
The Species Structure of Plankton Communities as a Response to Changes in the Trophic Gradient of the Mouth Areas of Large Tributaries to a Lowland Reservoir

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
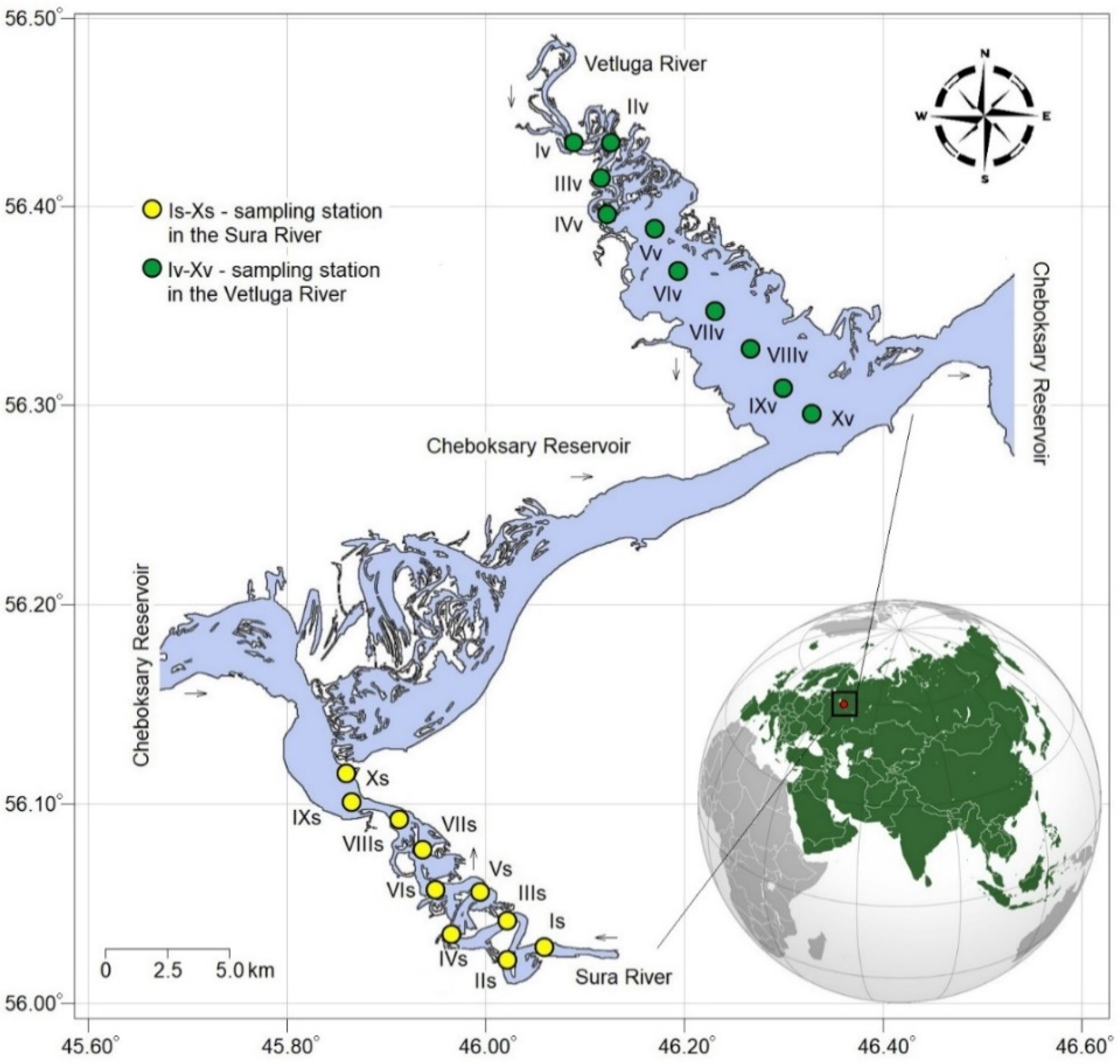
2.1. Study Area
2.2. Sample Collection and Environmental Indicators
2.3. Species Identification
2.4. Trophic State Index (TSI) and Data Analysis
3. Results
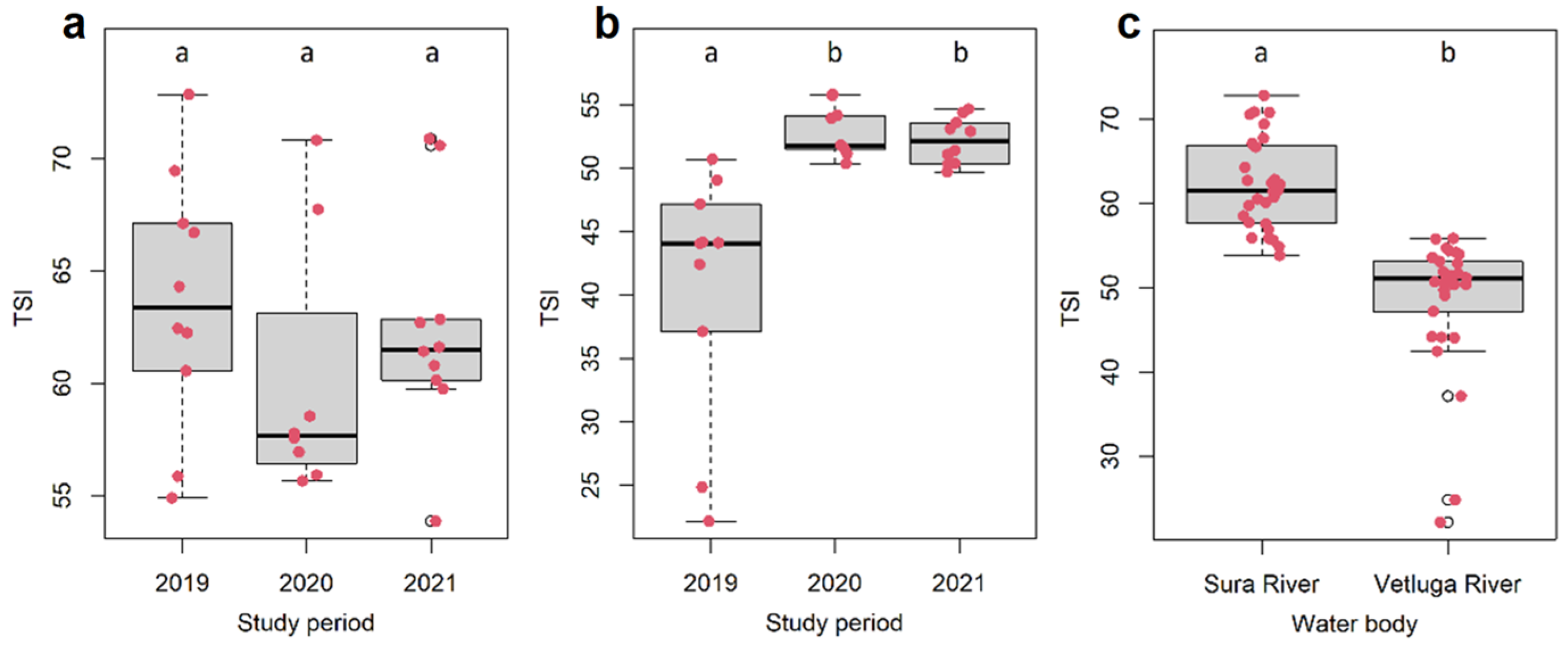
3.1. Environmental Indicators and Trophic State Index (TSI) Values
3.2. Taxonomical Structure of Plankton Communities
3.2.1. Phytoplankton Species Composition
3.2.2. Zooplankton Species Composition
3.3. Dominant Taxa
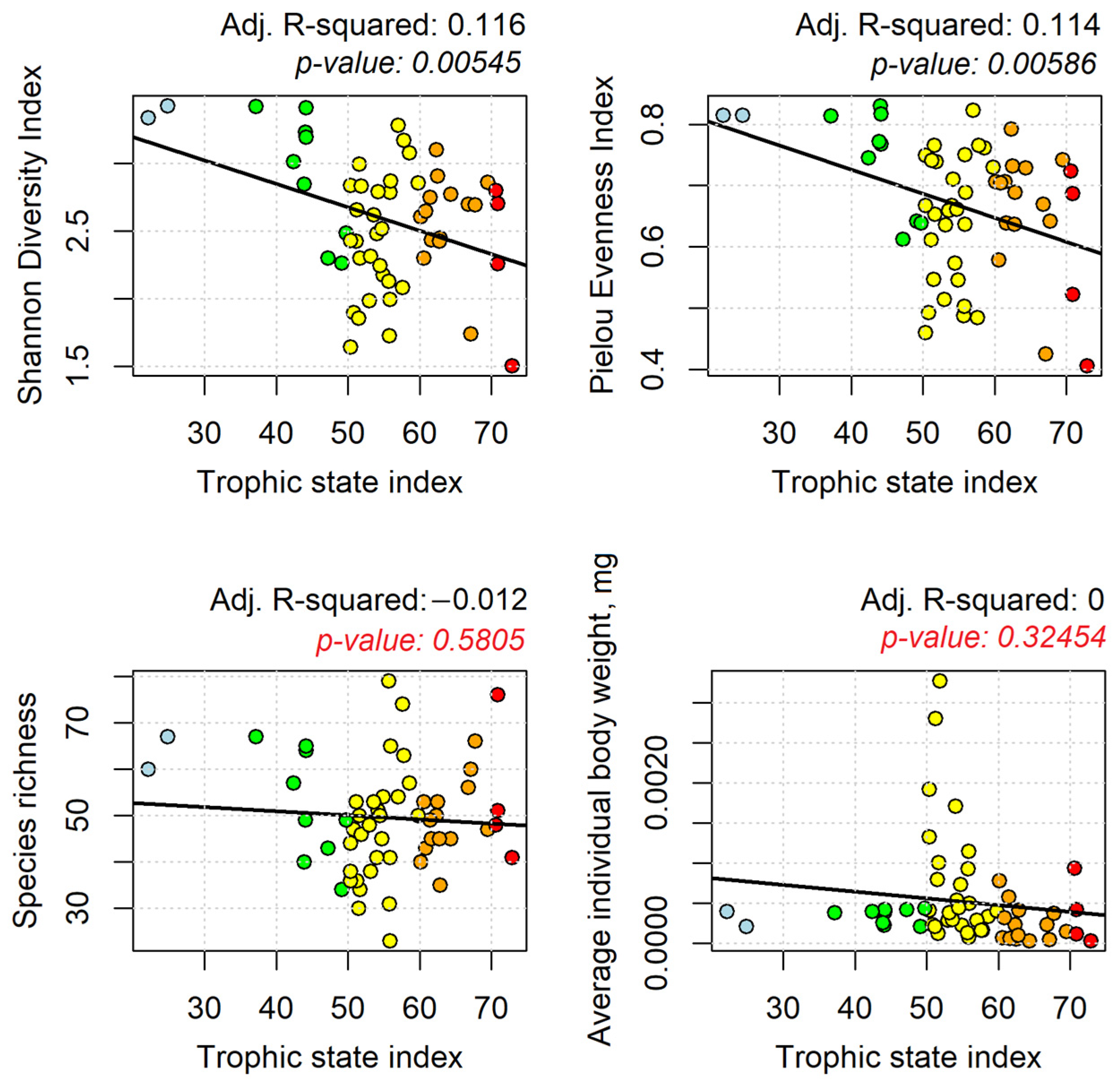
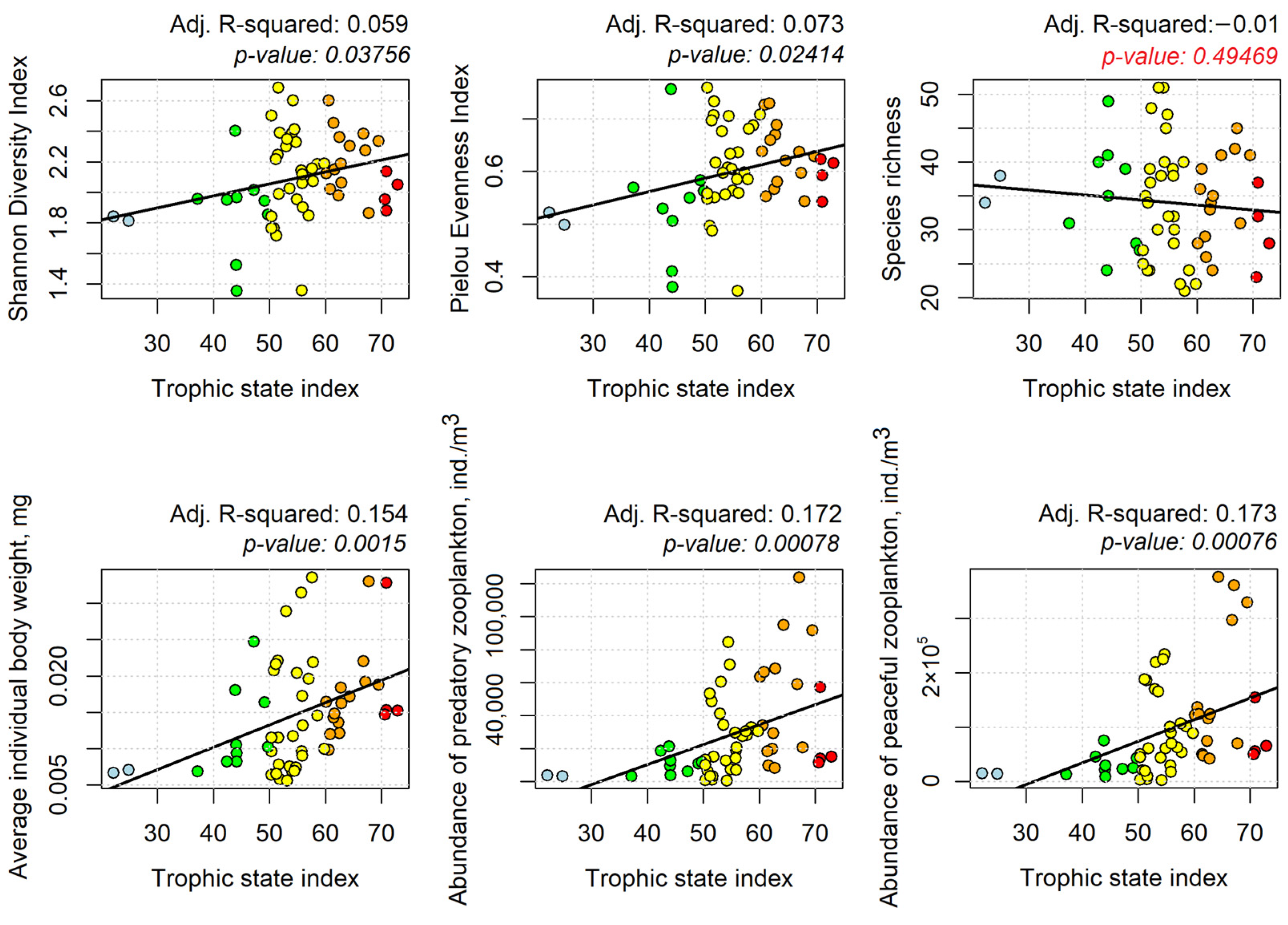
3.4. Correlation between Phyto- and Zooplankton Diversity Indicators and TSI
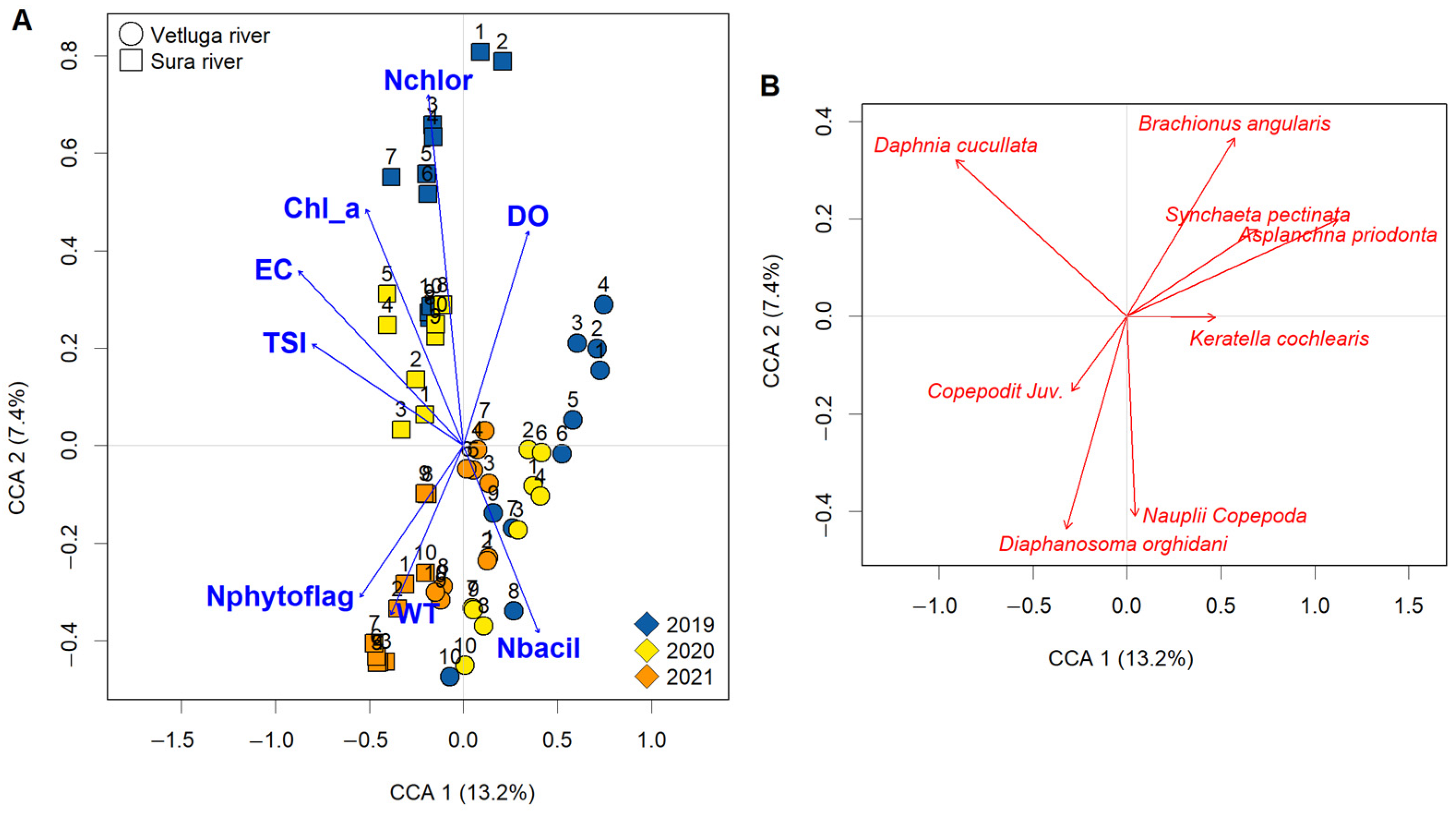
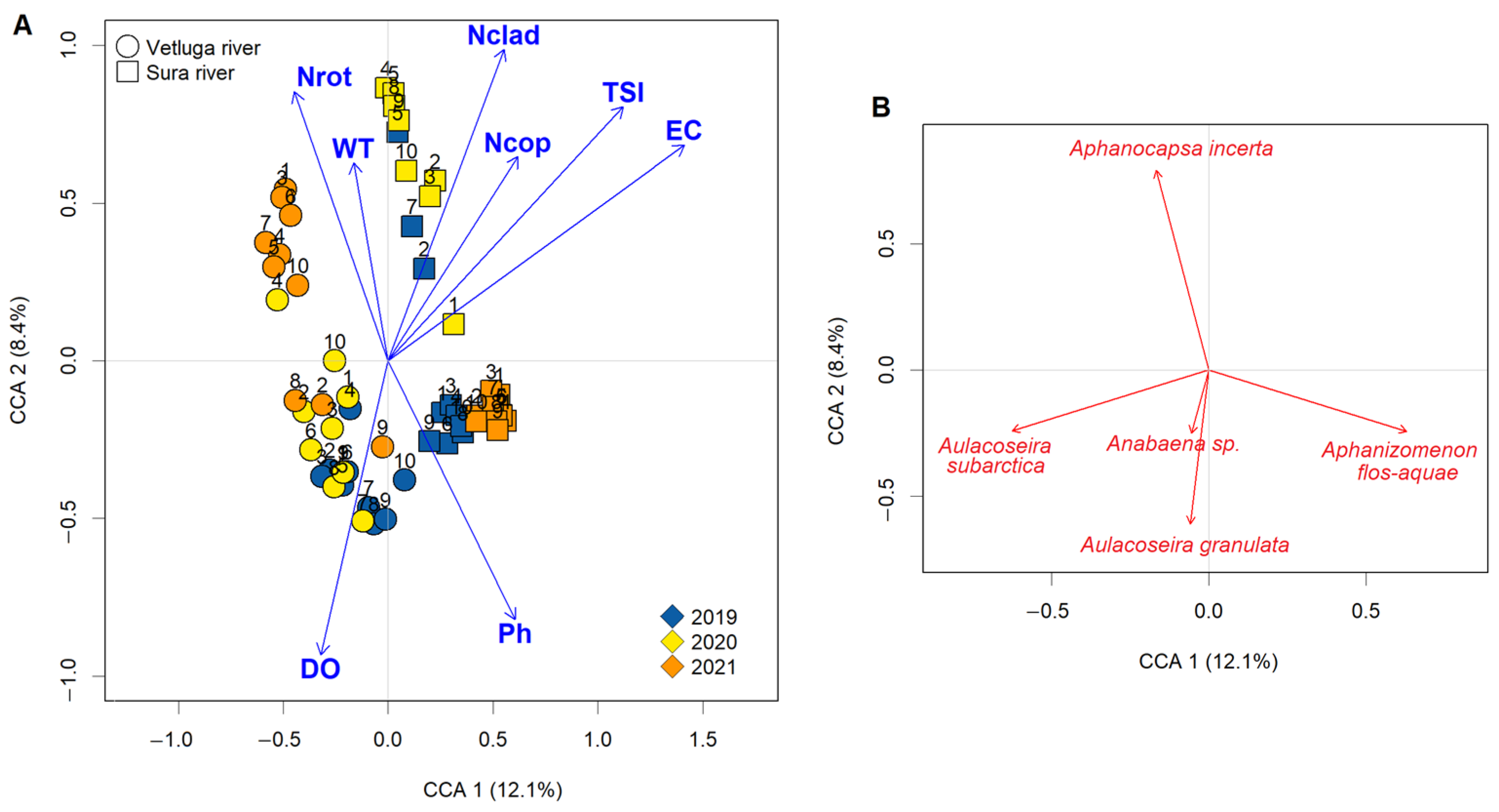
3.5. CCA of the Species Structure and Variation Partitioning
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chaurasia, S.; Gupta, R. Study on trophic state index of River Mandakini at Chitrakoot, India. Int. J. Adv. Res. Eng. Appl. Sci. 2016, 5, 34–43. [Google Scholar]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Ecology: Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [Green Version]
- Naeem, S.; Duffy, J.E.; Zavaleta, E. The functions of biological diversity in an age of extinction. Science 2012, 336, 1401–1406. [Google Scholar] [CrossRef] [Green Version]
- Gagic, V.; Bartomeus, I.; Jonsson, T.; Taylor, A.; Winqvist, C.; Fischer, C.; Slade, E.M.; Steffan-Dewenter, I.; Emmerson, M.; Potts, S.G.; et al. Functional identity and diversity of animals predict ecosystem functioning better than species-based indices. P. Roy. Soc. B-Biol. Sci. 2015, 282, 0142620. [Google Scholar] [CrossRef] [Green Version]
- Cloern, J.E. Our evolving conceptual model of the coastal eutrophication problem. Mar. Ecol. Prog. Ser. 2001, 210, 223–253. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.S.; Huszar, V.L.M.; Ovalle, A.R. Phytoplankton functional groups in a tropical estuary: Hydrological control and nutrient limitation. Estuar. Coasts. 2009, 32, 508–521. [Google Scholar] [CrossRef]
- Felipe-Lucia, M.R.; Soliveres, S.; Penone, C.; Fischer, M.; Ammer, C.; Boch, S.; Boeddinghaus, R.S.; Bonkowski, M.; Buscot, F.; Fiore-Donno, A.M.; et al. Land-use intensity alters networks between biodiversity, ecosystem functions, and services. Proc. Natl. Acad. Sci. USA 2020, 117, 28140–28149. [Google Scholar] [CrossRef]
