Google Scholar] [CrossRef]
- Yokoi, N.; Fukata, Y.; Sekiya, A.; Murakami, T.; Kobayashi, K.; Fukata, M. Identification of PSD-95 Depalmitoylating Enzymes. J. Neurosci. 2016, 36, 6431–6444. [Google Scholar] [CrossRef]
- Zhang, Y.; Matt, L.; Patriarchi, T.; Malik, Z.A.; Chowdhury, D.; Park, D.K.; Renieri, A.; Ames, J.B.; Hell, J.W. Capping of the N-terminus of PSD-95 by calmodulin triggers its postsynaptic release. EMBO J. 2014, 33, 1341–1353. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.-Y.; Man, H.-Y.; Ju, W.; Trimble, W.S.; MacDonald, J.F.; Wang, Y.T. Activation of Synaptic NMDA Receptors Induces Membrane Insertion of New AMPA Receptors and LTP in Cultured Hippocampal Neurons. Neuron 2001, 29, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Musleh, W.; Bi, X.; Tocco, G.; Yaghoubi, S.; Baudry, M. Glycine-induced long-term potentiation is associated with structural and functional modifications of α-amino-3-hydroxyl-5-methyl-4-isoxazolepropionic acid receptors. Proc. Natl. Acad. Sci. USA 1997, 94, 9451–9456. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Specht, C.G.; Waites, C.L.; Butler-Munro, C.; Leal-Ortiz, S.; Foote, J.W.; Genoux, D.; Garner, C.; Montgomery, J.M. SAP97 directs NMDA receptor spine targeting and synaptic plasticity. J. Physiol. 2011, 589, 4491–4510. [Google Scholar] [CrossRef]
- Kim, E.; Sheng, M. PDZ domain proteins of synapses. Nat. Rev. Neurosci. 2004, 5, 771–781. [Google Scholar] [CrossRef]
- Han, J.; Wu, P.; Wang, F.; Chen, J. S-palmitoylation regulates AMPA receptors trafficking and function: A novel insight into synaptic regulation and therapeutics. Acta Pharm. Sin. B 2015, 5, 1–7. [Google Scholar] [CrossRef]
- Thomas, G.M.; Hayashi, T.; Huganir, R.L.; Linden, D.J. DHHC8-dependent PICK1 palmitoylation is required for induction of cerebellar long-term synaptic depression. J. Neurosci. 2013, 33, 15401–15407. [Google Scholar] [CrossRef]
- Keith, D.J.; Sanderson, J.L.; Gibson, E.S.; Woolfrey, K.M.; Robertson, H.R.; Olszewski, K.; Kang, R.; El-Husseini, A.; Dell’Acqua, M.L. Palmitoylation of A-Kinase anchoring protein 79/150 regulates dendritic endosomal targeting and synaptic plasticity mechanisms. J. Neurosci. 2012, 32, 7119–7136. [Google Scholar] [CrossRef]
- Li, M.-D.; Huang, D.-H.; Zheng, Y.-Q.; Tina, D.; OuYang, H.; Song, K.-X.; Huang, Z.; Lin, X.-S.; Zhu, X.-Q.; Huang, J.-J.; et al. HHC2-Mediated AKAP150 Palmitoylation Regulates Hippocampal Synaptic Plasticity and Fear Memory. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Sanderson, J.L.; Scott, J.D.; Dell’Acqua, M.L. Control of Homeostatic Synaptic Plasticity by AKAP-Anchored Kinase and Phosphatase Regulation of Ca2+-Permeable AMPA Receptors. J. Neurosci. 2018, 38, 2863–2876. [Google Scholar] [CrossRef]
- Wang, T.; Liu, Y.; Liu, X.; Wei, X.; Ding, X.; Mo, L.; Wang, H.; Zhou, W.; Xu, T.; Deng, F.; et al. Golgi-specific DHHC type zinc finger protein is decreased in neurons of intractable epilepsy patients and pentylenetetrazole-kindled rats. Neuroreport 2018, 29, 1157–1165. [Google Scholar] [CrossRef]
- Fang, C.; Deng, L.; Keller, C.A.; Fukata, M.; Fukata, Y.; Chen, G.; Luscher, B. GODZ-Mediated Palmitoylation of GABAA Receptors Is Required for Normal Assembly and Function of GABAergic Inhibitory Synapses. J. Neurosci. 2006, 26, 12758–12768. [Google Scholar] [CrossRef]
- Hayashi, T.; Rumbaugh, G.; Huganir, R.L. Differential Regulation of AMPA Receptor Subunit Trafficking by Palmitoylation of Two Distinct Sites. Neuron 2005, 47, 709–723. [Google Scholar] [CrossRef]
- Chowdhury, D.; Turner, M.; Patriarchi, T.; Hergarden, A.C.; Anderson, D.; Zhang, Y.; Sun, J.; Chen, C.-Y.; Ames, J.B.; Hell, J.W. Ca2+/calmodulin binding to PSD-95 mediates homeostatic synaptic scaling down. EMBO J. 2018, 37, 122–138. [Google Scholar] [CrossRef]
- Kalashnikova, E.; Lorca, R.; Kaur, I.; Barisone, G.A.; Li, B.; Ishimaru, T.; Trimmer, J.S.; Mohapatra, D.P.; Díaz, E. SynDIG1: An Activity-Regulated, AMPA- Receptor-Interacting Transmembrane Protein that Regulates Excitatory Synapse Development. Neuron 2010, 65, 80–93. [Google Scholar] [CrossRef]
- Kaur, I.; Yarov-Yarovoy, V.; Kirk, L.M.; Plambeck, K.E.; Barragan, E.V.; Ontiveros, E.S.; Diaz, E. Activity-Dependent Palmitoylation Controls SynDIG1 Stability, Localization, and Function. J. Neurosci. 2016, 36, 7562–7568. [Google Scholar] [CrossRef]
- Xia, Z.-X.; Shen, Z.-C.; Zhang, S.-Q.; Wang, J.; Nie, T.-L.; Deng, Q.; Chen, J.-G.; Wang, F.; Wu, P.-F. De-palmitoylation by N-(tert-Butyl) hydroxylamine inhibits AMPAR-mediated synaptic transmission via affecting receptor distribution in postsynaptic densities. CNS Neurosci. Ther. 2018, 25, 187–199. [Google Scholar] [CrossRef] [PubMed]
- El-Husseini, A.E.D.; Schnell, E.; Dakoji, S.; Sweeney, N.; Zhou, Q.; Prange, O.; Gauthier-Campbell, C.; Aguilera-Moreno, A.; Nicoll, R.A.; Bredt, D.S. Synaptic strength regulated by palmitate cycling on PSD-95. Cell 2002, 108, 849–863. [Google Scholar] [CrossRef]
