
Therapeutic Potential of Clusterin Inhibition in Human Cancer

Abstract
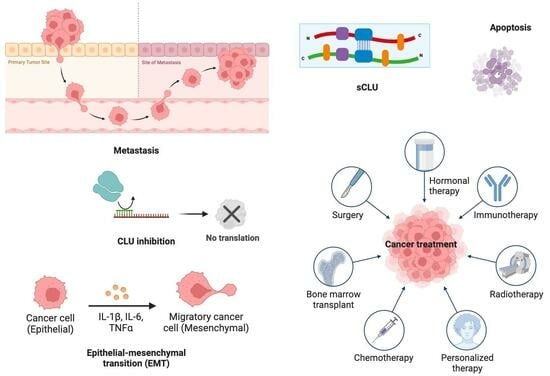
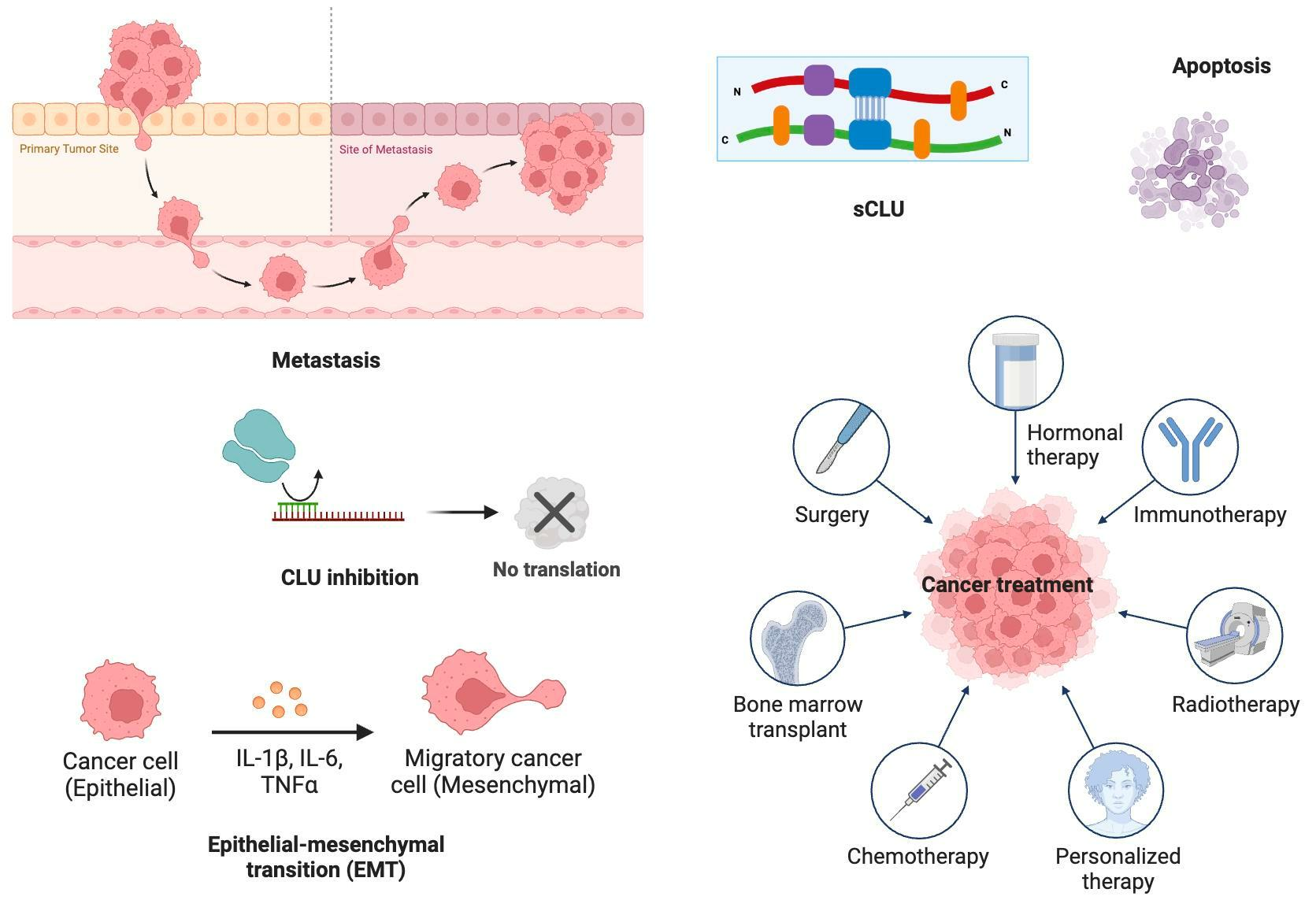
:
1. Introduction
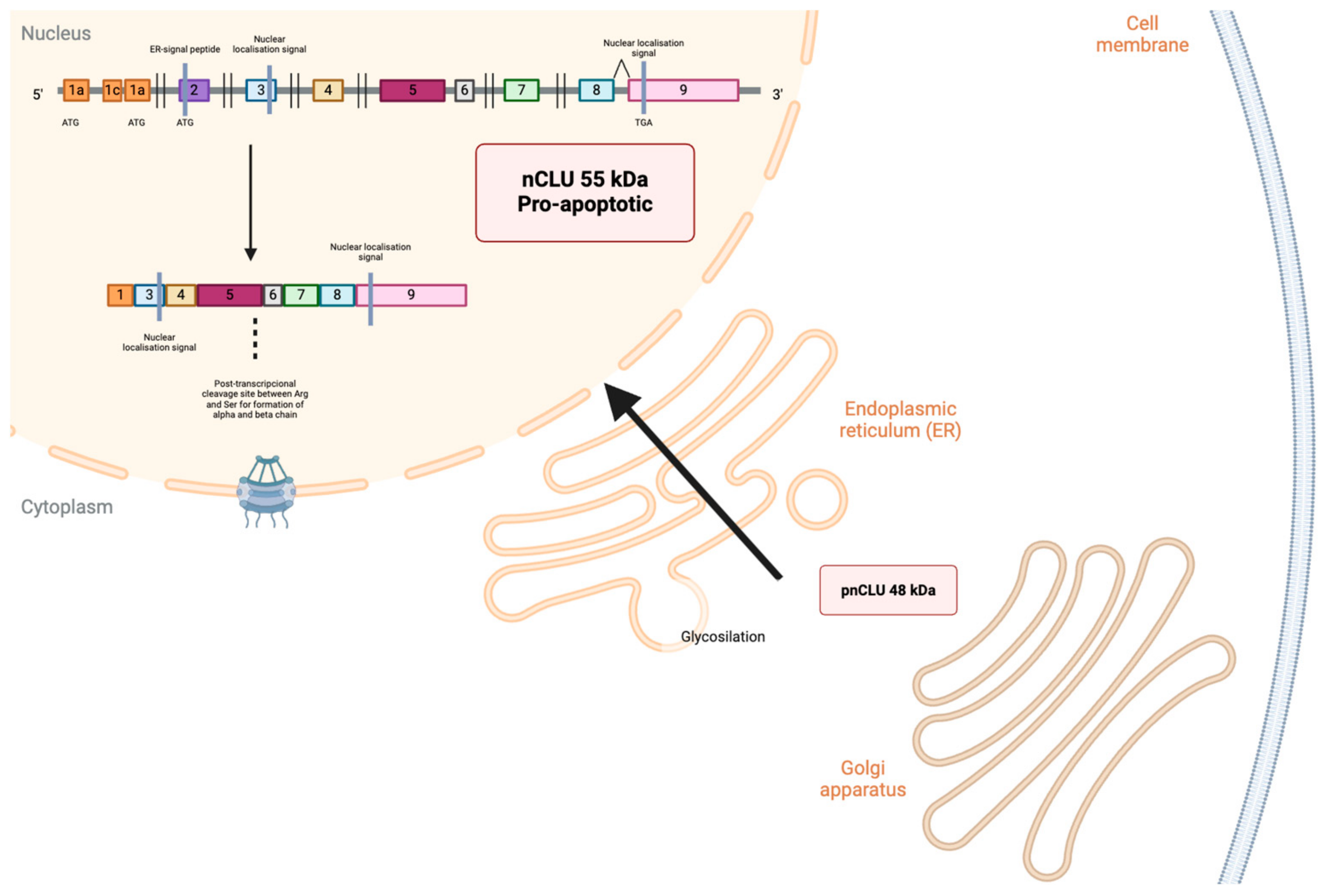
2. Isoforms and Regulation of CLU Expression Gene
3. Clusterin and Its Involvement in Cancer
3.1. Tumorigenesis
3.2. Cell Proliferation
3.3. Epithelial–Mesenchymal Transition and Metastasis
3.4. Chemoresistance and Chemosensitivity with Clusterin
4. Clusterin as a Biomarker and Therapeutic Target in Cancer
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hogg, S.D.; Embery, G. The isolation and partial characterization of a sulphated glycoprotein from human whole saliva which aggregates strains of Streptococcus sanguis but not Streptococcus mutans. Arch. Oral Biol. 1979, 24, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Jomary, C. Clusterin. Int. J. Biochem. Cell Biol. 2002, 34, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Trougakos, I.P.; Gonos, E.S. Regulation of clusterin/apolipoprotein J, a functional homologue to the small heat shock proteins, by oxidative stress in aging and age-related diseases. Free Radic. Res. 2006, 40, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Itahana, Y.; Piens, M.; Sumida, T.; Fong, S.; Muschler, J.; Desprez, P.Y. Regulation of clusterin expression in mammary epithelial cells. Exp. Cell Res. 2007, 313, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Trougakos, I.P.; Gonos, E.S. Clusterin/apolipoprotein J in human aging and cancer. Int. J. Biochem. Cell Biol. 2002, 34, 1430–1448. [Google Scholar] [CrossRef]
- Bettuzzi, S. Chapter 1: Introduction. Adv. Cancer Res. 2009, 104, 1–8. [Google Scholar] [CrossRef]
- Prochnow, H.; Gollan, R.; Rohne, P.; Hassemer, M.; Koch-Brandt, C.; Baiersdörfer, M. Non-secreted clusterin isoforms are translated in rare amounts from distinct human mRNA variants and do not affect Bax-mediated apoptosis or the NF-kappaB signaling pathway. PLoS ONE 2013, 8, e75303. [Google Scholar] [CrossRef]
- Athanas, K.M.; Mauney, S.L.; Woo, T.U. Increased extracellular clusterin in the prefrontal cortex in schizophrenia. Schizophr. Res. 2015, 169, 381–385. [Google Scholar] [CrossRef]
- Yu, B.; Yang, Y.; Liu, H.; Gong, M.; Millard, R.W.; Wang, Y.G.; Ashraf, M.; Xu, M. Clusterin/Akt Up-Regulation Is Critical for GATA-4 Mediated Cytoprotection of Mesenchymal Stem Cells against Ischemia Injury. PLoS ONE 2016, 11, e0151542. [Google Scholar] [CrossRef]
- Fandridis, E.; Apergis, G.; Korres, D.S.; Nikolopoulos, K.; Zoubos, A.B.; Papassideri, I.; Trougakos, I.P. Increased expression levels of apolipoprotein J/clusterin during primary osteoarthritis. In Vivo 2011, 25, 745–749. [Google Scholar]
- Suuronen, T.; Nuutinen, T.; Ryhänen, T.; Kaarniranta, K.; Salminen, A. Epigenetic regulation of clusterin/apolipoprotein J expression in retinal pigment epithelial cells. Biochem. Biophys. Res. Commun. 2007, 357, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Prikrylova Vranova, H.; Mareš, J.; Nevrlý, M.; Stejskal, D.; Zapletalová, J.; Hluštík, P.; Kaňovský, P. CSF markers of neurodegeneration in Parkinson’s disease. J. Neural Transm. 2010, 117, 1177–1181. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, G.; Beiser, A.S.; Preis, S.R.; Courchesne, P.; Chouraki, V.; Levy, D.; Seshadri, S. Plasma clusterin levels and risk of dementia, Alzheimer’s disease, and stroke. Alzheimers Dement. 2016, 3, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Schrijvers, E.M.; Koudstaal, P.J.; Hofman, A.; Breteler, M.M. Plasma clusterin and the risk of Alzheimer disease. JAMA 2011, 305, 1322–1326. [Google Scholar] [CrossRef]
- Chen, D.; Wang, Y.; Zhang, K.; Jiao, X.; Yan, B.; Liang, J. Antisense oligonucleotide against clusterin regulates human hepatocellular carcinoma invasion through transcriptional regulation of matrix metalloproteinase-2 and E-cadherin. Int. J. Mol. Sci. 2012, 13, 10594–10607. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Wang, Y.; Zhang, K.; Jiao, X.; Yan, B.; Liang, J. Regulation of chemosensitivity and migration by clusterin in non-small cell lung cancer cells. Cancer Chemother. Pharmacol. 2012, 69, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Lourda, M.; Trougakos, I.P.; Gonos, E.S. Development of resistance to chemotherapeutic drugs in human osteosarcoma cell lines largely depends on up-regulation of Clusterin/Apolipoprotein J. Int. J. Cancer 2007, 120, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, H.; Liu, Y.; Zhou, J.; He, W.; Quick, C.M.; Xie, D.; Smoller, B.R.; Fan, C.Y. Epigenetic and immunohistochemical characterization of the Clusterin gene in ovarian tumors. Arch. Gynecol. Obstet. 2013, 287, 989–995. [Google Scholar] [CrossRef]
- Tellez, T.; García-Aranda, M.; Redondo, M. The role of clusterin in carcinogenesis and its potential utility as a therapeutic target. Curr. Med. Chem. 2016, 23, 4297–4308. [Google Scholar] [CrossRef]
- Trougakos, I.P.; Pawelec, G.; Tzavelas, C.; Ntouroupi, T.; Gonos, E.S. Clusterin/Apolipoprotein J up-regulation after zinc exposure, replicative senescence, or differentiation of human hematopoietic cells. Biogerontology 2006, 7, 375–382. [Google Scholar] [CrossRef]
- Available online: http://www.ensembl.org/Homo_sapiens/Gene/Summary?g=ENSG00000120885,r=8:27596917-27615031 (accessed on 4 January 2024).
- Available online: http://vega.sanger.ac.uk/Homo_sapiens/Gene/Summary?g=OTTHUMG00000102114,r=8:27596917-27615031 (accessed on 4 January 2024).
- Bonacini, M.; Coletta, M.; Ramazzina, I.; Naponelli, V.; Modernelli, A.; Davalli, P.; Bettuzzi, S.; Rizzi, F. Distinct promoters, subjected to epigenetic regulation, drive the expression of two clusterin mRNAs in prostate cancer cells. Biochim. Biophys. Acta 2015, 1849, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, F.; Coletta, M.; Bettuzzi, S. Chapter 2: Clusterin (CLU): From one gene and two transcripts to many proteins. Adv. Cancer Res. 2009, 104, 9–23. [Google Scholar] [PubMed]
- Yu, J.T.; Tan, L. The role of clusterin in Alzheimer’s disease: Pathways, pathogenesis, and therapy. Mol. Neurobiol. 2012, 45, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Foster, E.M.; Dangla-Valls, A.; Lovestone, S.; Ribe, E.M.; Buckley, N.J. Clusterin in Alzheimer’s Disease: Mechanisms, Genetics, and Lessons from Other Pathologies. Front. Neurosci. 2019, 13, 164. [Google Scholar] [CrossRef] [PubMed]
