Highlights
- Mean chlorophyll under 0.5–1 μgChla/L becomes robust and resilient in a salt lagoon.
- Both nitrogen and phosphorus loads contribute to eutrophication in a Mediterranean salt lagoon.
- Key factors in the Mar Menor (MM): phosphorus water–sediment relationship and deep water plants.
- Recovery nitrate-polluted aquifer makes the MM more robust and resilient.
- High risk of massive mortalities when mean chlorophyll is higher than 5 μgChla/L.
Abstract
Eutrophication is a significant threat to aquatic ecosystems worldwide, and the Mar Menor hypersaline lagoon exemplifies a coastal lagoon at risk of algal blooms due to excessive nutrient loads, nitrogen, and phosphorus. These nutrients originate from various sources within the lagoon’s catchment area, including urban, agricultural, and livestock activities. Regular and occasional loads—during flood events—produce algal blooms that can significantly reduce the water oxygen content and cause massive mortalities, as observed in recent years. A daily algal growth model (Mmag) was developed to better understand the processes and determine key elements such as the phosphorus water–sediment interchange and deep water plants that effect the entire ecosystem and algal growth. The analysis developed can be applied in other wetlands around the world facing similar challenges. In the Mar Menor, both nitrogen and phosphorus have high relevance depending on the period of the year and the phosphorus legacy in the sediments. Floods are the main phosphorus input to the lagoon (80–90%), which goes to the sediment and is released after during the warm months in the following years. At the end of summer, phosphorus released from the sediment and the regular nitrate inputs to the lagoon increase the algal bloom risk. The good status of deep water plants, which reduces the phosphorus release in summer, is a key element to reduce the algal bloom risk. An integrated set of measures is required to reduce the mean chlorophyll to levels under 1 or 0.5 µgChla/L that can make the Mar Menor more robust and resilient.
1. Introduction
Eutrophication is a significant threat to lakes and lagoons worldwide and is a global concern that is severely deteriorating aquatic ecosystems [1]. Nutrient excess is considered as the key factor for eutrophication [2,3,4], specifically nitrogen (N) and phosphorus (P) excess [5], from land sources such as agricultural and livestock practices and also urban discharges [6]. Excessive N and P loads to lakes and lagoons enrich the water [7], producing an increase in the plant growth, including algal bloom in the upper layers and an algae biological decomposition in deep water that reduces the oxygen content to hypoxic or anoxic conditions, finally producing heavy mortalities due to the lack of oxygen [1,8,9,10].
Algal growth depends on various factors, including irradiance, temperature, nutrient enrichment, species interaction, salinity, and hydrodynamics [1,11,12,13]. In some cases, nutrients are the key factor for algal growth [2]. Adequate irradiance (light availability) and water temperature are essential prerequisites for maintaining the photosynthetic and survival rates of algal blooms. Irradiance has shown a stronger correlation with phytoplankton growth compared to temperature, regardless of the nutrient concentration, and it exhibits higher sensitivity to growth than temperature [14]. Light, temperature, and inorganic nutrients significantly influence the biochemical processes of organisms and are considered major factors controlling seagrass growth [15]. During certain conditions, such as summer, when there is high irradiance and temperature, the availability of nutrients becomes the main factor for algal bloom development. To better understand these processes and the influence of internal nutrients, particularly phosphorus, on lake and lagoon management and restoration, one-dimensional water quality models have been developed in recent years [16].
In many cases, phosphorus is considered as the main driver for eutrophication and algal bloom formation [17]. However, nutrient proportions (N:P ratio) and forms can have an influence on biodiversity [18]. There are numerous examples where the eutrophication process is associated to the N presence or to the N concentration increase in the water [18,19,20,21,22,23,24,25], in contrast with the general view that is solely related to the phosphorus presence. Consequently, a P-only reduction strategy has been argued because of the fact that P (point source pollution) is often less costly to control than N (diffuse pollution) [18], which cannot be enough to avoid algal blooms, especially if the internal P loads are significant in the lake or lagoon [16].
The Mar Menor (MM) is a salt coastal lagoon that has suffered algal blooms with massive mortalities in recent years due to the nutrient loads from its catchment area [26]. This coastal area has been in a critical ecological condition since 1980, when intensive irrigation agriculture expanded to the surroundings of this salted lagoon [27]. This is a clear example of a singular natural ecosystem threatened by human activities [28], as it is happening in many lakes and lagoons worldwide. Related to the nutrient sources when produced, this lagoon exhibits a high level of complexity. On one hand, there are nutrient loads originating from agricultural and livestock practices (diffuse pollution) as well as from urban areas (point pollution). On the other hand, there are regular nutrient loads throughout the year and occasional nutrient loads during floods. Also, climate change scenarios can significantly affect the MM in the future due to the changes in the deep water plants’ response to the temperature [29].
The novelty of this work lies in establishing the mathematical connection between the nutrient loads generated in the catchment area and the eutrophication process in a coastal lagoon. This is obtained by combining a basin model with a daily lagoon model and analyzing, with a global and integrate overview, all the nutrient transport processes in the catchment and the internal factors in the lagoon. The connection between the internal lagoon factors and the nutrient sources from the catchment enables us to determine how each nutrient source impacts the lagoon and the effectiveness when each source is reduced. Understanding the relationship between the nutrient loads and the response of algal blooms is a crucial element to establish measures for preventing algal blooms in the MM lagoon, so the main purpose of this research is to identify the key factors that produce algal blooms in the Mar Menor in order to help its managers in formulating measures to reduce the risk of algal blooms.
2. Study Case and Data
Study Area
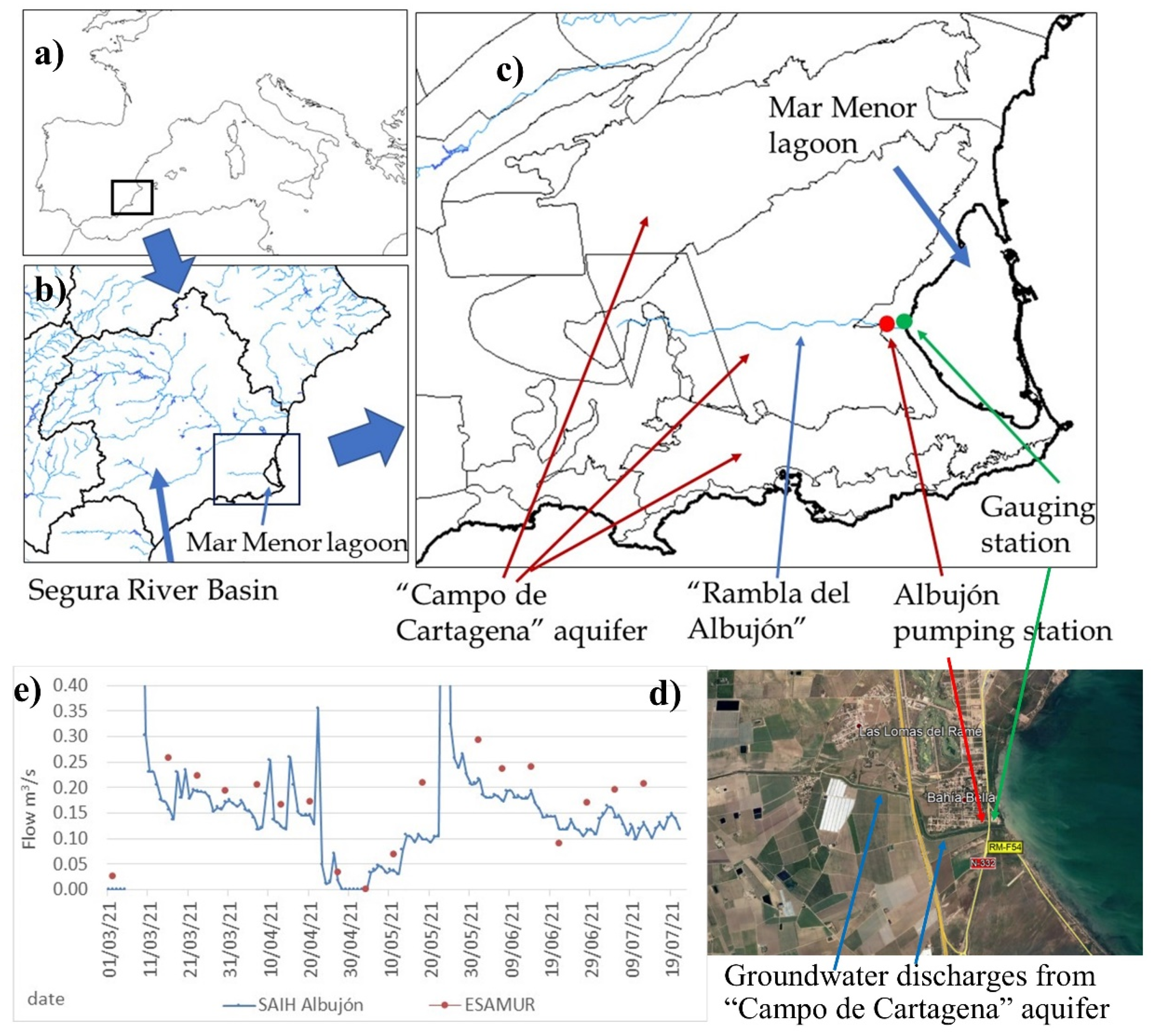
The Mar Menor (MM) is one of the largest (135 km2) hypersaline (38–46 psu, mean 44 psu) coastal lagoons in the Mediterranean [30], and it is usually more saline than Mediterranean Sea (38–40 psu), which gives the lagoon an extraordinary environmental value (Figure 1a) and constitutes it as a refuge for some species such as Pinna nobilis, a Mediterranean-endemic filter-feeding mollusk that has a massive mortality rate in most of the Spanish Mediterranean coast [31,32], relegating its populations to coastal lagoons or estuaries with salinities beyond the 36.5–39.7 psu [33]. The lagoon was designated as a Ramsar wetland in 1994 and it is located on the southeastern coast of Spain (37.7° N/−0.78° W) in a semi-arid region in the Segura River Basin District (RBD) (Figure 1b).
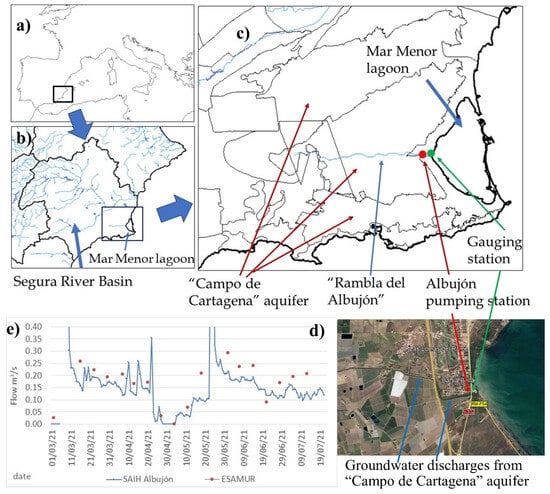
Figure 1.