- Spinelli, M.; Fusco, S.; Mainardi, M.; Scala, F.; Natale, F.; Lapenta, R.; Mattera, A.; Rinaudo, M.; Puma, D.D.L.; Ripoli, C.; et al. Brain insulin resistance impairs hippocampal synaptic plasticity and memory by increasing GluA1 palmitoylation through FoxO3a. Nat. Commun. 2017, 8, 2009. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.-C.; Xia, Z.-X.; Liu, J.-M.; Zheng, J.-Y.; Luo, Y.-F.; Yang, H.; Li, M.-D.; Cao, T.; Liu, H.-P.; Jin, G.-L.; et al. APT1-Mediated Depalmitoylation Regulates Hippocampal Synaptic Plasticity. J. Neurosci. 2022, 42, 2662–2677. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Lin, Y.; Luo, J.; Yu, J.; Cheng, Y.; Wu, X.; Lin, L.; Lin, Y. Maternal High-Fat Diet Multigenerationally Impairs Hippocampal Synaptic Plasticity and Memory in Male Rat Offspring. Endocrinology 2021, 162, bqaa214. [Google Scholar] [CrossRef]
- Steinberg, J.P.; Takamiya, K.; Shen, Y.; Xia, J.; Rubio, M.E.; Yu, S.; Jin, W.; Thomas, G.M.; Linden, D.J.; Huganir, R.L. Targeted In Vivo Mutations of the AMPA Receptor Subunit GluR2 and Its Interacting Protein PICK1 Eliminate Cerebellar Long-Term Depression. Neuron 2006, 49, 845–860. [Google Scholar] [CrossRef]
- Itoh, M.; Yamashita, M.; Kaneko, M.; Okuno, H.; Abe, M.; Yamazaki, M.; Natsume, R.; Yamada, D.; Kaizuka, T.; Suwa, R.; et al. Deficiency of AMPAR–Palmitoylation Aggravates Seizure Susceptibility. J. Neurosci. 2018, 38, 10220–10235. [Google Scholar] [CrossRef]
- Iizumi, M.; Oota-Ishigaki, A.; Yamashita, M.; Hayashi, T. Reduced Effect of Anticonvulsants on AMPA Receptor Palmitoylation-Deficient Mice. Front. Pharmacol. 2021, 12, 711737. [Google Scholar] [CrossRef]
- Hayashi, T.; Thomas, G.M.; Huganir, R.L. Dual Palmitoylation of NR2 Subunits Regulates NMDA Receptor Trafficking. Neuron 2009, 64, 213–226. [Google Scholar] [CrossRef]
- Mattison, H.A.; Hayashi, T.; Barria, A. Palmitoylation at Two Cysteine Clusters on the C-Terminus of GluN2A and GluN2B Differentially Control Synaptic Targeting of NMDA Receptors. PLoS ONE 2012, 7, e49089. [Google Scholar] [CrossRef]
- Hubalkova, P.; Ladislav, M.; Vyklicky, V.; Smejkalova, T.; Krausova, B.H.; Kysilov, B.; Krusek, J.; Naimová, Z.; Korinek, M.; Chodounska, H.; et al. Palmitoylation Controls NMDA Receptor Function and Steroid Sensitivity. J. Neurosci. 2021, 41, 2119–2134. [Google Scholar] [CrossRef]
- Stein, V.; House, D.R.C.; Bredt, D.S.; Nicoll, R.A. Postsynaptic Density-95 Mimics and Occludes Hippocampal Long-Term Potentiation and Enhances Long-Term Depression. J. Neurosci. 2003, 23, 5503–5506. [Google Scholar] [CrossRef]
- Schnell, E.; Sizemore, M.; Karimzadegan, S.; Chen, L.; Bredt, D.S.; Nicoll, R.A. Direct interactions between PSD-95 and stargazin control synaptic AMPA receptor number. Proc. Natl. Acad. Sci. USA 2002, 99, 13902–13907. [Google Scholar] [CrossRef]
- Jeyifous, O.; Lin, E.I.; Chen, X.; Antinone, S.E.; Mastro, R.; Drisdel, R.; Reese, T.S.; Green, W.N. Palmitoylation regulates glutamate receptor distributions in postsynaptic densities through control of PSD95 conformation and orientation. Proc. Natl. Acad. Sci. USA 2016, 113, E8482–E8491. [Google Scholar] [CrossRef]
- Purkey, A.M.; Woolfrey, K.M.; Crosby, K.C.; Stich, D.G.; Chick, W.S.; Aoto, J.; Dell’Acqua, M.L. AKAP150 Palmitoylation Regulates Synaptic Incorporation of Ca2+ -Permeable AMPA Receptors to Control LTP. Cell Rep. 2018, 25, 974–987.e4. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Crosby, K.C.; Feng, A.; Purkey, A.M.; Aronova, M.A.; Winters, C.A.; Crocker, V.T.; Leapman, R.D.; Reese, T.S.; Dell’Acqua, M.L. Palmitoylation of A-kinase anchoring protein 79/150 modulates its nanoscale organization, trafficking, and mobility in postsynaptic spines. Front. Synaptic Neurosci. 2022, 14, 1004154. [Google Scholar] [CrossRef] [PubMed]
- DeSouza, S.; Fu, J.; States, B.A.; Ziff, E.B. Differential Palmitoylation Directs the AMPA Receptor-Binding Protein ABP to Spines or to Intracellular Clusters. J. Neurosci. 2002, 22, 3493–3503. [Google Scholar] [CrossRef] [PubMed]
- Misra, C.; Restituito, S.; Ferreira, J.; Rameau, G.A.; Fu, J.; Ziff, E.B. Regulation of synaptic structure and function by palmitoylated AMPA receptor binding protein. Mol. Cell. Neurosci. 2010, 43, 341–352. [Google Scholar] [CrossRef]
- Yamazaki, M.; Fukaya, M.; Abe, M.; Ikeno, K.; Kakizaki, T.; Watanabe, M.; Sakimura, K. Differential palmitoylation of two mouse glutamate receptor interacting protein 1 forms with different N-terminal sequences. Neurosci. Lett. 2001, 304, 81–84. [Google Scholar] [CrossRef]
- Lu, W.; Ziff, E.B. PICK1 Interacts with ABP/GRIP to Regulate AMPA Receptor Trafficking. Neuron 2005, 47, 407–421. [Google Scholar] [CrossRef]
- Shepherd, J.; Bear, M.F. New views of Arc, a master regulator of synaptic plasticity. Nat. Neurosci. 2011, 14, 279–284. [Google Scholar] [CrossRef]
