
Research on the Ecological Restoration Effects of a Vallisneria natans (Lour.) Hara-Dominated Multitrophic Level Ecosystem


 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Design
2.3. Measurement Methods
2.4. Data Analysis
3. Results
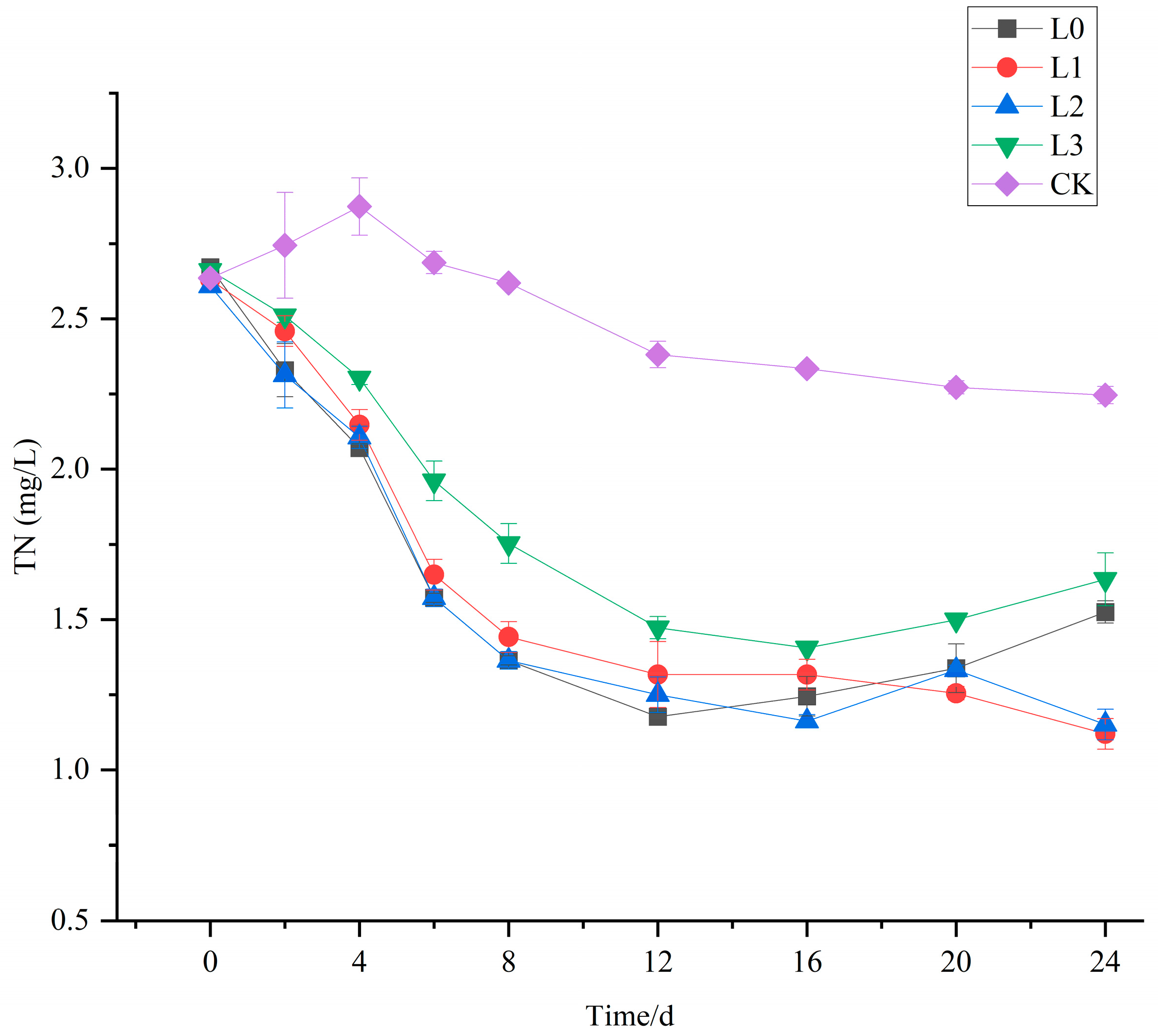
3.1. The Purification Effect of V. natans and B. aeruginosa Configuration on TN in Water Bodies
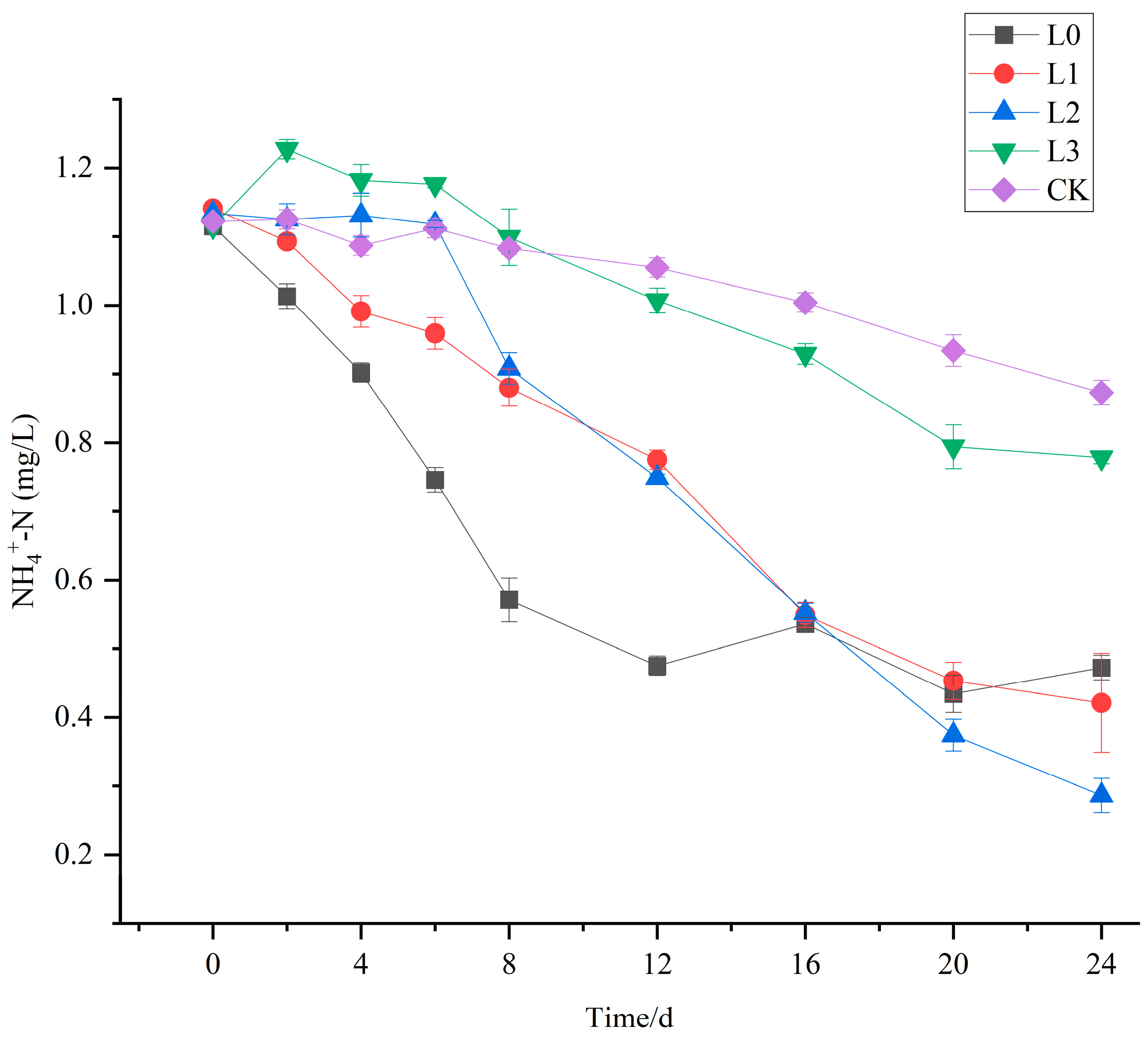