(a) Mar Menor location, (b) Segura River Basin, (c) “Campo de Cartagena” aquifer and “Rambla del Albujón”, and the (d) final stretch of Rambla del Albujón with groundwater discharge areas marked (e) river flows (m3/s) to the Mar Menor from real-time gauging station (SAIH Albujón) and weekly measurements (ESAMUR).
The inland watershed is drained via “Rambla del Albujón” (Figure 1c) and several ephemerons rivers that only produce surface runoff during rainfall events. It has 1244 km2 with around 75% of agricultural land [26] that converts this area into one of the main national and international producers of agricultural food in Europe [34]. In the catchment area, it includes the “Campo de Cartagena” (CC) aquifer, a sedimentary multi-layered aquifer heavy polluted by nitrates (Figure 1c), with a mean value of 200 mgNO3/L and 250 mgNO3/L in the nearest area to the MM [35]. The aquifer is drained via the “Rambla del Albujón” in its final stretch [36] (3.3 hm3/year) and have groundwater direct discharges to the MM (6.9–8.5 hm3/year) [37,38] with high nitrate levels (250 mgNO3/L).
The water and nutrients inputs to the MM come from different mechanisms divided in regular inputs and occasional inputs (floods). Regular inputs are direct groundwater discharges and surface river flows in the “Rambla del Albujón” (Figure 1c), which contains Waste Water Treatment Plants (WTP) discharges (“Torre Pacheco” and “Los Alcázares”) with a low nitrate level and high phosphorus, as well as subsurface runoff from the CC aquifer, with a high nitrate content (250 mgNO3/L) and without phosphorus.
Surface regular flows in the “Rambla del Albujón” are measured with a recent real-time gauging station (SAIH Albujón, Figure 1d) and direct weekly measurements (ESAMUR, “Entidad de Saneamiento y Depuración de la Región de Murcia”). River flows vary from 0.10 to 0.15 m3/s (3.1 y 4.7 hm3/year) (Figure 1e), and its mean nitrate concentration is 180 mgNO3/L and 0.5 mgP/L for the total phosphorus. Total loads from the surface discharges are estimated in around 672 tnNO3/year and 1.9 tnP/year (Table 1), which are similar values as reported by the Segura River Basin Authority.

Table 1.
Nutrient loads to the Mar Menor per year and flood event, nitrate and total phosphorus.
During rainfall events, the nitrate concentration drop down to around 25–30 mgNO3/L and the total phosphorus concentration is around 0.5 or 1.0 mgP/L. The high amount of total phosphorus is possible to come from urban overflows or from the irrigation fields as phosphorus attached to the sediments, as is observed in other agricultural areas with a heavy historical application of phosphorus [39]. Occasional phosphorus loads during rainfall events can be 10–20 times higher than the regular loads. This fact produces a large amount of available phosphorus in the ecosystem during the following days after rainfall events, so there is a very high risk of algal blooms, and consequently, of mortality. Sentinel Satellite images of the MM [40] show an increase in the chlorophyll content 3–10 days after small rainfall events and how it requires around 1 month to recover the initial status [37].
The Mar Menor mean depth is 4.7 m, with a maximum depth around 7 m [41], and the water temperature has an annual cycle from 11 °C in February to 30 °C in August. The MM is connected to the Mediterranean Sea by three inlets: “Las Encañizadas”, “El Estacio” and “Marchamalo”. One of them, “El Estacio”, was widened in the 1970s with a 4–5 m depth to be navigable [42], which produced a sharp reduction in the salinity and temperature in the lagoon due to the greater interchange of water with the Mediterranean Sea [43]. The reduction in salinity also produced that Cymodocea nodosa (phanerogam) was replaced by Caulerpa prolifera (alga) in a large area of the lagoon and the transformation of the entire ecosystem. Currently, Caulerpa prolifera extends over the entire lagoon relegating Cymodocea nodosa only to the sandy bar next to the coast [44,45].
There are many estimations of the lagoon volume from 580 hm3 [46], 610 hm3 [47], and 630 hm3 [48] to 725 hm3 [49]. The lagoon has a large interchange flow with the Mediterranean Sea, where the total water inflow is estimated around 920 hm3/year via the “El Estacio” and “Marchalano” sea gates (Table 2). Also, there is a positive water balance and net inflow from the Mediterranean Sea of 171 hm3/year [49] to compensate the evaporation of 183 hm3/year [50] from the lagoon that produces the increase in the salinity.
Table 2.
Water interchanges between Mar Menor and Sea (adapted from [49]).
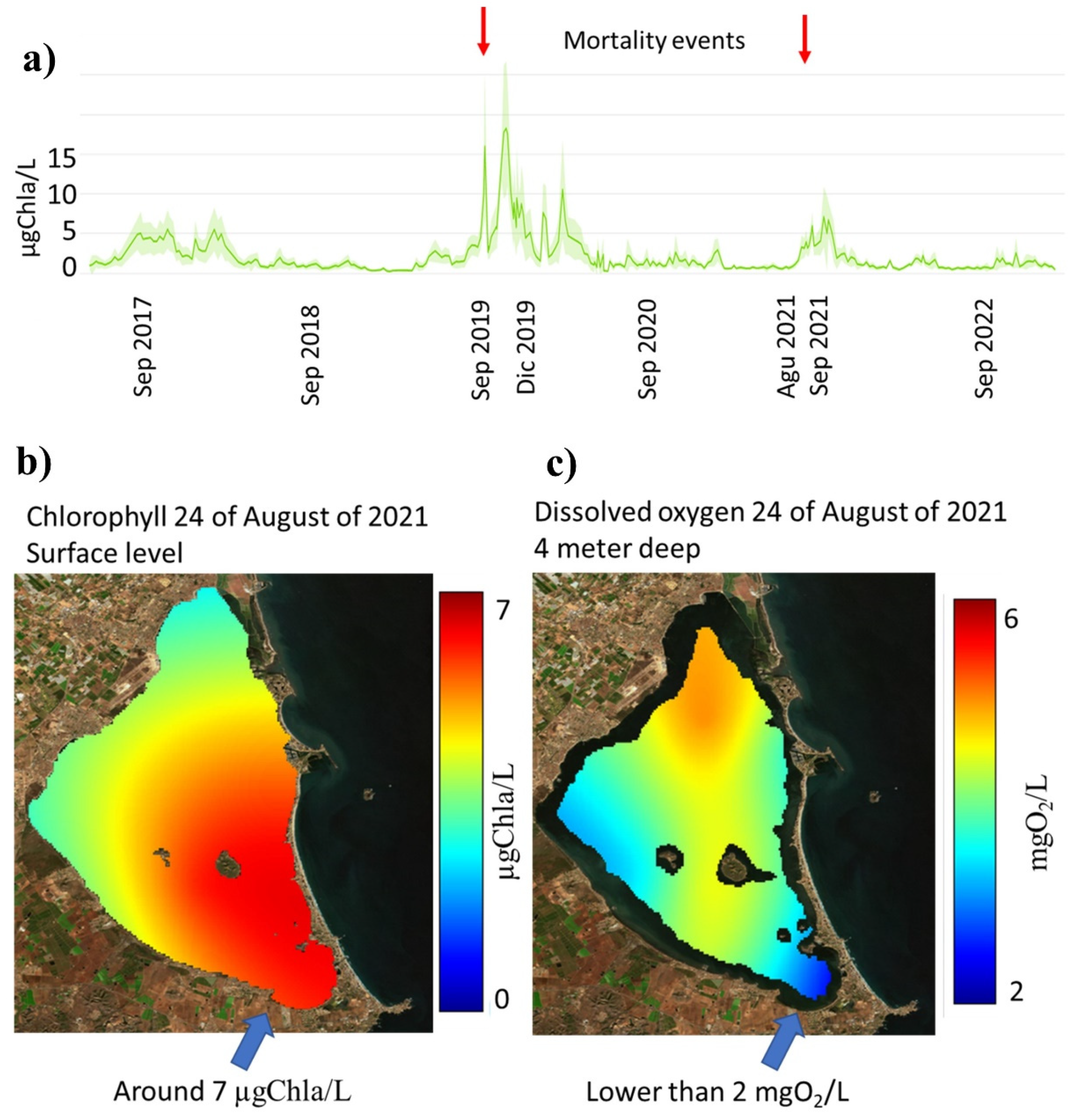
Mean chlorophyll levels in the MM (Figure 2a) are historically lower than 5 µgChla/L (1 µgChla/L = 1 mgChla/m3) [51,52]. When this mean level is overpassed, there is a high risk of mortalities in the MM, because in a specific area (the data from the Mar Menor scientific server [53]), chlorophyll can be greater than 7 or 10 µgChla/L (Figure 2b), and this can then produce phytoplankton degradation that consumes oxygen in deep water. If the oxygen levels are under 2 mgO2/L (Figure 2c), a massive mortality process can be produced, such as the mortalities which occurred in 2019 and 2021 [44,54]. Based on historical records, mean values lower than 2 µgChla/L are associated with a low risk of algal bloom in the lagoon, which is well related to the threshold defined under the Water Frame Work Directive (WFD) for the MM, which is 1.8 µgChla/L for a Good Status and 1.1 µgChla/L for a Very Good Status (Royal Decree 817/2015) [55].
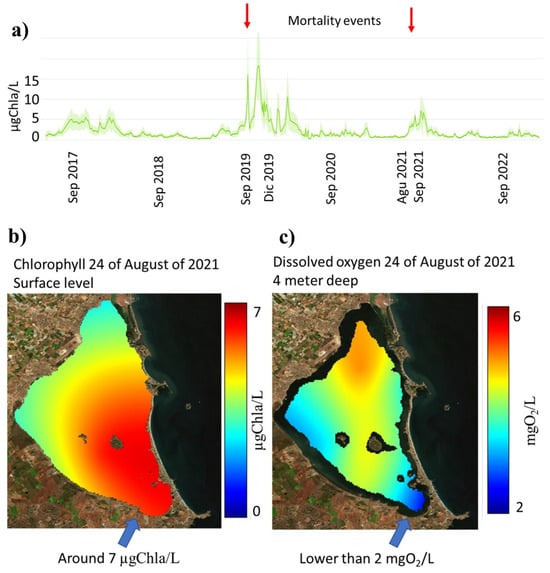
Figure 2.
(a) Mean chlorophyll values in Mar Menor, (b) chlorophyll map on 24 August 2021, and (c) dissolved oxygen map at 19 August 2021 when a mortality event occurs (data from Mar Menor scientific server [53]).