- Rohne, P.; Prochnow, H.; Koch-Brandt, C. The CLU-files: Disentanglement of a mystery. Biomol. Concepts 2016, 7, 1–15. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, W.; Kundu, S.; Jang, T.L.; Yang, X.; Pins, M.; Smith, N.; Jovanovic, B.; Xin, D.; Liang, L.; et al. The leader sequence triggers and enhances several functions of clusterin and is instrumental in the progression of human prostate cancer in vivo and in vitro. BJU Int. 2006, 98, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Poon, S.; Treweek, T.M.; Wilson, M.R.; Easterbrook-Smith, S.B.; Carver, J.A. Clusterin is an extracellular chaperone that specifically interacts with slowly aggregating proteins on their off-folding pathway. FEBS Lett. 2002, 513, 259–266. [Google Scholar] [CrossRef]
- Bettuzzi, S.; Scorcioni, F.; Astancolle, S.; Davalli, P.; Scaltriti, M.; Corti, A. Clusterin (SGP-2) transient overexpression decreases proliferation rate of SV40-immortalized human prostate epithelial cells by slowing down cell cycle progression. Oncogene 2002, 21, 4328–4334. [Google Scholar] [CrossRef]
- Pucci, S.; Bonanno, E.; Pichiorri, F.; Angeloni, C.; Spagnoli, L.G. Modulation of different clusterin isoforms in human colon tumorigenesis. Oncogene 2004, 23, 2298–2304. [Google Scholar] [CrossRef]
- Caccamo, A.E.; Scaltriti, M.; Caporali, A.; D’Arca, D.; Corti, A.; Corvetta, D.; Sala, A.; Bettuzzi, S. Ca2+ depletion induces nuclear clusterin, a novel effector of apoptosis in immortalized human prostate cells. Cell Death Differ. 2005, 12, 101–104. [Google Scholar] [CrossRef]
- Caccamo, A.E.; Scaltriti, M.; Caporali, A.; D’Arca, D.; Scorcioni, F.; Astancolle, S.; Mangiola, M.; Bettuzzi, S. Cell detachment and apoptosis induction of immortalized human prostate epithelial cells are associated with early accumulation of a 45 kDa nuclear isoform of clusterin. Biochem. J. 2004, 382, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Leskov, K.S.; Klokov, D.Y.; Li, J.; Kinsella, T.J.; Boothman, D.A. Synthesis and functional analyses of nuclear clusterin, a cell death protein. J. Biol. Chem. 2003, 278, 11590–11600. [Google Scholar] [CrossRef]
- Li, N.; Zoubeidi, A.; Beraldi, E.; Gleave, M.E. GRP78 regulates clusterin stability, retrotranslocation and mitochondrial localization under ER stress in prostate cancer. Oncogene 2013, 32, 1933–1942. [Google Scholar] [CrossRef]
- Nizard, P.; Tetley, S.; Le Dréan, Y.; Watrin, T.; Le Goff, P.; Wilson, M.R.; Michel, D. Stress-Induced Retrotranslocation of Clusterin/ApoJ into the Cytosol. Traffic 2007, 8, 554–565. [Google Scholar] [CrossRef]
- Scaltriti, M.; Bettuzzi, S.; Sharrard, R.M.; Caporali, A.; Caccamo, A.E.; Maitland, N.J. Clusterin overexpression in both malignant and nonmalignant prostate epithelial cells induces cell cycle arrest and apoptosis. Br. J. Cancer 2004, 91, 1842–1850. [Google Scholar] [CrossRef]
- Scaltriti, M.; Santamaria, A.; Paciucci, R.; Bettuzzi, S. Intracellular clusterin induces G2-M phase arrest and cell death in PC-3 prostate cancer cells1. Cancer Res. 2004, 64, 6174–6182. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Mathis, K.W.; Lee, I.K. The physiological roles of apolipoprotein J/clusterin in metabolic and cardiovascular diseases. Rev. Endocr. Metab. Disord. 2014, 15, 45–53. [Google Scholar] [CrossRef]
- Zoubeidi, A.; Chi, K.; Gleave, M. Targeting the cytoprotective chaperone, clusterin, for treatment of advanced cancer. Clin. Cancer Res. 2010, 16, 1088–1093. [Google Scholar] [CrossRef]
- Goetz, E.M.; Shankar, B.; Zou, Y.; Morales, J.C.; Luo, X.; Araki, S.; Bachoo, R.; Mayo, L.D.; Boothman, D.A. ATM dependent IGF-1 induction regulates secretory clusterin expression after DNA damage and in genetic instability. Oncogene Nat. Publ. Group 2011, 30, 3745–3754. [Google Scholar] [CrossRef]
- Michel, D.; Chatelain, G.; North, S.; Brun, G. Stress-induced transcription of the clusterin/apoJ gene. Biochem. J. 1997, 50, 45–50. [Google Scholar] [CrossRef]
- Trougakos, I.P. The Molecular Chaperone Apolipoprotein J/Clusterin as a Sensor of Oxidative Stress: Implications in Therapeutic Approaches—A Mini-Review. Gerontology 2013, 59, 514–523. [Google Scholar] [CrossRef] [PubMed]
- July, L.V.; Akbari, M.; Zellweger, T.; Jones, E.C.; Goldenberg, S.L.; Gleave, M.E. Clusterin expression is significantly enhanced in prostate cancer cells following androgen withdrawal therapy. Prostate 2002, 50, 179–188. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, P.C.; Shlyakhtenko, A.; Mir, A.A.; Vinson, C. Clustering of DNA sequences in human promoters. Genome Res. 2004, 14, 1562–1574. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Choi, W.S. Proapoptotic role of nuclear clusterin in brain. Anat. Cell Biol. 2011, 44, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.; Taillefer, D.; Lakins, J.; Pineault, J.; Chader, G.; Tenniswood, M. Molecular characterization of human TRPM-2/clusterin, a gene associated with sperm maturation, apoptosis and neurodegeneration. Eur. J. Biochem. 1994, 221, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Deb, M.; Sengupta, D.; Rath, S.K.; Kar, S.; Parbin, S.; Shilpi, A.; Pradhan, N.; Bhutia, S.K.; Roy, S.; Patra, S.K. Clusterin gene is predominantly regulated by histone modifications in human colon cancer and ectopic expression of the nuclear isoform induces cell death. Biochim. Biophys. Acta 2015, 1852, 1630–1645. [Google Scholar] [CrossRef]
- Hellebrekers, D.M.; Melotte, V.; Viré, E.; Langenkamp, E.; Molema, G.; Fuks, F.; Herman, J.G.; Van Criekinge, W.; Griffioen, A.W.; van Engeland, M. Identification of epigenetically silenced genes in tumor endothelial cells. Cancer Res. 2007, 67, 4138–4148. [Google Scholar] [CrossRef]
- Serrano, A.; Redondo, M.; Tellez, T.; Castro-Vega, I.; Roldan, M.J.; Mendez, R.; Rueda, A.; Jiménez, E. Regulation of clusterin expression in human cancer via DNA methylation. Tumour Biol. 2009, 30, 286–291. [Google Scholar] [CrossRef]
- Handy, D.E.; Castro, R.; Loscalzo, J. Epigenetic modifications: Basic mechanisms and role in cardiovascular disease. Circulation 2011, 123, 2145–2156. [Google Scholar] [CrossRef]
- Liao, F.T.; Lee, Y.J.; Ko, J.L.; Tsai, C.C.; Tseng, C.J.; Sheu, G.T. Hepatitis delta virus epigenetically enhances clusterin expression via histone acetylation in human hepatocellular carcinoma cells. J. Gen. Virol. 2009, 90, 1124–1134. [Google Scholar] [CrossRef]