- Zhang, H.; Bramham, C.R. Arc/Arg3.1 function in long-term synaptic plasticity: Emerging mechanisms and unresolved issues. Eur. J. Neurosci. 2021, 54, 6696–6712. [Google Scholar] [CrossRef]
- Barylko, B.; Wilkerson, J.R.; Cavalier, S.H.; Binns, D.D.; James, N.G.; Jameson, D.M.; Huber, K.M.; Albanesi, J.P. Palmitoylation and Membrane Binding of Arc/Arg3.1: A Potential Role in Synaptic Depression. Biochemistry 2017, 57, 520–524. [Google Scholar] [CrossRef]
- Cho, E.; Kim, D.-H.; Hur, Y.-N.; Whitcomb, D.; Regan, P.; Hong, J.-H.; Kim, H.; Suh, Y.H.; Cho, K.; Park, M. Cyclin Y inhibits plasticity-induced AMPA receptor exocytosis and LTP. Sci. Rep. 2015, 5, 12624. [Google Scholar] [CrossRef]
- Seo, J.; Hwang, H.; Choi, Y.; Jung, S.; Hong, J.-H.; Yoon, B.-J.; Rhim, H.; Park, M. Myristoylation-dependent palmitoylation of cyclin Y modulates long-term potentiation and spatial learning. Prog. Neurobiol. 2022, 218, 102349. [Google Scholar] [CrossRef]
- Yan, P.; Liu, H.; Zhou, T.; Sun, P.; Wang, Y.; Wang, X.; Zhang, L.; Wang, T.; Dong, J.; Zhu, J.; et al. Crosstalk of Synapsin1 palmitoylation and phosphorylation controls the dynamicity of synaptic vesicles in neurons. Cell Death Dis. 2022, 13, 786. [Google Scholar] [CrossRef]
- Rizo, J. Mechanism of neurotransmitter release coming into focus. Protein Sci. 2018, 27, 1364–1391. [Google Scholar] [CrossRef]
- Vardar, G.; Salazar-Lázaro, A.; Zobel, S.; Trimbuch, T.; Rosenmund, C. Syntaxin-1A modulates vesicle fusion in mammalian neurons via juxtamembrane domain dependent palmitoylation of its transmembrane domain. eLife 2022, 11, e78182C. [Google Scholar] [CrossRef]
- Tyagarajan, S.K.; Fritschy, J.M. Gephyrin: A master regulator of neuronal function? Nat. Rev. Neurosci. 2014, 15, 141–156. [Google Scholar] [CrossRef]
- Zaręba-Kozioł, M.; Bartkowiak-Kaczmarek, A.; Roszkowska, M.; Bijata, K.; Figiel, I.; Halder, A.; Kamińska, P.; Müller, F.; Basu, S.; Zhang, W.; et al. S-Palmitoylation of Synaptic Proteins as a Novel Mechanism Underlying Sex-Dependent Differences in Neuronal Plasticity. Int. J. Mol. Sci. 2021, 22, 6253. [Google Scholar] [CrossRef]
- Hohoff, C.; Zhang, M.; Ambrée, O.; Kravchenko, M.; Buschert, J.; Kerkenberg, N.; Gorinski, N.; Galil, D.A.; Schettler, C.; Werth, K.L.V.; et al. Deficiency of the palmitoyl acyltransferase ZDHHC7 impacts brain and behavior of mice in a sex-specific manner. Anat. Embryol. 2019, 224, 2213–2230. [Google Scholar] [CrossRef]
- Jia, L.; Chisari, M.; Maktabi, M.H.; Sobieski, C.; Zhou, H.; Konopko, A.M.; Martin, B.R.; Mennerick, S.J.; Blumer, K.J. A Mechanism Regulating G Protein-coupled Receptor Signaling That Requires Cycles of Protein Palmitoylation and Depalmitoylation. J. Biol. Chem. 2014, 289, 6249–6257. [Google Scholar] [CrossRef]
- Shipston, M.J. Ion Channel Regulation by Protein Palmitoylation. J. Biol. Chem. 2011, 286, 8709–8716. [Google Scholar] [CrossRef]
- Kilpatrick, C.L.; Murakami, S.; Feng, M.; Wu, X.; Lal, R.; Chen, G.; Du, K.; Luscher, B. Dissociation of Golgi-associated DHHC-type Zinc Finger Protein (GODZ)- and Sertoli Cell Gene with a Zinc Finger Domain-β (SERZ-β)-mediated Palmitoylation by Loss of Function Analyses in Knock-out Mice. J. Biol. Chem. 2016, 291, 27371–27386. [Google Scholar] [CrossRef]
- Hines, R.; Kang, R.; Goytain, A.; Quamme, G.A. Golgi-specific DHHC Zinc Finger Protein GODZ Mediates Membrane Ca2+ Transport. J. Biol. Chem. 2010, 285, 4621–4628. [Google Scholar] [CrossRef] [PubMed]
- Goytain, A.; Hines, R.; Quamme, G.A. Huntingtin-interacting Proteins, HIP14 and HIP14L, Mediate Dual Functions, Palmitoyl Acyltransferase and Mg2+ Transport. J. Biol. Chem. 2008, 283, 33365–33374. [Google Scholar] [CrossRef] [PubMed]
- Shimell, J.J.; Globa, A.; Sepers, M.D.; Wild, A.R.; Matin, N.; Raymond, L.A.; Bamji, S.X. Regulation of hippocampal excitatory synapses by the Zdhhc5 palmitoyl acyltransferase. J. Cell Sci. 2021, 134, jcs254276. [Google Scholar] [CrossRef] [PubMed]
- Meitzen, J.; Britson, K.A.; Tuomela, K.; Mermelstein, P.G. The expression of select genes necessary for membrane-associated estrogen receptor signaling differ by sex in adult rat hippocampus. Steroids 2017, 142, 21–27. [Google Scholar] [CrossRef]
- Kerkenberg, N.; Hohoff, C.; Zhang, M.; Lang, I.; Schettler, C.; Ponimaskin, E.; Wachsmuth, L.; Faber, C.; Baune, B.T.; Zhang, W. Acute stress reveals different impacts in male and female Zdhhc7-deficient mice. Anat. Embryol. 2021, 226, 1613–1626. [Google Scholar] [CrossRef]
- Mukai, J.; Dhilla, A.; Drew, L.J.; Stark, K.L.; Cao, L.; MacDermott, A.B.; Karayiorgou, M.; Gogos, J.A. Palmitoylation-dependent neurodevelopmental deficits in a mouse model of 22q11 microdeletion. Nat. Neurosci. 2008, 11, 1302–1310. [Google Scholar] [CrossRef]
