
Effect of Simulated Eutrophication of Peatlands on the Microbiome of Utricularia vulgaris L.


Abstract
1. Introduction
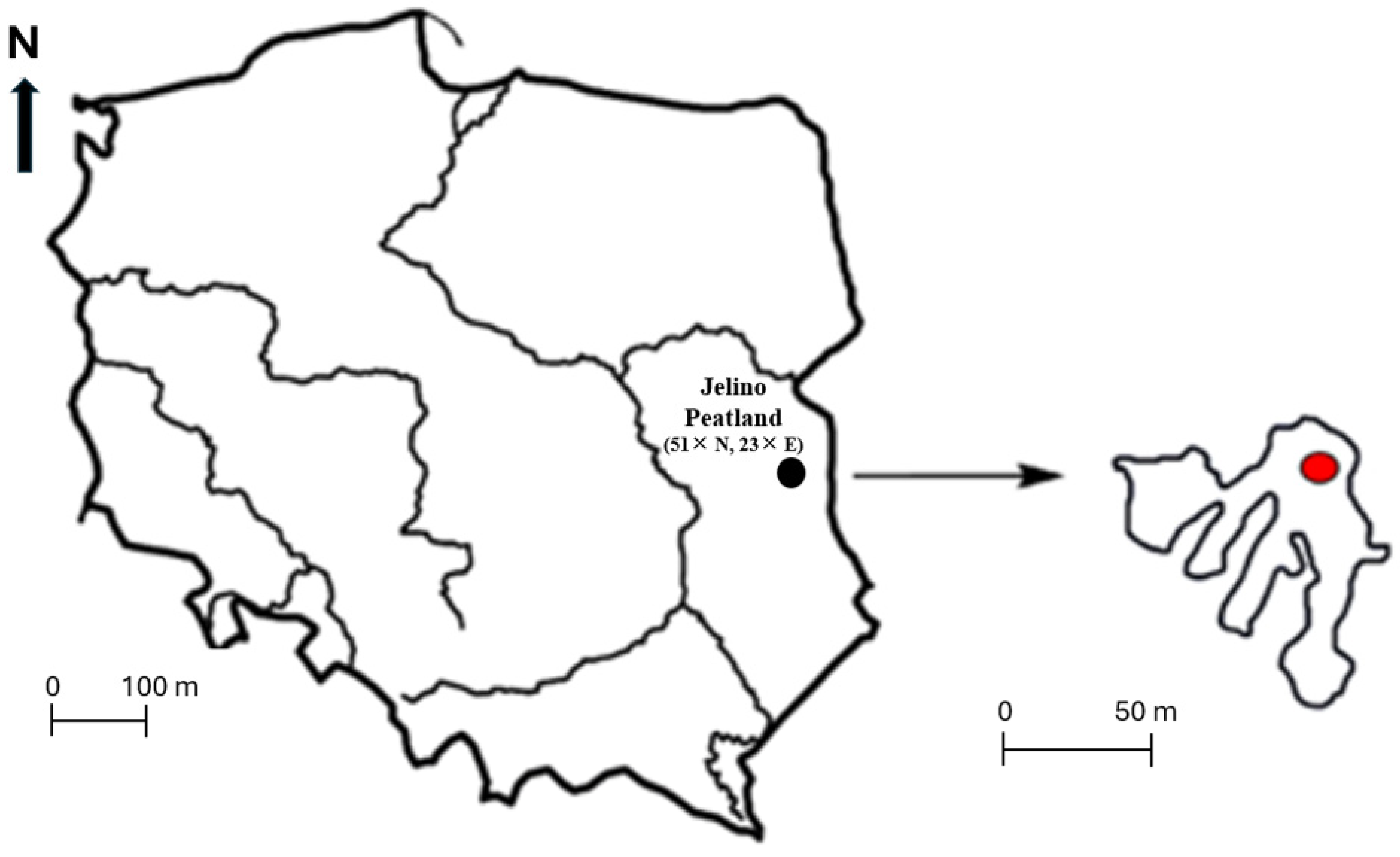
2. Materials and Methods
2.1. Experiment and Laboratory Analyses
2.2. Statistical Analysis
3. Results
3.1. Environmental Variables
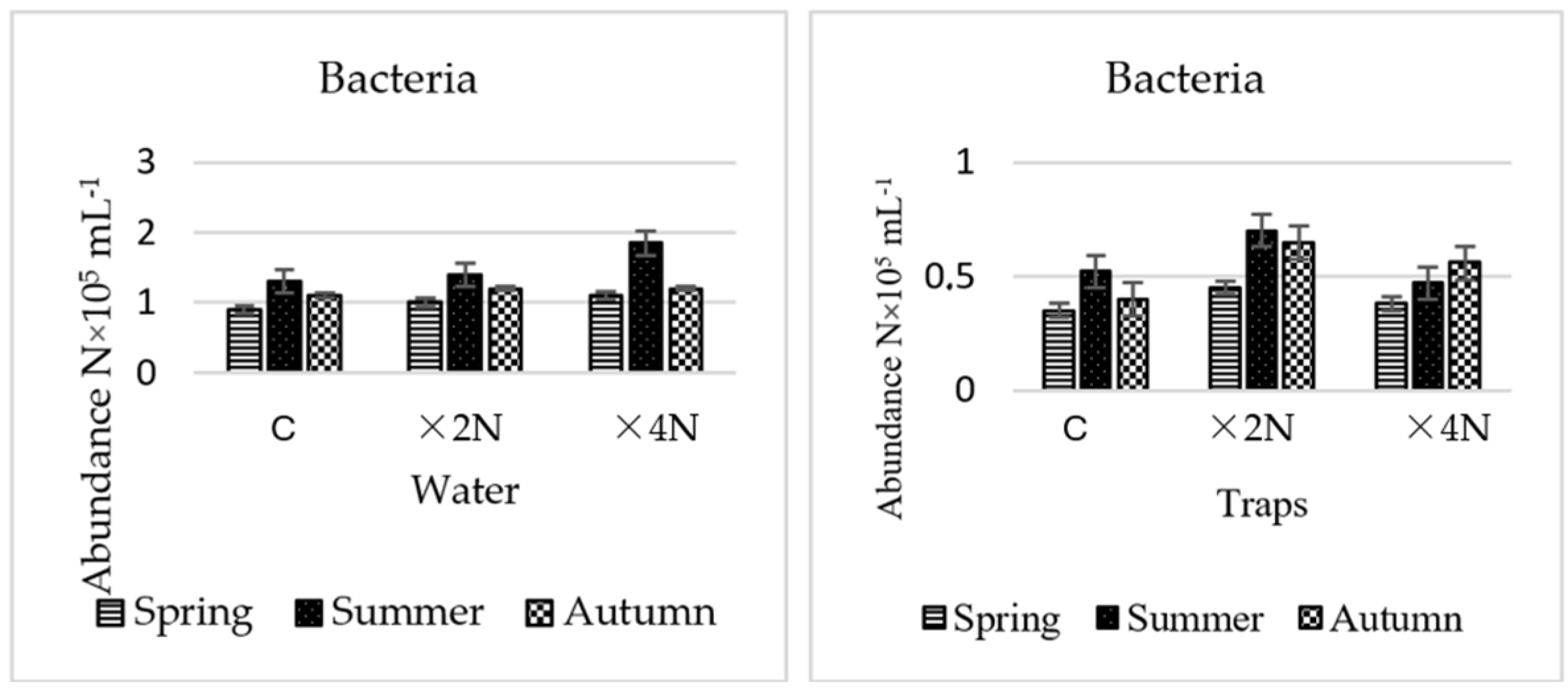
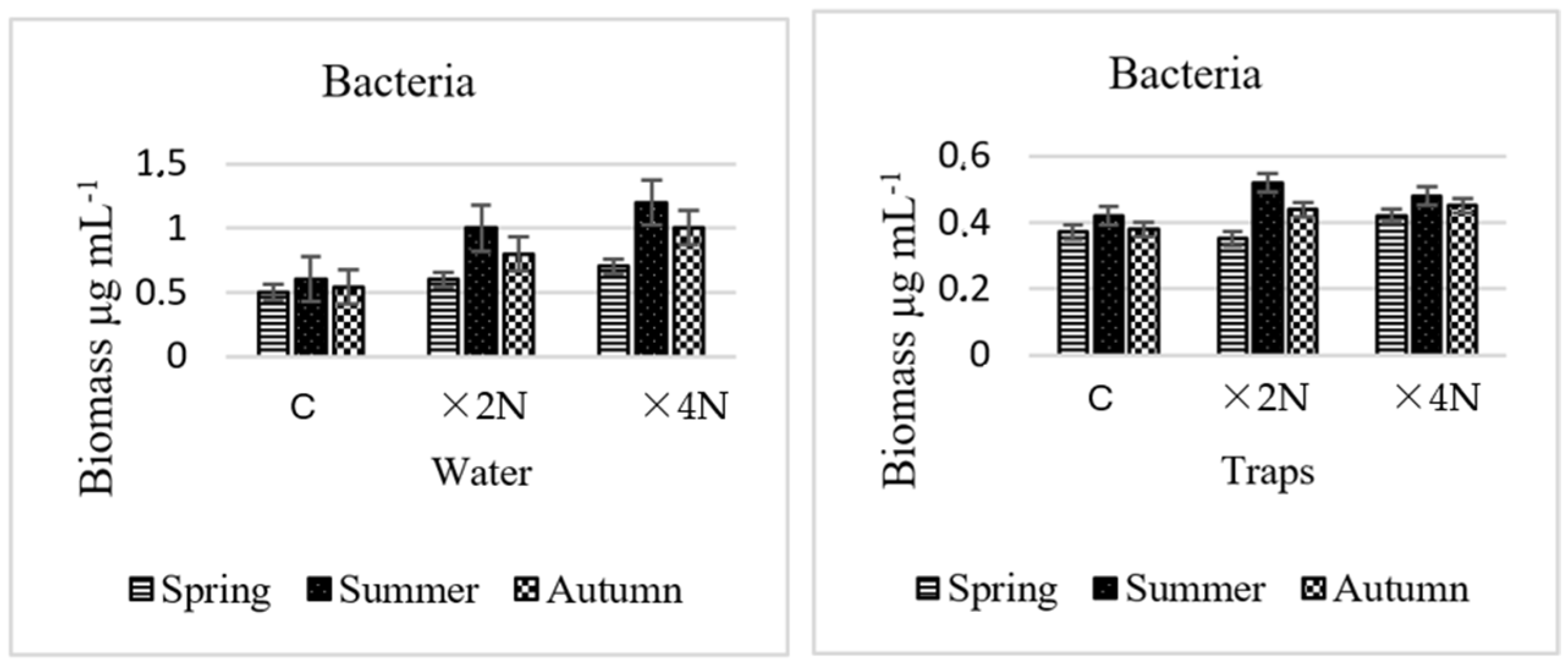
3.2. Abundance and Biomass of Bacteria
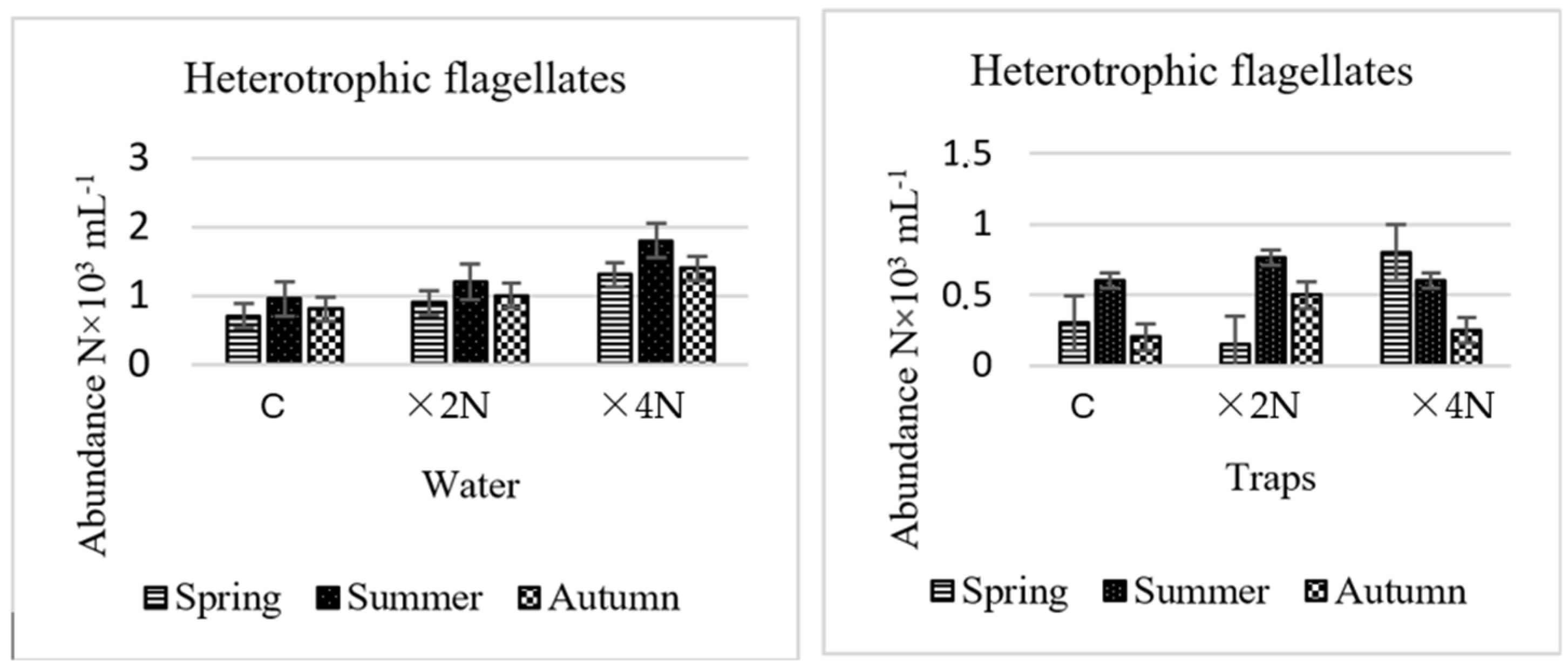
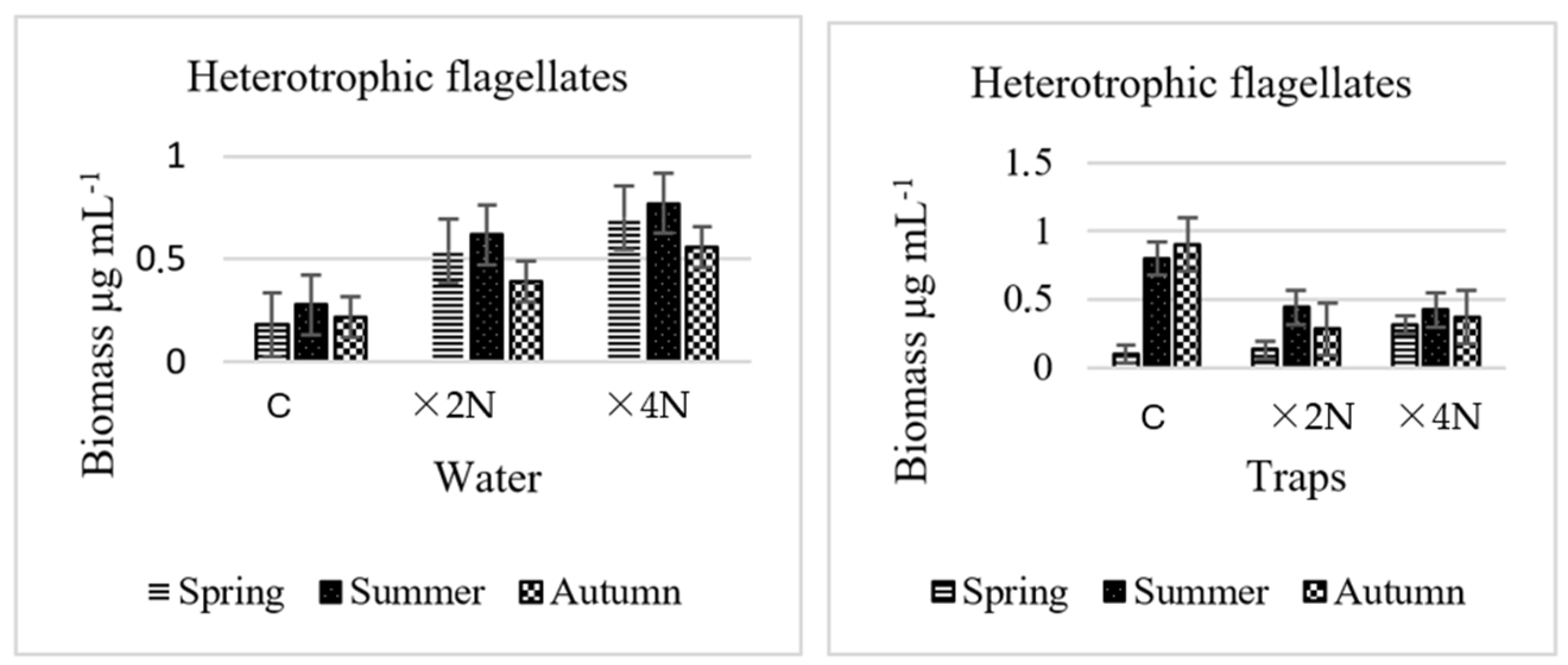
3.3. Abundance and Biomass of Heterotrophic Flagellates
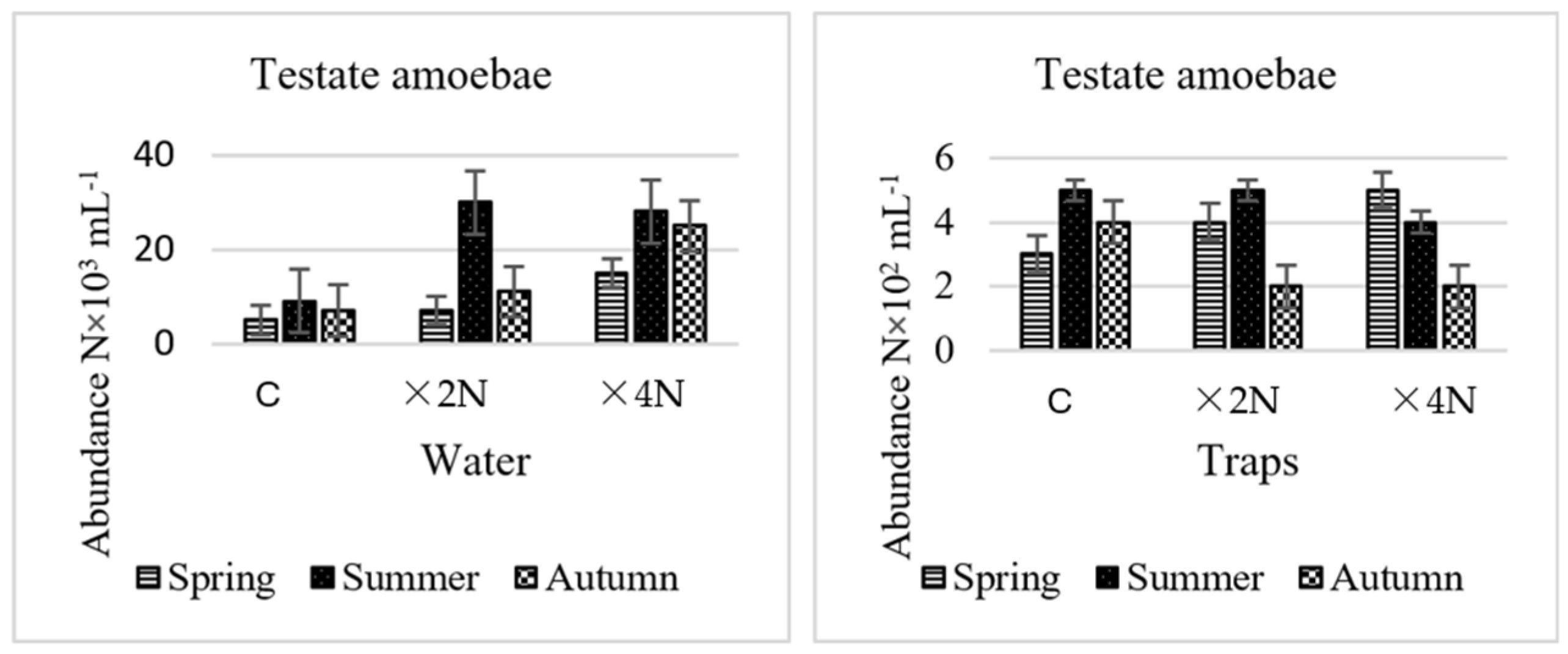
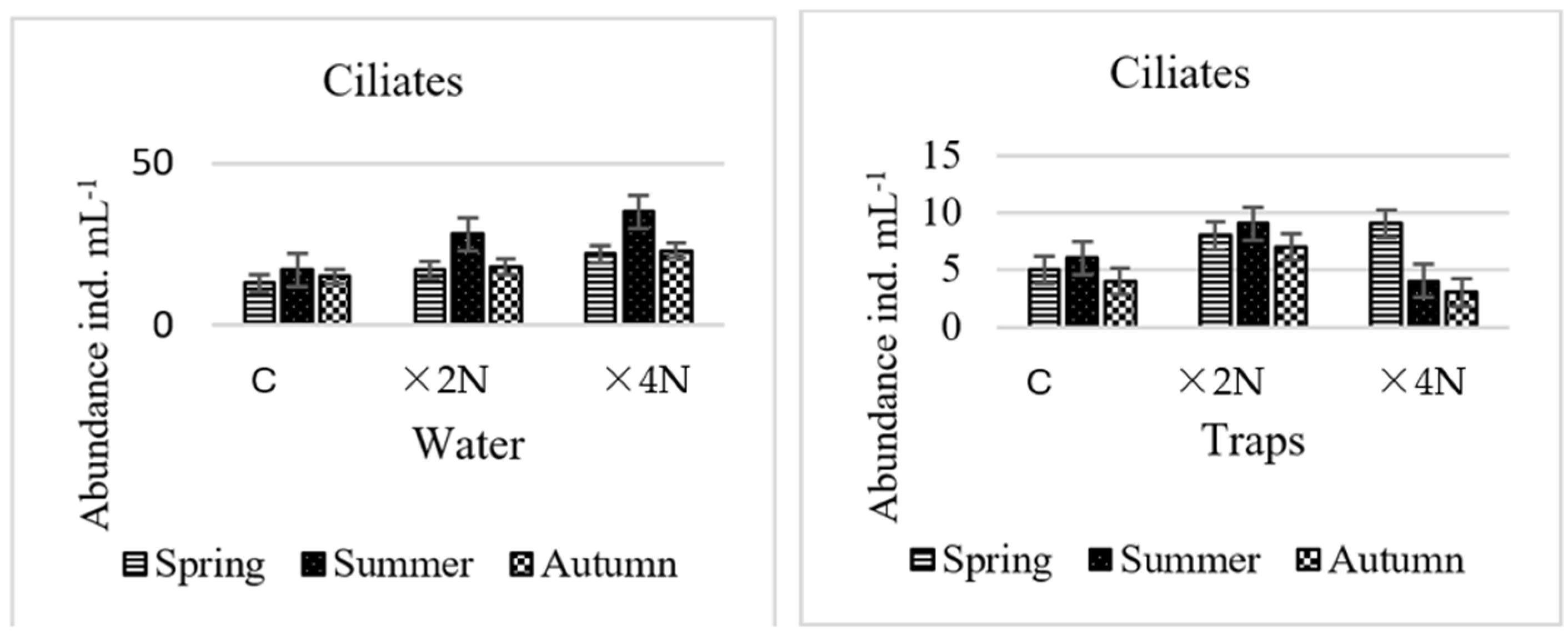
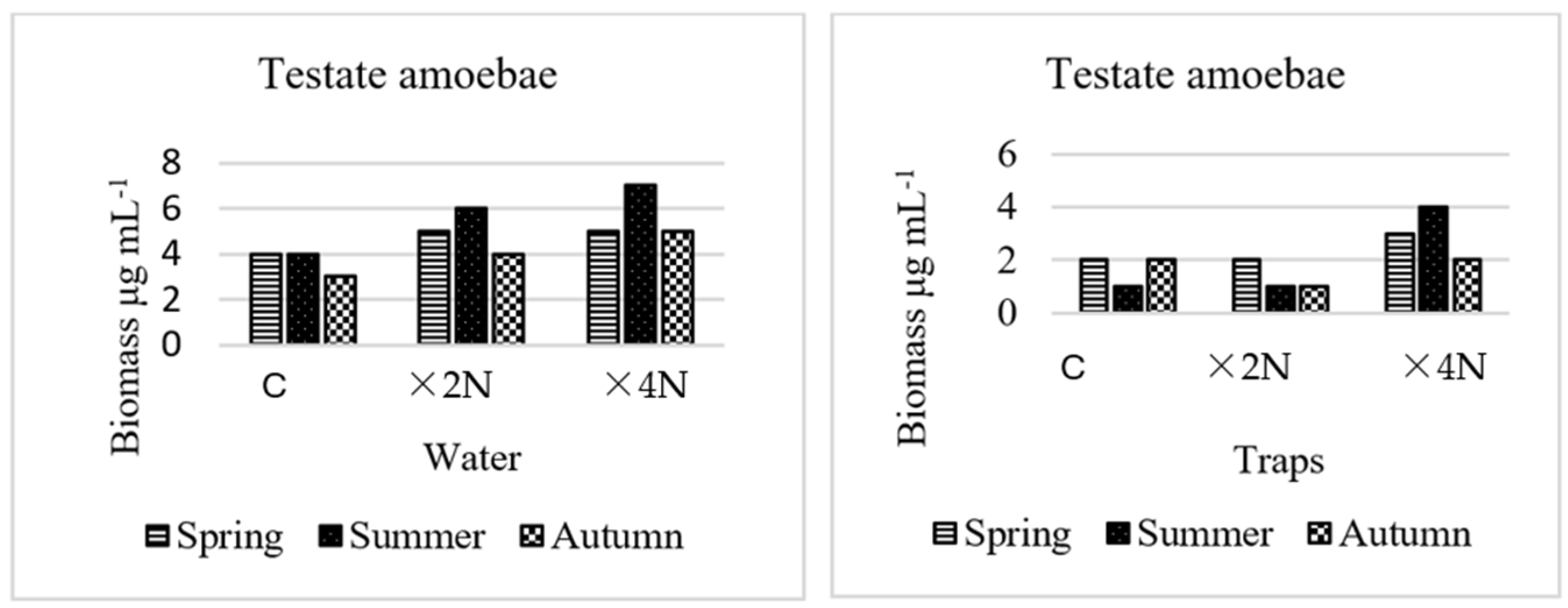
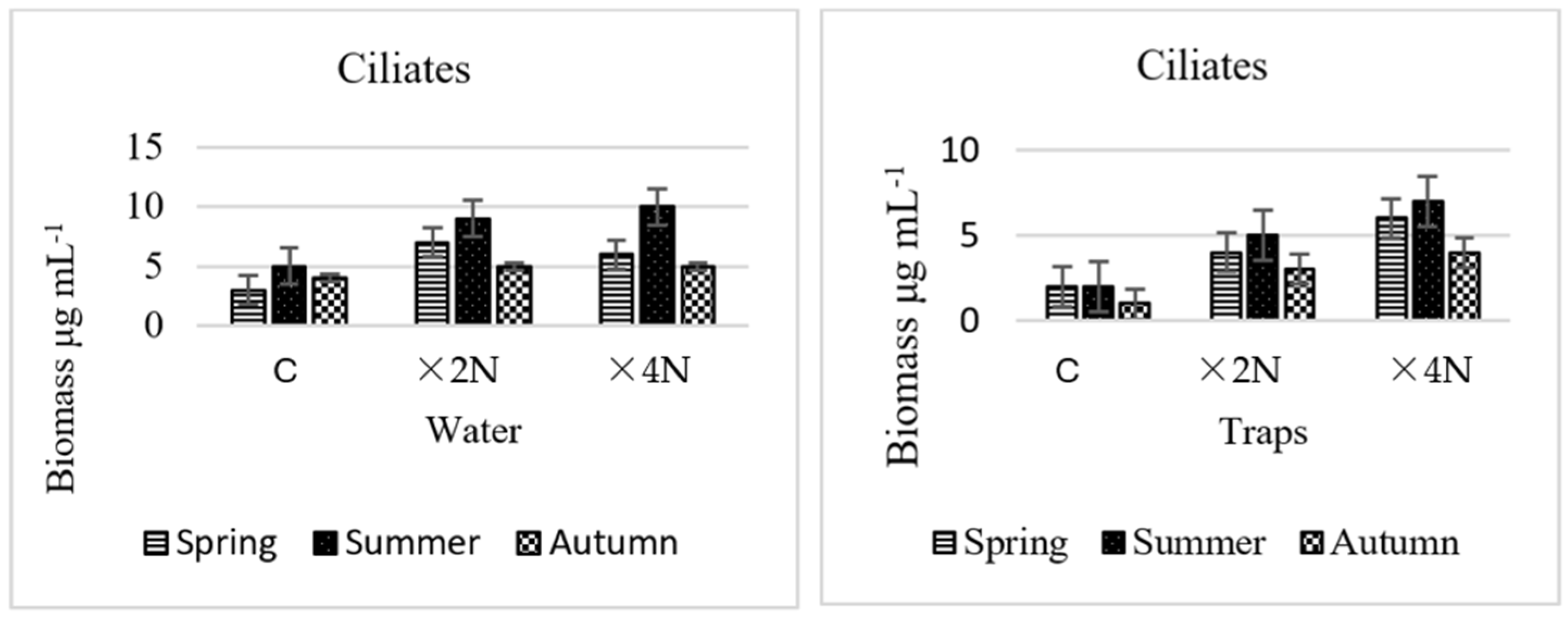
3.4. Abundance and Biomass of Testate Amoebae and Ciliates
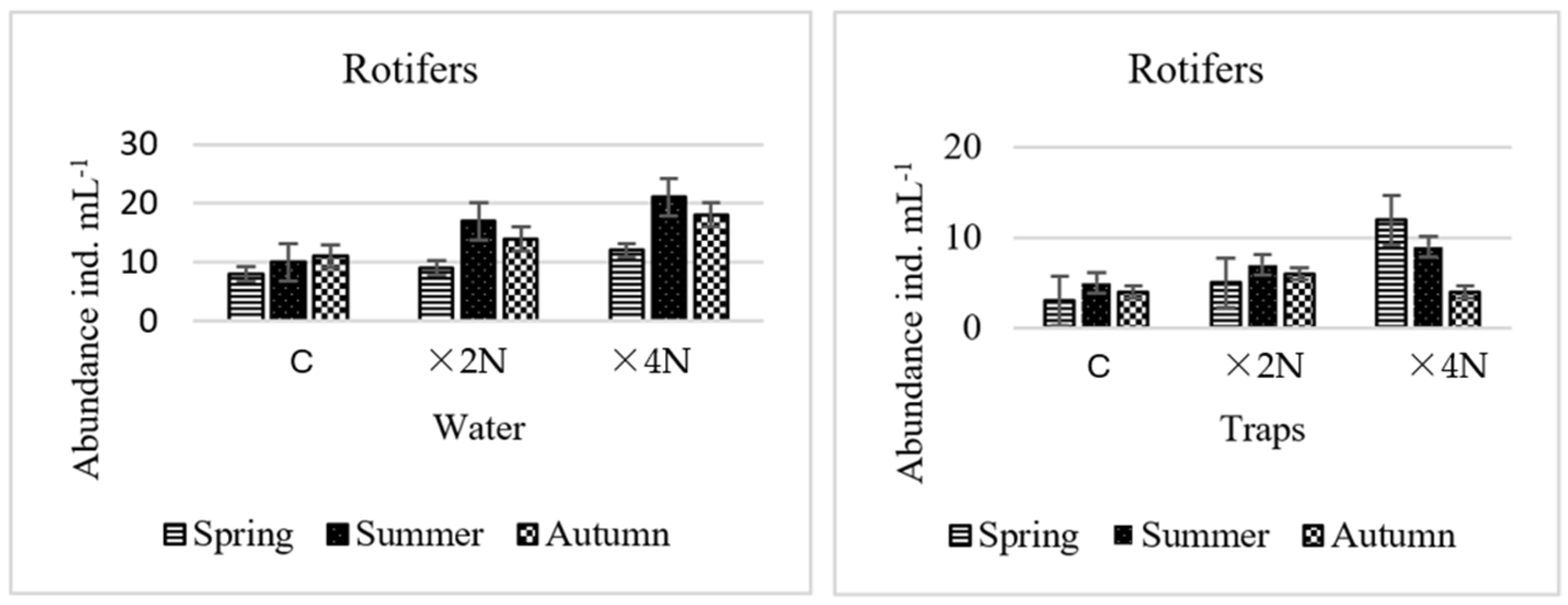
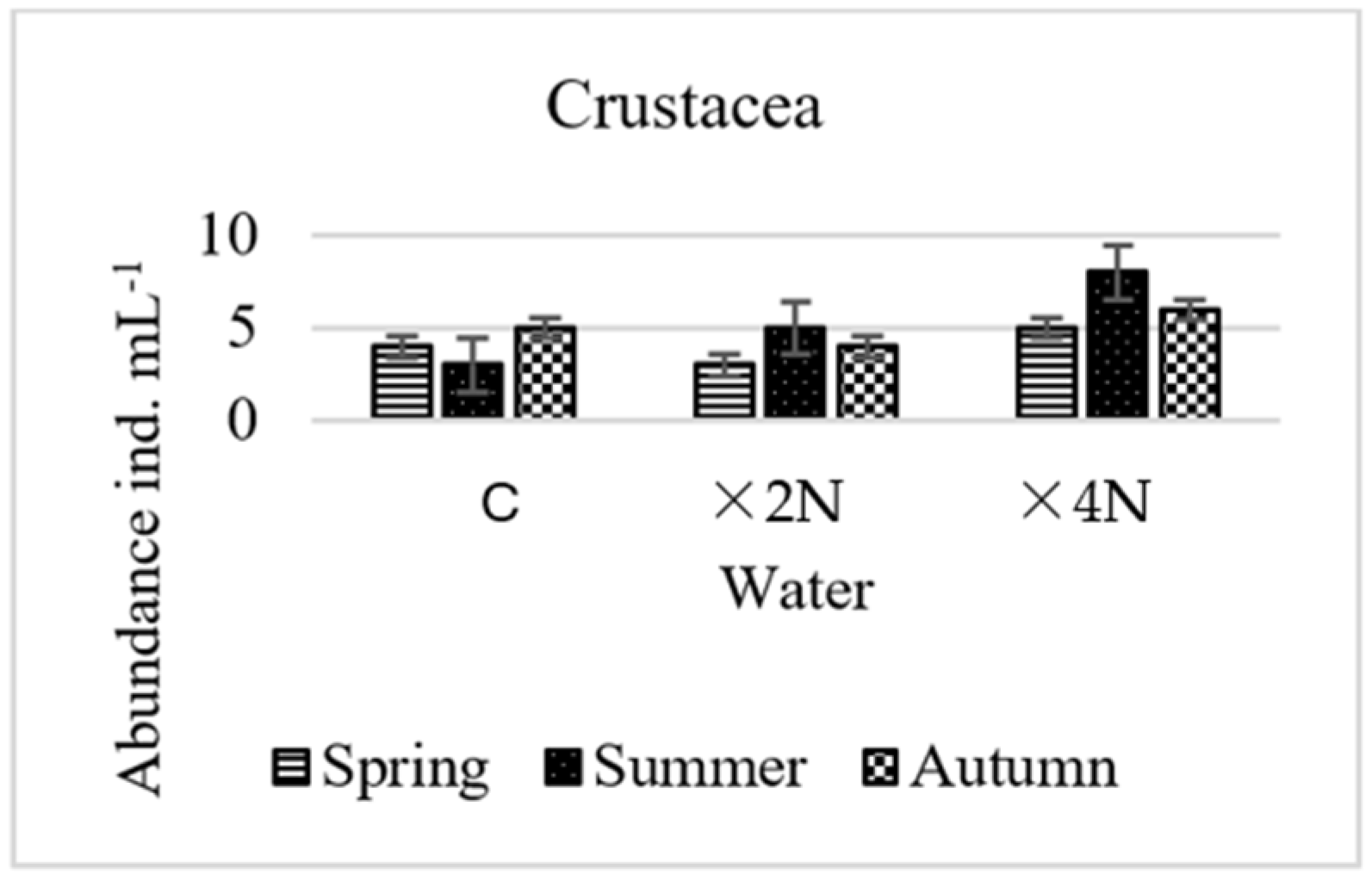
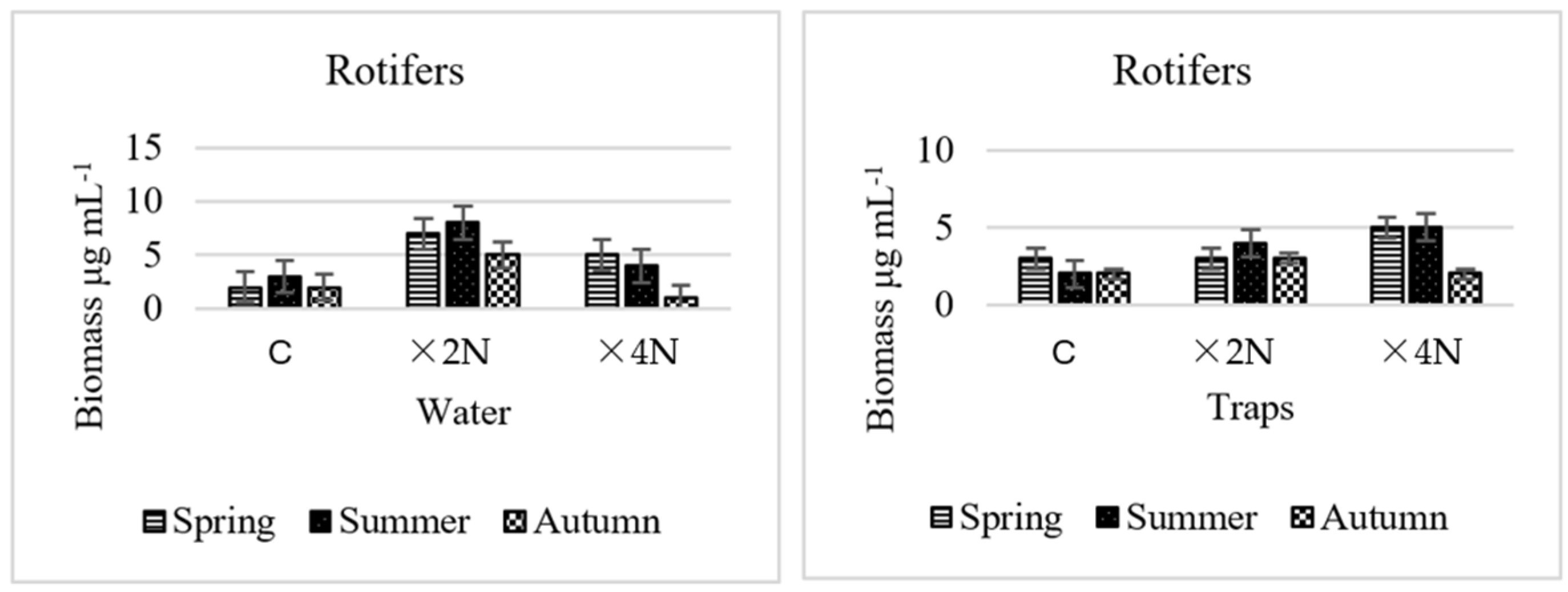
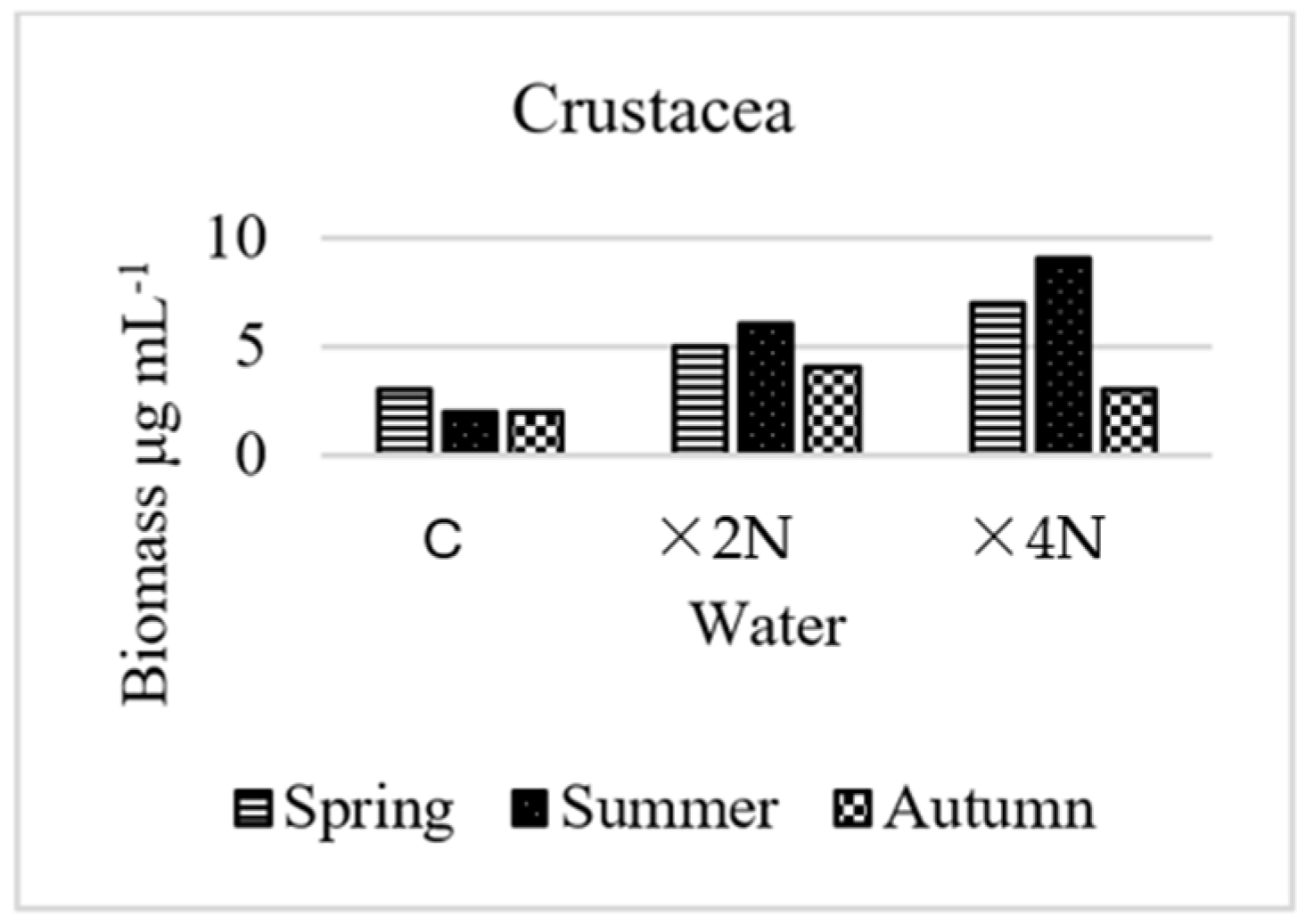
3.5. Abundance and Biomass of Small Metazoa
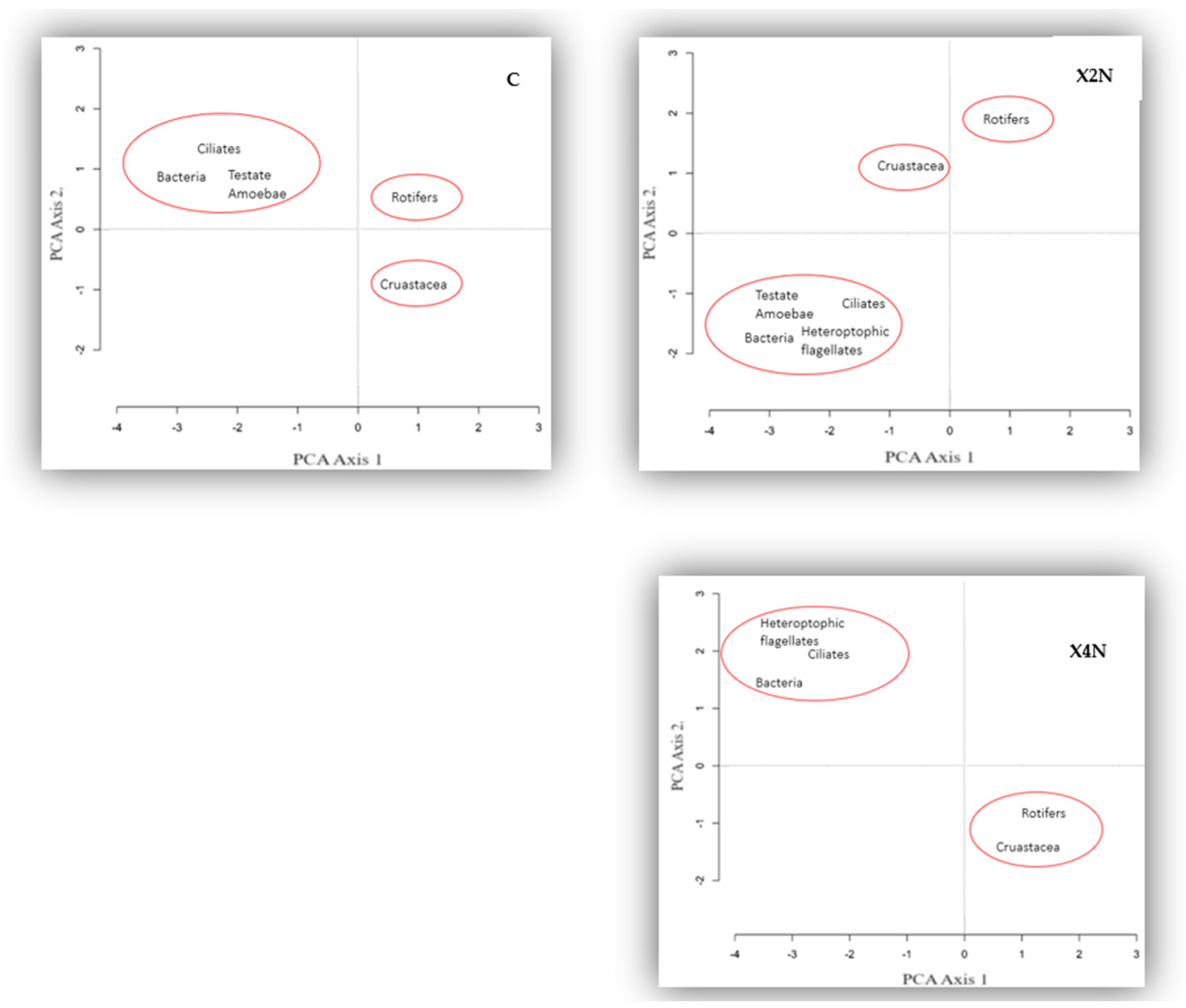
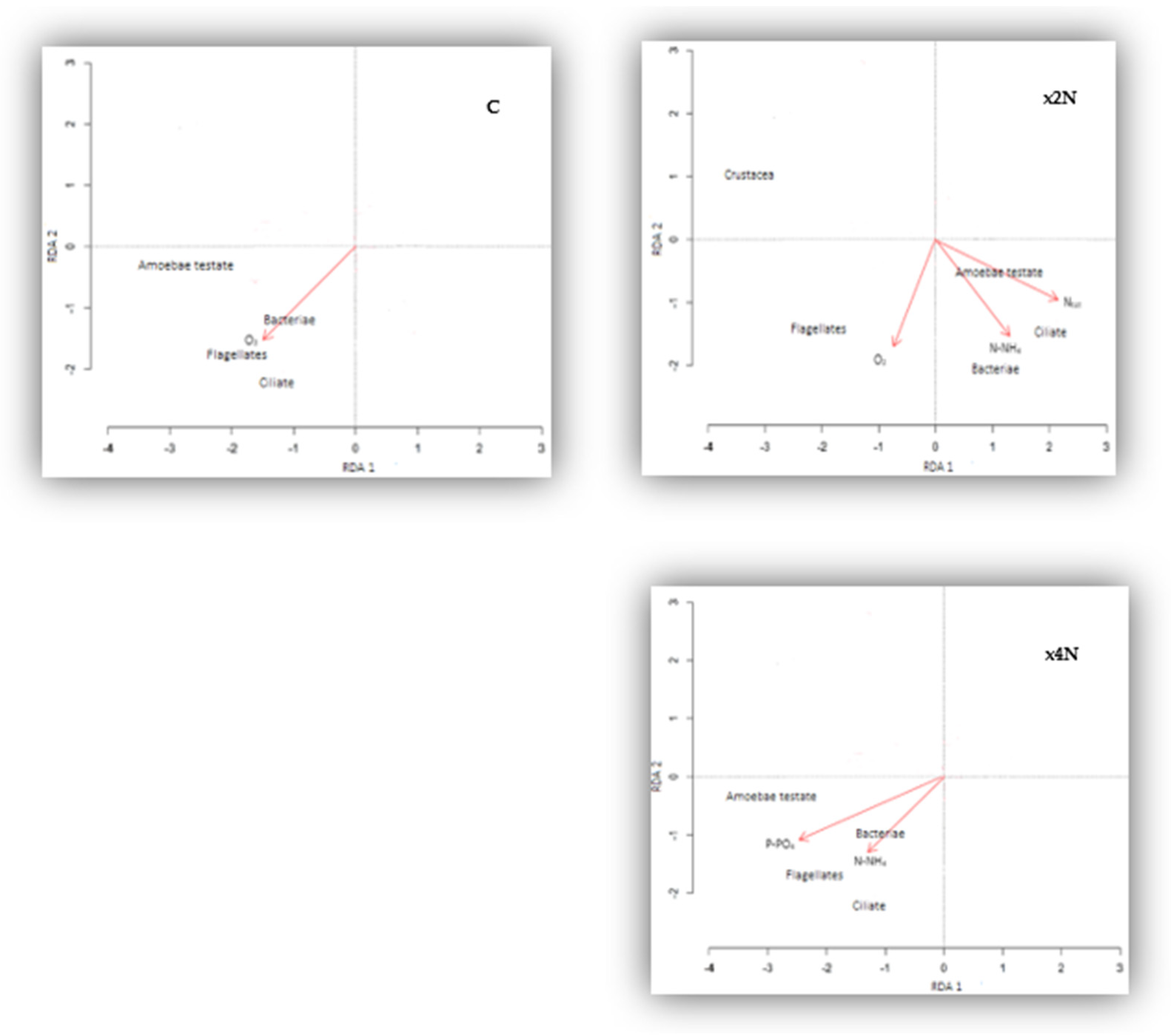
3.6. PCA, Redundancy Analysis (RDA), and Correlations
3.7. Correlations between Food Web Components
4. Discussion
4.1. Changes in Physical and Chemical Parameters Accompanying Eutrophication Processes
4.2. Effect of Biogenic Compounds on Heterotrophic Bacteria and Heterotrophic Flagellates
4.3. Effect of Biogenic Compounds on Testate Amoebae and Ciliates
4.4. Effect of Biogenic Compounds on Small Metazoa
4.5. Effect of Biogenic Substances on Food Web Structure
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ahmad, S.; Liu, H.; Günther, A.; Couwenberg, J.; Lennartz, B. Long-term rewetting of degraded peatlands restores hydrological buffer function. Sci. Total. Environ. 2020, 749, 141571. [Google Scholar] [CrossRef] [PubMed]
- Jiroušek, M.; Poulíčková, A.; Kintrová, K.; Opravilová, V.; Hájková, P.; Rybníček, K.; Kočí, M.; Bergová, K.; Hnilica, R.; Mikulášková, E.; et al. Long-term and contemporary environmental conditions as determinants of the species composition of bog organisms. Freshw. Biol. 2013, 58, 2196–2207. [Google Scholar] [CrossRef]
- Jurasinski, G.; Ahmad, S.; Anadon-Rosell, A.; Berendt, J.; Beyer, F.; Bill, R.; Blume-Werry, G.; Couwenberg, J.; Günther, A.; Joosten, H.; et al. From understanding to sustainable use of peatlands: The Wetscapes approach. Soil Syst. 2020, 4, 14. [Google Scholar] [CrossRef]
- Bragazza, L.; Buttler, A.; Habermacher, J.; Brancaleoni, L.; Gerdol, R.; Fritze, H.; Hanajík, P.; Laiho, R.; Johnson, D. High nitrogen deposition alters the decomposition of bog plant litter and reduces carbon accumulation. Glob. Chang. Biol. 2012, 18, 1163–1172. [Google Scholar] [CrossRef]
- Temmink, R.J.M.; Lamers, L.P.M.; Angelini, C.; Bouma, T.J.; Fritz, C.; van de Koppel, J.; Lexmond, R.; Rietkerk, M.; Silliman, B.R.; Joosten, H.; et al. Recovering wetland biogeomorphic feedback to restore the world’s biotic carbon hotspots. Science 2022, 6, 376–1479. [Google Scholar] [CrossRef]
- Basińska, A.M.; Reczuga, M.K.; Gąbka, M.; Stróżecki, M.; Luców, D.; Samson, M.; Urbaniak, M.; Leśny, J.; Chojnicki, B.H.; Gilbert, D.; et al. Experimental warming, and precipitation reduction affect the biomass of microbial communities in Sphagnum peatland. Ecol. Indic. 2020, 112, 106059. [Google Scholar] [CrossRef]
- Alexander, T.J.; Vonlanthen, P.; Seehausen, O. Does eutrophication-driven evolution change aquatic ecosystems? Philos. Trans. R. Soc. B 2017, 19, 372–1712. [Google Scholar] [CrossRef]