- Rauhala, H.E.; Porkka, K.P.; Saramäki, O.R.; Tammela, T.L.; Visakorpi, T. Clusterin is epigenetically regulated in prostate cancer. Int. J. Cancer 2008, 123, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Rosemblit, N.; Chen, C.L. Regulators for the rat clusterin gene: DNA methylation and cis-acting regulatory elements. J. Mol. Endocrinol. 1994, 13, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Mydlarz, W.; Uemura, M.; Ahn, S.; Hennessey, P.; Chang, S.; Demokan, S.; Sun, W.; Shao, C.; Bishop, J.; Krosting, J.; et al. Clusterin is a gene-specific target of microRNA-21 in head and neck squamous cell carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 868–877. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, Y.; Huang, Z.; Li, D.; Chen, X.; Cao, M.; Meng, Q.; Pang, H.; Sun, L.; Zhao, Y.; et al. miRNA-378 reverses chemoresistance to cisplatin in lung adenocarcinoma cells by targeting secreted clusterin. Sci. Rep. 2016, 6, 19455. [Google Scholar] [CrossRef]
- Buttyan, R.; Olsson, C.A.; Pintar, J.; Chang, C.; Bandyk, M.; Ng, P.Y.; Sawczuk, I.S. Induction of the TRPM-2 gene in cells undergoing programmed death. Mol. Cell Biol. 1989, 9, 3473–3481. [Google Scholar] [CrossRef]
- Leger, J.G.; Montpetit, M.L.; Tenniswood, M.P. Characterization and cloning of androgen-repressed mRNAs from rat ventral prostate. Biochem. Biophys. Res. Commun. 1987, 147, 196–203. [Google Scholar] [CrossRef]
- Leger, J.G.; Le Guellec, R.; Tenniswood, M.P. Treatment with antiandrogens induces an androgen-repressed gene in the rat ventral prostate. Prostate 1988, 13, 131–142. [Google Scholar] [CrossRef]
- Sensibar, J.A.; Sutkowski, D.M.; Raffo, A.; Buttyan, R.; Griswold, M.D.; Sylvester, S.R.; Kozlowski, J.M.; Lee, C. Prevention of cell death induced by tumor necrosis factor alpha in LNCaP cells by overexpression of sulfated glycoprotein-2 (clusterin). Cancer Res. 1995, 55, 2431–2437. [Google Scholar]
- Schwochau, G.B.; Nath, K.A.; Rosenberg, M.E. Clusterin protects against oxidative stress in vitro through aggregative and nonaggregative properties. Kidney Int. 1998, 53, 1647–1653. [Google Scholar] [CrossRef]
- Yang, C.R.; Leskov, K.; Hosley-Eberlein, K.; Criswell, T.; Pink, J.J.; Kinsella, T.J.; Boothman, D.A. Nuclear Clusterin/XIP8, an x-ray-induced Ku70-binding protein that signals cell death. Proc. Natl. Acad. Sci. USA 2000, 97, 5907–5912. [Google Scholar] [CrossRef] [PubMed]
- Leskov, K.S.; Araki, S.; Lavik, J.P.; Gomez, J.A.; Gama, V.; Gonos, E.S.; Trougakos, I.P.; Matsuyama, S.; Boothman, D.A. CRM1 protein-mediated regulation of nuclear clusterin (nCLU), an ionizing radiation-stimulated, Bax-dependent pro-death factor. J. Biol. Chem. 2011, 286, 40083–40090. [Google Scholar] [CrossRef]
- Essabbani, A.; Garcia, L.; Zonetti, M.J.; Fisco, T.; Pucci, S.; Chiocchia, G. Exon-skipping strategy by ratio modulation between cytoprotective versus pro-apoptotic clusterin forms increased sensitivity of LNCaP to cell death. PLoS ONE 2013, 8, e54920. [Google Scholar] [CrossRef] [PubMed]
- García-Aranda, M.; Serrano, A.; Redondo, M. Regulation of Clusterin Gene Expression. Curr. Protein Pept. Sci. 2017, 19, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Yoo, J.C.; Han, J.Y.; Hwang, E.M.; Kim, Y.S.; Jeong, E.Y.; Sun, C.H.; Yi, G.S.; Roh, G.S.; Kim, H.J.; et al. Human nuclear clusterin mediates apoptosis by interacting with Bcl-XL through C-terminal coiled coil domain. J. Cell Physiol. 2012, 227, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Shannan, B.; Seifert, M.; Leskov, K.; Willis, J.; Boothman, D.; Tilgen, W.; Reichrath, J. Challenge and promise: Roles for clusterin in pathogenesis, progression and therapy of cancer. Cell Death Differ. 2006, 13, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lv, X.; Chen, L.; Liu, Y. The role and function of CLU in cancer biology and therapy. Clin. Exp. Med. 2023, 23, 1375–1391. [Google Scholar] [CrossRef]
- Trougakos, I.P.; Lourda, M.; Antonelou, M.H.; Kletsas, D.; Gorgoulis, V.G.; Papassideri, I.S.; Zou, Y.; Margaritis, L.H.; Boothman, D.A.; Gonos, E.S. Intracellular clusterin inhibits mitochondrial apoptosis by suppressing stress signals activating p53 and stabilizing the cytosolic Ku70-Bax protein complex. Clin. Cancer Res. 2009, 15, 48–59. [Google Scholar] [CrossRef]
- Blume, A.J.; Foster, C.J. Adenylate cyclase of mouse neuroblastoma cells: Regulation by 2-chloroadenosine, prostaglandin E1, and the cations Mg2+, Ca2+, and Mn2+. J. Neurochem. 1976, 26, 305–311. [Google Scholar] [CrossRef]
- Nicholson, G.; Lawrence, A.; Ala, F.A.; Bird, G.W. Semi-quantitative assay of antigen site density by flow cytometry analysis. Transfus. Med. 1991, 1, 87–90. [Google Scholar] [CrossRef]
- Dews, M.; Fox, J.L.; Hultine, S.; Sundaram, P.; Wang, W.; Liu, Y.Y.; Furth, E.; Enders, G.H.; El-Deiry, W.; Schelter, J.M.; et al. The myc-miR-17~92 axis blunts TGF{beta} signaling and production of multiple TGF{beta}-dependent antiangiogenic factors. Cancer Res. 2010, 70, 8233–8246. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Wang, L.; Li, M.; Wang, X.; Zhang, L. Secreted clusterin (sCLU) regulates cell proliferation and chemosensitivity to cisplatin by modulating ERK1/2 signals in human osteosarcoma cells. World J. Surg. Oncol. 2014, 12, 255. [Google Scholar] [CrossRef]
- Wang, X.; Xie, J.; Lu, X.; Li, H.; Wen, C.; Huo, Z.; Xie, J.; Shi, M.; Tang, X.; Chen, H.; et al. Melittin inhibits tumor growth and decreases resistance to gemcitabine by downregulating cholesterol pathway gene CLU in pancreatic ductal adenocarcinoma. Cancer Lett. 2017, 399, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chesnokova, V.; Zonis, S.; Wawrowsky, K.; Tani, Y.; Ben-Shlomo, A.; Ljubimov, V.; Mamelak, A.; Bannykh, S.; Melmed, S. Clusterin and FOXL2 act concordantly to regulate pituitary gonadotroph adenoma growth. Mol. Endocrinol. 2012, 26, 2092–2103. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Peng, M.; Zhou, S.; Xiao, D.; Hu, X.; Xu, S.; Wu, J.; Yang, X. Metformin targets Clusterin to control lipogenesis and inhibit the growth of bladder cancer cells through SREBP-1c/FASN axis. Signal Transduct. Target. Ther. 2021, 6, 98. [Google Scholar] [CrossRef] [PubMed]
- Whitesell, L.; Lindquist, S.L. HSP90 and the chaperoning of cancer. Nat. Rev. Cancer 2005, 5, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Shim, Y.-J.; Kang, B.-H.; Jeon, H.-S.; Park, I.-S.; Lee, K.-U.; Lee, I.-K.; Park, G.-H.; Lee, K.-M.; Schedin, P.; Min, B.-H. Clusterin induces matrix metalloproteinase-9 expression via ERK1/2 and PI3K/Akt/NF-κB pathways in monocytes/macrophages. J. Leukoc. Biol. 2011, 90, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lu, J.; Zhou, S.; Wang, W.; Tan, G.; Zhang, Z.; Dong, Z.; Kang, T.; Tang, F. N, N′-dinitrosopiperazine-induced clusterin participates in nasopharyngeal carcinoma metastasis. Oncotarget 2016, 7, 5548–5563. [Google Scholar] [CrossRef] [PubMed]