- Padisák, J.; Reynolds, C.S. Selection of phytoplankton associations in Lake Balaton, Hungary, in response to eutrophication and restoration measures, with special reference to the cyanoprokaryotes. Hydrobiologia 1998, 384, 41–53. [Google Scholar] [CrossRef]
- Karadžić, V.; Subakov-Simić, G.; Krizmanić, J.; Natić, D. Phytoplankton and eutrophication development in the water supply reservoirs Garaši and Bukulja (Serbia). Desalination 2010, 255, 91–96. [Google Scholar] [CrossRef]
- Yang, B.; Jiang, Y.; He, W.; Liu, W.; Kong, X.; Jorgensen, S.E.; Xu, F. The tempo-spatial variations of phytoplankton diversities and their correlation with trophic state levels in a large eutrophic Chinese lake. Ecol. Ind. 2016, 66, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.L.; Pollard, A.I. Changes in the relationship between zooplankton and phytoplankton biomasses across a eutrophication gradient. Limnol. Oceanogr. 2018, 63, 2493–2507. [Google Scholar] [CrossRef]
- Shurganova, G.V.; Kudrin, I.A.; Yakimov, V.N.; Gavrilko, D.E.; Zhikharev, V.S.; Zolotareva, T.V. Spatial Distribution of Zooplankton on the Upper Part of the Cheboksary Reservoir. Inland Water Biol. 2018, 11, 317–325. [Google Scholar] [CrossRef]
- Shurganova, G.V.; Zhikharev, V.S.; Gavrilko, D.E.; Kudrin, I.A.; Zolotareva, T.V.; Yakimov, B.N.; Erina, O.N.; Tereshina, M.A. Zooplankton communities of the middle river part of the Cheboksary Reservoir and factors influencing their species structure. Pov. J. Ecol. 2019, 3, 384–395. [Google Scholar]
- Afonina, E.Y.; Tashlykova, N.A. Fluctuations in plankton community structure of endorheic soda lakes of southeastern Transbaikalia (Russia). Hydrobiologia 2020, 847, 1383–1398. [Google Scholar] [CrossRef]
- Frossard, V.; Aleya, L.; Henry, P.; Vallet, A.; Charlier, J.B. Impacts of nitrogen loads on the water and biota in a karst river (Loue River, France). Hydrobiologia 2020, 847, 2433–2448. [Google Scholar] [CrossRef]
- Liang, D.; Wang, Q.; Wei, N.; Tang, C.; Sun, X.; Yang, Y. Biological indicators of ecological quality in typical urban river-lake ecosystems: The planktonic rotifer community and its response to environmental factors. Ecol. Indic. 2020, 112, 106127. [Google Scholar] [CrossRef]
- Krupa, E.; Barinova, S.; Romanova, S.; Aubakirova, M.; Ainabaeva, N. Planktonic Invertebrates in the Assessment of Long-Term Change in Water Quality of the Sorbulak Wastewater Disposal System (Kazakhstan). Water 2020, 12, 3409. [Google Scholar] [CrossRef]