- García-Girón, J.; Bini, L.M.; Heino, J. Shortfalls in our understanding of the causes and consequences of functional and phylogenetic variation of freshwater communities across continents. Biol. Conserv. 2023, 282, 110082. [Google Scholar] [CrossRef]
- Le-Bagousse-Pinguet, J.; Soliveres, S.; Gross, N.; Maestre, F.T. Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality. Biol. Sci. 2019, 116, 8419–8424. [Google Scholar] [CrossRef]
- Le Moal, M.; Gascuel-Odoux, C.; Ménesguen, A.; Souchon, Y.; Étrillard, C.; Levain, A.; Moatar, F.; Pannard, A.; Souchu, P.; Lefebvre, A.; et al. Eutrophication: A new wine in an old bottle? Sci. Total Environ. 2019, 651, 1–11. [Google Scholar] [CrossRef]
- Heino, J.; Alahuhta, J.; Bini, L.M.; Cai, Y.; Heiskanen, A.S.; Hellsten, S.; Kortelainen, P.; Kotamäki, N.; Tolonen, K.; Vihervaara, P.; et al. Lakes in the era of global change: Moving beyond single-lake thinking in maintaining biodiversity and ecosystem services. Biol. Rev. 2021, 96, 89–106. [Google Scholar] [CrossRef]
- Mondragón-Camarillo, L.; Zaragoza, S.R. Food Webs. Plankton Communities 2022, 1, 1–24. [Google Scholar]
- Rolls, R.; Deane, D.; Johanson, S.; Heino, J.; Anderson, M.; Ellingsen, K. Biotic homogenisation and differentiation as directional change in beta diversity: Synthesising driver–response relationships to develop conceptual models across ecosystems. Biol. Rev. 2023, 98, 1388–1423. [Google Scholar] [CrossRef] [PubMed]
- Van Diggelen, R.; Middleton, B.; Bakker, J.; Grootjans, A.; Wassen, M. Fens and floodplains of the temperate zone: Present status, threats, conservation and restoration. Appl. Veg. Sci. 2006, 9, 157–162. [Google Scholar] [CrossRef]
- Schindler, D.W. Recent advances in the understanding and management of eutrophication. Limnol. Oceanogr. 2006, 51, 356–363. [Google Scholar] [CrossRef]
- Zak, D.; Wagner, C.; Payer, B.; Augustin, J.; Gelbrecht, J. Phosphorus mobilization in rewetted fens: The effect of altered peat properties and implications for their restoration. Ecol. Appl. 2010, 20, 1336–1349. [Google Scholar] [CrossRef] [PubMed]
- Walton, C.R.; Zak, D.; Audet, J.; Petersen, R.J.; Lange, J.; Oehmke, C.; Wichtmann, W.; Kreyling, J.; Grygoruk, M.; Jabłońska, E.; et al. Wetland buffer zones for nitrogen and phosphorus retention: Impacts of soil type, hydrology and vegetation. Sci. Total Environ. 2020, 727, 138709. [Google Scholar] [CrossRef]
- Giang, T.; Hieu, X.; Szmutzera, T.; Scholz, U.; Pecinka, A.; Schuberta, I. Metatranscriptome analysis reveals host-microbiome interactions in traps of carnivorous Genlisea species. Front. Microbiol. 2015, 6, 526. [Google Scholar]
- Płachno, B.J.; Adamec, L.; Kaińska, I. Relationship between trap anatomy and function in Australian carnivorous bladderworts (Utricularia) of the subgenus Polypompholyx. Aquat. Bot. 2015, 120, 290–296. [Google Scholar] [CrossRef]
- Adamec, L. Mineral nutrition of carnivorous plants: A review. Bot. Rev. 1997, 63, 273–299. [Google Scholar] [CrossRef]
- Adamec, L. Oxygen concentrations inside the traps of the carnivorous plants Utricularia and Genlisea (Lentibulariaceae). Ann. Bot. 2007, 100, 849–856. [Google Scholar] [CrossRef]
- Grodnitskaya, I.; Karpenko, L.V.; Knorre, A.; Syrtsov, S.N. Microbial activity of peat soils of boggy larch forests and bogs in the permafrost zone of central Evenkia. Eurasian Soil Sci. 2013, 46, 61–73. [Google Scholar] [CrossRef]