3.2. The Purification Effect of V. natans and B. aeruginosa Configuration on NH4+-N in Water Bodies
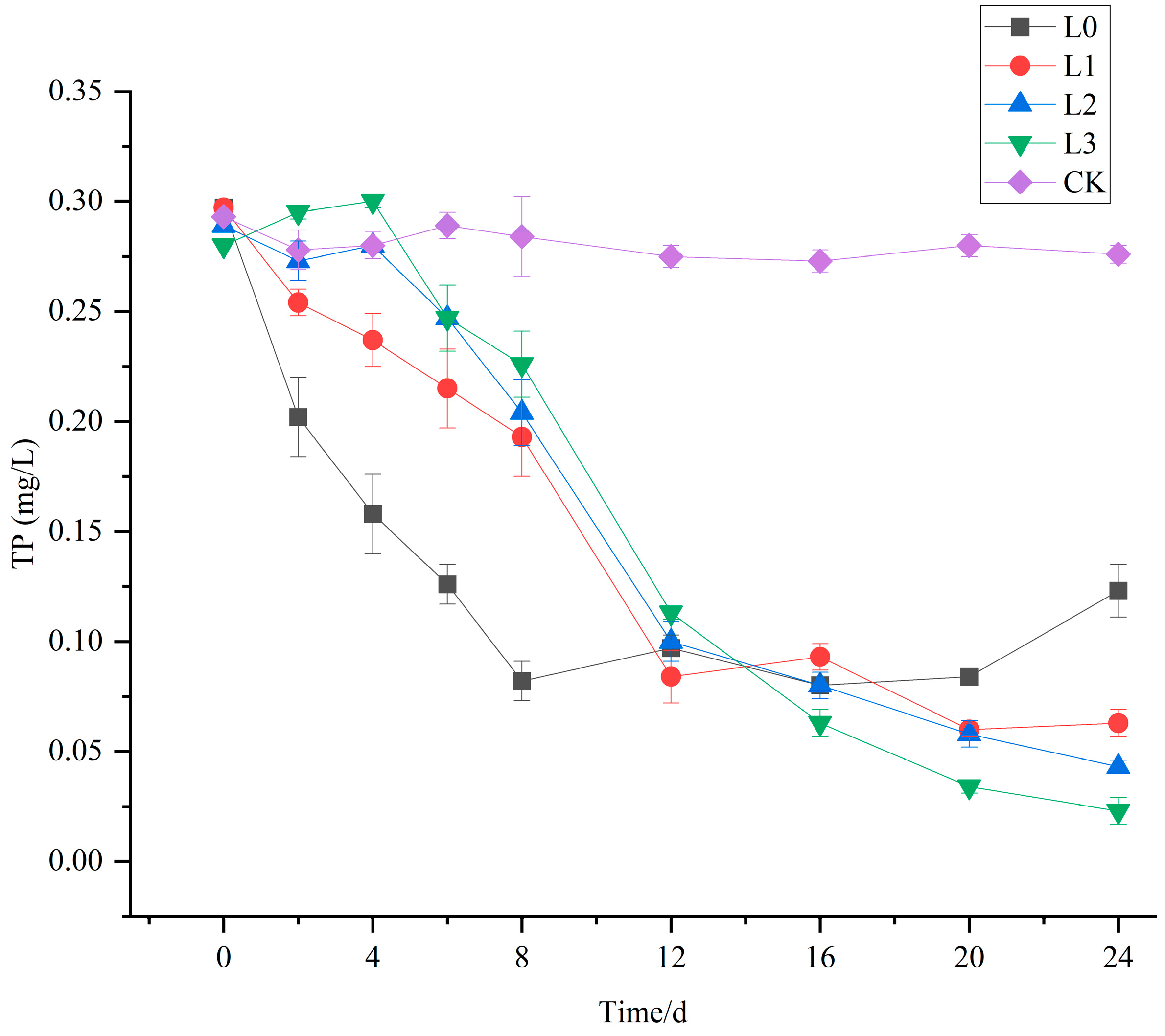
3.3. The Purification Effect of V. natans and B. aeruginosa Configuration on Total Phosphorus (TP) in Water Bodies
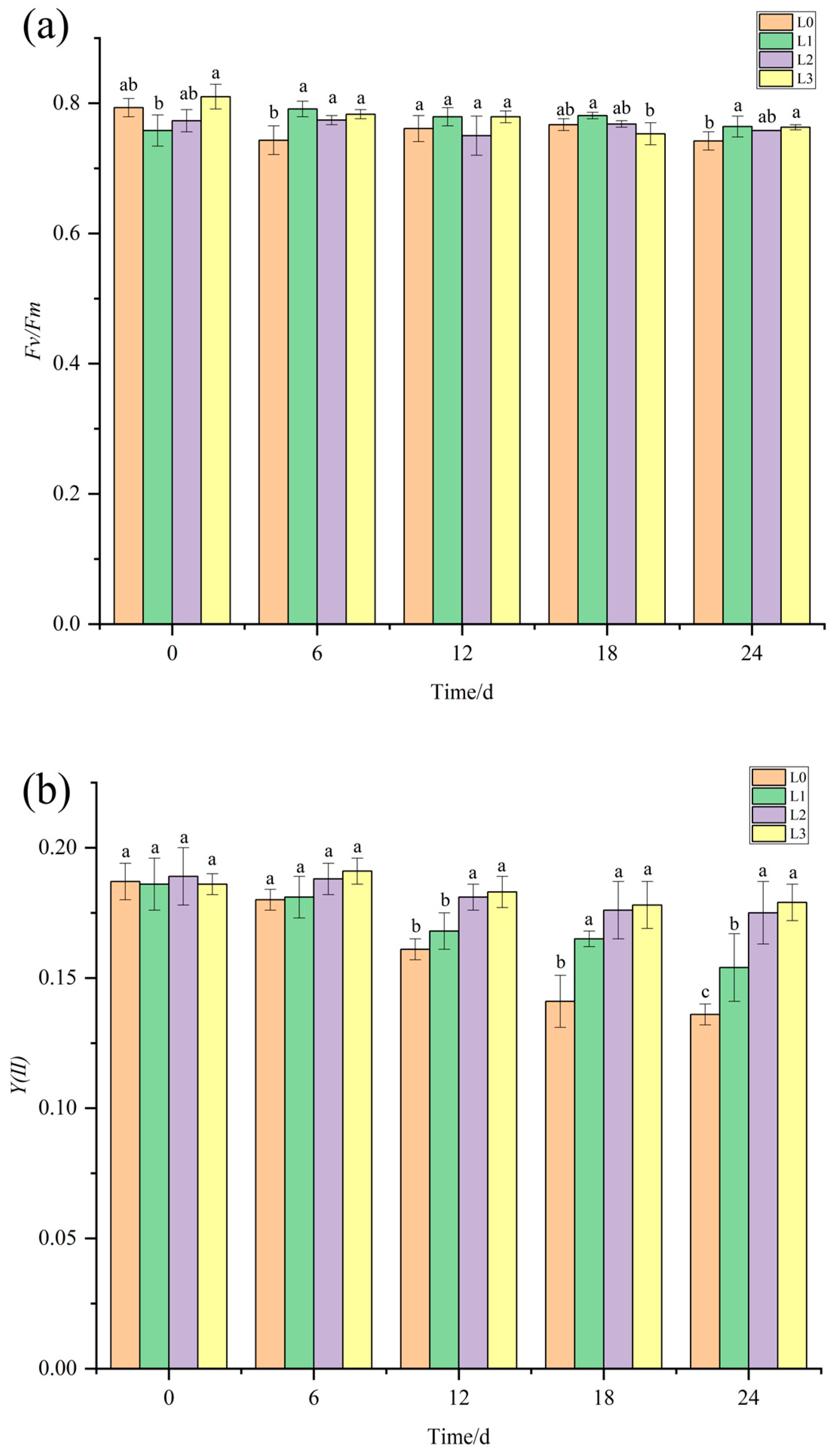
3.4. The Impact of V. natans and B. aeruginosa Configuration on the Photosynthetic Physiology of V. natans