- Bailes, J.; Soloviev, M. Insulin-Like Growth Factor-1 (IGF-1) and Its Monitoring in Medical Diagnostic and in Sports. Biomolecules 2021, 11, 217. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, C.; Chen, S.; Liu, J.; Fu, Y.; Luo, Y. Extracellular Hsp90alpha and clusterin synergistically promote breast cancer epithelial-to-mesenchymal transition and metastasis through LRP1. J. Cell Sci. 2019, 132, jcs228213. [Google Scholar] [CrossRef]
- Viard, I.; Wehrli, P.; Jornot, L.; Bullani, R.; Vechietti, J.-L.; French, L.E.; Schifferli, J.A.; Tschopp, J. Clusterin gene expression mediates resistance to apoptotic cell death induced by heat shock and oxidative stress. J. Investg. Dermatol. 1999, 112, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Cervellera, M.; Raschella, G.; Santilli, G.; Tanno, B.; Ventura, A.; Mancini, C.; Sevignani, C.; Calabretta, B.; Sala, A. Direct transactivation of the antiapoptotic gene apolipoprotein J (Clusterin) by B-MYB. J. Biol. Chem. 2000, 275, 21055–21060. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Bai, Y. IGF-1 activates the P13K/AKT signaling pathway by positively regulating secretory clusterin. Mol. Med. Rep. 2012, 6, 1433–1437. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, L.; Whyte, L.; Chatterjee, N.; Tenniswood, M. Effects of clusterin overexpression on metastatic progression and therapy in breast cancer. BMC Cancer 2010, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Meunier, A.; Cornet, F.; Campos, M. Bacterial cell proliferation: From molecules to cells. FEMS Microbiol. Rev. 2020, 45, fuaa046. [Google Scholar] [CrossRef]
- Wang, Z. Regulation of cell cycle progression by growth factor-induced cell signaling. Cells 2021, 10, 33270. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Shiraga, K.; Sato, S.; Punfa, W.; Naiki-Ito, A.; Yamashita, Y.; Shirai, T.; Takahashi, S. Apocynin, an NADPH oxidase inhibitor, suppresses rat prostate carcinogenesis. Cancer Sci. 2013, 104, 1711–1717. [Google Scholar] [CrossRef]
- Chen, Z.; Fan, Z.; Dou, X.; Zhou, Q.; Zeng, G.; Liu, L.; Chen, W.; Lan, R.; Liu, W.; Ru, G.; et al. Inactivation of tumor suppressor gene Clusterin leads to hyperactivation of TAK1-NF-kappaB signaling axis in lung cancer cells and denotes a therapeutic opportunity. Theranostics 2020, 10, 11520–11534. [Google Scholar] [CrossRef]
- Chun, Y.J. Knockdown of clusterin expression increases the in vitro sensitivity of human prostate cancer cells to paclitaxel. J. Toxicol. Environ. Health A 2014, 77, 1443–1450. [Google Scholar] [CrossRef]
- Zheng, W.; Yao, M.; Qian, Q.; Sai, W.; Qiu, L.; Yang, J.; Wu, W.; Dong, Z.; Yao, D. Oncogenic secretory clusterin in hepatocellular carcinoma: Expression at early staging and emerging molecular target. Oncotarget 2016, 8, 52321–52332. [Google Scholar] [CrossRef]
- Takeuchi, A.; Shiota, M.; Beraldi, E.; Thaper, D.; Takahara, K.; Ibuki, N.; Pollak, M.; Cox, M.E.; Naito, S.; Gleave, M.E.; et al. Insulin-like growth factor-I induces CLU expression through Twist1 to promote prostate cancer growth. Mol Cell Endocrinol. 2014, 384, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Men, C.; Xu, Y.; Zhao, K.; Luo, L.; Dong, D.; Yu, Q. Clusterin promotes growth and invasion of clear cell renal carcinoma cell by upregulation of S100A4 expression. Cancer Biomark. 2018, 21, 915–923. [Google Scholar] [CrossRef]
- Pajak, B.; Kania, E.; Gajkowska, B.; Orzechowski, A. Lipid rafts mediate epigallocatechin-3-gallate- and green tea extract-dependent viability of human colon adenocarcinoma COLO 205 cells; clusterin affects lipid rafts-associated signaling pathways. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2011, 62, 449–459. [Google Scholar]
- Tang, Z.; Wang, W.; Liu, Z.; Sun, X.; Liao, Z.; Chen, F.; Jiang, G.; Huo, G. Blocking ERK signaling pathway reduces MMP-9 expression to alleviate cerebral edema after traumatic brain injury in rats. J. South. Med. Univ. 2020, 40, 1018–1022. [Google Scholar]
- Yang, P.; Yang, Z.; Dong, Y.; Yang, L.; Peng, S.; Yuan, L.; Hu, X.; Chen, S.; Tang, H.; Yang, X.; et al. Clusterin is a prognostic biomarker of breast cancer and correlates with the immune microenvironment. Transl. Cancer Res. 2023, 12, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Tang, W.; Azizian, A.; Gaedcke, J.; Strobel, P.; Wang, L.; Cawley, H.; Ohara, Y.; Valenzuela, P.; Zhang, L.; et al. Dysregulation of the HNF1B/Clusterin axis enhances disease progression in a highly aggressive subset of pancreatic cancer patients. Carcinogenesis 2022, 43, 1198–1210. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. Different mechanisms of resistance to cancer drugs: A brief review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Zhang, H.; Kim, J.K.; Edwards, C.A.; Xu, Z.; Taichman, R.; Wang, C.-Y. Clusterin inhibits apoptosis by interacting with activated Bax. Nat. Cell Biol. 2005, 7, 909–915. [Google Scholar] [CrossRef]
- Tang, M.; Li, J.; Liu, B.; Song, N.; Wang, Z.; Yin, C. Clusterin expression in human testicular seminoma. Med. Hypotheses 2013, 81, 635–637. [Google Scholar] [CrossRef]
- Wang, X.; Zou, F.; Zhong, J.; Yue, L.; Wang, F.; Wei, H.; Yang, G.; Jin, T.; Dong, X.; Li, J.; et al. Secretory clusterin mediates resistance to oxaliplatin through the Gadd45a/PI3K/Akt signaling pathway in hepatocellular carcinoma. J. Cancer 2018, 9, 1403–1413. [Google Scholar] [CrossRef]
- Panico, F.; Rizzi, F.; Fabbri, L.M.; Bettuzzi, S.; Luppi, F. Clusterin (CLU) and lung cancer. Adv. Cancer Res. 2009, 105, 63–76. [Google Scholar]
- July, L.V.; Beraldi, E.; So, A.; Fazli, L.; Evans, K.; English, J.C.; Gleave, M.E. Nucleotide-based therapies targeting clusterin chemosensitize human lung adenocarcinoma cells both in vitro and in vivo. Mol. Cancer Ther. 2004, 3, 223–232. [Google Scholar] [CrossRef]
- Bi, J.; Guo, A.L.; Lai, Y.R.; Li, B.; Zhong, J.M.; Wu, H.Q.; Xie, Z.; He, Y.L.; Lv, Z.L.; Lau, S.H.; et al. Overexpression of clusterin correlates with tumor progression, metastasis in gastric cancer: A study on tissue microarrays. Neoplasm 2010, 57, 191. [Google Scholar] [CrossRef]
- Xie, D.; Lau, S.H.; Sham, J.S.; Wu, Q.L.; Fang, Y.; Liang, L.Z.; Che, L.H.; Zeng, Y.X.; Guan, X.Y. Up-regulated expression of cytoplasmic clusterin in human ovarian carcinoma. Cancer 2005, 103, 277–283. [Google Scholar] [CrossRef]
- Yang, G.F.; Li, X.M.; Xie, D. Overexpression of clusterin in ovarian cancer is correlated with impaired survival. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc. 2009, 19, 1342–1346. [Google Scholar] [CrossRef] [PubMed]