3. Methods and Models
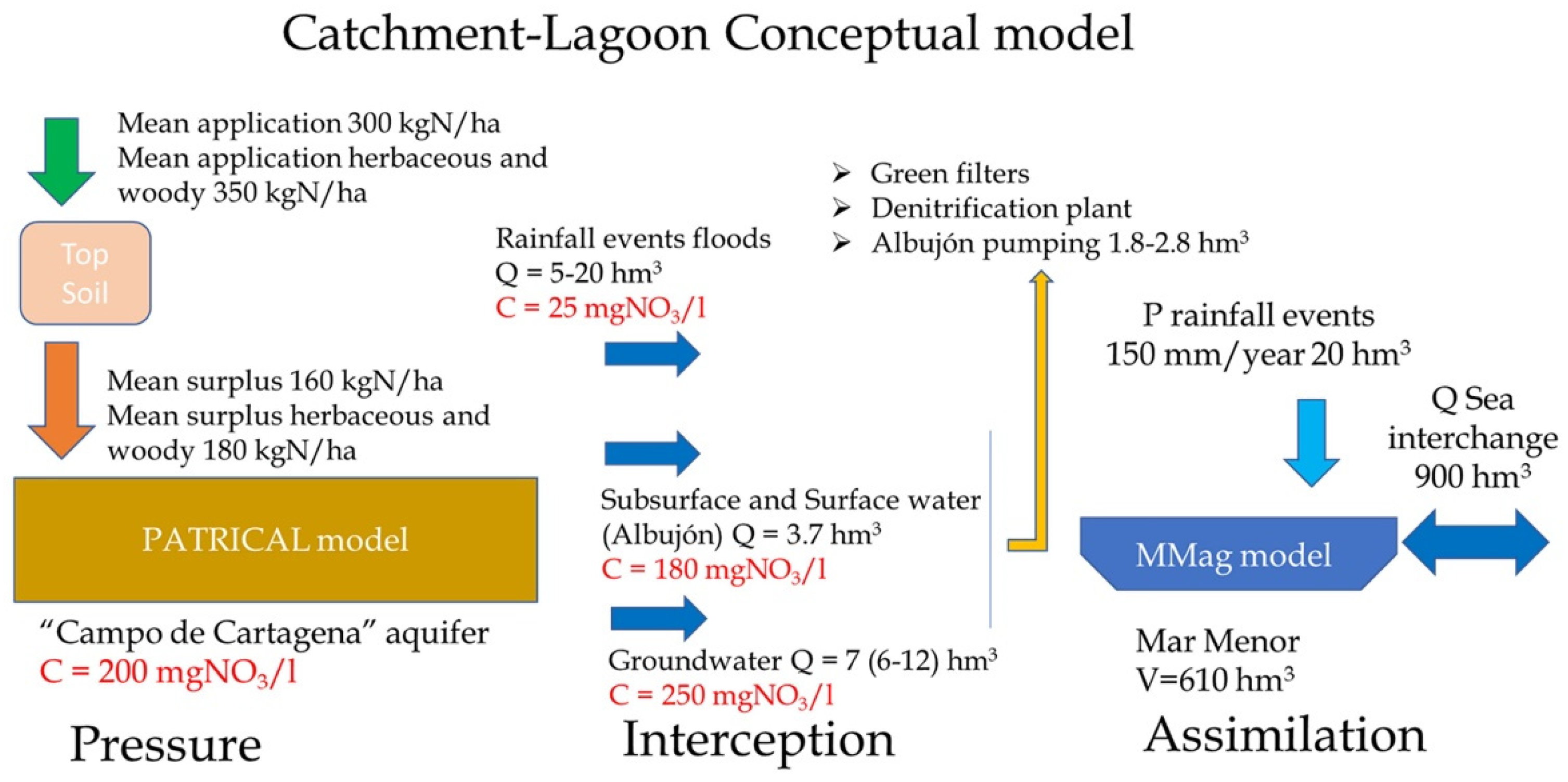
An integrated conceptual Catchment–Lagoon model (Figure 3) has developed and it reflects how nutrient loads, from nitrogen and phosphorus application in the agricultural fields and urban discharges, reach the Mar Menor lagoon, producing algal growth. Two models are used to analyze the relationship between the measures implemented in the catchment area and their effects in the MM lagoon. The first model, PATRICAL [56], establishes the connection between the nitrogen doses applied in agricultural fields and the resulting nitrate concentration in both the aquifer and surface runoff. The second model, Mmag (Mar Menor algal growth model), simulates daily algal growth based on the nutrient inputs to the lagoon.
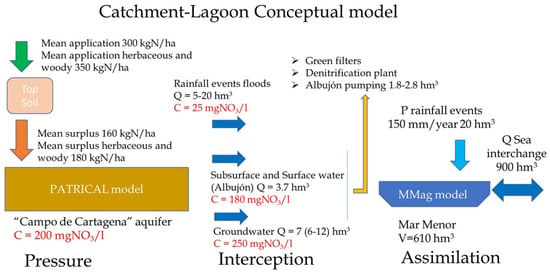
Figure 3.
Conceptual model Catchment–Lagoon, including model for the catchment PATRICAL and model for the lagoon MMag.
The conceptual Catchment–Lagoon model, related to the nutrient loads and its effect in the lagoon, has three main parts: pressure, interception, and assimilation. The first one, pressure, corresponds with the nutrient application in the catchment and how it arrives to the lagoon, which is simulated via the PATRICAL model. The second one, interception, corresponds with the different elements to remove or eliminate nitrate at the end of the catchment before it arrives to the Mar Menor, such as water pumping stations or green filters. The third one, assimilation, corresponds with the internal processes that occur in the lagoon with the nutrients and the algal bloom risk, which is simulated with the Mmag model.
The first part, pressure, is simulated via the PATRICAL model, which is a nitrogen transport model for large basins widely used in Spain; in the context of WFD implementation, it is used to determine the aquifer nitrate levels, necessary measures to recovery aquifers, and the time needed. The model is a large-scale (medium/large RBs), conceptual, monthly, and spatially distributed (grid 1 × 1 km2) water balance [57] and water quality model [56], for multi-decadal periods of 50–100 years. It is also used to assess the measures to recovery nitrate-polluted aquifers and their effect in environmental ecosystems [39].
Based on the Spanish Nitrogen Balance [58]’s average application of nitrogen, in herbaceous and woody crops, it is around 350 kgN/ha (300 kgN/ha over the entire aquifer) [37] and produces a nitrogen surplus in the form of pressure around 180 kgN/ha (160 kgN/ha over the entire aquifer) (Figure 3). This pressure is concordant with the nitrate concentration values calculated for the “Campo de Cartagena” aquifer via the PATRICAL model. Both model results and the data observed in the groundwater network indicate that the mean nitrate level in the aquifer is around 200 mgNO3/L and 250 mgNO3/L in the area near the contact with the MM.
Over the “Campo de Cartagena” aquifer, there is around 400 pig farms with 800,000 pigs that produces around 5700 tN/year. Total manures, pigs, and others are around 8000 tN/ha, which represents a pressure of 160 kgN/ha in the agricultural land over the aquifer. Since pig manure is not a fertilizer appreciated by farmers and there is no evidence of the massive use of it as a fertilizer in this area, the not-well-solved management of this manure can be a potential pressure over the aquifer representing 110–120 kgN/ha, which represents the largest part (65%) of the current nitrogen surplus calculated in this area (185 kgN/ha).
The second part, interception, corresponds with the available infrastructures to remove nutrients, such as pumping stations and green filters. Currently, there is only a water pumping station called “Bombeo del Albujón” at the end of the “Rambla del Albujón” (Figure 3) that can elevate around 1.8 hm3/year of water for reuse in irrigation. This station can take water from the drainage network, which drains the CC aquifer in this area, and from the final stretch of the “Rambla del Albujón”.
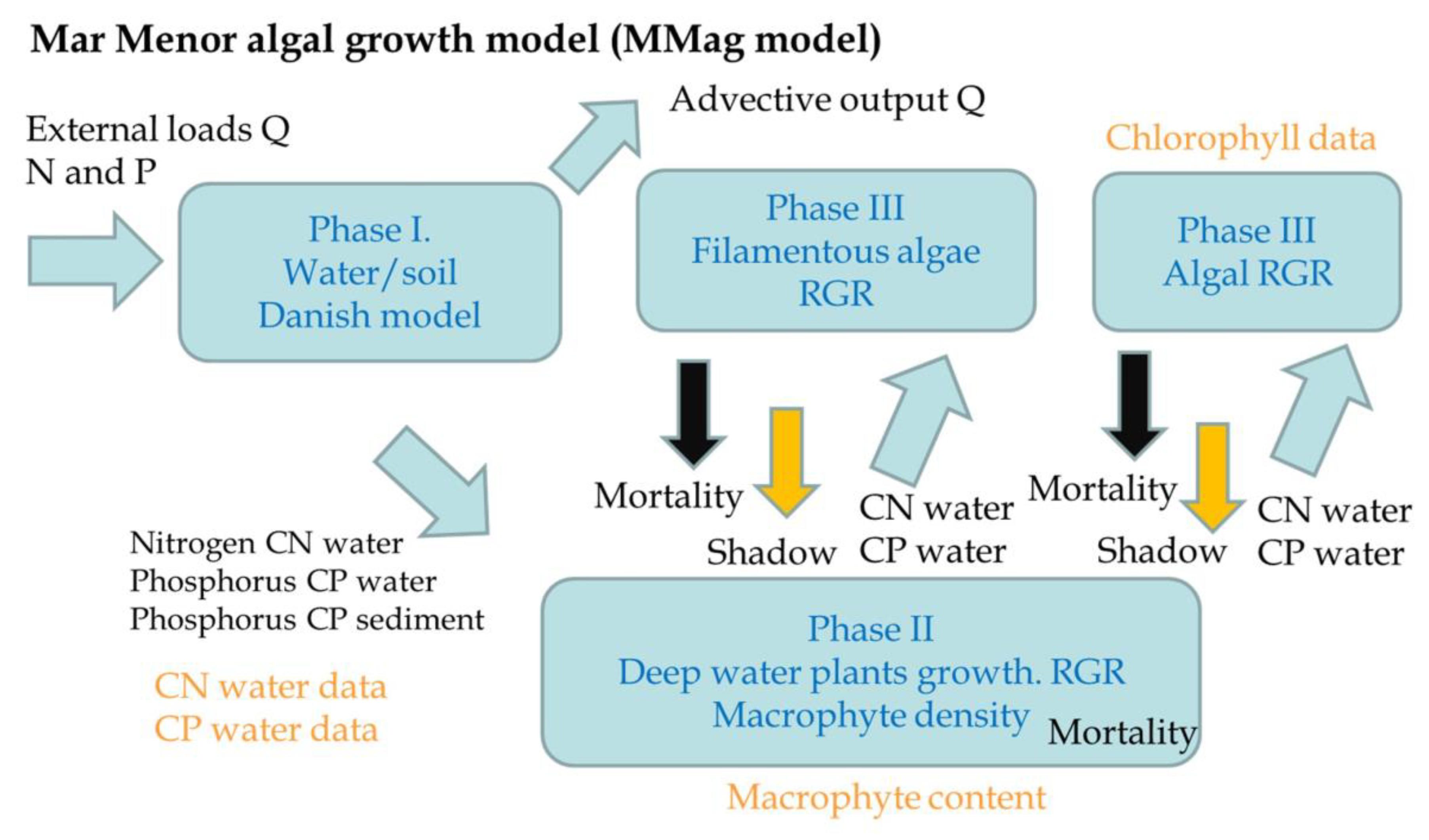
The third part, assimilation, is simulated via the MMag model, which is a daily model specifically developed to simulate algal growth in a well-mixed lagoon, considering the main nutrient inputs of nitrogen and phosphorus under stationary conditions (Figure 4). The model includes three main phases: phase I, the water–soil interaction based on an adapted version of the Danish model [59] for salt water, where the phosphorus and nitrogen content in water can be compared with real observations in water; phase II, the deep water plants’ growth that includes a combination of both Cymodocea nodosa (plant) and Caulerpa proligera (alga) growth, which is based on the Relative Growth Rate (RGR) and mortality rate that depend on water temperature [60] and salinity; and phase III, the upper layer algal growth, which is also based in the RGR and the mortality rate for both filamentous algae such as Chaetomorpha linum (“Cabello de Angel” in Spanish) or Enteromorpha intestinalis (“Ova marina” in Spanish), and other microscopic marine algae (phytoplankton), which is compared with chlorophyll measurements. The model was calibrated with the best data available on the nitrogen, phosphorus, and chlorophyll concentration in the MM. Model details and equations are included in the Supplementary Material.