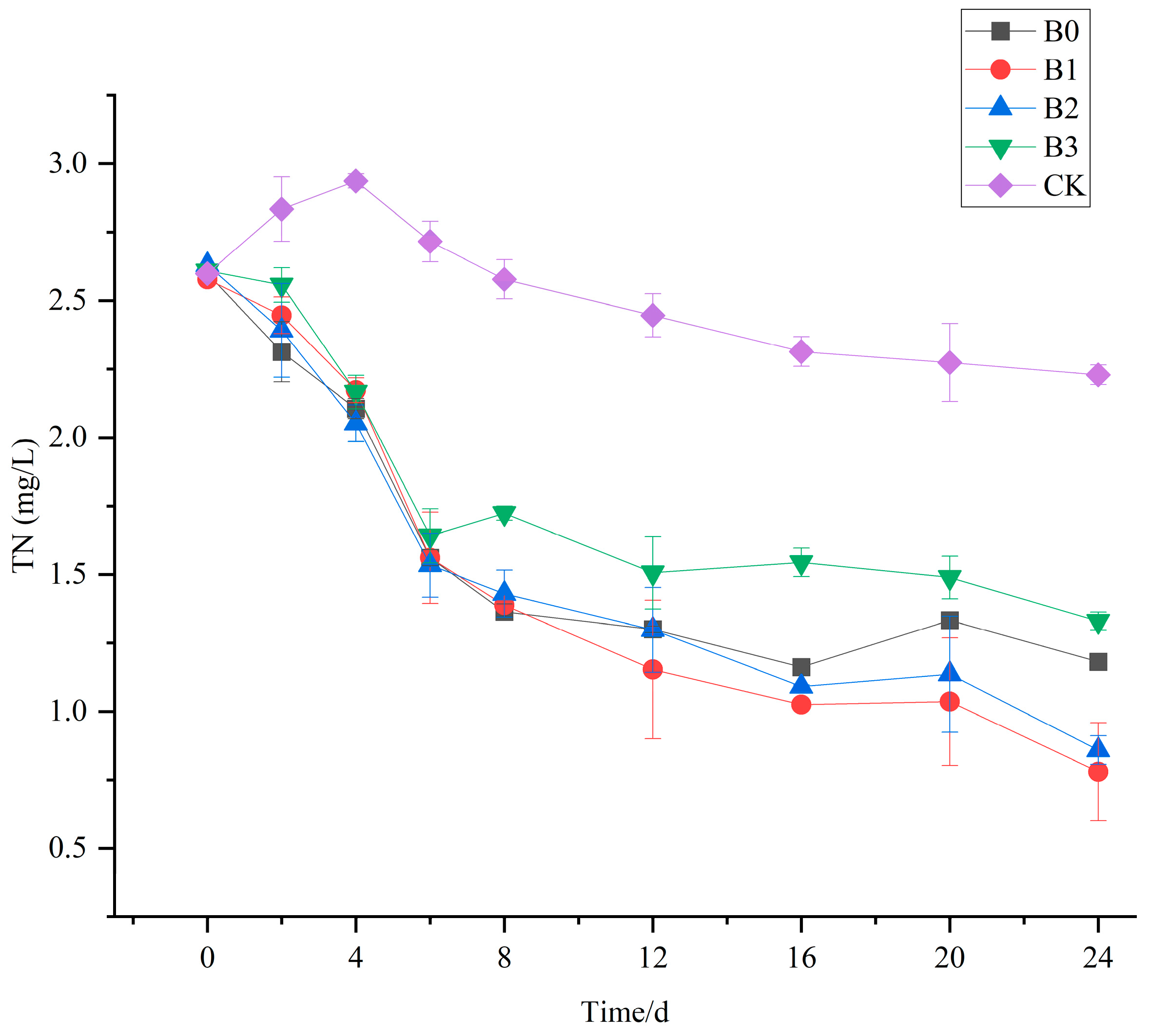
3.5. The Purification Effect of V. natans, B. aeruginosa, and H. cumingii Configuration on TN in Water Bodies
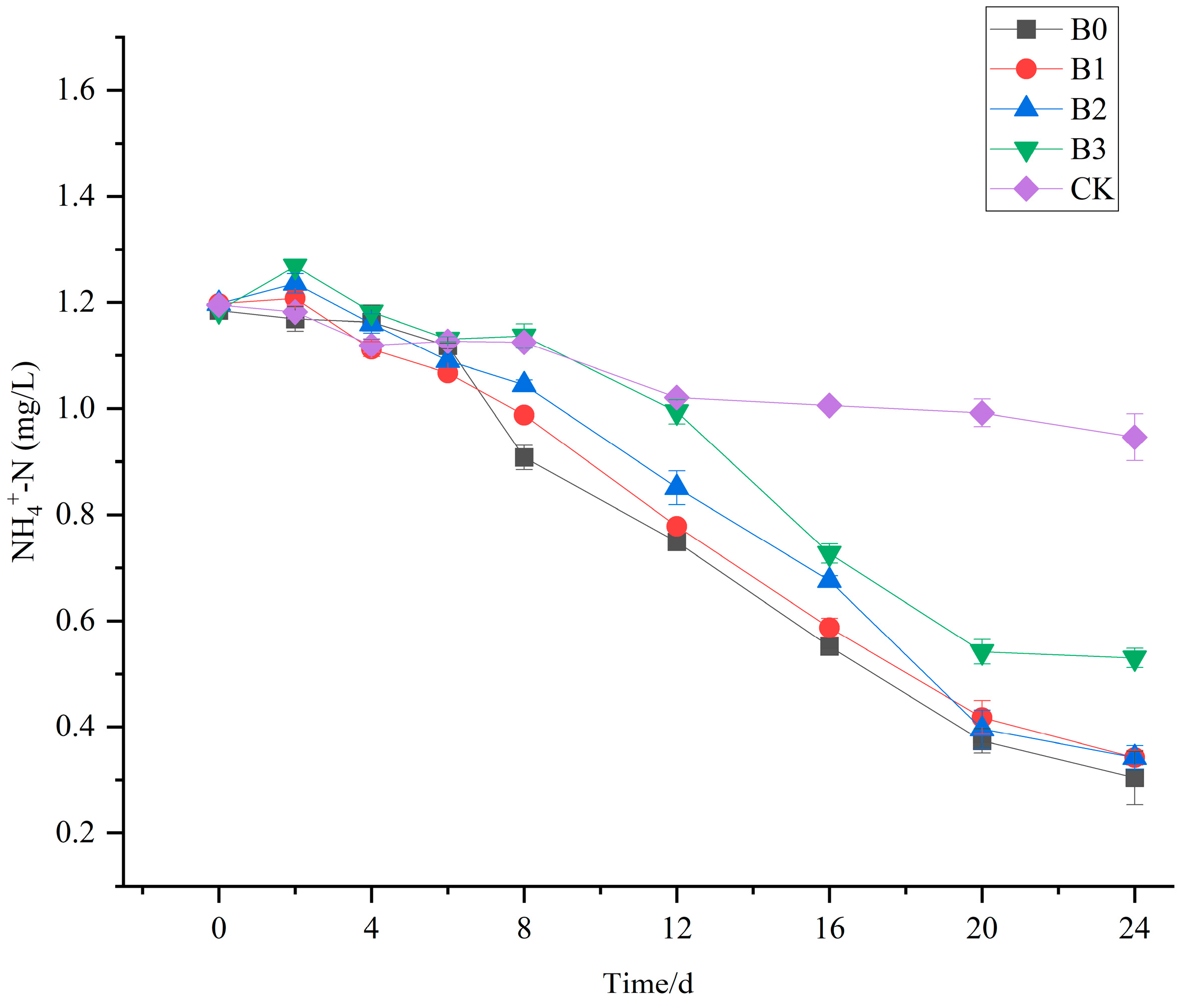
3.6. The Purification Effect of V. natans, B. aeruginosa, and H. cumingii Configuration on NH4+-N in Water Bodies
3.7. The Purification Effect of V. natans, B. aeruginosa, and H. cumingii Configuration on TP in Water Bodies