- Mukai, J.; Tamura, M.; Fénelon, K.; Rosen, A.M.; Spellman, T.J.; Kang, R.; MacDermott, A.B.; Karayiorgou, M.; Gordon, J.A.; Gogos, J.A. Molecular Substrates of Altered Axonal Growth and Brain Connectivity in a Mouse Model of Schizophrenia. Neuron 2015, 86, 680–695. [Google Scholar] [CrossRef]
- Young, F.B.; Butland, S.L.; Sanders, S.S.; Sutton, L.M.; Hayden, M.R. Putting proteins in their place: Palmitoylation in Huntington disease and other neuropsychiatric diseases. Prog. Neurobiol. 2012, 97, 220–238. [Google Scholar] [CrossRef]
- Milnerwood, A.J.; Parsons, M.P.; Young, F.B.; Singaraja, R.R.; Franciosi, M.V.; Bergeron, S.; Hayden, M.R.; Raymond, L.A. Memory and synaptic deficits in Hip14/DHHC17 knockout mice. Proc. Natl. Acad. Sci. USA 2013, 110, 20296–20301. [Google Scholar] [CrossRef]
- Sanders, S.S.; Parsons, M.P.; Mui, K.K.N.; Southwell, A.L.; Franciosi, S.; Cheung, D.; Waltl, S.; Raymond, L.A.; Hayden, M.R. Sudden death due to paralysis and synaptic and behavioral deficits when Hip14/Zdhhc17 is deleted in adult mice. BMC Biol. 2016, 14, 108. [Google Scholar] [CrossRef]
- Singaraja, R.R.; Huang, K.; Sanders, S.S.; Milnerwood, A.J.; Hines, R.; Lerch, J.P.; Franciosi, S.; Drisdel, R.C.; Vaid, K.; Young, F.B.; et al. Altered palmitoylation and neuropathological deficits in mice lacking HIP14. Hum. Mol. Genet. 2011, 20, 3899–3909. [Google Scholar] [CrossRef]
- Nelson, J.C.; Witze, E.; Ma, Z.; Ciocco, F.; Frerotte, A.; Randlett, O.; Foskett, J.K.; Granato, M. Acute Regulation of Habituation Learning via Posttranslational Palmitoylation. Curr. Biol. 2020, 30, 2729–2738.e4. [Google Scholar] [CrossRef]
- Rankin, C.H.; Abrams, T.; Barry, R.J.; Bhatnagar, S.; Clayton, D.F.; Colombo, J.; Coppola, G.; Geyer, M.A.; Glanzman, D.L.; Marsland, S.; et al. Habituation revisited: An updated and revised description of the behavioral characteristics of habituation. Neurobiol. Learn. Mem. 2009, 92, 135–138. [Google Scholar] [CrossRef]
- Koster, K.P.; Francesconi, W.; Berton, F.; Alahmadi, S.; Srinivas, R.; Yoshii, A. Developmental NMDA receptor dysregulation in the infantile neuronal ceroid lipofuscinosis mouse model. eLife 2019, 8, e40316. [Google Scholar] [CrossRef]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA receptor subunit diversity: Impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef]
- Sapir, T.; Segal, M.; Grigoryan, G.; Hansson, K.M.; James, P.; Segal, M.; Reiner, O. The Interactome of Palmitoyl-Protein Thioesterase 1 (PPT1) Affects Neuronal Morphology and Function. Front. Cell. Neurosci. 2019, 13, 92. [Google Scholar] [CrossRef]
- Koster, K.P.; Flores-Barrera, E.; Artur de la Villarmois, E.; Nguyen, T.T.; Niqula, A.; Noriega-Gonzalez, L.Y.; Fyke, Z.; Caballero, A.; Cologna, S.M.; Tseng, K.Y.; et al. Loss of Depalmitoylation Exaggerates Synaptic Upscaling and Leads to Neuroinflammation in a Lysosomal Storage Disease. bioRxiv 2021. [Google Scholar] [CrossRef]
- Tong, J.; Gao, J.; Feng, B.; Zhao, X.; Li, J.; Qi, Y.; Zhang, Q.; Lu, C.; Peng, S. PPT1 Deciency-Induced GABAAR Hyperpalmitoylation Impairs Synaptic Transmission and Memory Formation. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Demontis, G.C.; Pezzini, F.; Margari, E.; Bianchi, M.; Longoni, B.; Doccini, S.; Lalowski, M.M.; Santorelli, F.M.; Simonati, A. Electrophysiological Profile Remodeling via Selective Suppression of Voltage-Gated Currents by CLN1/PPT1 Overexpression in Human Neuronal-Like Cells. Front. Cell. Neurosci. 2020, 14, 569598. [Google Scholar] [CrossRef]
- Kim, S.-J.; Zhang, Z.; Sarkar, C.; Tsai, P.-C.; Lee, Y.-C.; Dye, L.; Mukherjee, A.B. Palmitoyl protein thioesterase-1 deficiency impairs synaptic vesicle recycling at nerve terminals, contributing to neuropathology in humans and mice. J. Clin. Investig. 2008, 118, 3075–3086. [Google Scholar] [CrossRef]
- Attinger, A.; Wang, B.; Keller, G.B. Visuomotor Coupling Shapes the Functional Development of Mouse Visual Cortex. Cell 2017, 169, 1291–1302.e14. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Grimwood, P.D.; Morris, R.G.M. An Evaluation of the Hypothesis. Annu. Rev. Neurosci 2000, 23, 649–711. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, J.; Höfer, K.; Wong, A.M.; Cooper, J.; Birnbaum, S.G.; Hammer, R.E.; Hofmann, S.L. DHHC5 Interacts with PDZ Domain 3 of Post-synaptic Density-95 (PSD-95) Protein and Plays a Role in Learning and Memory. J. Biol. Chem. 2010, 285, 13022–13031. [Google Scholar] [CrossRef] [PubMed]
- Kouskou, M.; Thomson, D.M.; Brett, R.R.; Wheeler, L.; Tate, R.J.; Pratt, J.A.; Chamberlain, L.H. Disruption of the Zdhhc9 intellectual disability gene leads to behavioural abnormalities in a mouse model. Exp. Neurol. 2018, 308, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Wallis, T.P.; Venkatesh, B.G.; Narayana, V.K.; Kvaskoff, D.; Ho, A.; Sullivan, R.K.; Windels, F.; Sah, P.; Meunier, F.A. Saturated free fatty acids and association with memory formation. Nat. Commun. 2021, 12, 3443. [Google Scholar] [CrossRef]