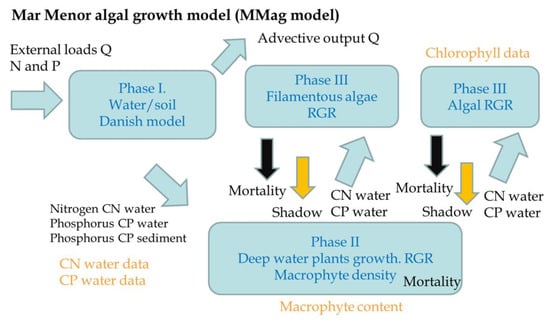
Figure 4.
Mar Menor algal growth model (MMag model) conceptual processes.
The intermediate results of the model are the deep water plants’ density and filamentous algae density, both in kg/m2, respectively indicate the amount of deep water plants in the soil and the quantity of filamentous algae in the water. The deep water plants’ density results are compared with the observed data of the amount of macrophytes in the Mar Menor.
4. Results and Discussion
4.1. Model Calibration and System Response
The model developed, the MMAg model, has been calibrated considering the current conditions to reproduce algal growth produced during the summer of 2021, when specific areas reached to 10 µgChla/L of the chlorophyll concentration. That year is repeated four times to check the stability of the model. The model is applied to a cell of the Mar Menor lagoon corresponding with 20% of the nearest area to the “Rambla del Albujón” mouth and includes the operation of the Albujón pumping station extracting around 1.8 hm3/year. A model calibration and sensitivity analysis is developed to determine how each parameter affects the model results, grading them from low to very high (Table 3).
Table 3.
Mmag model component parameters calibrated and parameters sensitivity.
The Danish model was originally developed for shallow fresh waters [59], hence the calibrated parameters obtained for the MM is five times higher than the original values and this result perfectly fits to a salt water lagoon, which is 5 to 10 times more soluble than fresh water [61]. By adapting the Danish model to the MM, it establishes a larger deposition of phosphorus in the sediments in winter with cool waters, and a larger release of phosphorus from the sediments in warm waters during summer. On the other hand, a nitrogen deposition indicates that 0.5% of the nitrogen available in water is deposited in the sediments each day.
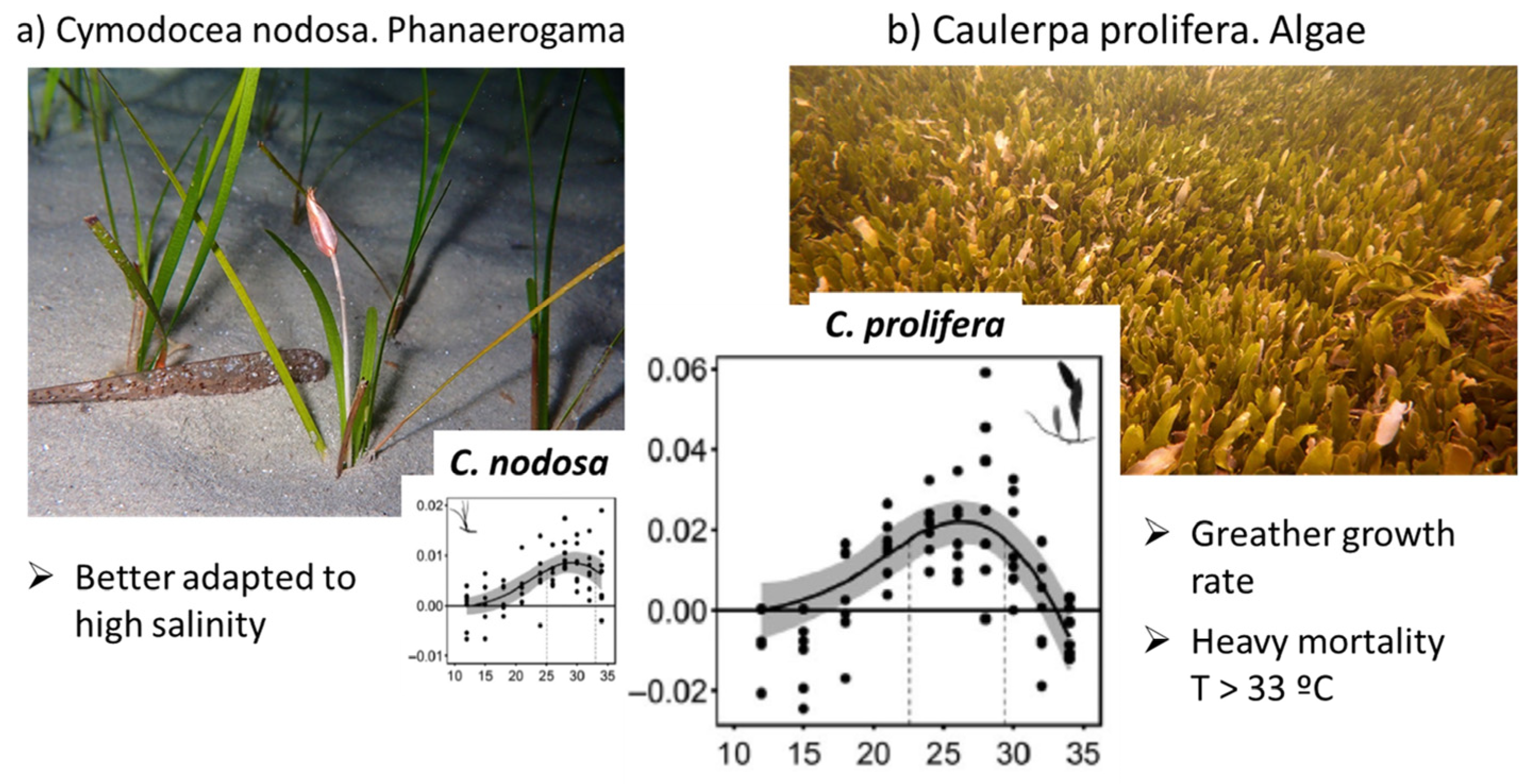
Model phase II corresponds with the deep water plants’ growth, such as phanerogams plants (Cymodocea nodosa) and algae (Caulerpa prolifera). The literature considers (Figure 5) that RGR for Cymodocea nodosa varies from 0.00 at a 15 °C water temperature to 0.01 (10 mg/g/day) at a 30° water temperature, and RGR for Caulerpa prolifera is twice as much as the previous case varying from 0.00 at 15 °C to 0.02 (20 mg/g/day) at 30 °C [60]. RGR is one of the most sensitive parameters of the model, becoming a key factor in the stability and behaviour of the model. The Cymodocea nodosa upper thermal limit is reported to be around 34 °C [62,63] or 34–35 °C [64], and for Caulerpa prolifera, it is reported to be 33 °C [60], which is one or two degrees lower than the other. It is recommended to develop specific studies to determine the RGR curves depending on water temperature and salinity for the Mar Menor deep water plant species.
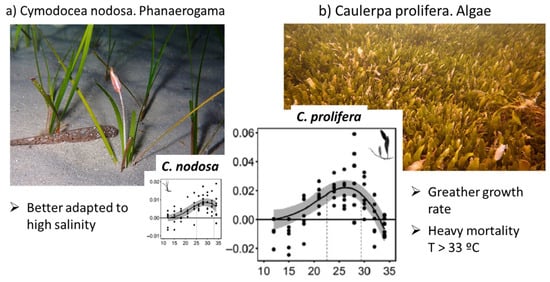
Figure 5.
Relative Growth Rate RGR for Cymodocea nodosa and Caulerpa prolifera (adapted from [60]).
Calibrated RGR for deep vegetation is 10 mg/d/day at 30 °C, which corresponds with the value of the Cymodocea nodosa, so the model results indicate that this species dominate the global response in the MM, as it is reported to have a larger presence of Cymodocea nodosa (phanerogam) [44] in the samples analyzed in the MM [65].
The third phase, corresponding with algal growth, has higher values of RGR than deep water plants, from 3 (filamentous) to 7 (algae) times greater, and its growth produces shadows that block light from entering into deeper waters, reducing the deep water plants’ growth and increasing its mortality. The algae mortality rate also has an important role with 0.6% per day for filamentous and 1.4% per day for algae. The phytoplankton concentration can be compared with the measured data of chlorophyll in the lagoon.