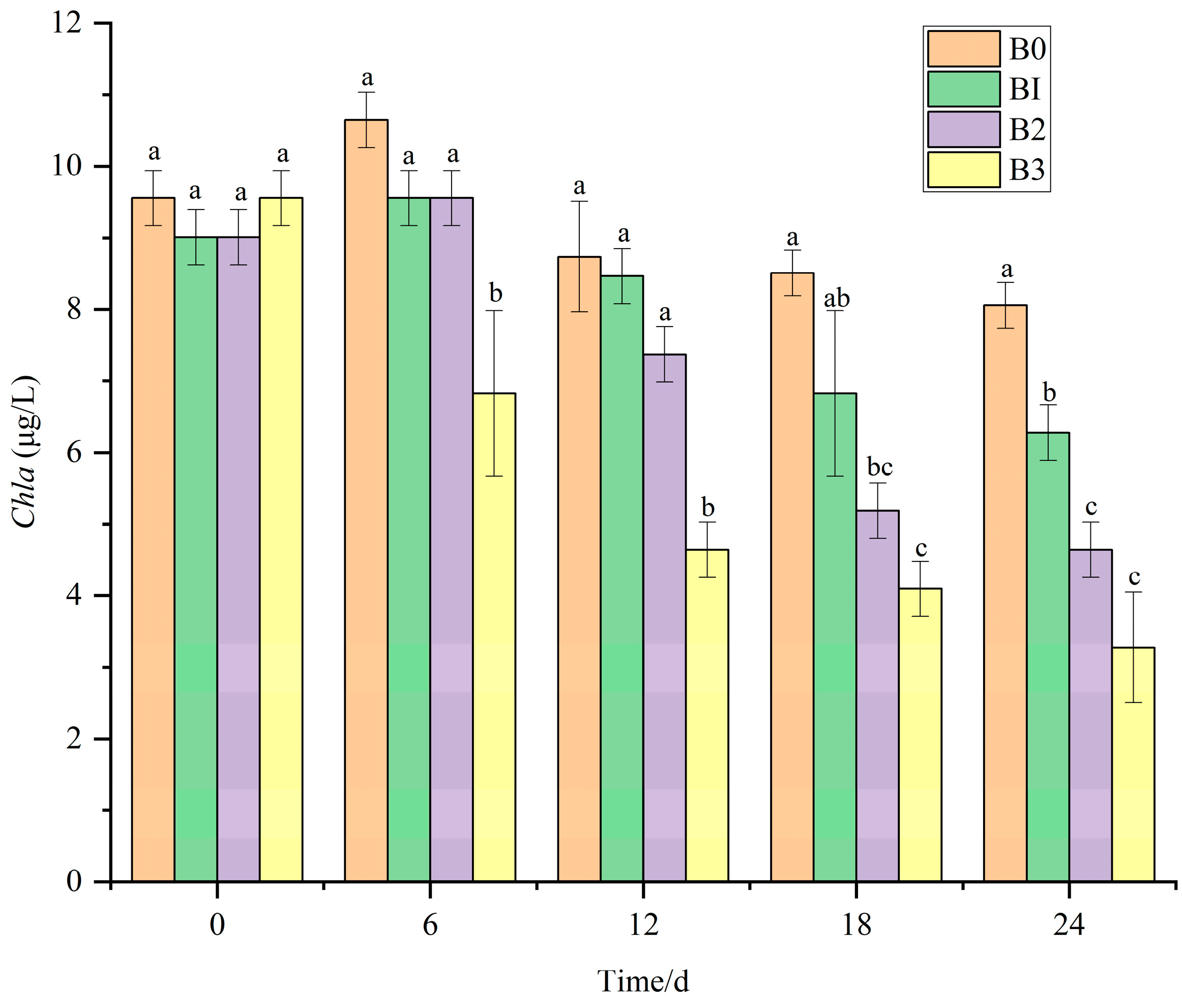
3.8. The Removal Effect of V. natans, B. aeruginosa, and H. cumingii Configuration on Chla in Water Bodies
4. Discussion
4.1. Mechanism of Nutrient Removal in V. natans and B. aeruginosa Configuration
4.2. Mechanism of Nutrient Removal in V. natans, B. aeruginosa, and H. cumingii Configuration
4.3. Interactions and Synergistic Restoration Effects among Organisms
5. Conclusions
- In the V. natans and B. aeruginosa configuration, the optimal biomass ratio of 15:2 resulted in the highest total nitrogen removal rate of 55.86%, ammonia nitrogen removal of 74.28%, and total phosphorus removal of 91.61%. The B. aeruginosa can scrape the attached algae on the leaves of V. natans, promote the photosynthesis of V. natans, and improve the absorption capacity of V. natans to nutrients in the water body.