- Lacombe, R.J.S.; Giuliano, V.; Chouinard-Watkins, R.; Bazinet, R.P. Natural Abundance Carbon Isotopic Analysis Indicates the Equal Contribution of Local Synthesis and Plasma Uptake to Palmitate Levels in the Mouse Brain. Lipids 2018, 53, 481–490. [Google Scholar] [CrossRef]
- Smith, M.E.; Cisbani, G.; Metherel, A.H.; Bazinet, R.P. The majority of brain palmitic acid is maintained by lipogenesis from dietary sugars and is augmented in mice fed low palmitic acid levels from birth. J. Neurochem. 2022, 161, 112–128. [Google Scholar] [CrossRef]
- Vesga-Jiménez, D.J.; Martin, C.; Barreto, G.E.; Aristizábal-Pachón, A.F.; Pinzón, A.; González, J. Fatty Acids: An Insight into the Pathogenesis of Neurodegenerative Diseases and Therapeutic Potential. Int. J. Mol. Sci. 2022, 23, 2577. [Google Scholar] [CrossRef]
- Melo, H.M.; Silva, G.D.S.S.D.; Sant’Ana, M.R.; Teixeira, C.V.L.; Clarke, J.R.; Coreixas, V.S.M.; de Melo, B.C.; Fortuna, J.T.; Forny-Germano, L.; Ledo, J.H.; et al. Palmitate Is Increased in the Cerebrospinal Fluid of Humans with Obesity and Induces Memory Impairment in Mice via Pro-inflammatory TNF-α. Cell Rep. 2020, 30, 2180–2194.e8. [Google Scholar] [CrossRef]
- Oota-Ishigaki, A.; Takao, K.; Yamada, D.; Sekiguchi, M.; Itoh, M.; Koshidata, Y.; Abe, M.; Natsume, R.; Kaneko, M.; Adachi, T.; et al. Prolonged contextual fear memory in AMPA receptor palmitoylation-deficient mice. Neuropsychopharmacology 2022, 47, 2150–2159. [Google Scholar] [CrossRef]
- Cho, E.; Park, M. Palmitoylation in Alzheimer’s disease and other neurodegenerative diseases. Pharmacol. Res. 2016, 111, 133–151. [Google Scholar] [CrossRef] [PubMed]
- Koster, K.P.; Yoshii, A. Depalmitoylation by Palmitoyl-Protein Thioesterase 1 in Neuronal Health and Degeneration. Front. Synaptic Neurosci. 2019, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Sanders, S.S.; Hayden, M.R. Aberrant palmitoylation in Huntington disease. Biochem. Soc. Trans. 2015, 43, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Ivatt, R.M.; Sanchez-Martinez, A.; Godena, V.K.; Brown, S.; Ziviani, E.; Whitworth, A.J. Genome-wide RNAi screen identifies the Parkinson disease GWAS risk locus SREBF1 as a regulator of mitophagy. Proc. Natl. Acad. Sci. USA 2014, 111, 8494–8499. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zheng, F.; Hu, Y.; Yang, Y.; Li, Y.; Chen, G.; Wang, W.; He, M.; Zhou, R.; Ma, Y.; et al. ZDHHC8 critically regulates seizure susceptibility in epilepsy. Cell Death Dis. 2018, 9, 795. [Google Scholar] [CrossRef]
- Cervilla-Martínez, J.F.; Rodríguez-Gotor, J.J.; Wypijewski, K.J.; Fontán-Lozano, Á.; Wang, T.; Santamaría, E.; Fuller, W.; Mejías, R. Altered Cortical Palmitoylation Induces Widespread Molecular Disturbances in Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 14018. [Google Scholar] [CrossRef]
- Funahashi, S. Working Memory in the Prefrontal Cortex. Brain Sci. 2017, 7, 49. [Google Scholar] [CrossRef]
- Magnusson, K.R. Aging of the NMDA receptor: From a mouse’s point of view. Futur. Neurol. 2012, 7, 627–637. [Google Scholar] [CrossRef]
- Zamzow, D.R.; Elias, V.; Acosta, V.A.; Escobedo, E.; Magnusson, K.R. Higher Levels of Protein Palmitoylation in the Frontal Cortex across Aging Were Associated with Reference Memory and Executive Function Declines. eNeuro 2019, 6, e0310-18. [Google Scholar] [CrossRef]
- Cárcel-Trullols, J.; Kovács, A.D.; Pearce, D.A. Cell biology of the NCL proteins: What they do and don’t do. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 2242–2255. [Google Scholar] [CrossRef]
- Augustine, E.F.; Adams, H.R.; Reyes, E.D.L.; Drago, K.; Frazier, M.; Guelbert, N.; Laine, M.; Levin, T.; Mink, J.W.; Nickel, M.; et al. Management of CLN1 Disease: International Clinical Consensus. Pediatr. Neurol. 2021, 120, 38–51. [Google Scholar] [CrossRef]
- Mole, S.E.; Anderson, G.; Band, H.A.; Berkovic, S.F.; Cooper, J.D.; Holthaus, S.-M.K.; McKay, T.R.; Medina, D.L.; Rahim, A.; Schulz, A.; et al. Clinical challenges and future therapeutic approaches for neuronal ceroid lipofuscinosis. Lancet Neurol. 2018, 18, 107–116. [Google Scholar] [CrossRef]
- Saja, S.; Buff, H.; Smith, A.C.; Williams, T.S.; Korey, C.A. Identifying cellular pathways modulated by Drosophila palmitoyl-protein thioesterase 1 function. Neurobiol. Dis. 2010, 40, 135–145. [Google Scholar] [CrossRef]
- Aby, E.; Gumps, K.; Roth, A.; Sigmon, S.; Jenkins, S.E.; Kim, J.J.; Kramer, N.J.; Parfitt, K.D.; Korey, C.A. Mutations in palmitoyl-protein thioesterase 1 alter exocytosis and endocytosis at synapses in Drosophila larvae. Fly 2013, 7, 267–279. [Google Scholar] [CrossRef]
- Brady, S.; Morfini, G. A Perspective on Neuronal Cell Death Signaling and Neurodegeneration. Mol. Neurobiol. 2010, 42, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.B.; Chen, A.; Kaminsky, S.M.; Crystal, R.G.; Sondhi, D. Advances in the treatment of neuronal ceroid lipofuscinosis. Expert Opin. Orphan Drugs 2019, 7, 473–500. [Google Scholar] [CrossRef] [PubMed]