- Waite, I.R.; Pan, Y.; Edwards, P.M. Assessment of multi-stressors on compositional turnover of diatom, invertebrate and fish assemblages along an urban gradient in Pacific Northwest streams (USA). Ecol. Indic. 2020, 112, 106047. [Google Scholar] [CrossRef] [Green Version]
- Zhikharev, V.S.; Gavrilko, D.E.; Shurganova, G.V. Zooplankton community structure in mouth areas of different rivers (tributaries of the lowland Cheboksary Reservoir, European Russia). IOP Conf. Series Earth Environ. Sci. 2021, 834, 012063. [Google Scholar] [CrossRef]
- Telesh, I.V.; Khlebovich, V.V. Principal processes within the estuarine salinity gradient: A review. Mar. Pollut. Bull. 2010, 61, 149–155. [Google Scholar] [CrossRef]
- Gabyshev, V.A.; Ivanova, A.P.; Tsarenko, P.M. Diversity and features of the spatial structure of algal communities of water bodies and watercourses in the Lena River estuary. Inland Water Biol. 2019, 12, 1–9. [Google Scholar] [CrossRef]
- Krylov, A.V.; Tsvetkov, A.I.; Malin, M.I.; Romanenko, A.V.; Poddubnii, S.A.; Otjukova, N.G. Communities of hydrobionts and the physical-chemical characteristics of the estuary area of inflow of a flat water basin. Inland Water Biol. 2010, 3, 59–69. [Google Scholar] [CrossRef]
- Sakharova, E.G. Phytoplankton of Ecotone Zones of the Rybinsk Reservoir; Extended Abstract of Cand. Sci. (Biol.). Ph.D. Thesis, Papanin Institute for Biology of Inland Waters Russian Academy of Sciences, Borok, Russia, 25 May 2017; p. 23. (In Russian). [Google Scholar]
- Grabowska, M.; Ejsmont-Karabin, J.; Karpowicz, M. Reservoir-river relationships in lowland, shallow, eutrophic systems: An impact of zooplankton from hypertrophic reservoir on river zooplankton. Pol. J. Ecol. 2013, 4, 759–768. [Google Scholar]
- Tews, J.; Brose, U.; Grimm, V.; Tielborger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Heino, J.; Melo, A.S.; Siqueira, T.; Soininen, J.; Valanko, S.; Bini, L.M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshw. Biol. 2015, 60, 845–869. [Google Scholar] [CrossRef]
- Maishanu, H.M.; Mainarasa, M.; Magami, I.M. Assessment of productivity status using Carlson’s TSI and fish diversity of Gorongo Dam, Sokoto State, Nigeria. Path Sci. 2018, 4, 2001–2006. [Google Scholar] [CrossRef] [Green Version]
- Özen, A.; Sorf, M.; Trochine, C.; Liboriussen, L.; Beklioglu, M.; Søndergaard, M.; Lauridsen, T.L.; Johansson, L.S.; Jeppesen, E. Long-term effects of warming and nutrients on microbes and other plankton in mesocosms. Freshw. Biol. 2013, 58, 483–493. [Google Scholar] [CrossRef]