- Sirovà, D.; Bàrta, J.; Šimek, K.; Posch, T.; Pech, J.; Stone, J.; Borovec, J.; Adamec, L.; Vrba, J. Hunters or farmers? Microbiome characteristics help elucidate the diet composition in an aquatic carnivorous plant. Microbiome 2018, 7, 225. [Google Scholar] [CrossRef]
- Donohue, I.; Jackson, A.L.; Pusch, M.T.; Irvine, K. Nutrient enrichment homogenizes lake benthic assemblages at local and regional scales. Ecology 2009, 90, 3470–3477. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.C.; Housley, L.; Back, J.A.; King, R.S. Freshwater eutrophication drives sharp reductions in temporal beta diversity. Ecology 2018, 99, 47–56. [Google Scholar] [CrossRef]
- Zhang, H.; Väliranta, M.; Piilo, S.; Amesbury, M.J.; Aquino-López, M.A.; Roland, T.P.; Salminen-Paatero, S.; Paatero, J.; Lohila, A.; Tuittila, E.S. Decreased carbon accumulation feedback driven by climate-induced drying of two southern boreal bogs over recent centuries. Glob. Chang. Biol. 2020, 26, 2435–2448. [Google Scholar] [CrossRef]
- Geng, M.D.; Zhang, W.; Hu, T.; Wang, R.; Cheng, X.; Wang, J. Eutrophication causes microbial community homogenization via modulating generalist species. Water Res. 2022, 210, 118003. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Song, W.; Warren, A. Periphytic ciliate colonization: Annual cycle and responses to environmental conditions. Aquat. Microb. Ecol. 2005, 39, 159–170. [Google Scholar] [CrossRef][Green Version]
- Krasuska, U.; Glinka, A.; Gniazdowska, A. Carnivorous plant menu. KOSMOS-Probl. Nauk Biol. 2012, 61, 635–646. [Google Scholar]
- Tran, T.D.; Cao, H.X.; Jovtchev, G.; Novák, P.; Giang, T.; Vu, H.; Macas, J.; Schubert, J.; Fuchs, J. Chromatin organization and cytological features of carnivorous Genlisea species with large genome size differences. Front. Plant. Sci. 2015, 6, 151809. [Google Scholar] [CrossRef]
- Friday, L.E.; Quarmby, C. Uptake and translocation of prey-derived 15N and 32P in Utricularia vulgaris L. New Phytol. 1994, 126, 273–281. [Google Scholar] [CrossRef]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identification and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–984. [Google Scholar] [CrossRef]
- Caron, D.A. Technique for enumeration of heterotrophic and phototrophic nanoplankton, using epifluorescence microscopy and comparison with other procedures. Appl. Environ. Microbiol. 1983, 46, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitt. Internat. Verein. Limnol. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Gilbert, D.; Amblard, C.; Bourdier, G.; Francez, A.J. The microbial loop at the surface of a peatland: Structure, functioning and impact of nutrients inputs. Microb. Ecol. 1998, 35, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Bottrell, H.H.; Duncan, A.; Gliwicz, Z.M.; Grygierek, E.; Herzig, A.; Hillbricht-Ilkowska, A.; Kurasawa, H.; Larsson, P.; Weglenska, T. A reviewer of some problems in zooplankton production studies. IBP 1976, 24, 419–456. [Google Scholar]
- Golterman, H.L. Methods for Chemical Analysis of Fresh Waters; Blackwell Scientific: Oxford, UK; Edinburgh, UK, 1969; p. 172. [Google Scholar]
- Lundin, L.; Nilsson, T.; Jordan, S.; Lode, E.; Strömgren, M. Impacts of rewetting on peat, hydrology, and water chemical composition over 15 years in two finished peat extraction areas in Sweden. Wetl. Ecol. Manag. 2017, 25, 405–419. [Google Scholar] [CrossRef]