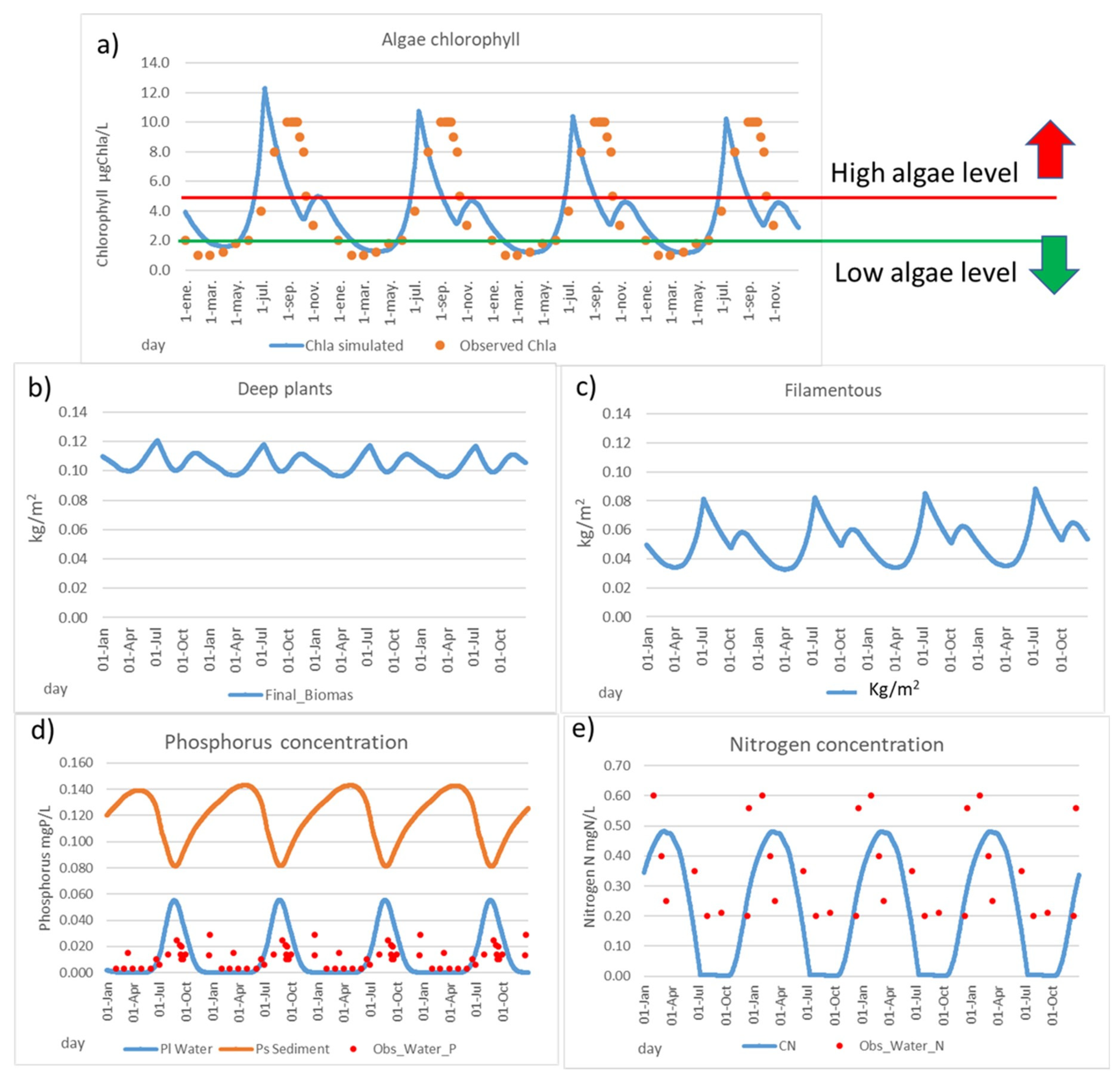
The results show the great importance of understanding the phosphorus release from sediment to water during the warm months in summer (Figure 6d). The amount of available phosphorus in winter is very reduced, where only the phosphorus that arrives from “Rambla del Albujón” is available before sedimentation is produced, due to the main direction of phosphorus changing to sedimentation in the cold water months, so phosphorus clearly limits algal growth in winter. On the other hand, the adapted Danish model produces a heavy release of phosphorus from the sediment to water in the warm months. In these months, a large amount of phosphorus is available in the ecosystem, so a rapid growth is produced; therefore, the available nitrogen in water is decreasing (Figure 6e), as well it is observed in the reduced number of available data. Hence, nitrogen can become the limiting factor in these months. This greater amount of available phosphorus in the warm months, combined with the nitrogen presence, produces a high risk of algal bloom in the warm months (Figure 6a).
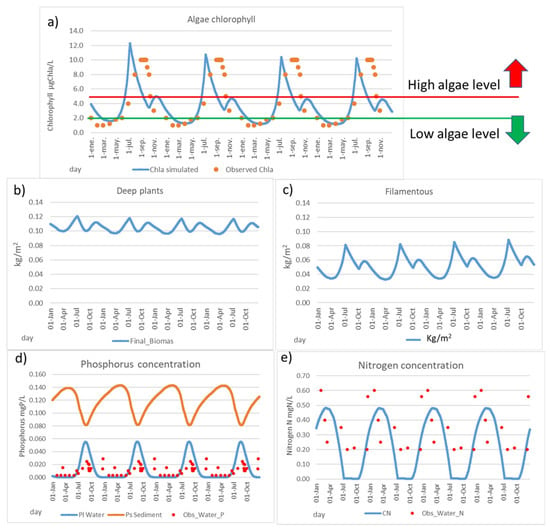
Figure 6.
Calibrated Mmag model. (a) Chlorophyll content (µgChla/L), (b) deep water plants density (kg/m2), (c) filamentous density (kg/m2), (d) phosphorus concentration in water and soil (sediment), soil equivalent in the water column (mgP/L), and (e) nitrogen content in water (mgN/L).
Based on the model results, most parts of available phosphorus in water come from the sediment (94%), so the internal load has a high relevant role in the behaviour of the system nowadays. These results are coherent with a large and increasing amount of phosphorus reported in the sediment in the last years from 293 mgP/kg in 2017 to 405 mgP/kg in 2020 [65]. Considering that the typical phosphorus content in soil can range from poor soils with 150 µg/g (=mg/kg) to rich soils with 700 µg/g [66], it can be considered that the MM sediments are currently rich in phosphorus content, as phosphorus legacy.
The deep water plants’ density (kg/m2) obtained is around 0.09–0.11 kg/m2 (Figure 6b), of which the value is very close to the total macrophyte content in the MM, as estimated in 2018 with 0.08 kg/m2 [45]. The amount of deep water plants has varied strongly during the last years due to the heavy rainfall (DANA) produced in December of 2016 and the posterior environmental crisis in the MM, mainly in the Cymodocea nodosa, but not affecting the Caulerpa prolifera located near the sea barrier [44]. The well-developed cover by Cymodocea nodosa in 2014 disappeared due to the DANA in 2016 to only 1400 tn. After the DANA, it has been in a recuperation process until 2018 with 6200 tn [45].
Deep water plants are one of the most important processes related to the algal blooms of the MM. In the model, the presence of a well-developed deep vegetation increases the nutrient retention and reduces the algal bloom risk. On the other hand, if deep water plants disappear, the algal bloom risk increases highly, as is also indicated by other authors mentioning marine benthic macrophyte meadows can be an important element in the resistance of the ecosystem to algal blooms [45]. Also, various deep water plants with different responses to temperature and salinity can produce diverse responses in the ecosystem [67].
In relation with the nutrient inputs, considering both nitrogen and phosphorus are necessary to increase the chlorophyll levels in the lagoon, a large release of phosphorus is produced in summer (high water temperature) from the sediments, so there is always enough phosphorus in the water during this period. Therefore, nitrogen becomes the limiting factor and changes in the external nitrogen inputs have a significant effect in the final chlorophyll levels in summer. The main phosphorus inputs from rainfall events have a significant effect in the chlorophyll levels through the increasing phosphorus legacy in the sediments and their subsequent release in the following summers.
Considering only continuous annual external inputs to the lagoon, the N:P ratio (=287) is extremely high due to the large amount of nitrogen. If it is also included, the phosphorus loads during floods the N:P ratio is reduced and ranges from 30 to 130. However, when it is included, the internal load of phosphorus from the sediment N:P ratio decreases to around 14. In all cases, the N:P ratio is more consistent with the green algae presence in the MM.
Controlling the nutrient inputs has allowed many lakes, lagoons, rivers, estuaries, and coastal ecosystems to recover rapidly from eutrophication. However, in some systems, recovery has been slow or negligible [12]. In addition, when a lake has a large internal loading of phosphorus, removing the external phosphorus source might not be an effective restoration technique for this lake [16]. Therefore, in the case of the MM in the present, controlling the regular nitrogen inputs and reducing the large amount of phosphorus inputs during floods, in order to reduce the phosphorus legacy and the internal load, are the key elements to reduce the algal bloom risk.
4.2. Measures to Reduce Algal Bloom Risk
The measures applied to eutrophic lake restoration are from the external measures to reduce the nutrient loads to the internal measures designed to either retain nutrients in the sediments or remove them from the lake ecosystem [68]. Multiple strategies have to be developed in the Mar Menor [69], so a set of different types of measures will be applied in the next years in the catchment and the MM.
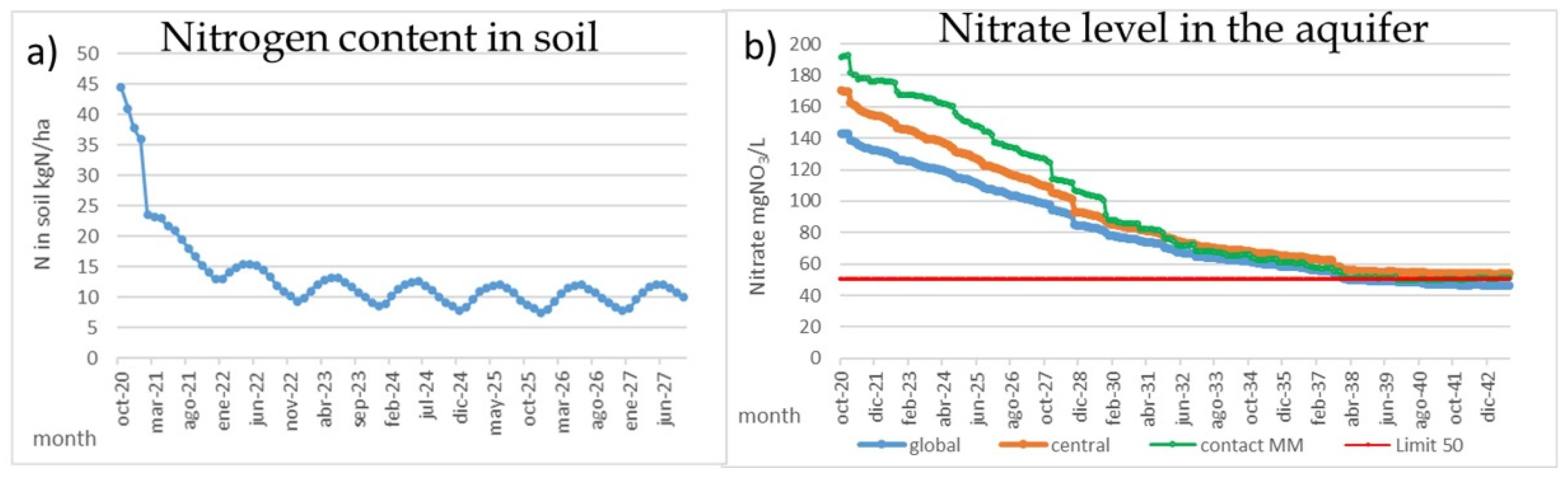
The most effective measure for the MM is to reduce the nitrate pollution in the “Campo de Cartagena” aquifer and accomplish the WFD requirements for groundwaters, which dictates that nitrate levels must be under 50 mgNO3/L. Based on the PATRICAL model results, this can be obtained by reducing the nitrogen surplus by means of improving the irrigation systems and reducing half the amount of nitrogen applied in this area. It is demonstrated that current nitrogen application, both mineral and manures, produces a large amount of nitrogen excess and heavy pollutes the aquifer. This reduction has two significant effects. The first one is a great reduction in a few years, around 2–3 years, in the amount of nitrogen and phosphorus in soil (Figure 7a) that could be mobilized during rainfall events and reach to the MM, increasing the algal bloom risk after floods. The second one is the gradual reduction in the CC nitrate levels to 100 mgNO3/L in around 6 years and 50 mgNO3/L in around 12 years (Figure 7b), so the nitrate loads from the aquifer to the MM will significantly lower in the medium-term, with 50 mgNO3/L for the direct groundwater discharges and 30 mgNO3/L for the surface waters.