- Al-Maghrabi, J.A.; Butt, N.S.; Anfinan, N.; Sait, K.; Sait, H.; Bajouh, O.; Khabaz, M.N. Clusterin immunoexpression is associated with early stage endometrial carcinomas. Acta Histochemical. 2016, 118, 430–434. [Google Scholar] [CrossRef]
- Fuzio, P.; Valletti, A.; Napoli, A.; Napoli, G.; Cormio, G.; Selvaggi, L.; Liuni, S.; Pesole, G.; Maiorano, E.; Perlino, E. Regulation of the expression of CLU isoforms in endometrial proliferative diseases. Int. J. Oncol. 2013, 42, 1929–1944. [Google Scholar] [CrossRef]
- Won, Y.S.; Lee, S.J.; Yeo, S.G.; Park, D.C. Effects of female sex hormones on clusterin expression and paclitaxel resistance in endometrial cancer cell lines. Int. J. Med. Sci. 2012, 9, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Park, D.C.; Yeo, S.G.; Shin, E.Y.; Mok, S.C.; Kim, D.H. Clusterin confers paclitaxel resistance in cervical cancer. Gynecol. Oncol. 2006, 103, 996–1000. [Google Scholar] [CrossRef]
- Niu, Z.; Li, X.; Hu, B.; Li, R.; Wang, L.; Wu, L.; Wang, X. Small interfering RNA targeted to secretory clusterin blocks tumor growth, motility, and invasion in breast cancer. Acta Biochim. Biophys. Sin. 2012, 44, 991–998. [Google Scholar] [CrossRef]
- Li, J.; Jia, L.; Zhao, P.; Jiang, Y.; Zhong, S.; Chen, D. Stable knockdown of clusterin by vectorbased RNA interference in a human breast cancer cell line inhibits tumour cell invasion and metastasis. J. Int. Med. Res. 2012, 40, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Redondo, M.; Villar, E.; Torres-Muñoz, J.; Tellez, T.; Morell, M.; Petito, C.K. Overexpression of clusterin in human breast carcinoma. Am. J. Pathol. 2000, 157, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Sun, B.; Zhao, X.; Cui, Y.; Xu, S.; Dong, X.; Zhao, J.; Meng, J.; Jia, X.; Chi, J. Secreted CLU is associated with the initiation of triple-negative breast cancer. Cancer Biol. Ther. 2012, 13, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.H.; Wang, Y.; Chun, B.; Li, C.X.; Wu, L. Secretory clusterin (sCLU) overexpression is associated with resistance to preoperative neoadjuvant chemotherapy in primary breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 1337–1344. [Google Scholar] [PubMed]
- Redondo, M.; Tellez, T.; Roldan, M.J. The role of clusterin (CLU) in malignant transformation and drug resistance in breast carcinomas. Adv. Cancer Res. 2009, 105, 21–43. [Google Scholar] [PubMed]
- Mazzarelli, P.; Pucci, S.; Spagnoli, L.G. CLU and colon cancer. The dual face of CLU: From the normal to the malignant phenotype. Adv. Cancer Res. 2009, 105, 45–61. [Google Scholar] [PubMed]
- Redondo, M.; Rodrigo, I.; Alcaide, J.; Tellez, T.; Roldan, M.J.; Funez, R.; Diaz-Martin, A.; Rueda, A.; Jiménez, E. Clusterin expression is associated with decreased disease-free survival of patients with colorectal carcinomas. Histopathology 2010, 56, 932–936. [Google Scholar] [CrossRef]
- Sun, B.; Moibi, J.A.; Mak, A.; Xiao, Z.; Roa, W.; Moore, R.B. Response of bladder carcinoma cells to TRAIL and antisense oligonucleotide, Bcl-2 or clusterin treatments. J. Urol. 2009, 181, 1361–1371. [Google Scholar] [CrossRef]
- Miyake, H.; Gleave, M.; Kamidono, S.; Hara, I. Overexpression of clusterin in transitional cell carcinoma of the bladder is related to disease progression and recurrence. Urology 2002, 59, 150–154. [Google Scholar] [CrossRef]
- Hazzaa, S.M.; Elashry, O.M.; Afifi, I.K. Clusterin as a diagnostic and prognostic marker for transitional cell carcinoma of the bladder. Pathol. Oncol. Res. POR 2010, 16, 101–109. [Google Scholar] [CrossRef]
- Lau, S.H.; Sham, J.S.; Xie, D.; Tzang, C.H.; Tang, D.; Ma, N.; Hu, L.; Wang, Y.; Wen, J.M.; Xiao, G.; et al. Clusterin plays an important role in hepatocellular carcinoma metastasis. Oncogene 2006, 25, 1242–1250. [Google Scholar] [CrossRef]
- Xiu, P.; Dong, X.; Dong, X.; Xu, Z.; Zhu, H.; Liu, F.; Wei, Z.; Zhai, B.; Kanwar, J.R.; Jiang, H.; et al. Secretory clusterin contributes to oxaliplatin resistance by activating Akt pathway in hepatocellular carcinoma. Cancer Sci. 2013, 104, 375–382. [Google Scholar] [CrossRef]
- Patarat, R.; Riku, S.; Kunadirek, P.; Chuaypen, N.; Tangkijvanich, P.; Mutirangura, A.; Puttipanyalears, C. The expression of FLNA and CLU in PBMCs as a novel screening marker for hepatocellular carcinoma. Sci. Rep. 2021, 11, 14838. [Google Scholar] [CrossRef]
- Gao, G.; Luan, X. Diagnostic performance of clusterin in hepatocellular carcinoma: A meta-analysis. Int. J. Biol. Markers 2022, 37, 404–411. [Google Scholar] [CrossRef]
- Rasmy, H.S.; Mohammed, H.A.; Mohammed, E.S.; Ahmed, A.S.M.; Isaac, A. Serum clusterin as a promising diagnostic and prognostic marker for hepatocellular carcinoma after locoregional treatment. Egypt. J. Immunol. 2022, 29, 26–40. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Chen, Y.H.; Tan, Q.Q.; Liu, X.B.; Tan, C. Clusterin is upregulated by erastin, a ferroptosis inducer and exerts cytoprotective effects in pancreatic adenocarcinoma cells. Anti-Cancer Drugs 2023, 35, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.J.; Motoo, Y.; Su, S.B.; Mouri, H.; Ohtsubo, K.; Matsubara, F.; Sawabu, N. Expression of clusterin in human pancreatic cancer. Pancreas 2002, 25, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Hoeller, C.; Pratscher, B.; Thallinger, C.; Winter, D.; Fink, D.; Kovacic, B.; Sexl, V.; Wacheck, V.; Gleave, M.E.; Pehamberger, H.; et al. Clusterin regulates drug-resistance in melanoma cells. J. Investig. Dermatol. 2005, 124, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- He, L.R.; Liu, M.Z.; Li, B.K.; Rao, H.L.; Liao, Y.J.; Zhang, L.J.; Guan, X.Y.; Zeng, Y.X.; Xie, D. Clusterin as a predictor for chemoradiotherapy sensitivity and patient survival in esophageal squamous cell carcinoma. Cancer Sci. 2009, 100, 2354–2360. [Google Scholar] [CrossRef]
- Bijian, K.; Mlynarek, A.M.; Balys, R.L.; Jie, S.; Xu, Y.; Hier, M.P.; Black, M.J.; Di Falco, M.R.; LaBoissiere, S.; Alaoui-Jamali, M.A. Serum proteomic approach for the identification of serum biomarkers contributed by oral squamous cell carcinoma and host tissue microenvironment. J. Proteome Res. 2009, 8, 2173–2185. [Google Scholar] [CrossRef] [PubMed]
- Redondo, M.; Fùnez, R.; Esteban, F. Apoptosis in the Development and Treatment of Laryngeal Cancer: Role of p53, Bcl-2 and Clusterin. In Apoptosis in Carcinogenesis and Chemotherapy; Chen, G.G., Lai, P.B., Eds.; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Wellmann, A.; Thieblemont, C.; Pittaluga, S.; Sakai, A.; Jaffe, E.S.; Siebert, P.; Raffeld, M. Detection of differentially expressed genes in lymphomas using cDNA arrays: Identification of clusterin as a new diagnostic marker for anaplastic large-cell lymphomas. Blood 2000, 96, 398–404. [Google Scholar] [CrossRef]