- Jeppesen, E.; Nõges, P.; Davidson, T.A.; Haberman, J.; Nõges, T.; Blank, K.; Lauridsen, T.L.; Søndergaard, M.; Sayer, C.; Laugaste, R.; et al. Zooplankton as indicators in lakes: A scientific-based plea for including zooplankton in the ecological quality assessment of lakes according to the European Water Framework Directive (WFD). Hydrobiologia 2011, 676, 279–297. [Google Scholar] [CrossRef]
- Rall, B.C.; Guill, C.; Brose, U. Food-web connectance and predator interference dampen the paradox of enrichment. Oikos 2008, 117, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Rip, J.M.K.; McCann, K.S. Cross-ecosystem differences in stability and the principle of energy flux. Ecol. Lett. 2011, 14, 733–740. [Google Scholar] [CrossRef]
- Okhapkin, A.G.; Yulova, G.A.; Startseva, N.A. Composition and structure of phytoplankton from certain flood-plain lakes of the cultural landscape (as exemplified by Nizhniy Novgorod). Bot. Zh. 2002, 87, 78–88. [Google Scholar]
- Reynolds, C.S. Ecology of Phytoplankton; Cambrige University Press: New York, NY, USA, 2006; p. 551. [Google Scholar]
- Korneva, L.G. Phytoplankton of Volga River Basin Reservoirs; Kostroma Printing House: Kostroma, Russia, 2015; p. 284. (In Russian) [Google Scholar]
- Chislock, M.F.; Doster, E.; Zitomer, R.A.; Wilson, A.E. Eutrophication: Causes, Consequences, and Controls in Aquatic Ecosystems. Nat. Educ. Knowl. 2013, 4, 10. [Google Scholar]
- Tian, W.; Zhang, H.; Zhao, L.; Xiong, Y.; Huang, H. Effects of Environmental Factors on the Temporal Stability of Phytoplankton Biomass in a Eutrophic Man-Made Lake. Water 2016, 8, 582. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Li, Z.; Li, L.; Lu, F.; Liu, Y.; Lu, H.; Fan, Y. Phytoplankton alpha diversity indices response the trophic state variation in hydrologically connected aquatic habitats in the Harbin Section of the Songhua River. Sci. Rep. 2020, 10, 21337. [Google Scholar] [CrossRef]
- Dunck, B.; Felisberto, S.A.; Nogueira, I.D. Effects of freshwater eutrophication on species and functional beta diversity of periphytic algae. Hydrobiologia 2019, 837, 195–204. [Google Scholar] [CrossRef]
- Li, Y.; Mengdie, G.; Jinlei, Y.; Yingxun, D.; Min, X.; Weizhen, Z.; Jianjun, W.; Haojie, S.; Rong, W.; Feizhou, C. Eutrophication decrease compositional dissimilarity in freshwater plankton communities. Sci. Total Environ. 2022, 821, 153434. [Google Scholar] [CrossRef]