- McColgan, P.; Tabrizi, S.J. Huntington’s disease: A clinical review. Eur. J. Neurol. 2018, 25, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Saudou, F.; Humbert, S. The Biology of Huntingtin. Neuron 2016, 89, 910–926. [Google Scholar] [CrossRef]
- Lontay, B.; Kiss, A.; Virág, L.; Tar, K. How Do Post-Translational Modifications Influence the Pathomechanistic Landscape of Huntington’s Disease? A Comprehensive Review. Int. J. Mol. Sci. 2020, 21, 4282. [Google Scholar] [CrossRef]
- Yanai, A.; Huang, K.; Kang, R.; Singaraja, R.; Arstikaitis, P.; Gan, L.; Orban, P.C.; Mullard, A.; Cowan, C.M.; Raymond, L.; et al. Palmitoylation of huntingtin by HIP14is essential for its trafficking and function. Nat. Neurosci. 2006, 9, 824–831. [Google Scholar] [CrossRef]
- Sutton, L.M.; Sanders, S.S.; Butland, S.; Singaraja, R.; Franciosi, S.; Southwell, A.L.; Doty, C.N.; Schmidt, M.E.; Mui, K.K.; Kovalik, V.; et al. Hip14l-deficient mice develop neuropathological and behavioural features of Huntington disease. Hum. Mol. Genet. 2012, 22, 452–465. [Google Scholar] [CrossRef]
- Lemarié, F.L.; Caron, N.S.; Sanders, S.S.; Schmidt, M.E.; Nguyen, Y.T.; Ko, S.; Xu, X.; Pouladi, M.A.; Martin, D.D.; Hayden, M.R. Rescue of aberrant huntingtin palmitoylation ameliorates mutant huntingtin-induced toxicity. Neurobiol. Dis. 2021, 158, 105479. [Google Scholar] [CrossRef]
- Kang, R.; Wang, L.; Sanders, S.S.; Zuo, K.; Hayden, M.R.; Raymond, L.A. Altered Regulation of Striatal Neuronal N-Methyl-D-Aspartate Receptor Trafficking by Palmitoylation in Huntington Disease Mouse Model. Front. Synaptic Neurosci. 2019, 11, 3. [Google Scholar] [CrossRef]
- Virlogeux, A.; Scaramuzzino, C.; Lenoir, S.; Carpentier, R.; Louessard, M.; Genoux, A.; Lino, P.; Hinckelmann, M.-V.; Perrier, A.L.; Humbert, S.; et al. Increasing brain palmitoylation rescues behavior and neuropathology in Huntington disease mice. Sci. Adv. 2021, 7, eabb0799. [Google Scholar] [CrossRef]
- Ferrer, I.; Goutan, E.; Marın, C.; Rey, M.J.; Ribalta, T. Brain-derived neurotrophic factor in Huntington disease. Brain Res. 2000, 866, 257–261. [Google Scholar] [CrossRef]
- Gauthier, L.R.; Charrin, B.C.; Borrell-Pagès, M.; Dompierre, J.P.; Rangone, H.; Cordelières, F.P.; De Mey, J.; MacDonald, M.E.; Leßmann, V.; Humbert, S.; et al. Huntingtin Controls Neurotrophic Support and Survival of Neurons by Enhancing BDNF Vesicular Transport along Microtubules. Cell 2004, 118, 127–138. [Google Scholar] [CrossRef]
- Graham, R.K.; Ehrnhoefer, D.E.; Hayden, M.R. Caspase-6 and neurodegeneration. Trends Neurosci. 2011, 34, 646–656. [Google Scholar] [CrossRef]
- Skotte, N.H.; Sanders, S.S.; Singaraja, R.; Ehrnhoefer, D.E.; Vaid, K.; Qiu, X.; Kannan, S.; Verma, C.; Hayden, M. Palmitoylation of caspase-6 by HIP14 regulates its activation. Cell Death Differ. 2017, 24, 433–444. [Google Scholar] [CrossRef]
- Cline, E.N.; Bicca, M.A.; Viola, K.L.; Klein, W.L. The Amyloid-β Oligomer Hypothesis: Beginning of the Third Decade. J. Alzheimer’s Dis. 2018, 64, S567–S610. [Google Scholar] [CrossRef]
- Soto, C.; Pritzkow, S. Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases. Nat. Neurosci. 2018, 21, 1332–1340. [Google Scholar] [CrossRef]
- Mizumaru, C.; Saito, Y.; Ishikawa, T.; Yoshida, T.; Yamamoto, T.; Nakaya, T.; Suzuki, T. Suppression of APP-containing vesicle trafficking and production of β-amyloid by AID/DHHC-12 protein. J. Neurochem. 2009, 111, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, R.; Barren, C.; Kovacs, D.M. Palmitoylation of Amyloid Precursor Protein Regulates Amyloidogenic Processing in Lipid Rafts. J. Neurosci. 2013, 33, 11169–11183. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, R.; Fenn, R.H.; Barren, C.; Tanzi, R.E.; Kovacs, D.M. Palmitoylated APP Forms Dimers, Cleaved by BACE1. PLoS ONE 2016, 11, e0166400. [Google Scholar] [CrossRef] [PubMed]
- Vetrivel, K.S.; Meckler, X.; Chen, Y.; Nguyen, P.D.; Seidah, N.; Vassar, R.; Wong, P.C.; Fukata, M.; Kounnas, M.Z.; Thinakaran, G. Alzheimer Disease Aβ Production in the Absence of S-Palmitoylation-dependent Targeting of BACE1 to Lipid Rafts. J. Biol. Chem. 2009, 284, 3793–3803. [Google Scholar] [CrossRef]
- Bustos, F.J.; Ampuero, E.; Jury, N.; Aguilar, R.; Falahi, F.; Toledo, J.; Ahumada, J.; Lata, J.; Cubillos, P.; Henriques, B.; et al. Epigenetic editing of the Dlg4/PSD95 gene improves cognition in aged and Alzheimer’s disease mice. Brain 2017, 140, 3252–3268. [Google Scholar] [CrossRef]
- Dore, K.; Carrico, Z.; Alfonso, S.; Marino, M.; Koymans, K.; Kessels, H.W.; Malinow, R. PSD-95 protects synapses from β-amyloid. Cell Rep. 2021, 35, 109194. [Google Scholar] [CrossRef]
- Otte, C.; Gol, S.M.; Penninx, B.W.; Pariante, C.M.; Etkin, A.; Fava, M.; Mohr, D.C.; Schatzberg, A.F. Major depressive disorder. Nat. Rev. Dis. Prim. 2016, 2, 16065. [Google Scholar] [CrossRef]