- Ma, J.; Gao, W.; Gao, J. sCLU as prognostic biomarker and therapeutic target in osteosarcoma. Bioengineered 2019, 10, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, F.; Bettuzzi, S. Clusterin (CLU) and prostate cancer. Adv. Cancer Res. 2009, 105, 1–19. [Google Scholar] [PubMed]
- Téllez, T.; Martin-García, D.; Redondo, M.; García-Aranda, M. Clusterin Expression in Colorectal Carcinomas. Int. J. Mol. Sci. 2023, 24, 14641. [Google Scholar] [CrossRef] [PubMed]
- Miyake, H.; Gleave, M.E.; Arakawa, S.; Kamidono, S.; Hara, I. Introducing the clusterin gene into human renal cell carcinoma cells enhances their metastatic potential. J. Urol. 2002, 167, 2203–2208. [Google Scholar] [CrossRef]
- Shi, H.; Deng, J.H.; Wang, Z.; Cao, K.Y.; Zhou, L.; Wan, H. Knockdown of clusterin inhibits the growth and migration of renal carcinoma cells and leads to differential gene expression. Mol. Med. Rep. 2013, 8, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Beheshti Namdar, A.; Kabiri, M.; Mosanan Mozaffari, H.; Aminifar, E.; Mehrad-Majd, H. Circulating clusterin levels and cancer risk: A systematic review and meta-analysis. Cancer Control 2022, 29. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.; Miyake, H.; Gleave, M.; Fujisawa, M. Effect of targeting clusterin using OGX-011 on antitumor activity of temsirolimus in a human renal cell carcinoma model. Target Oncol. 2016, 12, 69–79. [Google Scholar] [CrossRef]
- Yamanaka, K.; Gleave, M.; Muramaki, M.; Hara, I.; Miyake, H. Enhanced radiosensitivity by inhibition of the anti-apoptotic gene clusterin using antisense oligodeoxynucleotide in a human bladder cancer model. Oncol. Rep. 2005, 13, 885. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Z.; Zhang, K.; Liu, X.; Cao, W.; Zhang, L.; Zhang, S.; Yan, B.; Wang, Y.; Xia, C. Clusterin confers gemcitabine resistance in pancreatic cancer. World J. Surgical. Oncol. 2011, 9, 59. [Google Scholar] [CrossRef]
- Cao, C.; Shinohara, E.T.; Li, H.; Niermann, K.J.; Kim, K.W.; Sekhar, K.R.; Gleave, M.; Freeman, M.; Lu, B. Clusterin as a therapeutic target for radiation sensitization in a lung cancer model. Int. J. Radiat. Oncol. Biol. Phys. 2005, 63, 1228–1236. [Google Scholar] [CrossRef]
- Zellweger, T.; Chi, K.; Miyake, H.; Adomat, H.; Kiyama, S.; Skov, K.; Gleave, M.E. Enhanced radiation sensitivity in prostate cancer by inhibition of the cell survival protein clusterin. Clin Cancer Res. 2002, 8, 3276–3284. [Google Scholar] [PubMed]
- So, A.; Chi, K.; Miyake, H.; Adomat, H.; Kiyama, S.; Skov, K.; Gleave, M.E. Knockdown of the cytoprotective chaperone, clusterin, chemosensitizes human breast cancer cells both in vitro and in vivo. Mol. Cancer Ther. 2005, 4, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Zwain, I.; Amato, P. Clusterin protects granulosa cells from apoptotic cell death during follicular atresia. Exp. Cell Res. 2000, 257, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Kususda, Y.; Miyake, H.; Gleave, M.E.; Fujisawa, M. Clusterin inhibition using OGX-011 synergistically enhances antitumour activity of sorafenib in a human renal cell carcinoma model. Br. J. Cancer 2012, 106, 1945–1952. [Google Scholar] [CrossRef]
- Hassan, M.K.; Watari, H.; Han, Y.; Mitamura, T.; Hosaka, M.; Wang, L.; Tanaka, S.; Sakuragi, N. Clusterin is a potential molecular predictor for ovarian cancer patient’s survival: Targeting clusterin improves response to paclitaxel. J. Exp. Clin. Cancer Res. CR 2011, 30, 113. [Google Scholar] [CrossRef]
- Blumenstein, B.; Saad, F.; Hotte, S.; Chi, K.N.; Eigl, B.; Gleave, M.; Jacobs, C. Reduction in serum clusterin is a potential therapeutic biomarker in patients with castration-resistant prostate cancer treated with custirsen. Cancer Med. 2013, 2, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Higano, C.S. Potential use of custirsen to treat prostate cancer. OncoTargets Ther. 2013, 6, 785–797. [Google Scholar] [CrossRef]
- Chi, K.N.; Hotte, S.J.; Yu, E.Y.; Tu, D.; Eigl, B.J.; Tannock, I.; Saad, F.; North, S.; Powers, J.; Gleave, M.E.; et al. Randomized phase II study of docetaxel and prednisone with or without OGX-011 in patients with metastatic castration-resistant prostate cancer. J. Clin. Oncol. 2010, 28, 4247–4254. [Google Scholar] [CrossRef]
- Chia, S.; Dent, S.; Ellard, S.; Ellis, P.M.; Vandenberg, T.; Gelmon, K.; Powers, J.; Walsh, W.; Seymour, L.; Eisenhauer, E.A. Phase II trial of OGX-011 in combination with docetaxel in metastatic breast cancer. Clin. Cancer Res. 2009, 15, 708–713. [Google Scholar] [CrossRef]
- Laskin, J.J.; Nicholas, G.; Lee, C.; Gitlitz, B.; Vincent, M.; Cormier, Y.; Stephenson, J.; Ung, Y.; Sanborn, R.; Pressnail, B.; et al. Phase I/II trial of custirsen (OGX-011), an inhibitor of clusterin, in combination with a gemcitabine and platinum regimen in patients with previously untreated advanced non-small cell lung cancer. J. Thorac. Oncol. 2012, 7, 579–586. [Google Scholar] [CrossRef]
- Ma, X.; Zou, L.; Li, X.; Chen, Z.; Lin, Q.; Wu, X. MicroRNA-195 regulates docetaxel resistance by targeting clusterin in prostate cancer. Biomed. Pharmacother. 2018, 99, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, X.; Jiang, Y.; Jia, X.; Guo, Y. miR-217-5p inhibits invasion and metastasis of prostate cancer by targeting clusterin. Mamm Genome. 2021, 32, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Davalli, P.; Rizzi, F.; Caldara, G.F.; Davoli, S.; Corti, A.; Silva, A.; Astancolle, S.; Vitale, M.; Bettuzzi, S.; Arcari, M.; et al. Chronic administration of green tea extract to TRAMP mice induces the collapse of Golgi apparatus in prostate secretory cells and results in alterations of protein post-translational processing. Int. J. Oncol. 2011, 39, 1521–1527. [Google Scholar] [PubMed]
- Zhu, Y.; Chen, P.; Gao, Y.; Ta, N.; Zhang, Y.; Cai, J.; Zhao, Y.; Liu, S.; Zheng, J. MEG3 activated by Vitamin D inhibits colorectal cancer cells proliferation and migration via regulating clusterin. EBioMedicine 2018, 30, 148–157. [Google Scholar] [CrossRef]
- Xiong, J.; Wang, S.; Chen, T.; Shu, X.; Mo, X.; Chang, G.; Chen, J.J.; Li, C.; Luo, H.; Lee, J.D. Verteporfin blocks Clusterin which is required for survival of gastric cancer stem cell by modulating HSP90 function. Int. J. Biol. Sci. 2019, 15, 312–324. [Google Scholar] [CrossRef]
- Available online: https://clinicaltrials.gov/study/NCT02412462?intr=clusterin&page=2&rank=11 (accessed on 10 January 2024).