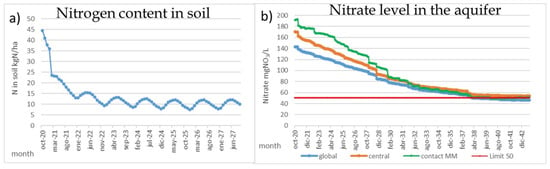
Figure 7.
Reduction in nitrogen content in soil (kg/ha) and nitrate levels (NO3/L) in the aquifer after reduction in fertilizer application in the “Campo de Cartagena”. Results obtained with PATRICAL model.
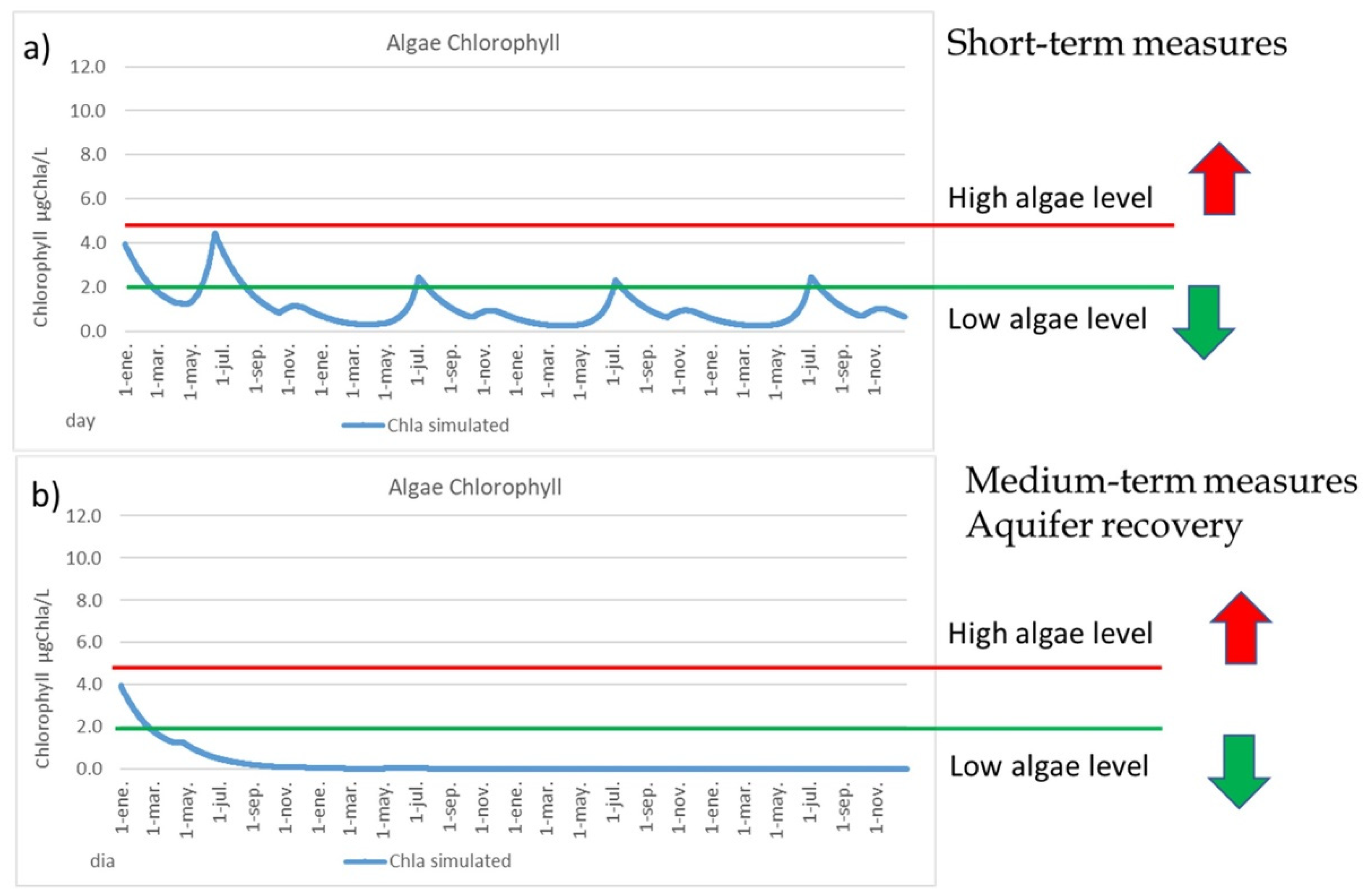
While the medium-term measures reduced the nitrate levels in the aquifer, other measures can also apply in the short-time, such as artificial wetlands and green filters around the MM, a denitrification system for the surface runoff of the “Rambla del Albujón”, an improvement of the WWTPs and an increase of the Albujón pumping station capacity (from current 1.8 hm3/year to 2.8 hm3/year). None of these measures are enough by itself to reduce the risk of algal blooms in the MM, so a combination of them is necessary, and together, can reduce the nitrate levels in the surface runoff from 180 mgNO3/L to around 50 mgNO3/L. Under these conditions, there is a small reduction in the deep water plants’ density, due to the lesser amount of nutrients that reach the lagoon; the phosphorus content in soil is reduced to around 20%, which is associated with a lower deep water plants’ density; there is a heavy reduction in the filamentous plant density, to less than half; and finally, a great reduction in the chlorophyll levels is produced. Most of the time, the chlorophyll levels are below 2 µgChla/L (Figure 8a), so the algal bloom risk will be low. Only in the warmer months of July and August will the chlorophyll be around 2 µgChla/L. However, under these conditions, there is still a remaining algal bloom risk in the MM if a flood occurs with a large contribution of nutrients, so the MM is still vulnerable.
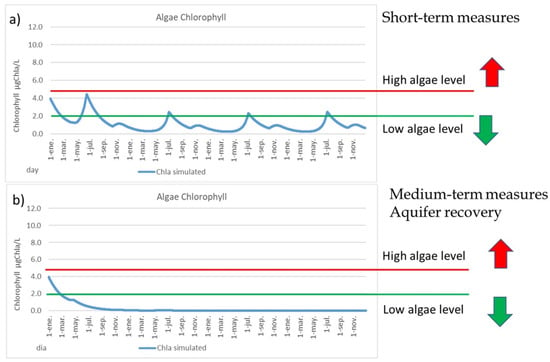
Figure 8.
(a) Chlorophyll content (µgChla/L) for the short-term measures. (b) Chlorophyll content (µgChla/L) for the medium-term measures.
Applying both type of measures, in short-term and in medium-term, the chlorophyll levels and algal bloom risks are very low (Figure 8b), the filamentous algae disappears and the phosphorus content in soil is strongly reduced, so the phosphorus legacy in the MM is gradually reduced. In this scenario, the PATRICAL model indicates that the amount of nutrients in agricultural soil are four times lower, so the nutrients contributions during flood events will be lower to the lagoon, and then the MM will be more robust and resilient against flood events that may occur.
5. Conclusions
The Mar Menor coastal lagoon is one of the largest hypersaline lagoon in the Mediterranean that is threatened by excessive nutrient loads, nitrogen, and phosphorus from its contributive basin. In the last few years, different events were characterized by a heavy rise of chlorophyll levels above 5 µgChla/L, which have produced massive mortality in many species, so this chlorophyll level can be considered as a high risk of algal blooms. Mean chlorophyll levels lower than 2 µgChla/L can be considered as a low level of phytoplankton for the MM lagoon. Although, to make the lagoon more robust and resilient, the mean chlorophyll level should be lower than 0.5–1 µgChla/L.
Solar irradiance, nitrogen, and phosphorus are necessary for algal bloom. Nitrogen comes from the “Campo de Cartagena” aquifer, a heavy nitrate-polluted aquifer (200 mgNO3/L). Floods are the main external phosphorus source that can contribute 10 to 20 times the annual load from the regular sources, increasing the phosphorus legacy in the sediments of the MM. This phosphorus will be released to the water in the following years during the warm months, where around 94% of phosphorus inputs to the water come from internal loads from the sediments, increasing the algal bloom risk in these months.
The Mar Menor daily model of algal growth (MMag) shows the great importance of deep water plants in the behavior of the MM and its resilient capacity. A well-developed layer of deep water plants absorbs and retains more nutrients and avoids the phosphorus release in summer, significantly reducing the algal bloom risk. In winter, especially in the cold water months, phosphorus tends to move from water to the sediment, increasing the phosphorus content in soil, the phosphorus legacy, and limiting the algal growth. On the other hand, during the warm months in summer, there is an important phosphorus release from soil to the water, so a large amount of phosphorous is available for filamentous and algal growth, becoming nitrate as the limiting factor in these months.
Limiting the regular nitrogen inputs and occasional phosphorus inputs during floods are the main factors to reduce the algal blooms’ risk in the Mar Menor. Measures to recover the MM need to reduce the mean chlorophyll level below 1 or 0.5 µgChla/L to make the system more resilient in the face of large nutrient loads that are produced during flood events, especially phosphorus loads.
The best measure to reduce the nutrient inputs to the MM is to decrease the nitrate levels in the “Campo de Cartagena” aquifer, following the requirements of the Water Framework Directive that will require between 6 and 12 years, due to aquifer inertia. This situation can be obtained by means of improving agricultural practices, including reducing the nutrient doses to half, which also reduces the amount of nitrogen and phosphorus content in soil, which can be mobilized during flood events.
During the intermediate period before aquifer recovery takes place, implementing short-term measures such as artificial wetlands, green filters, denitrification plants, and increasing the volume pumped in the “Albujón” pumping station help reduce the nitrogen loads entering the MM and mitigate the risk of algal blooms under normal conditions. However, it is important to note that there is still a risk of algal blooms if severe flood events occur.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15203569/s1.
Funding
This work was supported by the Spanish Ministry for the Ecological Transition and the Demographic Challenge.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
The author agreed with the content and gave explicit consent to submit. Consent was obtained from the responsible authorities at the institute/organization where the work has been carried out, before the work was submitted.
Data Availability Statement
Data available on request due to restrictions.
Acknowledgments
The author would like to acknowledge the Segura River Basin District-Confederación Hidrográfica del río Segura-; the General Directorate for Water of the Spanish Ministry for the Ecological Transition and Demographic Challenge -Dirección General del Auga del Ministerio para la Transición Ecológica y el Reto Demográfico- that funded this research; and also, the contributions of the student Nele Röse to this research.
Conflicts of Interest
The author declares no conflict of interest.