- Yohn, C.N.; Gergues, M.M.; Samuels, B.A. The role of 5-HT receptors in depression. Mol. Brain 2017, 10, 28. [Google Scholar] [CrossRef]
- Moncrieff, J.; Cooper, R.E.; Stockmann, T.; Amendola, S.; Hengartner, M.P.; Horowitz, M.A. The serotonin theory of depression: A systematic umbrella review of the evidence. Mol. Psychiatry 2022. [Google Scholar] [CrossRef]
- Gorinski, N.; Bijata, M.; Prasad, S.; Wirth, A.; Galil, D.A.; Zeug, A.; Bazovkina, D.; Kodaurova, E.; Kulikova, E.; Ilchibaeva, T.; et al. Attenuated palmitoylation of serotonin receptor 5-HT1A affects receptor function and contributes to depression-like behaviors. Nat. Commun. 2019, 10, 3924. [Google Scholar] [CrossRef]
- Stapel, B.; Gorinski, N.; Gmahl, N.; Rhein, M.; Preuss, V.; Hilfiker-Kleiner, D.; Frieling, H.; Bleich, S.; Ponimaskin, E.; Kahl, K.G. Fluoxetine induces glucose uptake and modifies glucose transporter palmitoylation in human peripheral blood mononuclear cells. Expert Opin. Ther. Targets 2019, 23, 883–891. [Google Scholar] [CrossRef]
- Liu, L.; Dai, L.; Xu, D.; Wang, Y.; Bai, L.; Chen, X.; Li, M.; Yang, S.; Tang, Y. Astrocyte secretes IL-6 to modulate PSD-95 palmitoylation in basolateral amygdala and depression-like behaviors induced by peripheral nerve injury. Brain Behav. Immun. 2022, 104, 139–154. [Google Scholar] [CrossRef]
- Charych, E.I.; Jiang, L.-X.; Lo, F.; Sullivan, K.; Brandon, N.J. Interplay of Palmitoylation and Phosphorylation in the Trafficking and Localization of Phosphodiesterase 10A: Implications for the Treatment of Schizophrenia. J. Neurosci. 2010, 30, 9027–9037. [Google Scholar] [CrossRef]
- Pinner, A.L.; Tucholski, J.; Haroutunian, V.; McCullumsmith, R.E.; Meador-Woodruff, J.H. Decreased protein S-palmitoylation in dorsolateral prefrontal cortex in schizophrenia. Schizophr. Res. 2016, 177, 78–87. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Q.; Qi, Y.; Gao, J.; Li, W.; Lv, L.; Chen, G.; Zhang, Z.; Yue, X.; Peng, S. Enzymatic activity of palmitoyl-protein thioesterase-1 in serum from schizophrenia significantly associates with schizophrenia diagnosis scales. J. Cell. Mol. Med. 2019, 23, 6512–6518. [Google Scholar] [CrossRef]
- Zhao, Y.; He, A.; Zhu, F.; Ding, M.; Hao, J.; Fan, Q.; Li, P.; Liu, L.; Du, Y.; Liang, X.; et al. Integrating genome-wide association study and expression quantitative trait locus study identifies multiple genes and gene sets associated with schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 81, 50–54. [Google Scholar] [CrossRef]
- Maulik, P.K.; Mascarenhas, M.N.; Mathers, C.D.; Dua, T.; Saxena, S. Prevalence of intellectual disability: A meta-analysis of population-based studies. Res. Dev. Disabil. 2011, 32, 419–436. [Google Scholar] [CrossRef]
- Moeschler, J.B.; Shevell, M.; Saul, R.A.; Chen, E.; Freedenberg, D.L.; Hamid, R.; Jones, M.C.; Stoler, J.M.; Tarini, B.A.; Committee on Genetics. Comprehensive Evaluation of the Child With Intellectual Disability or Global Developmental Delays. Pediatrics 2014, 134, e903–e918. [Google Scholar] [CrossRef]
- Ramos, A.K.S.; Caldas-Rosa, E.C.C.; Ferreira, B.M.; Versiani, B.R.; Moretti, P.N.; de Oliveira, S.F.; Pic-Taylor, A.; Mazzeu, J.F. ZDHHC9 X-linked intellectual disability: Clinical and molecular characterization. Am. J. Med. Genet. Part A 2023, 91, 599–604. [Google Scholar] [CrossRef]
- Mansouri, M.R.; Marklund, L.; Gustavsson, P.; Davey, E.; Carlsson, B.; Larsson, C.; White, I.; Gustavson, K.-H.; Dahl, N. Loss of ZDHHC15 expression in a woman with a balanced translocation t(X;15)(q13.3;cen) and severe mental retardation. Eur. J. Hum. Genet. 2005, 13, 970–977. [Google Scholar] [CrossRef]
- Raymond, F.L.; Tarpey, P.S.; Edkins, S.; Tofts, C.; O’Meara, S.; Teague, J.; Butler, A.; Stevens, C.; Barthorpe, S.; Buck, G.; et al. Mutations in ZDHHC9, Which Encodes a Palmitoyltransferase of NRAS and HRAS, Cause X-Linked Mental Retardation Associated with a Marfanoid Habitus. Am. J. Hum. Genet. 2007, 80, 982–987. [Google Scholar] [CrossRef] [PubMed]
- Tarpey, P.S.; Smith, R.; Pleasance, E.; Whibley, A.; Edkins, S.; Hardy, C.; O’Meara, S.; Latimer, C.; Dicks, E.; Menzies, A.; et al. A systematic, large-scale resequencing screen of X-chromosome coding exons in mental retardation. Nat. Genet. 2009, 41, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Branchi, I.; Bichler, Z.; Berger-Sweeney, J.; Ricceri, L. Animal models of mental retardation: From gene to cognitive function. Neurosci. Biobehav. Rev. 2003, 27, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Hamel, L.D.; Reddy, K.D.; Farh, L.; Rettew, L.M.; Sanchez, P.R.; Deschenes, R.J. Mutations in the X-linked Intellectual Disability Gene, zDHHC9, Alter Autopalmitoylation Activity by Distinct Mechanisms. J. Biol. Chem. 2014, 289, 18582–18592. [Google Scholar] [CrossRef]