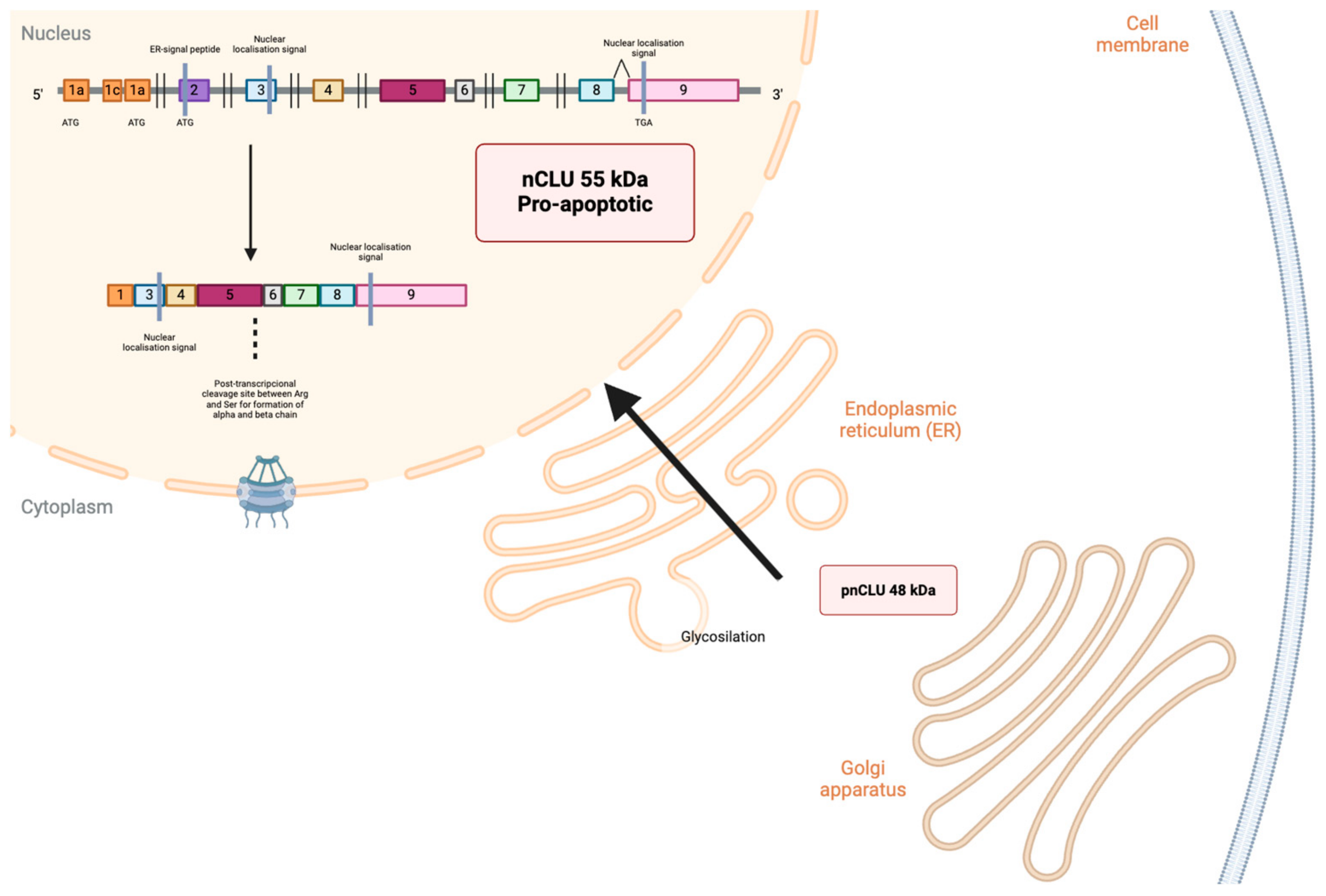

{kind=link}
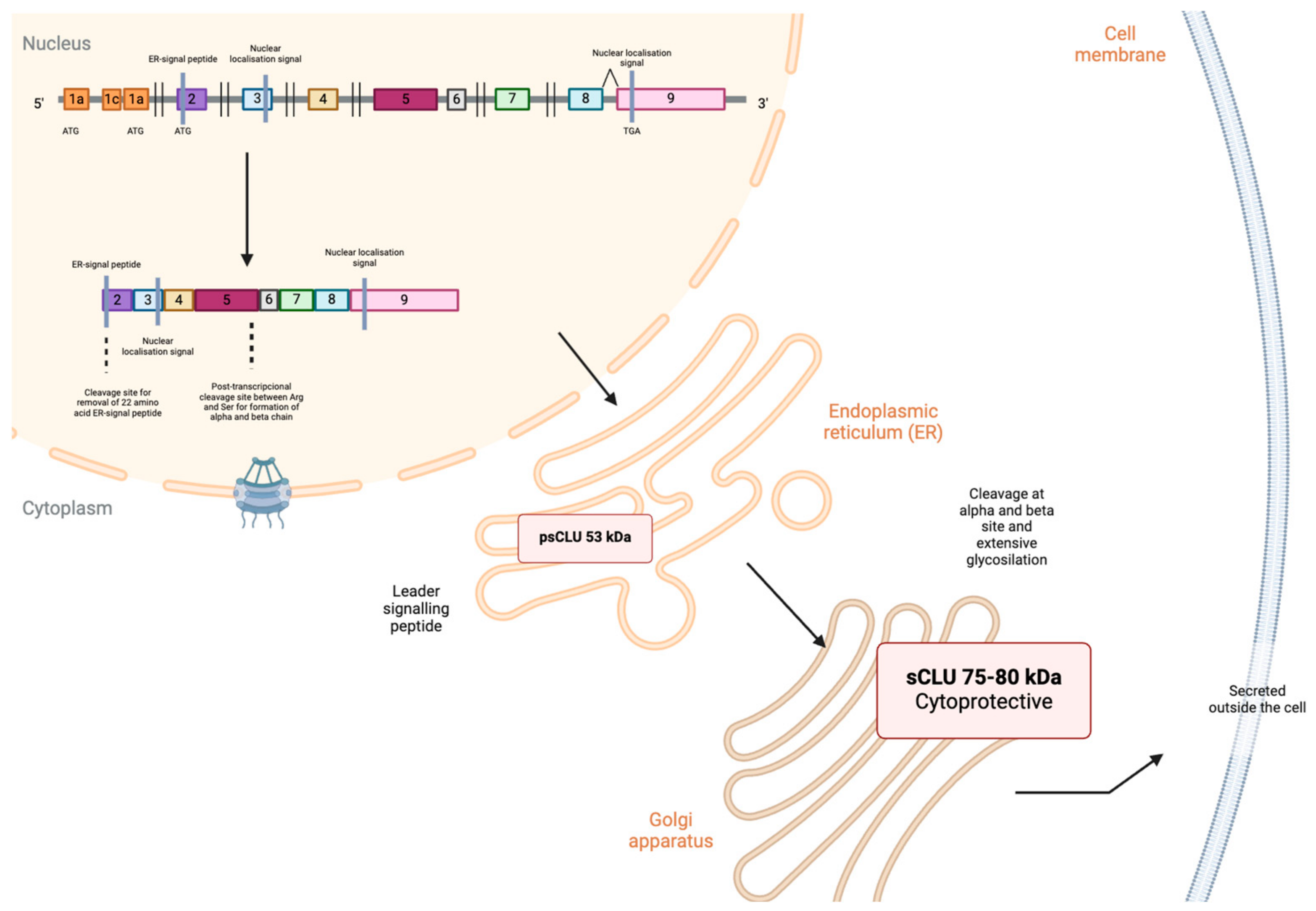{kind=link}
{kind=link}
| Biological Processes | nCLU | sCLU |
|---|---|---|
| Tumorigenesis | The proapoptotic isoform nCLU of CLU induces apoptosis in breast and prostate cancer, through specific interactions with proteins like Ku70 and Bcl-XL [70,71]. | sCLU also plays a crucial role by interacting with protein complexes like Ku70-bax, acting as a Bax retention factor in the cytosol, inhibiting its proapoptotic function. Under normal conditions, inhibition of CLU weakens this complex, allowing Bax to translocate to the mitochondria, triggering cytochrome c release, and activating caspase 9, initiating apoptosis [72]. |
| Cell Proliferation | c-Myc, a transcription factor encoded by the oncogene MYC involved in tumorigenesis, inhibits the expression of nCLU by upregulating the microRNA cluster miRNA-17 ~ 92 and attenuating the TGF-β axis, thus promoting angiogenesis and tumor growth in colon cancer [69]. | Blocking sCLU using apocynin, a substance that inhibits NADPH oxidase, halts the MEK-ERK1/2 pathway, resulting in reduced cell proliferation [73]. |
| Melittin inhibits sCLU, inactivating both the cholesterol/NF-κB/Bcl-2 axis and the cholesterol/p-ERK axis to suppress tumor growth in pancreatic cancer [74]. | ||
| sCLU exhibits an antiproliferative property by inactivating the TAK1/NF-κB axis, preventing the transforming growth factor beta receptor 1 (TGFBR1) from recruiting the TNF receptor-activating factor 6 (TRAF6)/TAK1-binding protein 2 (TAB2)/TGF-β-activated kinase 1 (TAK1) complex to inhibit tumor proliferation and growth in human non-small cell lung cancer (NSCLC) [75]. | ||
| Metformin, a conventional medication for type II diabetes, exhibits antitumor effects. Metformin suppresses sCLU, thereby hindering tumor growth through the inactivation of the SREBP-1c/fatty acid synthase (FASN) axis [76]. | ||
| sCLU promotes cell growth and proliferation by upregulating the expression of the calcium-binding protein S100A4 in renal cancer [77]. | ||
| Chemoresistance and Chemosensitivity | The stress response induced by treatments such as radiotherapy and chemotherapy lead to the overexpression of sCLU, a cytoprotective chaperone, which, by binding to activated Bax, it impedes the discharge of cytochrome c and prevents apoptosis [78]. | |
| Epithelial–Mesenchymal Transition and Metastasis | In nasopharyngeal carcinoma, CLU undergoes positive regulation by N, N′-dinitrosopiperazine (DNP), triggering MMP-9 and VEGF expression, thus facilitating to metastasis [79]. | |
| In breast cancer, CLU collaborates with eHsp90α to activate key signaling pathways, promoting EMT, migration, and tumor metastasis [80]. | ||
| In colon cancer, CLU interaction with platelets activates the p38MAPK pathway and positively regulates MMP-9, facilitating invasion [81]. | ||
| Studies on prostate cancer it has been demonstrated that miRNA-217-5p exerts control over the processes of invasion and migration by specifically targeting CLU [82]. |
| Types of Cancer | Expression of CLU In Vitro | Expression of CLU In Vivo |
|---|---|---|
| Non-small cell lung | Non-small cell lung cancer cell lines show overexpression upon treatment with chemotherapy or radiotherapy. ASO therapy sensitizes cells to these treatments and decreases their metastatic potential [103] | Patients exhibiting positive CLU expression tend to experience improved overall disease-free survival compared to those with negative CLU expression [104]. |
| More than 80% of the tumors are immunoreactive for CLU [104]. | ||
| Gastric | Overexpression of sCLU correlates significantly with metastasis, tumor invasion, and TNM stage. In addition, it correlates with unfavorable survival for advanced-stage gastric cancers [105]. | |
| Ovarian | Elevated sCLU levels show an inverse relationship with the tumor apoptotic index and are detected more frequently in metastatic lesions than in primary tumors [106]. | |
| Increased sCLU expression is associated with increased biological aggressiveness and decreased survival [107]. | ||
| Endometrial | When CLU is expressed in endometrial tumors, it is associated with a lower stage, supporting its role in the diagnosis of endometrial carcinoma [108]. | |
| There has been detected higher mRNA expression in both neoplastic and hyperplastic tissues compared to a normal endometrium. In this regard, an increase in mRNA expression of the specific sCLU isoform has been observed in neoplastic and hyperplastic endometrial diseases, but an increase in CLU protein has not been detected. Furthermore, specific CLU immunoreactivity has been observed in all glandular cells of the endometrium compared to other cellular compartments where CLU immunoreactivity was lower or absent [109]. | ||
| Increased CLU expression enhances paclitaxel resistance in endometrial cancer [110,111]. | ||