References
- Kakade, A.; Salama, E.-S.; Han, H.; Zheng, Y.; Kulshrestha, S.; Jalalah, M.; Harraz, F.A.; Alsareii, S.A.; Li, X. World eutrophic pollution of lake and river: Biotreatment potential and future perspectives. Environ. Technol. Innov. 2021, 23, 101604. [Google Scholar] [CrossRef]
- Bonilla, S.; Aguilera, A.; Aubriot, L.; Huszar, V.; Almanza, V.; Haakonsson, S.; Izaguirre, I.; O’Farrell, I.; Salazar, A.; Becker, V.; et al. Nutrients and not temperature are the key drivers for cyanobacterial biomass in the Americas. Harmful Algae 2023, 121, 102367. [Google Scholar] [CrossRef] [PubMed]
- García, J. Ecological engineering practice as a global strategy to prevent eutrophication and microalgae blooms. Ecol. Eng. 2021, 161, 106152. [Google Scholar] [CrossRef]
- Le Moal, M.; Gascuel-Odoux, C.; Ménesguen, A.; Souchon, Y.; Étrillard, C.; Levain, A.; Moatar, F.; Pannard, A.; Souchu, P.; Lefebvre, A.; et al. Eutrophication: A new wine in an old bottle? Sci. Total Environ. 2019, 651, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fink, G.; Alcamo, J.; Flörke, M.; Reder, K. Phosphorus loadings to the world’s largest lakes: Sources and trends. Glob. Biogeochem. Cycles 2018, 32, 617–634. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, Y.; Zhang, Y.Y. Plant Nutrients and Trace Elements from the Changjiang Watersheds to the East China Sea. In Ecological Continuum from the Changjiang (Yangtze River) Watersheds to the East China Sea Continental Margin. Estuaries of the World; Zhang, J., Ed.; Springer: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Saturday, A.; Kangume, S.; Bamwerinde, W. Content and dynamics of nutrients in the surface water of shallow Lake Mulehe in Kisoro District, South–western Uganda. Appl. Water Sci. 2023, 13, 150. [Google Scholar] [CrossRef]
- Carpenter, S.R. Eutrophication of aquatic ecosystems: Bistability and soil phosphorus. Proc. Natl. Acad. Sci. USA 2005, 102, 10002–10005. [Google Scholar] [CrossRef]
- Häder, D.-P.; Banaszak, A.T.; Villafañe, V.E.; Narvarte, M.A.; González, R.A.; Helbling, E.W. Anthropogenic pollution of aquatic ecosystems: Emerging problems with global implications. Sci. Total Environ. 2020, 713, 136586. [Google Scholar] [CrossRef]
- Liang, Z.; Xu, Y.; Qiu, Q.; Liu, Y.; Lu, W.; Wagner, T. A framework to develop joint nutrient criteria for lake eutrophication management in eutrophic lakes. J. Hydrol. 2021, 594, 125883. [Google Scholar] [CrossRef]
- Yang, X.E.; Wu, X.; Hao, H.L.; He, Z.L. Mechanisms and assessment of water eutrophication. J. Zhejiang Univ. Sci. B 2008, 9, 197–209. [Google Scholar] [CrossRef]
- Zhao, J.; Gao, Q.; Liu, Q.; Fu, G. Lake eutrophication recovery trajectories: Some recent findings and challenges ahead. Ecol. Indic. 2020, 110, 105878. [Google Scholar] [CrossRef]
- Zouiten, H.; Díaz, C.Á.; Gómez, A.G.; Cortezón, J.A.R.; Alba, J.G. An advanced tool for eutrophication modeling in coastal lagoons: Application to the Victoria lagoon in the north of Spain. Ecol. Model. 2013, 265, 99–113. [Google Scholar] [CrossRef]
- Edwards, K.F.; Thomas, M.K.; Klausmeier, C.A.; Litchman, E. Phytoplankton growth and the interaction of light and temperature: A synthesis at the species and community level. Limnol. Oceanogr. 2016, 61, 1232–1244. [Google Scholar] [CrossRef]
- Lee, K.-S.; Park, S.R.; Kim, Y.K. Effects of irradiance, temperature, and nutrients on growth dynamics of seagrasses: A review. J. Exp. Mar. Biol. Ecol. 2007, 350, 144–175. [Google Scholar] [CrossRef]
- Tasnim, B.; Fang, X.; Hayworth, J.S.; Tian, D. Simulating Nutrients and Phytoplankton Dynamics in Lakes: Model Development and Applications. Water 2021, 13, 2088. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; El-Naggar, A.H.; Alaswad, S.O.; Elsayed, M.; Li, M.; Li, W. Enhancement of biodiesel yield from a halophilic green microalga isolated under extreme hypersaline conditions through stepwise salinity adaptation strategy. Bioresour. Technol. 2020, 310, 123462. [Google Scholar] [CrossRef]
- Glibert, P.M. Eutrophication, harmful algae and biodiversity—Challenging paradigms in a world of complex nutrient changes. Mar. Pollut. Bull. 2017, 124, 591–606. [Google Scholar] [CrossRef]
- Glibert, P.M.; Wilkerson, F.; Dugdale, R.C.; Parker, A.E.; Alexander, J.A.; Blaser, S.; Murasko, S. Microbial communities from San Francisco Bay Delta respond differently to oxidized and reduced nitrogen substrates—Even under conditions that would otherwise suggest nitrogen sufficiency. Front. Mar. Sci. 2014, 1, 17. [Google Scholar] [CrossRef]
- Baldia, S.F.; Evangelista, A.D.; Aralar, E.V.; Santiago, A.E. Nitrogen and phosphorus utilization in the cyanobacterium Microcystis aeruginosa isolated from Laguna de Bay, Philippines. J. Appl. Phycol. 2007, 19, 607–613. [Google Scholar] [CrossRef]
- Lehman, E.M. Seasonal occurrence and toxicity of Microcystis in impoundments of the Huron River, Michigan, USA. Water Res. 2007, 41, 795–802. [Google Scholar] [CrossRef]
- Ha, J.H.; Hidaka, T.; Tsuno, H. Quantification of toxic Microcystis and evaluation of its dominance ratio in blooms using real-time PCR. Environ. Sci. Technol. 2009, 43, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Gobler, C.J.; Davis, T.W.; Coyne, K.J.; Boyer, G.L. The interactive influences of nutrient loading and zooplankton grazing on the growth and toxicity of cyanobacteria blooms in a eutrophic lake. Harmful Algae 2007, 6, 119–133. [Google Scholar] [CrossRef]
- Moisander, P.H.; Ochiai, M.; Lincoff, A. Nutrient limitation of Microcystis aeruginosa in northern California Klamath River reservoirs. Harmful Algae 2009, 8, 889–897. [Google Scholar] [CrossRef]
- Davis, T.W.; Harke, M.J.; Marcoval, M.A.; Goleski, J.; Orano-Dawson, C.; Berry, D.L.; Gobler, C.J. Effects of nitrogenous compounds and phosphorus on the growth of toxic and non-toxic strains of Microcystis during cyanobacterial blooms. Aquat. Microb. Ecol. 2010, 61, 149–162. [Google Scholar] [CrossRef]
- López-Ballesteros, A.; Trolle, D.; Srinivasan, R.; Senent-Aparicio, J. Assessing the effectiveness of potential best management practices for science-informed decision support at the watershed scale: The case of the Mar Menor coastal lagoon, Spain. Sci. Total Environ. 2023, 859, 160144. [Google Scholar] [CrossRef] [PubMed]
- Maté-Sánchez-Val, M.; Aparicio-Serrano, G. The impact of marine pollution on the probability of business failure: A case study of the Mar Menor lagoon. J. Environ. Manag. 2023, 332, 117381. [Google Scholar] [CrossRef]
- Conesa, H.M.; Jiménez-Cárceles, F.J. The Mar Menor lagoon (SE Spain): A singular natural ecosystem threatened by human activities. Mar. Pollut. Bull. 2007, 54, 839–849. [Google Scholar] [CrossRef]
- Lloret, J.; Marín, A.; Marín-Guirao, L. Is coastal lagoon eutrophication likely to be aggravated by global climate change? Estuar. Coast. Shelf Sci. 2008, 78, 403–412. [Google Scholar] [CrossRef]
- Gilabert, J. Seasonal plankton dynamics in a Mediterranean hypersaline coastal lagoon: The Mar Menor. J. Plankton Res. 2001, 23, 207–218. [Google Scholar] [CrossRef]
- Vázquez-Luis, M.; Álvarez, E.; Barrajón, A.; García-March, J.R.; Grau, A.; Hendriks, I.E.; Jiménez, S.; Kersting, D.; Moreno, D.; Pérez, M.; et al. S.O.S. Pinna nobilis: A Mass Mortality Event in Western Mediterranean Sea. Front. Mar. Sci. 2017, 4, 220. [Google Scholar] [CrossRef]
- Cabanellas-Reboredo, M.; Vázquez-Luis, M.; Mourre, B.; Álvarez, E.; Deudero, S.; Amores, Á.; Addis, P.; Ballesteros, E.; Barrajón, A.; Coppa, S.; et al. Tracking a mass mortality outbreak of pen shell Pinna nobilis populations: A collaborative effort of scientists and citizens. Sci. Rep. 2019, 9, 13355. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Melendreras, E.; Gomariz-Castillo, F.; Alonso-Sarría, F.; Martín, F.J.G.; Murcia, J.; Canales-Cáceres, R.; Esplá, A.A.R.; Barberá, C.; Giménez-Casalduero, F. The relict population of Pinna nobilis in the Mar Menor is facing an uncertain future. Mar. Pollut. Bull. 2022, 185, 114376. [Google Scholar] [CrossRef] [PubMed]
- Castejón-Porcel, G.; Espín-Sánchez, D.; Ruiz-Álvarez, V.; García-Marín, R.; Moreno-Muñoz, D. Runoff water as a resource in the Campo de Cartagena (Region of Murcia): Current possibilities for use and benefits. Water 2018, 10, 456. [Google Scholar] [CrossRef]
- Medidas Para la Recuperación del Acuífero del Campo de Cartagena y Reducción de los Aportes de Nitrógeno al Mar Menor. Instituto de Ingeniería del Agua y Medio Ambiente. Universitat Politècnica de València. IIAMA-UPV. 2020. Available online: https://www.chsegura.es/export/sites/chs/descargas/cuenca/marmenor/docsdescarga/InformeUPVSeguraCampoCartagena_20210914.pdf (accessed on 1 June 2023).
- Alcolea, A.; Contreras, S.; Huninkb, J.E.; García-Aróstegui, J.L.; Jiménez-Martínez, J. Hydrogeological modelling for the watershed management of the Mar Menor coastal lagoon (Spain). Sci. Total Environ. 2019, 663, 901–914. [Google Scholar] [CrossRef]
- Servicios de Consultoría y Asistencia Técnica para la Simulación Bajo Distintos Escenarios y Mediante Modelo Hidrológico, de la Evolución del Contenido en Nitratos de la Masa de Agua Subterránea 070.052 Campo de Cartagena. (TTMM: Varios. Provincias: Murcia y Alicante). Instituto de Ingeniería del Agua y Medio Ambiente. Universitat Politècnica de València. IIAMA-UPV. 2021. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwiS9IrJw--BAxWomlYBHdGHBRAQFnoECAgQAQ&url=https%3A%2F%2Fwww.chsegura.es%2Fexport%2Fsites%2Fchs%2Fdescargas%2Fcuenca%2Fmarmenor%2Fdocsdescarga%2FInformeUPVSeguraCampoCartagena_20210914.pdf&usg=AOvVaw3Odm17Z-nHQ-7gnh_J1s-p&opi=89978449 (accessed on 1 June 2023).