- Mieczan, T.; Adamczuk, M.; Pawlik-Skowrońska, B.; Toporowska, M. Eutrophication of peatbogs: Consequences of P and N enrichment for microbial and metazoan communities in mesocosm experiments. Aqua. Microb. Ecol. 2015, 74, 121–141. [Google Scholar] [CrossRef]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef]
- Bragazza, L.; Freeman, C.; Jones, T.; Rydin, H.; Limpens, J.; Fenner, N.; Ellis, T.; Gerdol, R.; Hàjek, M.; Hàjek, T.; et al. Atmospheric nitrogen deposition promotes carbon loss from peat bogs. Proc. Natl. Acad. Sci. USA 2006, 103, 19386–19389. [Google Scholar] [CrossRef]
- Bubier, J.; Moore, T.M.; Bledzki, L. Effects of nutrient addition on vegetation and carbon cycling in an ombrothropic bog. Glob. Chang. Biol. 2007, 13, 1167–1186. [Google Scholar] [CrossRef]
- Lin, H.Y.; Yeh, W.Y.; Tsai, S.F.; Chiang, K.P.; Lin, J.Y.; Tsao, C.C.; Lin, H.J. Biological protective effects against vibrio infections in grouper larvae using the Strombidium sp. NTOU1, a marine ciliate amenable for scaled-up culture and with an excellent bacteriovorous ability. Front. Mar. Sci. 2020, 7, 373. [Google Scholar] [CrossRef]
- Wu, Y.; Blodau, C.; Moore, T.R.; Bubier, J.; Juutinen, S.T.; Larmola, T. Effects of experimental nitrogen deposition on peatland carbon pools and fluxes: A modelling analysis. Biogeosciences 2015, 12, 79–101. [Google Scholar] [CrossRef]
- Sirova, D.; Borovec, J.; Picek, T.; Adamec, L.; Nedbalova, L.; Vrba, J. Ecological implications of organic carbon dynamics in the traps of aquatic carnivorous Utricularia plants. Funct. Plant Biol. 2011, 38, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Peltoniemi, K.; Fritze, H.; Laiho, R. Response of fungal and actinobacterial communities to water-level drawdown in boreal peatland sites. Soil Biol. Biochem. 2009, 41, 1902–1914. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lee, S.H.; Freeman, C.; Fenner, N.; Kang, H. Comparative analysis of soil microbial communities and their responses to the short-term drought in bog, fen, and riparian wetlands. Soil Biol. Biochem. 2008, 40, 2874–2880. [Google Scholar] [CrossRef]
- Mitchell, E.A.D.; Gilbert, D.; Amblard, C.; Grosverinier, P.; Gobat, J.M. Structure of microbial communities in Sphagnum peatlands and effect of atmospheric carbon dioxide enrichment. Microb. Ecol. 2003, 46, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Chróst, J.R.; Siuda, I. Microbial production, utilization, and enzymatic degradation of organic matter in the upper trophogenic layer in the pelagial zone of lakes along a eutrophication gradient. Limnol. Oceanogr. 2006, 51, 749–762. [Google Scholar] [CrossRef]
- Lin, X.; Green, S.; Tfaily, M.M.; Prakash, O.; Konstantinidis, K.T.; Corbett, J.E.; Chanton, J.P.; Cooper, W.T.; Kostka, J.E. Microbial community structure and activity linked to contrasting biogeochemical gradients in bog and fen environments of the Glacial Lake Agassiz Peatland. Appl. Environ. Microbiol. 2012, 78, 7023–7031. [Google Scholar] [CrossRef]
- Larmola, T.; Tuittila, E.S.; Tiirola, M.; Nykanen, H.; Martikainen, M.J.; Yrjälä, K.; Tuomivirta, T.; Fritze, H. The role of Sphagnum mosses in the methane cycling of a boreal mire. Ecology 2010, 91, 2356–2365. [Google Scholar] [CrossRef]
- Chróst, R.J.; Adamczewski, T.; Kalinowska, K.; Skowrońska, A. Abundance and structure of microbial loop components (bacteria and protists) in lakes of different trophic status. J. Microbiol. Biotechnol. 2009, 19, 858–868. [Google Scholar] [CrossRef]