- In the V. natans, B. aeruginosa, and H. cumingii configuration, the optimal biomass ratio of 15:2:10 achieved a total nitrogen removal rate of 69.75%, ammonia nitrogen removal of 71.45%, and a Chla removal rate of 50.48%, showing significant inhibition of phytoplankton.
- The V. natans, B. aeruginosa, and H. cumingii configuration demonstrated superior water quality compared to the V. natans and B. aeruginosa configuration, with a 13.89% increase in total nitrogen removal following the addition of H. cumingii.
- After adding the H. cumingii to the system, there was no significant effect on the removal of ammonia nitrogen in the water body, and the total phosphorus concentration showed a changing trend of first increasing and then decreasing; the higher the density of the H. cumingii, the greater the upward trend. Therefore, in the practical application stage, the density of H. cumingii should be carefully selected.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ministry of Ecology and Environment of the People’s Republic of China. China Ecological Environment Status Bulletin 2021; Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China, 2022. (In Chinese)
- Sun, G.; Sheng, L.X. Ecological engineering for eutrophication control in lake. Chin. J. Appl. Ecol. 2001, 12, 590–592. (In Chinese) [Google Scholar]
- Wang, M.; Zhang, H.; Zeng, H.X.; Song, X.; Zhou, S.; Li, Y.Q. Research progress on the causes, current status and remediation technology ofwater eutrophication. J. Anhui Agric. Sci. 2022, 50, 1–6+11. (In Chinese) [Google Scholar]
- Tang, P.; Yu, L.J.; Peng, Z.X.; Fan, P.Y.; Li, T.M.; Ren, K.Y. Research progresses on algae inhibition by allelopathy of aquatic plants. J. Biol. 2021, 38, 104–108. (In Chinese) [Google Scholar]
- Zhang, Q.; Song, Q.; Zhu, Y.; Cui, X.H. Research on overwintering abilities of seven submerged plants under artificial conditions. J. Shanghai Jiao Tong Univ. Agric. Sci. 2015, 33, 85–89. (In Chinese) [Google Scholar]
- Liu, J.K.; Xie, P. Direct control of microcystis bloom through the use of planktivorous carp-closure experiments and lake fishery. Ecol. Sci. 2003, 22, 193–196. (In Chinese) [Google Scholar]
- Zhou, L.H.; Gu, X.H.; Zeng, Q.F.; Mao, Z.G.; Gao, H.M. Comparative purification effect of different density snail and macrophytes on aquaculture tail water. Resour. Enviroment Yangtze Basin 2011, 20, 173–178. (In Chinese) [Google Scholar]
- Chen, L.S.; Hong, H.P.; Hong, R.H.; Liu, Q.G. Discussion on water purification function and rational fishery management of silver carp and bighead carp in Qiandao Lake. Aquat. Technol. Intell. 2005, 6, 259–261. (In Chinese) [Google Scholar]
- Covich, A.P.; Palmer, M.A.; Crowl, T.A. The role of benhic invertebrate species in freshwater ecosystems: Zoobenthic species influence energy flows and nutrient cycling. BioScience 1999, 49, 119–127. [Google Scholar] [CrossRef]
- Wang, M.; Du, X.H.; Jin, K.; Cai, X.H.; Hu, X.M. Study on purifying effect of different proportions of hydrophytes and benthic organisms on water quality in Lakeside Wetland. Acta Agric. Jiangxi 2016, 28, 70–73. (In Chinese) [Google Scholar]
- Gao, H.; Qian, X.; Wu, H.; Li, H.; Pan, H.; Han, C. Combined effects of submerged macrophytes and aquatic animals on the restoration of a eutrophic water body-a case study of Gonghu Bay, Lake Taihu. Ecol. Eng. 2017, 102, 15–23. [Google Scholar] [CrossRef]
- Szabó, S.; Peeters, E.T.H.M.; Várbíró, G.; Borics, G.; Lukács, B.A. Phenotypic plasticity as a clue for invasion success of the submerged aquatic plant Elodea nuttallii. Plant Biol. 2019, 21, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Li, K.Y.; Wen, M.Z.; Yang, H.W.; Li, Y.M.; Liu, Z.W. Mutualistic relationship between freshwater sanils aquatic macrophytes. Acta Ecol. Sin. 2007, 27, 5427–5432. (In Chinese) [Google Scholar]
- Li, P.; Zhang, X.F.; Mo, S.Q. Effects of mussel (Anodonta woodiana), submerged macrophyte (Vallisneria natans) and their coexistence on water quality. Chin. J. Ecol. 2016, 35, 1589–1594. (In Chinese) [Google Scholar]
- Xu, M.Y.; Liu, W.; Tan, G.W.; Huang, S.B.; Wu, J.H.; Lu, Y.F.; Chen, P. Effect of aquatics combination system in purifying eutrophic scenic waters. J. Yangtze River Sci. Reasearch Inst. 2019, 36, 27–31. (In Chinese) [Google Scholar]
- Li, Y.; Li, Z.N.; Yang, Y.H.; Lin, J.X.; Sun, L.P. Study on ecological remediation effect of urban landscape water quality improvement: A case study of Shiwan Park Landscape Lake in Foshan. Guangdong Chem. Ind. 2021, 48, 117–120. (In Chinese) [Google Scholar]