- Suchora, M.; Szczurowska, A.; Niedźwiecki, M. Trophic state indexes and phytoplankton in the trophic status assessment of waters of a small retention reservoir at an early stage of operation. J. Ecol. Eng. 2017, 18, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, L.; Poikane, S.; Lyche Solheim, A.; Phillips, G.; Borics, G.; Catalan, J.; Hoyos, C.D.; Drakare, S.; Dudley, B.J.; Järvinen, M.; et al. Strength and uncertainty of phytoplankton metrics for assessing eutrophication impacts in lakes. Hydrobiologia 2013, 704, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Machado, K.B.; Bini, L.M.; Melo, A.S.; De Andrade, A.T.; De Almeida, M.F.; Carvalho, P.; Teresa, F.B.; De Oliveira Roque, F.; Bortolini, J.C.; Padial, A.A.; et al. Functional and taxonomic diversities are better early indicators of eutrophication than composition of freshwater phytoplankton. Hydrobiologia 2022. [Google Scholar] [CrossRef]
- Mineeva, N.; Lazareva, V.; Livinov, A.; Stepanova, I.; Chuiko, G.; Papchenkov, V.; Korneva, L.; Shcherbina, G.; Pryanichnikova, E.; Perova, S.; et al. Chapter 2. The Volga River. In Rivers of Europe; Elsevier: Amsterdam, The Netherlands, 2021; pp. 27–79. [Google Scholar]
- Krylov, A. (Ed.) Hydroecology of Lowland Water Reservoir Tributaries Mouth; Filigran: Yaroslavl, Russia, 2015; p. 466. (In Russian) [Google Scholar]
- Guidance on the Chemical Analysis of Sea and Fresh Water in the Ecological Monitoring of Fishery Water Bodies and the Environment of the World Ocean That Is Promising for Fishing; RFRIFO: Moscow, Russia, 2003.
- Borics, G.; Abonyi, A.; Salmaso, N.; Ptacnik, R. Freshwater phytoplankton diversity: Models, drivers and implications for ecosystem properties. Hydrobiologia 2021, 848, 53–75. [Google Scholar] [CrossRef]
- Malashenkov, D.V.; Dashkova, V.; Zhakupova, K.; Vorobjev, I.A.; Barteneva, N.S. Comparative analysis of freshwater phytoplankton communities in two lakes of Burabay National Park using morphological and molecular approaches. Sci. Rep. 2021, 11, 16130. [Google Scholar] [CrossRef] [PubMed]
- Vodeneeva, E.L.; Kulizin, P.V. Algae of the Mordovian Nature Reserve. In Annotated List of Species; Flora and Fauna of Nature Reserves: Moscow, Russia, 2019; Volume 134, p. 62. (In Russian) [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2021; Available online: http://www.algaebase.org (accessed on 29 July 2021).
- Okhapkin, A.; Sharagina, E.; Kulizin, P.; Startseva, N.; Vodeneeva, E. Phytoplankton community structure in highly-mineralized small gypsum karst lake (Russia). Microorganisms 2022, 10, 386. [Google Scholar] [CrossRef] [PubMed]