- Siragusa, A.J.; Swenson, J.E.; Casamatta, D.A. Culturable bacteria present in the fluid of the hooded-pitcher plant Sarracenia minor based on 16S rDNA gene sequence data. Microb. Ecol. 2007, 54, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.H. Bladder function in Utricularia purpurea (Lentibulariaceae): Is carnivory important? Am. J. Bot. 2001, 88, 170–176. [Google Scholar] [CrossRef]
- Sirová, D.; Adamec, L.; Vrba, J. Enzymatic activities in traps of four aquatic species of the carnivorous genus Utricularia. New Phytol. 2003, 159, 669–675. [Google Scholar] [CrossRef]
- Buosi, P.R.B.; Pauleto, G.M.; Lansac-Toha, F.A.; Velho, L.F.M. Ciliate community associated with aquatic macrophyte roots: Effects of nutrient enrichment on the community composition and species richness. Eur. J. Protistol. 2011, 47, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Sirova, D.; Borovec, J.; Černá, B.; Rejmánková, E.; Adamec, L. Microbial community development in the traps of aquatic Utricularia species. Aquat. Bot. 2009, 90, 129–136. [Google Scholar] [CrossRef]
- Auer, B.; Arndt, H. Taxonomic composition, and biomass of heterotrophic flagellates in relation to lake trophy and season. Freshw. Biol. 2001, 46, 959–972. [Google Scholar] [CrossRef]
- Davidson, E.D.; Janssens, I.A. Temperature sensitivity of soil C decomposition and feedback to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef]
- Płachno, B.J.; Łukaszek, M.; Wolowski, K.; Adamec, M.; Stolarczyk, P. Aging of Utricularia traps and variability of microorganisms associated with that microhabita. Aquat. Bot. 2012, 97, 101–110. [Google Scholar] [CrossRef]
- Mieczan, T.; Bartkowska, A. The effect of experimentally simulated climate warming on the microbiome of carnivorous plants—A microcosm experiment. Glob. Ecol. Conserv. 2022, 34, e02040. [Google Scholar] [CrossRef]
- Adamec, L. Soil fertilization enhances growth of the carnivorous plant Genlisea violacea. Biologia 2008, 63, 201–203. [Google Scholar] [CrossRef]
- Koenig, I.; Feldmeyer-Christe, E.; Mitchell, E.A.D. Comparative ecology of vascular plant, bryophyte, and testate amoeba communities in four Sphagnum peatlands along an altitudinal gradient in Switzerland. Ecol. Indic. 2015, 54, 48–59. [Google Scholar] [CrossRef]
- Marcisz, K.; Marcisz, D.; Colombaroli, V.E.; Jassey, W.; Tinner, P.; Kołaczek, M.; Gałka, M.; Karpińska-Kołaczek, M.; Słowiński, M.; Lamentowicz, M. A novel testate amoebae trait-based approach to infer environmental disturbance in Sphagnum peatlands. Sci. Rep. 2016, 6, 33907. [Google Scholar] [CrossRef] [PubMed]
- Lamentowicz, M.; Gałka, M.; Marcisz, K.; Słowiński, M.; Kajukało-Drygalska, K.; Dayras, M.D.; Jassey, V.E.J. Unveiling tipping points in long-term ecological records from Sphagnum—Dominated peatlands. Biol. Lett. 2019, 15, 20190043. [Google Scholar]
- Carballeira, R.; Pontevedra-Pombala, X. Diversity of Testate Amoebae as an Indicator of the Conservation Status of Peatlands in Southwest Europe. Ecol. Biogeogr. Evol. Biol. Peatl. 2021, 13, 269. [Google Scholar] [CrossRef]
- Jassey, V.E.; Shimano, S.; Dupuy, C.; Toussaint, M.L.; Gilbert, D. Characterizing the feeding habits on the testate amoebae Hyalosphenia papilio and Nebela tincta along narrow “fen-bog” gradient using digestive vacuole content and 13C and 15N isotopic analyses. Protist 2011, 163, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Mazei, Y.; Tsyganov, A.N.; Bubnova, O.A. Structure of community of testate amoebae in a sphagnum dominated bog in upper sura flow (Middle Volga Territory). Biol. Bull. 2007, 4, 462–474. [Google Scholar] [CrossRef]