- Singh, N.K.; Gourevitch, J.D.; Wemple, B.C.; Watson, K.B.; Rizzo, D.M.; Polasky, S.; Ricketts, T.H. Optimizing wetland restoration to improve water quality at a regional scale. Environ. Res. Lett. 2019, 14, 064006. [Google Scholar] [CrossRef]
- GB3838-2002; Environmental Quality Standards for Surface Water. China Standards Press: Beijing, China, 2002. (In Chinese)
- State Environmental Protection Administration. Analytical Methods for Water and Wastewater Monitoring, 4th ed.; China Environmental Science Press: Beijing, China, 2002; pp. 101–280. (In Chinese) [Google Scholar]
- Wu, J. Effect of Shading and High Temperature Treatment on the Morphology and Physiology of Three Vanilla Plants; Jiangxi Agricultural University: Nanchang, China, 2022. (In Chinese) [Google Scholar]
- Li, P.M.; Gao, H.Y.; Reto, J.S. Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study. J. Plant Physiol. Mol. Biol. 2005, 31, 559–566. (In Chinese) [Google Scholar]
- Huang, Z.X.; Zhang, Y.J.; Ma, H.F.; Wang, C.; Luo, S.T.; Huo, H.C.; Dong, Y. A comparative study on removal efficiency of four submerged plants on phosphorus and nitrogen in eutrophic water under different planting density. Ecol. Sci. 2011, 30, 102–106. (In Chinese) [Google Scholar]
- Quiroz-Flores, A.D.J.; Miranda-Arce, M.G.; Ramírez-García Armora, P.; Lot-Helgueras, A. Phosphorus accumulation within the sediments of hydrophytes communities in the Tochac lake, Hidalgo, Mexico. Polibotánica 2019, 46, 233–240. [Google Scholar] [CrossRef][Green Version]
- Zhao, D.; Xu, W.J.; Li, Y.; Chen, H. Purification of eutrophicated water by Vallisneria spinulosa. Chin. Agric. Sci. Bull. 2010, 26, 189–192. (In Chinese) [Google Scholar]
- Zhang, Q. Effect of Copper Rust and Aurx on Water Quality and Attached Bacterial Diversity. Master’s Thesis, Hebei University, Baoding, China, 2020. (In Chinese). [Google Scholar]
- Li, X.J.; He, S.Z.; Chang, X.X.; Tan, F.G.; Shen, Y.; Lu, F.T.; Hou, W.T. Role of snails, mussels and submerged plants in purifying micro-polluted water. Chin. J. Environ. Eng. 2016, 10, 95–102. (In Chinese) [Google Scholar]
- Wei, Y.C.; Pu, P.M. Abatement of the nitrogen and phosphorus concent-ration in water by the snail Bellamya aeruginosa in Lake Taihu. Resour. Environ. Yangtze Basin 1999, 1, 88–90. (In Chinese) [Google Scholar]
- Li, M.; Zhang, K.F.; Ren, J.; Chen, S.F.; Xie, P.L. Test of water purification in the reservoir for benthic animals. In Proceedings of the 2019 Chinese Society of Environmental Science Annual Conference of Science and Technology-Environmental Engineering Technology Innovation and Application Sub-Forum, Xi’an, China, 24 August 2019. (In Chinese). [Google Scholar]
- Ma, S.B.; Zhu, J.Y.; Lu, K.H.; Yang, W. Impact of freshwater gastropod (Bellamya aeruginosa) on the zooplankton community in the water body experiencing a cyanobacterial bloom. Chin. J. Ecol. 2016, 35, 1872–1878. (In Chinese) [Google Scholar]
- Li, Q.G.; Wei, T.; Zhang, G.M.; Xian, G. Effects of Hyriopsis cumingii on nitrogen and phosphorus releases from sediment and on wetland microorganisms in Baiyangdian Wetland. Res. Environ. Sci. 2020, 33, 2318–2325. (In Chinese) [Google Scholar]
- Salgado, J.; Sayer, C.D.; Brooks, S.J.; Davidson, T.A.; Okamura, B. Connectivity and zebra mussel invasion offer short-term buffering of eutrophication impacts on floodplain lake landscape biodiversity. Divers. Distrib. 2019, 25, 1334–1347. [Google Scholar] [CrossRef]
- Smith, R.; Higgins, S.N.; Vander Zanden, M.J.; Joppa, L.N.; Vadeboncoeur, Y. The effect of dreissenid invasions on chlorophyll and the chlorophyll: Total phosphorus ratio in north-temperate lakes. Can. J. Fish. Aquat. Sci. 2011, 68, 319–329. [Google Scholar]
- Zhang, X.F.; Liu, Z.W.; Jeppesen, E.; Taylor, W.D. Effects of deposit-feeding tubificid worms and filter-feeding bivalves on benthic-pelagic coupling: Implications for the restoration of eutrophic shallow lakes. Water Res. 2014, 50, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Han, S.S.; Wen, Y.M. Phosphorus release and affecting factors in the sediments of eutrophic water. Chin. J. Ecol. 2004, 2, 98–101. (In Chinese) [Google Scholar]
- Liu, J.S.; Shi, H.; Li, Y.Y. Effect of Cd2+ stress on photosynthetic and chlorophyll fluorescence properties in cucumber seedlings. Res. Soil Water Conserv. 2011, 18, 187–190+196. (In Chinese) [Google Scholar]
- Zhang, A.L.; Pan, Y.Z.; Jiang, B.B.; Liu, Q. Effect of shading on photosynthetic and chlorophyll fluorescence parameters in Lilium spp. J. Southwest China Norm. Univ. 2011, 36, 163–167. (In Chinese) [Google Scholar]