| Breast | Studies with the MDA-MB-231 cell line show how sCLU silencing significantly inhibits cell proliferation and drastically reduces cell invasion, cell progression and metastatic potential [112,113]. | Unlike benign lesions, atypical hyperplasias, intraductal carcinomas, and invasive carcinomas are characterized by CLU overexpression [114]. |
| Overexpression of sCLU is observed in a higher percentage of triple-negative breast cancer [115] and is associated with a negative estrogen and progesterone receptor status [114]. | ||
| Likewise, overexpression in the stroma tends to directly correlate with resistance to preoperative neoadjuvant chemotherapy in the primary tumor and inversely with the apoptosis rate, indicating that gene expression may not be necessary for apoptotic cell death [114,116,117]. | ||
| Colon | sCLU is overexpressed, while nCLU is downregulated [118]. Likewise, increased sCLU expression was predominantly observed in the cytoplasm of highly invasive tumors and metastatic lymph nodes [31], indicating that CLU expression might serve as a marker to identify patients with more aggressive tumors who could potentially benefit from targeted treatments [119]. | |
| Bladder | Treatment with the antisense oligonucleotide (ASO) targeting negative regulation of Bcl-2 and CLU increases the sensitivity of partially resistant bladder carcinoma cells to the tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) [120]. | The recurrence-free survival time of patients with overexpression of CLU was shorter than that of patients with normal CLU expression [121,122]. |
| Hepatocellular | High levels of sCLU are associated with migration, invasion, and metastasis [15] due to increased MMP-2 expression and decreased E-cadherin expression [123]. | |
| Furthermore, sCLU overexpression contributes to oxaliplatin resistance [15]. | ||
| In peripheral blood mononuclear cells (PBMC) from hepatocellular carcinoma patients, CLU has been proposed as a prospective detection biomarker along with other genes for its sensitivity and specificity [124]. The combination of CLU and AFP further improves diagnostic performance [125]. | ||
| The initial levels of CLU are higher for patients with progressive disease than for those with partial or complete response, respectively [126,127]. | ||
| Pancreatic | The inducer of ferroptosis, a type of cell death characterized by the accumulation of reactive oxygen species (ROS), interferes with apoptotic cell death by regulating CLU [128]. | CLU expression in stages I and II is not significantly associated with apoptosis. Moreover, patients with positive CLU expression present better survival rates [129]. |
| Melanomas | Increased expression is linked to heightened drug resistance and extended survival of tumor cells, whereas suppression diminishes resistance and lowers the survival rate of melanoma cells, both in laboratory settings and within living organisms [130]. | |
| Esophageal squamous cells | Elevated CLU expression is associated with unfavorable outcomes in locoregional, overall, and distant progression-free survival. Additionally, individuals exhibiting CLU overexpression in both epithelium and stroma tend to have shorter survival times [131,132]. | |
| Head and neck | Overexpression of CLU has been observed, but its implications have not yet been determined [133]. | Although CLU is detected in a low proportion of laryngeal carcinomas, it seems to exert a significant role in local invasiveness [133]. |
| Anaplastic large cell lymphomas | The role of CLU is unknown, but its expression within this lymphoma type provides an additional marker for diagnosis [134]. | |
| CLU expression does not correlate with the expression of anaplastic lymphoma kinase-1 (ALK-1). In reactive lymphoid tissues, only fibroblastic reticular cells and follicular dendritic cells exhibit positive expression [134]. | ||
| Osteosarcoma | sCLU overexpression is associated with metastasis and chemotherapy resistance [135]. | |
| Prostate | The expression of CLU increases in advanced stages of cancer, and its suppression sensitizes cells to chemotherapeutic drugs [136]. | It has been observed that CLU expression decreases considerably compared to benign tissues [137]. |
| Renal | The introduction of the CLU gene enhances the metastatic potential of renal cell cancer [138], while the removal of the CLU gene inhibits growth and migration [139]. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-García, D.; García-Aranda, M.; Redondo, M. Therapeutic Potential of Clusterin Inhibition in Human Cancer. Cells 2024, 13, 665. https://doi.org/10.3390/cells13080665
Martín-García D, García-Aranda M, Redondo M. Therapeutic Potential of Clusterin Inhibition in Human Cancer. Cells. 2024; 13(8):665. https://doi.org/10.3390/cells13080665
Chicago/Turabian StyleMartín-García, Desirée, Marilina García-Aranda, and Maximino Redondo. 2024. "Therapeutic Potential of Clusterin Inhibition in Human Cancer" Cells 13, no. 8: 665. https://doi.org/10.3390/cells13080665