- Mitchell, E.A.D.; Charman, D.J.; Warner, B.G. Testate amoebae analysis in ecological and paleoecological studies of wetlands: Past, present and future. Biodivers. Conserv. 2008, 17, 2115–2137. [Google Scholar] [CrossRef]
- Lamentowicz, M.; Kajukało-Drygalska, K.; Kołaczek, P.; Jassey, V.E.J.; Gąbka, M.; Karpińska-Kołaczek, M. Testate amoebae taxonomy and trait diversity are coupled along an openness and wetness gradient in pine-dominated Baltic bogs. Eur. J. Protistol. 2020, 73, 125674. [Google Scholar] [CrossRef]
- Davis, S.R.; Wilkinson, D.M. The conservation management value of testate amoebae as ‘restoration’ indicators: Speculations based on two damaged raised mires in northwest England. Holocene 2004, 14, 135–143. [Google Scholar] [CrossRef]
- Lamentowicz, M.; Mitchell, E.A.D. Testate Amoebae as Ecological and Palaeohydrological Indicators in Peatlands—The Polish Experience; Taylor & Francis Group: London, UK, 2007; pp. 85–90. [Google Scholar]
- Marcisz, K.; Jassey, V.E.J.; Kosakyan, A.; Krashevska, V.; Lahr, D.J.G.; Lara, E.; Lamentowicz, M.; Macumber, A.; Mazei, Y.; Mitchell, E.A.D.; et al. Testate amoebae functional traits and their use in paleoecology. Front. Ecol. Evol. 2020, 8, 575–966. [Google Scholar] [CrossRef]
- Warner, B.; Asada, T.; Quinn, N. Seasonal influences on the ecology of testate amoebae (Protozoa) in a small Sphagnum peatland in Southern Ontario, Canada. Microb. Ecol. 2007, 54, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Jassey, V.E.; Signarbieux, C.; Hättenschwiler, S.; Bragazza, L.; Buttler, A.; Delarue, F.; Fournier, B.; Gilbert, D.; Laggoun-Défarge, F.; Lara, E.; et al. An unexpected role for mixotrophs in the response of peatland carbon cycling to climate warming. Sci. Rep. 2015, 5, 16931. [Google Scholar] [CrossRef] [PubMed]
- Nicolau, A.; Martins, M.J.; Mota, M.; Lima, N. Effect of copper in the protistan community of activated sludge. Chemosphere 2005, 58, 605–614. [Google Scholar] [CrossRef]
- Payne, R.J. Can testate amoeba-based palaeohydrology be extended to fens? J. Quat. Sci. 2011, 26, 15–27. [Google Scholar] [CrossRef]
- Ravindran, C.; Irudayarajan, L.; Raveendran, H.P. Possible beneficial interactions of ciliated protozoans with coral health and resilience. Appl. Environ. Microbiol. 2023, 89, 10. [Google Scholar] [CrossRef] [PubMed]
- Kexin, Z.; Muqi, X.; Biao, L.; Hong, C. Characteristics of microfauna and their relationships with the performance of an activated sludge plant in China. J. Environ. Sci. 2007, 20, 482–486. [Google Scholar]
- Nguyen-Viet, H.; Gilbert, D.; Mitchell, E.A.D.; Badot, P.M.; Bernard, N. Effects of experimental lead pollution on the microbial communities associated with Sphagnum fallax (Bryophyta). Microb. Ecol. 2007, 54, 232–241. [Google Scholar] [CrossRef]
- Wilkinson, D.M.; Mitchell, E.A.D. Testate amoebae and nutrient cycling with particular reference to soils. Geomicrobiol. J. 2010, 27, 520–533. [Google Scholar] [CrossRef]
- Mieczan, T.; Bielańska-Grajner, I.; Tarkowska-Kukuryk, M. Hydrochemical and microbiological distinction and function of ombrotrophic peatland lagg as ecotone between Sphagnum peatland and forest catchment (Poleski National Park, eastern Poland). Ann. Limnol.-Int. J. Limnol. 2012, 48, 323–336. [Google Scholar] [CrossRef][Green Version]
- Zingel, P.; Huitu, E.; Makela, S.; Arvola, L. The abundance and diversity of planktonic ciliates in 12 boreal lakes of varying trophic state. Arch. Hydrobiol. 2002, 155, 315–333. [Google Scholar] [CrossRef]