- Gao, L.N. Comparison of chlorophyll fluorescence characteristics between two submerged macrophytes in Jiuzhaigou National Nature Reserve. Acta Agric. Zhejiangensis 2017, 29, 951–958. (In Chinese) [Google Scholar]
- Xu, H.J.; Ling, Q.F.; Yang, C.G.; Lin, J.H.; Wu, L.K.; Lu, J.M. Preliminary study on the elimination effects of algae by three species of freshwater bivalve. J. Hydroecol. 2010, 31, 72–75. (In Chinese) [Google Scholar]
- Hu, L.; Pan, X.J.; Peng, J.H.; Zou, X.; Wan, C.Y. In situ enclosure test on the effect of Hyriopsis cumingii hanging culture on eutrophication control in water bodies. In Proceedings of the 2016 Annual Meeting of the Chinese Society for Environmental Sciences, Haikou, China, 14 October 2016. (In Chinese). [Google Scholar]
- Anesio, A.M.; Abreu, P.C.; Biddanda, B.A. The role of free and attached microorganisms in the decomposition of estuarine macrophyte detritus. Estuar. Coast. Shelf Sci. 2003, 56, 197–201. [Google Scholar] [CrossRef]
- Steffan, S.A.; Chikaraishi, Y.; Dharampal, P.S.; Pauli, J.N.; Guédot, C.; Ohkouchi, N. Unpacking brown food-webs: Animal trophic identity reflects rampant microbivory. Ecol. Evol. 2017, 7, 3532–3541. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.D.; Taylor, B.R. Abrupt population increase by the leaf-shredding caddisfly Pycnopsyche guttifer profoundly changes leaf litter decomposition in streams of eastern Canada. Fundam. Appl. Limnol. 2021, 195, 153–171. [Google Scholar] [CrossRef]
- Tang, J.Y.; Cao, P.P.; Xu, C.; Liu, M.S. Effects of aquatie plants during their decayand decomposition on water quality. Chin. J. Appl. Ecol. 2013, 24, 83–89. (In Chinese) [Google Scholar]
- Wu, S.; He, S.; Zhou, W.; Gu, J.; Huang, J.; Gao, L.; Zhang, X. Decomposition characteristics of three different kinds of aquatic macrophytes and their potential application as carbon resource in constructed wetland. Environ. Pollut. 2017, 231, 1122–1133. [Google Scholar] [CrossRef]
- Zhuang, M.F.; He, S.B.; Liu, W.R. Research on nitrogen enhanced removal by Iris pseudacorus residual self-consumption constructed wetlands. Technol. Water Treat. 2019, 45, 106–112. (In Chinese) [Google Scholar]
- Xian, Q.M.; Chen, H.D.; Liu, H.L.; Zou, H.X.; Yin, D.Q. Isolation and identification of antialgal compounds from the leaves of Vallisneria spiralis L. by activity-guided fractionation. Environ. Sci. Pollut. Res. 2006, 13, 233–237. [Google Scholar] [CrossRef]
- Gao, Y.N.; Liu, B.Y.; Wang, J.; He, F.; Liang, W.; Xu, D.; Zhang, L.P.; Wu, Z.B. Allelopathic effects of phenolic compounds released by Vallisneria spiralis on Microcystis aeruginosa. J. Lake Sci. 2011, 23, 761–766. (In Chinese) [Google Scholar]
- Qin, B.Q.; Gao, G.; Hu, W.P.; Wu, Q.L.; Hu, C.H.; Liu, Z.W.; Gu, X.H.; Zhu, G.W.; Chen, F.Z. Reflections on the theory and practice of shallow lake ecosystem restoration. J. Lake Sci. 2005, 17, 9–16. (In Chinese) [Google Scholar]
- Zhong, Y.X.; Hu, H.Y.; Qian, Y. Effect of environment and nutrient factors on the content of nitrogen and phosphorus in two duckweeds species Spirodela polyrrhiza and Lemna aequinoctialis. Environ. Sci. 2005, 26, 67–71. (In Chinese) [Google Scholar]
- Wang, Y.L.; Xiao, Y.; Pan, H.Y.; Fu, C.Z.; Gao, S.X. Analysis of nutrient composition and comprehensive utilization of submersed aquatic macrophytes (Vallisneria natans). J. Ecol. Rural Environ. 2006, 22, 45–47. (In Chinese) [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Han, Z.; Jiang, R.; Cai, Q.; Zhang, M.; Wang, R.; Liu, Y.; Shao, L.; Wu, M.; Zhang, J.; et al. Research on the Ecological Restoration Effects of a Vallisneria natans (Lour.) Hara-Dominated Multitrophic Level Ecosystem. Water 2024, 16, 1050. https://doi.org/10.3390/w16071050
Li B, Han Z, Jiang R, Cai Q, Zhang M, Wang R, Liu Y, Shao L, Wu M, Zhang J, et al. Research on the Ecological Restoration Effects of a Vallisneria natans (Lour.) Hara-Dominated Multitrophic Level Ecosystem. Water. 2024; 16(7):1050. https://doi.org/10.3390/w16071050
Chicago/Turabian StyleLi, Bin, Zheng Han, Ruitong Jiang, Qingjie Cai, Meiqiong Zhang, Rong Wang, Yuchao Liu, Liu Shao, Meiqin Wu, Jianheng Zhang, and et al. 2024. "Research on the Ecological Restoration Effects of a Vallisneria natans (Lour.) Hara-Dominated Multitrophic Level Ecosystem" Water 16, no. 7: 1050. https://doi.org/10.3390/w16071050
APA StyleLi, B., Han, Z., Jiang, R., Cai, Q., Zhang, M., Wang, R., Liu, Y., Shao, L., Wu, M., Zhang, J., He, W., Liu, J., & He, P. (2024). Research on the Ecological Restoration Effects of a Vallisneria natans (Lour.) Hara-Dominated Multitrophic Level Ecosystem. Water, 16(7), 1050. https://doi.org/10.3390/w16071050