- Modelo de Flujo. Acuífero Cuaternario del Campo de Cartagena: “Cuantificación, Control de la Calidad y Seguimiento Piezométrico de la Descarga de Agua Subterránea del Acuífero Cuaternario del Campo de Cartagena al Mar Menor. 2021 Tragsatec. Available online: https://www.chsegura.es/static/marmenor/DescargasMarMenorCuaternarioCampoCartagena.zip (accessed on 1 June 2023).
- Pérez-Martín, M.; Benedito-Castillo, S. Fertigation to Recover Nitrate-Polluted Aquifer and Improve a Long Time Eutrophicated Lake, Spain. Sci. Total Environ. 2023, 894, 165020. [Google Scholar] [CrossRef]
- López-Andreu, F.J.; López-Morales, J.A.; Juárez, J.F.A.; Alcaraz, R.; Hernández, M.D.; Erena, M.; Domínguez-Gómez, J.A.; Galiano, S.G. Monitoring System of the Mar Menor Coastal Lagoon (Spain) and Its Watershed Basin Using the Integration of Massive Heterogeneous Data. Sensors 2022, 22, 6507. [Google Scholar] [CrossRef]
- Erena, M.; Domínguez, J.A.; Aguado, F.; Soria, J.; Galiano, G. Monitoring Coastal Lagoon Water Quality through Remote Sensing: The Mar Menor as a Case Study. Water 2019, 11, 1468. [Google Scholar] [CrossRef]
- Angel, P.-R.; Marcos, C.; Gilabert, J. The Ecology of the Mar Menor Coastal Lagoon: A Fast Changing Ecosystem under Human Pressure. Coastal Lagoons. In Ecosystem Processes and Modeling for Sustainable Use and Development; Gönenç, I.E., Wolflin, J.P., Eds.; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- García-Oliva, M.; Marcos, C.; Umgiesser, G.; McKiver, W.; Ghezzo, M.; De Pascalis, F.; Pérez-Ruzafa, A. Modelling the impact of dredging inlets on the salinity and temperature regimes in coastal lagoons. Ocean Coast. Manag. 2019, 180, 104913. [Google Scholar] [CrossRef]
- Ruiz, J.M.; Albentsosa, M.; Aldeguer, B.; Alvarez-Rogel, J.; Yebra, L. Informe de Evolución y Estado Actual del Mar Menor en Relación al Proceso de Eutrofización y Sus Causas 2020; Technical Report Instituto Español de Oceanografía (IEO); Spanish Institute of Oceanography: Madrid, Spain, 2020; p. 165. Available online: https://www.miteco.gob.es/es/prensa/informe-ieo-mar-menor.aspx (accessed on 11 December 2022).
- Bernardeau-Esteller, J.; Sandoval-Gil, J.M.; Belando, M.D.; Ramos-Segura, A.; García-Muñoz, R.; Marín-Guirao, L.; Ruiz, J.M. The Role of Cymodocea nodosa and Caulerpa prolifera Meadows as Nitrogen Sinks in Temperate Coastal Lagoons. Diversity 2023, 15, 172. [Google Scholar] [CrossRef]
- Arévalo, L. El Mar Menor como sistema forzado por el Mediterráneo. Boletín Instituo Español Oceanogr. 1988, 5, 61–95. [Google Scholar]
- IEO. Estudio Oceanográfico Integral de alta Resolución de la Laguna Costera del Mar Menor, Murcia. Primer Informe Técnico: Campaña MM1116 (Otoño). 2018. Available online: https://www.ulpgc.es/sites/default/files/ArchivosULPGC/noticia/2017/Feb/estudio-ieo-mar-menor.pdf (accessed on 1 June 2023).
- Martínez-Alvarez, V.; Gallego-Elvira, B.; Maestre-Valero, J.; Tanguy, M. Simultaneous solution for water, heat and salt balances in a Mediterranean coastal lagoon (Mar Menor, Spain). Estuar. Coast. Shelf Sci. 2011, 91, 250–261. [Google Scholar] [CrossRef]
- López-Castejón, F. Caracterización de la Hidrodinámica del Mar Menor y los Flujos de Intercambio con el Mediterráneo Mediante Datos in situ y Modelado Numérico. Ph.D. Thesis, Technical University of Cartagena, Cartagena, Spain, 2017. [Google Scholar] [CrossRef]
- Senent-Aparicio, J.; López-Ballesteros, A.; Nielsen, A.; Trolle, D. A holistic approach for determining the hydrology of the mar menor coastal lagoon by combining hydrological & hydrodynamic models. J. Hydrol. 2021, 603, 127150. [Google Scholar] [CrossRef]
- Torrente, M.D.B.; Ruiz, J.M.; Muñoz, R.G.; Segura, A.R.; Esteller, J.B.; Casero, J.J.; Guirao, L.M.; Moreno, P.G.; Navarro, I.F.; Nuez, E.F.; et al. Collapse of macrophytic communities in a eutrophicated coastal lagoon. Front. Mar. Sci. 2019. [Google Scholar] [CrossRef]
- IEO. Informe de Síntesis Sobre el Estado Actual del Mar Menor y sus Causas en Relación a los Contenidos de Nutrients. 2019. Available online: https://www.ieo.es/documents/10640/7708614/1_Informe+sobre+la+evolucion+del+Mar+Menor+y+sus+causas_noviembre+2019+final+B.pdf/564902d9-d41e-4e45-b730-9e561411c977 (accessed on 1 June 2023).
- UPCT (Universidad Politécnica de Cartagena). Servidor de Datos Científicos del Mar Menor (SDC). 2023. Available online: https://marmenor.upct.es/ (accessed on 10 February 2023).
- Ruiz, J.M.; Clemente-Navarro, P.; Mercado, J.M.; Fraile-Nuez, E.; Albentosa, M.; Marín-Guirao, L.; Santos, J. Nuevo Evento de Mortalidad Masiva de Organismos Marinos en el Mar Menor: Contexto y Factores. Instituto Español de Oceanografía IEO. September 2021. Available online: https://www.ieo.es/documents/10640/7708614/3_informe+IEO_FINAL2_MGB_060921.pdf/cb4859a4-4da4-4855-82ec-8fb4167790c3 (accessed on 1 June 2023).
- Royal Decrete 817/2015, de 11 de Septiembre, por el que se Establecen los Criterios de Seguimiento y Evaluación del Estado de las Aguas Superficiales y las Normas de Calidad Ambiental. Available online: https://www.boe.es/buscar/pdf/2015/BOE-A-2015-9806-consolidado.pdf (accessed on 1 June 2023).
- Pérez-Martín, M.A.; Estrela, T.; Del-Amo, P. Measures required to reach the nitrate objectives in groundwater based on a long-term nitrate model for large river basins (Júcar, Spain). Sci. Total Environ. 2016, 566–567, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martín, M.A.; Estrela, T.; Andreu, J.; Ferrer, J. Modeling Water Resources and River-Aquifer Interaction in the Júcar River Basin, Spain. Water Resour. Manag. 2014, 28, 4337–4358. [Google Scholar] [CrossRef]
- Mapama. “Balance del Nitrógeno en la Agricultura Española, 2019”. Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente. 2021. Available online: https://www.mapa.gob.es/es/agricultura/temas/medios-de-produccion/01_modelnationallevelncpsa_ae_110n_spain_junio2021_tcm30-620003.xlsx (accessed on 1 June 2023).
- Jensen, J.P.; Pedersen, A.R.; Jeppesen, E.; Søndergaard, M. An empirical model describing the seasonal dynamics of phosphorus in 16 shallow eutrophic lakes after external loading reduction. Limnol. Oceanogr. 2006, 51, 791–800. [Google Scholar] [CrossRef]
- Savva, I.; Bennett, S.; Roca, G.; Jordà, G.; Marbà, N. Thermal tolerance of Mediterranean marine macrophytes: Vulnerability to global warming. Ecol. Evol. 2018, 8, 12032–12043. [Google Scholar] [CrossRef]
- Caraco, N.; Cole, J.; Likens, G.E. A comparison of phosphorus immobilization in sediments of freshwater and coastal marine systems. Biogeochemistry 1990, 9, 277–290. [Google Scholar] [CrossRef]
- Bennett, S.; Vaquer-Sunyer, R.; Jorda, G.; Forteza, M.; Roca, G.; Marbà, N. Thermal Performance of Seaweeds and Seagrasses Across a Regional Climate Gradient. Front. Mar. Sci. 2022, 9, 733315. [Google Scholar] [CrossRef]
- Marbà, N.; Jordà, G.; Bennett, S.; Duarte, C.M. Seagrass Thermal Limits and Vulnerability to Future Warming. Front. Mar. Sci. 2022, 9, 860826. [Google Scholar] [CrossRef]
- Tsioli, S.; Orfanidis, S.; Papathanasiou, V.; Katsaros, C.; Exadactylos, A. Effects of salinity and temperature on the performance of Cymodocea nodosa and Ruppia cirrhosa: A medium-term laboratory study. Bot. Mar. 2018, 62, 97–108. [Google Scholar] [CrossRef]
- Servicio y desarrollo de la red de control y vigilancia de la calidad de las aguas litorales para el seguimiento del estado de las masas de agua costeras en la Región de Murcia. 2021 Ref: DT2021/014 2021 Servicio de planificación y evaluación ambiental (CARM). Informe Final Taxon.
- Wild, A. (Ed.) Russell’s Soil Conditions and Plant Growth; Longman Scientific and Technical: Harlow, UK, 1988. [Google Scholar]
- McGlathery, K.; Sundbäck, K.; Anderson, I. Eutrophication in shallow coastal bays and lagoons: The role of plants in the coastal filter. Mar. Ecol. Prog. Ser. 2007, 348, 1–18. [Google Scholar] [CrossRef]
- Jilbert, T.; Couture, R.-M.; Huser, B.J.; Salonen, K. Preface: Restoration of eutrophic lakes: Current practices and future challenges. Hydrobiologia 2020, 847, 4343–4357. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Campillo, S.; Fernández-Palacios, J.M.; García-Lacunza, A.; García-Oliva, M.; Ibañez, H.; Navarro-Martínez, P.C.; Pérez-Marcos, M.; Pérez-Ruzafa, I.M.; Quispe-Becerra, J.I.; et al. Long-Term Dynamic in Nutrients, Chlorophyll a, and Water Quality Parameters in a Coastal Lagoon During a Process of Eutrophication for Decades, a Sudden Break and a Relatively Rapid Recovery. Front. Mar. Sci. 2019, 6, 26. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).