- Baker, K.; Astle, D.E.; Scerif, G.; Barnes, J.; Smith, J.; Moffat, G.; Gillard, J.; Baldeweg, T.; Raymond, F.L. Epilepsy, cognitive deficits and neuroanatomy in males with ZDHHC 9 mutations. Ann. Clin. Transl. Neurol. 2015, 2, 559–569. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Ponimaskin, E. Palmitoylation as a Functional Regulator of Neurotransmitter Receptors. Neural Plast. 2018, 2018, 5701348. [Google Scholar] [CrossRef]
- Ho, G.P.; Selvakumar, B.; Mukai, J.; Hester, L.D.; Wang, Y.; Gogos, J.A.; Snyder, S.H. S-Nitrosylation and S-Palmitoylation Reciprocally Regulate Synaptic Targeting of PSD-95. Neuron 2011, 71, 131–141. [Google Scholar] [CrossRef]
- Pan, Y.; Xiao, Y.; Pei, Z.; Cummins, T.R. S-Palmitoylation of the sodium channel Nav1.6 regulates its activity and neuronal excitability. J. Biol. Chem. 2020, 295, 6151–6164. [Google Scholar] [CrossRef]
- Huang, K.; Kang, M.H.; Askew, C.; Kang, R.; Sanders, S.S.; Wan, J.; Davis, N.G.; Hayden, M.R. Palmitoylation and function of glial glutamate transporter-1 is reduced in the YAC128 mouse model of Huntington disease. Neurobiol. Dis. 2010, 40, 207–215. [Google Scholar] [CrossRef]
- Sobocińska, J.; Roszczenko-Jasinska, P.; Zaręba-Kozioł, M.; Hromada-Judycka, A.; Matveichuk, O.; Traczyk, G.; Łukasiuk, K.; Kwiatkowska, K. Lipopolysaccharide Upregulates Palmitoylated Enzymes of the Phosphatidylinositol Cycle: An Insight from Proteomic Studies. Mol. Cell. Proteom. 2018, 17, 233–254. [Google Scholar] [CrossRef]
- Mikic, I.; Planey, S.; Zhang, J.; Ceballos, C.; Seron, T.; von Massenbach, B.; Watson, R.; Callaway, S.; McDonough, P.M.; Price, J.H.; et al. A Live Cell, Image-Based Approach to Understanding the Enzymology and Pharmacology of 2-Bromopalmitate and Palmitoylation. Methods Enzymol. 2006, 414, 150–187. [Google Scholar]
- Wang, C.; Chen, X.; Shi, W.; Wang, F.; Du, Z.; Li, X.; Yao, Y.; Liu, T.; Shao, T.; Li, G.; et al. 2-Bromopalmitate impairs neural stem/progenitor cell proliferation, promotes cell apoptosis and induces malformation in zebrafish embryonic brain. Neurotoxicol. Teratol. 2015, 50, 53–63. [Google Scholar] [CrossRef]
- Zmuda, F.; Chamberlain, L.H. Regulatory effects of post-translational modifications on zDHHC S-acyltransferases. J. Biol. Chem. 2020, 295, 14640–14652. [Google Scholar] [CrossRef]
- Gauthier-Kemper, A.; Igaev, M.; Sündermann, F.; Janning, D.; Brühmann, J.; Moschner, K.; Reyher, H.-J.; Junge, W.; Glebov, K.; Walter, J.; et al. Interplay between phosphorylation and palmitoylation mediates plasma membrane targeting and sorting of GAP43. Mol. Biol. Cell 2014, 25, 3284–3299. [Google Scholar] [CrossRef]
- Salaun, C.; Greaves, J.; Chamberlain, L.H. The intracellular dynamic of protein palmitoylation. J. Cell Biol. 2010, 191, 1229–1238. [Google Scholar] [CrossRef]
- Lan, T.; Delalande, C.; Dickinson, B.C. Inhibitors of DHHC family proteins. Curr. Opin. Chem. Biol. 2021, 65, 118–125. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Class of Lipid Modification | Type of Lipid Modification | PTM Character | Lipid | Amino Acid | Linkage |
|---|---|---|---|---|---|
| S-acylation | S-palmitoylation | reversible | Palmitate (16:0) | cysteine | thioester |
| Stearate (18:0) | cysteine | thioester | |||
| Oleate (18:1) | cysteine | thioester | |||
| Arachidonate (20:4n-6) | cysteine | thioester | |||
| Eicosapentanoate (20:5n-3) | cysteine | thioester | |||
| N-myristoylation | N-myristoylation | stable | Myristate (14:0) | N-terminal glycine | amide |
| Isoprenylation | S-farnesylation | stable | Farnesyl (15:0) | cysteine | thioester |
| S-geranylgeranylation | stable | Geranylgeranyl (20:0) | cysteine | thioester | |
| Fatty acylation of secreted proteins by membrane bound O-acyl transferases | N-palmitoylation | stable | Palmitate (16:0) | N-terminal glycine | amide |
| O-acylation | stable | Octanoate (8:0) | cysteine | oxyester | |
| reversible | monounsaturated form of palmitate (palmitoleic 16:1Δ9) | cysteine | oxyester |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buszka, A.; Pytyś, A.; Colvin, D.; Włodarczyk, J.; Wójtowicz, T. S-Palmitoylation of Synaptic Proteins in Neuronal Plasticity in Normal and Pathological Brains. Cells 2023, 12, 387. https://doi.org/10.3390/cells12030387
Buszka A, Pytyś A, Colvin D, Włodarczyk J, Wójtowicz T. S-Palmitoylation of Synaptic Proteins in Neuronal Plasticity in Normal and Pathological Brains. Cells. 2023; 12(3):387. https://doi.org/10.3390/cells12030387
Chicago/Turabian StyleBuszka, Anna, Agata Pytyś, Domnic Colvin, Jakub Włodarczyk, and Tomasz Wójtowicz. 2023. "S-Palmitoylation of Synaptic Proteins in Neuronal Plasticity in Normal and Pathological Brains" Cells 12, no. 3: 387. https://doi.org/10.3390/cells12030387
APA StyleBuszka, A., Pytyś, A., Colvin, D., Włodarczyk, J., & Wójtowicz, T. (2023). S-Palmitoylation of Synaptic Proteins in Neuronal Plasticity in Normal and Pathological Brains. Cells, 12(3), 387. https://doi.org/10.3390/cells12030387