- De Paggi, S.J. New data on the distribution of Kellicottia bostoniensis (Rousselet, 1908) (Rotifera: Monogononta: Brachionidae): Its presence in Argentina. Zool. Anz. 2002, 241, 363–368. [Google Scholar] [CrossRef]
- Monchenko, V.I. Redescription of the oriental species Thermocyclops taihokuensis (Harada) (Copepoda: Cyclopoida) from its western most population. Zool. Middle East 2008, 43, 99–104. [Google Scholar] [CrossRef]
- Sukhikh, N.M.; Alekseev, V.R. Eurytemora caspica sp. nov. from the Caspian Sea—One more new species within the E. affinis complex (Copepoda: Calanoida, Temoridae). Proc. Zool. Inst. Russ. Acad. Sci. 2013, 317, 85–100. [Google Scholar] [CrossRef]
- Alekseev, V.R. Confusing invader: Acanthocyclops americanus (Copepoda: Cyclopoida) and its biological, anthropogenic and climate-dependent mechanisms of rapid distribution in Eurasia. Water 2021, 13, 1423. [Google Scholar] [CrossRef]
- Alonso, M. Fauna Iberica 7. In Crustacea Branchiopoda; Museo Nacional de Ciencias Naturales: Madrid, Spain, 1996; p. 486. [Google Scholar]
- Hudec, I. Anomopoda, Ctenopoda, Haplopoda, Onychopoda (Crustacea: Branchiopoda). Fauna Slovenska III; VEDA: Bratislava, Slovakia, 2010; p. 496. [Google Scholar]
- Błędzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe. Cladocera & Copepoda (Calanoida, Cyclopoida); Key to Species Identification, with Notes on Ecology, Distribution, Methods and Introduction to Data Analysis; Springer International Publishing: Cham, Switzerland, 2016; p. 918. [Google Scholar]
- Rogers, D.C.; Thorp, J.H. Thorp and Covich’s Freshwater Invertebrates: Volume 4: Keys to Palaearctic Fauna; Academic Press: Oxford, UK, 2019; p. 920. [Google Scholar]
- Rogers, D.C.; Thorp, J.H.K. (Eds.) Keys to Zooplankton and Zoobenthos of Fresh Waters of European Russia, Volume 1: Zooplankton; Tovarishchestvo Nauchn. Izd. KMK: Moscow, Russia, 2010. [Google Scholar]
- Jing, H.W.; Hua, L.; Sun, C.H.; Guo, J. Analysis on urban lakes’ eutrophication status in Beijing. Sci. Limnol. Sin. 2008, 20, 357–363. [Google Scholar]
- Zhang, Y.L.; Zhou, Y.Q.; Shi, K.; Qin, B.Q.; Yao, X.L.; Zhang, Y.B. Optical properties and composition changes in chromophoric dissolved organic matter along trophic gradients: Implications for monitoring and assessing lake eutrophication. Water Res. 2018, 131, 255–263. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Oxford, UK, 2012. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: http://www.R–project.org/ (accessed on 10 November 2022).