- Mieczan, T.; Tarkowska-Kukuryk, M.; Adamczuk, M.; Pęczuła, W.; Demetraki-Paleolog, A.; Niedźwiecki, M. Research of different types of peatbogs: Relationships of biocenosis structures and physico-chemical parameters. Pol. J. Environ. Stud. 2015, 1, 191–198. [Google Scholar]
- Andersen, R.; Wells, C.; Macrae, M.; Price, J. Nutrient mineralization and microbial functional diversity in a restored bog approach natural conditions 10 years post restoration. Soil Biol. Biochem. 2013, 64, 37–47. [Google Scholar] [CrossRef]
- Lukić, D.; Limberger, R.; Agatha, S.; Montagnes, D.J.; Weisse, T. Thermal performance of planktonic ciliates differs between marine and freshwaters: A case study providing guidance for climate change studies Scientific Significance Statement. Limnol. Oceanogr. 2022, 7, 520–526. [Google Scholar] [CrossRef]
- Andersen, R.; Chapman, S.J.; Artz, R.R.E. Microbial communities in natural and disturbed peatlands:a review. Soil Biol. Biochem. 2013, 57, 979–994. [Google Scholar] [CrossRef]
- Šimek, K.; Pitsch, G.; Salcher, M.M.; Sirová, D.; Shabarova, T.; Adamec, L. Ecological traits of the algae-bearing Tetrahymena utriculariae (Ciliophora) from traps of the aquatic carnivorous plant Utricularia reflexa. J. Eukaryot. Microbiol. 2017, 64, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Weisse, T.; Anderson, R.; Arndt, H.; Calbet, A.; Hansen, P.J.; Montagnes, D.J.S. Functional ecology of aquatic phagotrophic protists—Concepts, limitations, and perspectives. Eur. J. Protistol. 2016, 55, 50–74. [Google Scholar] [CrossRef] [PubMed]
- Arndt, H. Rotifers as predators on components of the microbial web (bacteria, heterotrophic flagellates, ciliates)—A review. Hydrobiologia 1993, 255, 231–246. [Google Scholar] [CrossRef]
- Kim, S.; Kim, J.; Joo, J.; Choi, J. Response of the rotifer community to human-induced changes in the trophic state of a reservoir. Oceanol. Hydrobiol. Stud. 2020, 49, 329–344. [Google Scholar] [CrossRef]
- Płachno, B.J.; Faber, J.; Jankun, A. Cuticular discontinuities in glandular hairs of Genlisea St.-Hil. in relation to their functions. Acta Bot. Gall. 2005, 152, 125–130. [Google Scholar] [CrossRef]
- Jeppesen, E.; Nõges, P.; Davidson, T.A.; Haberman, J.; Nõges, T.; Blank, K.; Lauridsen, T.L.; Søndergaard, M.; Sayer, C.; Laugaste, R.; et al. Zooplankton as indicators in lakes:a scientificbased plea for including zooplankton in the ecological quality assessment according to the European Water Framework Directive (WFD). Hydrobiologia 2011, 676, 279–297. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, B.; Trivedi, C.; Eldrige, D.; Abades, S.; Alfaro, F.; Bastide, F.; Berhe, A.; Cutler, N.; Gallardo, A.; et al. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Sauvadet, M.; Chauvat, M.; Cluzeau, D.; Maron, P.; Villenave, C.; Bertrand, I. The dynamics of soil micro-food web structure and functions vary according to litter quality. Soil Biol. Biochem. 2016, 95, 262–274. [Google Scholar] [CrossRef]
- Jassey, V.E.; Meyer, C.; Dupuy, C.; Bernard, N.; Mitchell, E.A.D.; Toussaint, M.L.; Metian, M.; Chatelain, A.P.; Gilbert, D. To what extent do food preferences explain the trophic position of heterotrophic and mixotrophic microbial consumers in Sphagnum peatland? Microb. Ecol. 2013, 66, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Dolan, J.R. Doing a Protist Book. Protist 2013, 164, 154–159. [Google Scholar] [CrossRef]