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. vegan: Community Ecology Package. R Package Version 2.6-2. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 11 November 2022).
- Maselli, V.; Pellegrini, C.; Del Bianco, F.; Mercorella, A.; Nones, M.; Crose, L.; Guerrero, M.; Nittrouer, J.A. River morphodynamic evolution under dam-induced backwater: An example from the Po River (Italy). J. Sediment. Res. 2018, 88, 1190–1204. [Google Scholar] [CrossRef] [Green Version]
- Liro, M. Dam reservoir backwater as a field-scale laboratory of human-induced changes in river biogeomorphology: A review focused on gravel-bed rivers. Sci. Total Environ. 2019, 651, 2899–2912. [Google Scholar] [CrossRef]
- McCabe, D.J. Rivers and Streams: Life in Flowing Water. Nat. Educ. Knowl. 2010, 1, 4. [Google Scholar]
- Deng, A.; Ye, C.; Liu, W. Spatial and seasonal patterns of nutrients and heavy metals in twenty-seven rivers draining into the South China Sea. Water 2018, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Jayme-Torres, G.; Hansen, A.M. Nutrient loads in the river mouth of the Río Verde basin in Jalisco, Mexico: How to prevent eutrophication in the future reservoir? Environ. Sci. Pollut. Res. Int. 2018, 25, 20497–20509. [Google Scholar] [CrossRef] [PubMed]
- Tockner, K.; Baumgartner, C.; Schiemer, F.; Ward, J.V. Biodiversity of a Danubian Floodplain: Structural, Functional and Compositional Aspects. In Biodiversity in Wetlands: Assessment, Function and Conservation; Backhuys Publishers: Leiden, The Netherlands, 2000. [Google Scholar]
- Guti, G. Significance of side-tributaries and floodplains for the Danubian fish populations. Arch. Hydrobiol. Suppl. 2002, 13, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Bazarova, B.B.; Tashlykova, N.A.; Afonina, E.Y.; Kuklin, A.P.; Matafonov, P.V.; Tsybekmitova, G.T. Biodiversity of the Ingoda River ecosystem. IOP Conf. Ser. Earth Environ. Sci. 2022, 962, 012006. [Google Scholar] [CrossRef]
- Okhapkin, A.G.; Genkal, S.I.; Scharagina, E.M.; Vodeneeva, E.L. Structure and dynamics of phytoplankton in the Oka River mouth at the beginning of the 21st century. Inland Water Biol. 2014, 7, 357–365. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, W.; Baoqing, S. Distribution of nitrogen and phosphorus and estimation of nutrient fluxes in the water and sediments of Liangzi Lake, China. Environ. Sci. Pollut. Res. Int. 2020, 27, 7096–7104. [Google Scholar] [CrossRef]
- Korneva, L.G. Invasions of alien species of planktonic microalgae into the fresh waters of Holarctic (Review). Russ. J. Biol. Invasions 2014, 5, 65–81. [Google Scholar] [CrossRef]
- Kulizin, P.V.; Gusev, E.S.; Vodeneeva, E.L.; Okhapkin, A.G. Taxonomic composition and morphology of silica-scaled Chrysophytes of some left-bank Volga tributaries. Inland Water Biol. 2021, 14, 357–367. [Google Scholar] [CrossRef]
- Kaštovský, J.; Hauer, T.; Mareš, J.; Krautová, M.; Bešta, T.; Komárek, J.; Desortová, B.; Heteša, B.; Hindáková, A.; Houk, V.; et al. A review of the alien and expansive species of freshwater cyanobacteria and algae in the Czech Republic. Biol. Invasions 2010, 12, 3599–3625. [Google Scholar] [CrossRef]
- Krztoń, W.; Kosiba, J.; Pociecha, A.; Wilk-Woźniak, E. The effect of cyanobacterial blooms on bio- and functional diversity of zooplankton communities. Biodivers. Conserv. 2019, 28, 1815–1835. [Google Scholar] [CrossRef] [Green Version]
- Pinel-Alloul, B.; Chemli, A.; Taranu, Z.E.; Bertolo, A. Using the diversity, taxonomic and functional attributes of a zooplankton community to determine lake environmental typology in the natural southern boreal lakes (Québec, Canada). Water 2022, 13, 578. [Google Scholar] [CrossRef]
- Sługocki, L.; Czerniawski, R. Trophic state (TSISD) and mixing type significantly influence pelagic zooplankton biodiversity in temperate lakes (NW Poland). PeerJ 2018, 6, e5731. [Google Scholar] [CrossRef] [Green Version]
- Pomari, J.; Kane, D.D.; Nogueira, M.G. Application of multiple-use indices to assess reservoirs water quality and the use of plankton community data for biomonitoring purposes. Int. J. of Hyd. 2018, 2, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Dodson, S. Land use, primary productivity, and lake area as descriptors of zooplankton diversity. Ecology 2005, 86, 255–261. [Google Scholar] [CrossRef]
- Monakov, A.V. Feeding of Freshwater Invertebrates; Kenobi Productions: Ghent, Belgium, 2003. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Indicators | 2019 | 2020 | 2021 | |||