- Bielańska-Grajner, I.; Ejsmont-Karabin, J.; Iakovenko, N. Wrotki (Rotifera, Bdelloidea), Fauna Słodkowodna Polski; Uniwersytet Łódzki: Łódź, Poland, 2013; p. 153. [Google Scholar]
- Reczuga, M.K.; Lamentowicz, M.; Mulot, M.; Mitchell, E.A.D.; Buttler, A.; Chojnicki, B.; Słowiński, M.; Binet, P.; Chiapusio, G.; Gilbert, D.; et al. Predator-prey mass ratio drives microbial activity and its response to drought in Sphagnum peatlands. Ecol. Evol. 2018, 8, 5752–5764. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spring | Summer | Autumn | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | ×2N | ×4N | C | ×2N | ×4N | C | ×2N | ×4N | |
| T (°C) | 24 | 23.6 | 24.8 | 24.4 | 28.9 | 29.7 | 24.4 | 26 | 26 |
| pH | 5.4 | 6.2 | 6.51 | 6.06 | 6.06 | 6.13 | 6.13 | 6.57 | 6.6 |
| Cond. (μS cm−1) | 29.1 | 29.5 | 33.3 | 31 | 29.8 | 30.04 | 31 | 33.3 | 39.1 |
| O2 (mg L−1) | 8.09 | 7.9 | 7.7 | 8.29 | 7.7 | 7.65 | 7.66 | 7.66 | 6.46 |
| O2% | 97 | 94.2 | 92.4 | 101 | 93.5 | 92.2 | 96.1 | 92.3 | 83 |
| Ntot (mg N L−1) | 1.204 | 1.335 | 1.846 | 1.203 | 1.452 | 1.505 | 1.397 | 1.441 | 1.440 |
| N-NO3 (mg N L−1) | 0.072 | 0.342 | 0.353 | 0.105 | 0.220 | 0.250 | 0.114 | 0.148 | 0.198 |
| N-NH4 (mg N L−1) | <0.01 | 0.01 | 0.012 | 0.017 | <0.01 | 0.022 | 0.025 | 0.027 | 0.031 |
| Chl a (μg L−1) | 37 | 18 | 17 | 50 | 80 | 280 | 50 | 58 | 252 |
| COD (mg L−1) | 65.6 | 68.5 | 70 | 67 | 66 | 65 | 67 | 65 | 66.5 |
| BOD (mg L−1) | 40 | 41.5 | 42.5 | 39.5 | 40 | 40.5 | 40.5 | 39 | 40.5 |
| TOC (mg L−1) | 30 | 31 | 32 | 29.8 | 30.2 | 30.5 | 29.2 | 30.5 | 31.5 |
| SP | SU | AU | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| O2 | pH | Chl-a | Electrical Conductivity | N-NH4 | O2 | pH | Chl-a | Electrical Conductivity | N-NH4 | O2 | pH | Chl-a | Electrical Conductivity | N-NH4 | ||
| C | W | 0.62 | 0.68 | 0.37 | 0.46 | 0.43 | 0.39 | |||||||||
| T | 0.51 | 0.56 | ||||||||||||||
| ×2N | W | 0.57 | −0.44 | 0.56 | 0.42 | |||||||||||
| T | 0.22 | 0.67 | −0.43 | 0.36 | 0.39 | −0.21 | 0.21 | |||||||||
| ×4N | W | 0.5 | ||||||||||||||
| T | −0.24 | 0.46 | −0.69 | 0.54 | ||||||||||||
| SP | SU | AU | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B | HNF | TA | C | Cr | B | HNF | TA | C | Cr | B | HNF | TA | C | Cr | ||
| C | B | 0.48 | ||||||||||||||
| HNF | 0.56 | |||||||||||||||
| TA | 0.52 | 0.64 | 0.47 | 0.43 | ||||||||||||
| C | 0.45 | 0.57 | ||||||||||||||
| Cr | ||||||||||||||||
| ×2N | B | 0.67 | ||||||||||||||
| HNF | 0.78 | |||||||||||||||
| TA | 0.62 | 0.69 | 0.76 | |||||||||||||
| C | 0.48 | 0.43 | 0.58 | |||||||||||||
| Cr | ||||||||||||||||
| B | 0.69 | |||||||||||||||
| ×4N | HNF | 0.82 | ||||||||||||||
| TA | 0.59 | |||||||||||||||
| C | 0.53 | 0.62 | - | |||||||||||||
| Cr | ||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartkowska, A.; Mieczan, T. Effect of Simulated Eutrophication of Peatlands on the Microbiome of Utricularia vulgaris L. Water 2024, 16, 1046. https://doi.org/10.3390/w16071046
Bartkowska A, Mieczan T. Effect of Simulated Eutrophication of Peatlands on the Microbiome of Utricularia vulgaris L. Water. 2024; 16(7):1046. https://doi.org/10.3390/w16071046
Chicago/Turabian StyleBartkowska, Aleksandra, and Tomasz Mieczan. 2024. "Effect of Simulated Eutrophication of Peatlands on the Microbiome of Utricularia vulgaris L." Water 16, no. 7: 1046. https://doi.org/10.3390/w16071046
APA StyleBartkowska, A., & Mieczan, T. (2024). Effect of Simulated Eutrophication of Peatlands on the Microbiome of Utricularia vulgaris L. Water, 16(7), 1046. https://doi.org/10.3390/w16071046