|---|---|---|---|---|---|---|
| Sura River | Vetluga River | Sura River | Vetluga River | Sura River | Vetluga River | |
| Transparency (SD), m | 1.1 ± 0.1 | 1.1 ± 0.1 | 0.8 ± 0.1 | 0.9 ± 0.04 | 1.0 ± 0.1 | 0.9 ± 0.03 |
| Dissolved oxygen (DO), mg/L | 11.9 ± 0.5 | 8.7 ± 0.1 | 3.5 ± 0.04 | 8.5 ± 0.3 | 6.2 ± 0.3 | 8.3 ± 0.3 |
| pH | 9.3 ± 0.1 | 8.8 ± 0.01 | 7.8 ± 0.03 | 7.9 ± 0.1 | 8.5 ± 0.03 | 8.4 ± 0.04 |
| Elect. conductivity (EC), µS/cm | 630.6 ± 37.0 | 203.4 ± 11.8 | 693.9 ± 3.5 | 234.7 ± 10.0 | 701.3 ± 16.2 | 239.7 ± 3.8 |
| Water temperature (WT), °C | 22.2 ± 0.2 | 21.1 ± 0.1 | 21.8 ± 0.05 | 21.2 ± 0.1 | 24.5 ± 0.1 | 26.3 ± 0.3 |
| Chlorophyll-a (Chl-a), mg/L | 26.7 ± 2.6 | 6.3 ± 1.0 | 15.0 ± 2.5 | 10.4 ± 1.1 | 17.9 ± 2.5 | 11.8 ± 0.9 |
| Total phosphorus (TP), µg/L | 0.2 ± 0.04 | 0.04 ± 0.01 | 0.2 ± 0.04 | 0.1 ± 0.01 | 0.2 ± 0.03 | 0.1 ± 0.002 |
| Trophic Status | Dominant Taxa |
|---|---|
| oligotrophic | Aulacoseira subarctica (O. Müll.) E.Y. Haw.—22.6%; Aulacoseira granulata (Ehrenb.) Simonsen—10.1% |
| mesotrophic | Aulacoseira subarctica (O. Müll.) E.Y. Haw.—16.1%; Aulacoseira granulata (Ehrenb.) Simonsen—15.1% |
| slightly eutrophic | Aphanocapsa incerta (Lemmermann) G. Cronberg & Komárek—13.4%; Anabaena sp.—11.7% |
| medium eutrophic | Aphanocapsa incerta (Lemmermann) G. Cronberg & Komárek—15.2%; Aphanizomenon flos-aquae Ralfs ex Bornet & Flahault—10.9% |
| highly eutrophic | Aphanocapsa incerta (Lemmermann) G. Cronberg & Komárek—18.9% |
| Trophic Status | Dominant Taxa |
|---|---|
| oligotrophic | Synchaeta pectinata Ehrenberg, 1832—35.3%; Asplanchna priodonta Gosse, 1850—20.3%; Keratella cochlearis (Gosse, 1851)—11.9% |
| mesotrophic | Nauplius Copepoda—18.2%; Copepodite stages—13.0%; Brachionus angularis Gosse, 1851—10.9%; Asplanchna priodonta Gosse, 1850—10.0% |
| slightly eutrophic | Nauplius Copepoda—16.9%; Brachionus angularis Gosse, 1851—12.8%; Copepodite stages—11.8% |
| medium eutrophic | Nauplius Copepoda—14.7%; Brachionus angularis Gosse, 1851—14.1%; Copepodite stages—11.5%; Daphnia (Daphnia) cucullata Sars, 1862—11.4% |
| highly eutrophic | Nauplius Copepoda—19.3%; Brachionus angularis Gosse, 1851—17.6%; Daphnia (Daphnia) cucullata Sars, 1862—15.1%; Diaphanosoma orghidani Negrea, 1982—11.4% |
| Factors | Adjusted R2 | F | P | |
|---|---|---|---|---|
| Phytoplankton | Rotifera abundance (Nrot) | 0.015 | 1.88 | 0.001 * |
| Cladocera abundance (Nclad) | 0.015 | 1.83 | 0.002 * | |
| Copepoda abundance (Ncop) | 0.008 | 1.44 | 0.008 * | |
| Dissolved oxygen (DO) | 0.023 | 2.32 | 0.001 * | |
| Hydrogen indicator (pH) | 0.022 | 2.25 | 0.001 * | |
| Electrical conductivity (EC) | 0.048 | 3.81 | 0.001 * | |
| Water temperature (WT) | 0.017 | 2.00 | 0.001 * | |
| Trophic State Index (TSI) | 0.030 | 2.70 | 0.001 * | |
| Zooplankton | Bacillariophyta abundance (Nbacil) | 0.035 | 3.02 | 0.001 * |
| Chlorophyta abundance (Nchlor) | 0.034 | 2.94 | 0.003 * | |
| Phytoflagellate abundance (Nphytoflag) | 0.028 | 2.61 | 0.002 * | |
| Chlorophyll-a (Chl_a) | 0.027 | 2.57 | 0.001 * | |
| Electrical conductivity (EC) | 0.086 | 6.25 | 0.001 * | |
| Water temperature (WT) | 0.042 | 3.49 | 0.001 * | |
| Dissolved oxygen (DO) | 0.046 | 3.69 | 0.002 * | |
| Trophic State Index (TSI) | 0.078 | 5.75 | 0.001 * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhikharev, V.; Vodeneeva, E.; Kudrin, I.; Gavrilko, D.; Startseva, N.; Kulizin, P.; Erina, O.; Tereshina, M.; Okhapkin, A.; Shurganova, G. The Species Structure of Plankton Communities as a Response to Changes in the Trophic Gradient of the Mouth Areas of Large Tributaries to a Lowland Reservoir. Water 2023, 15, 74. https://doi.org/10.3390/w15010074
Zhikharev V, Vodeneeva E, Kudrin I, Gavrilko D, Startseva N, Kulizin P, Erina O, Tereshina M, Okhapkin A, Shurganova G. The Species Structure of Plankton Communities as a Response to Changes in the Trophic Gradient of the Mouth Areas of Large Tributaries to a Lowland Reservoir. Water. 2023; 15(1):74. https://doi.org/10.3390/w15010074
Chicago/Turabian StyleZhikharev, Vyacheslav, Ekaterina Vodeneeva, Ivan Kudrin, Dmitry Gavrilko, Natalia Startseva, Pavel Kulizin, Oxana Erina, Maria Tereshina, Alexander Okhapkin, and Galina Shurganova. 2023. "The Species Structure of Plankton Communities as a Response to Changes in the Trophic Gradient of the Mouth Areas of Large Tributaries to a Lowland Reservoir" Water 15, no. 1: 74. https://doi.org/10.3390/w15010074