
Relative Contribution of Fungal Communities to Carbon Loss and Humification Process in Algal Sludge Aerobic Composting

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Materials, Design, and Sampling Collection
2.2. Determination of Compost Samples
2.3. Bioinformatic Analyses
2.4. Statistical Analyses
3. Results and Discussion
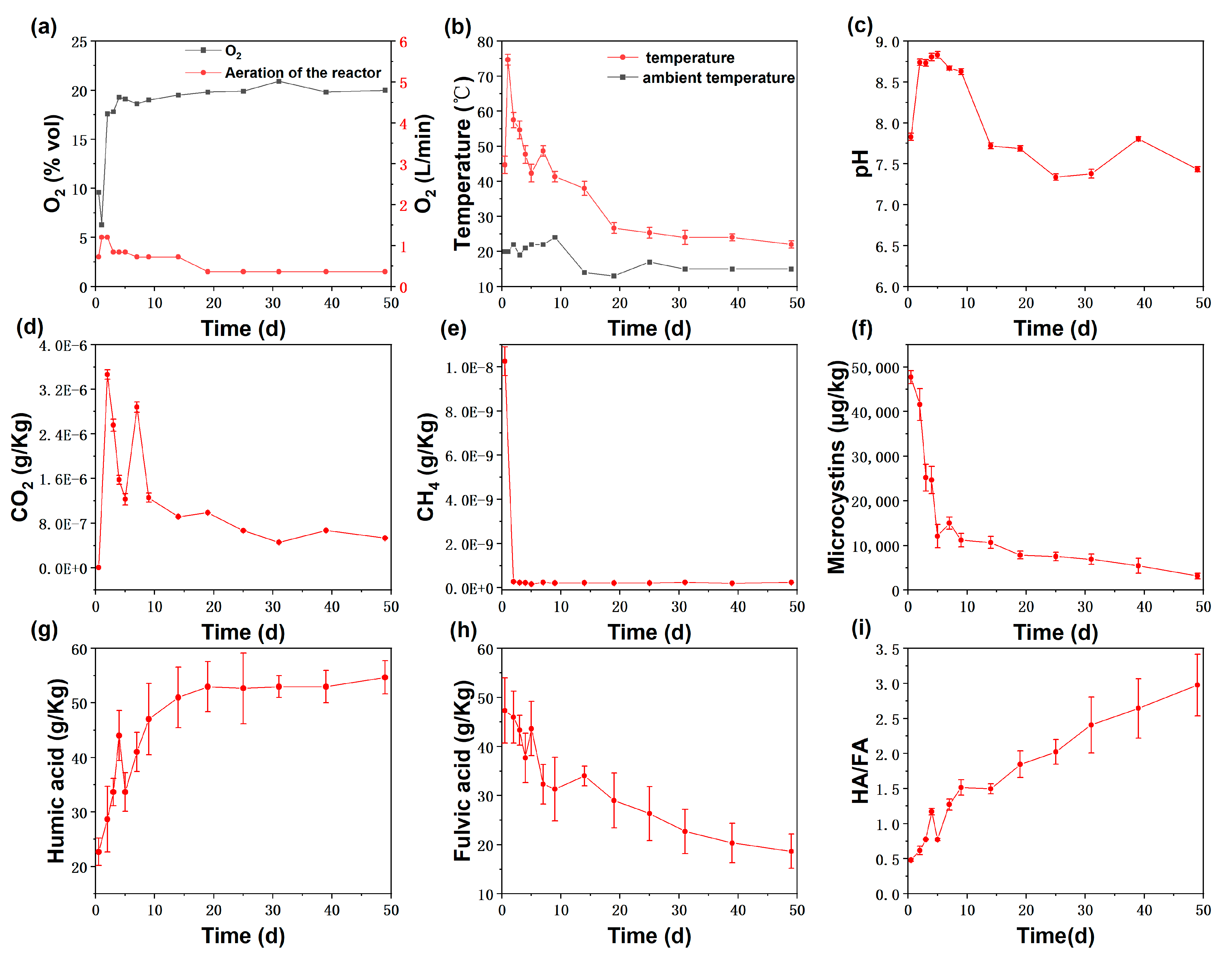
3.1. Physicochemical Changes during Composting
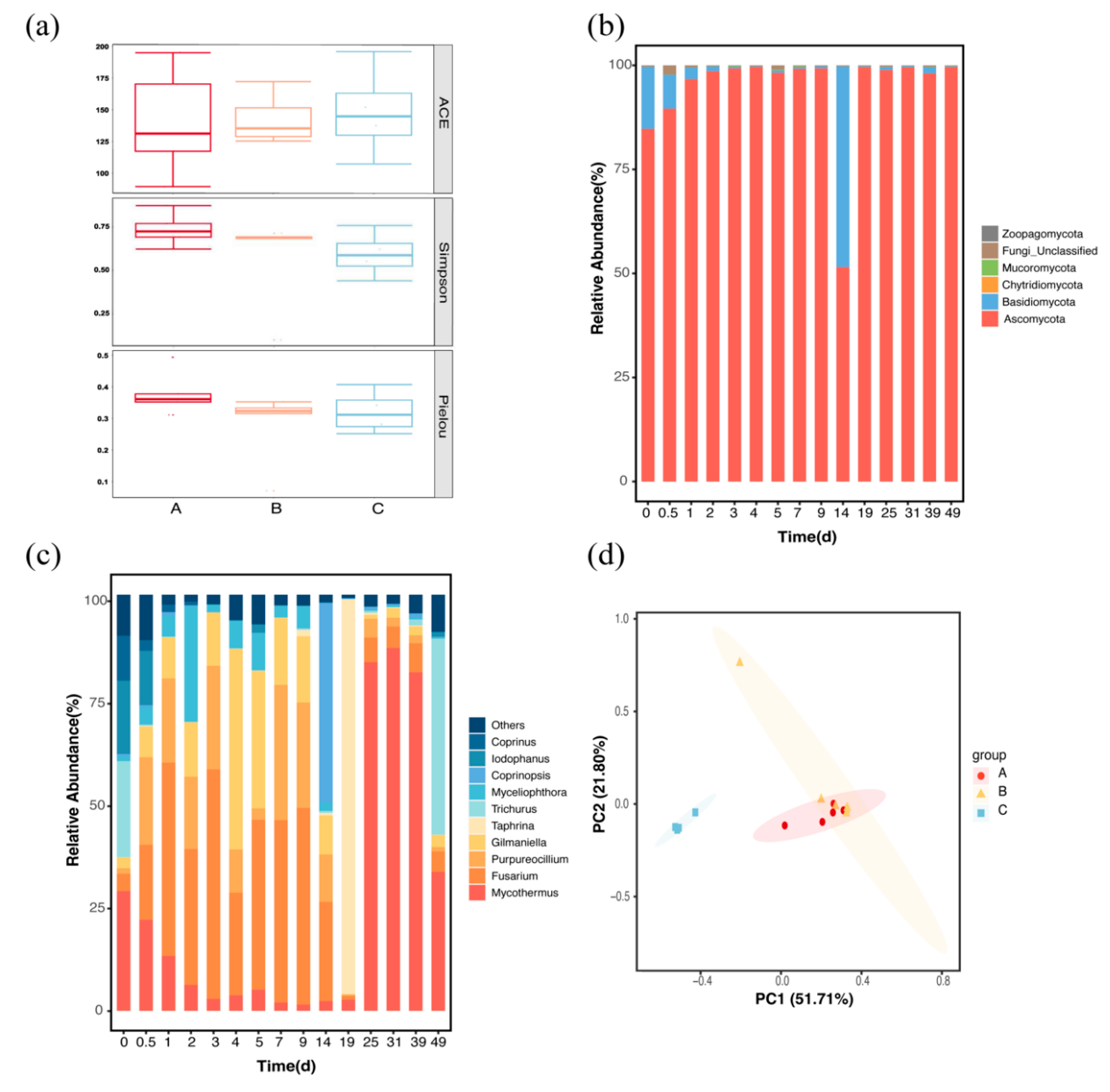
3.2. Dynamics of Fungal Communities
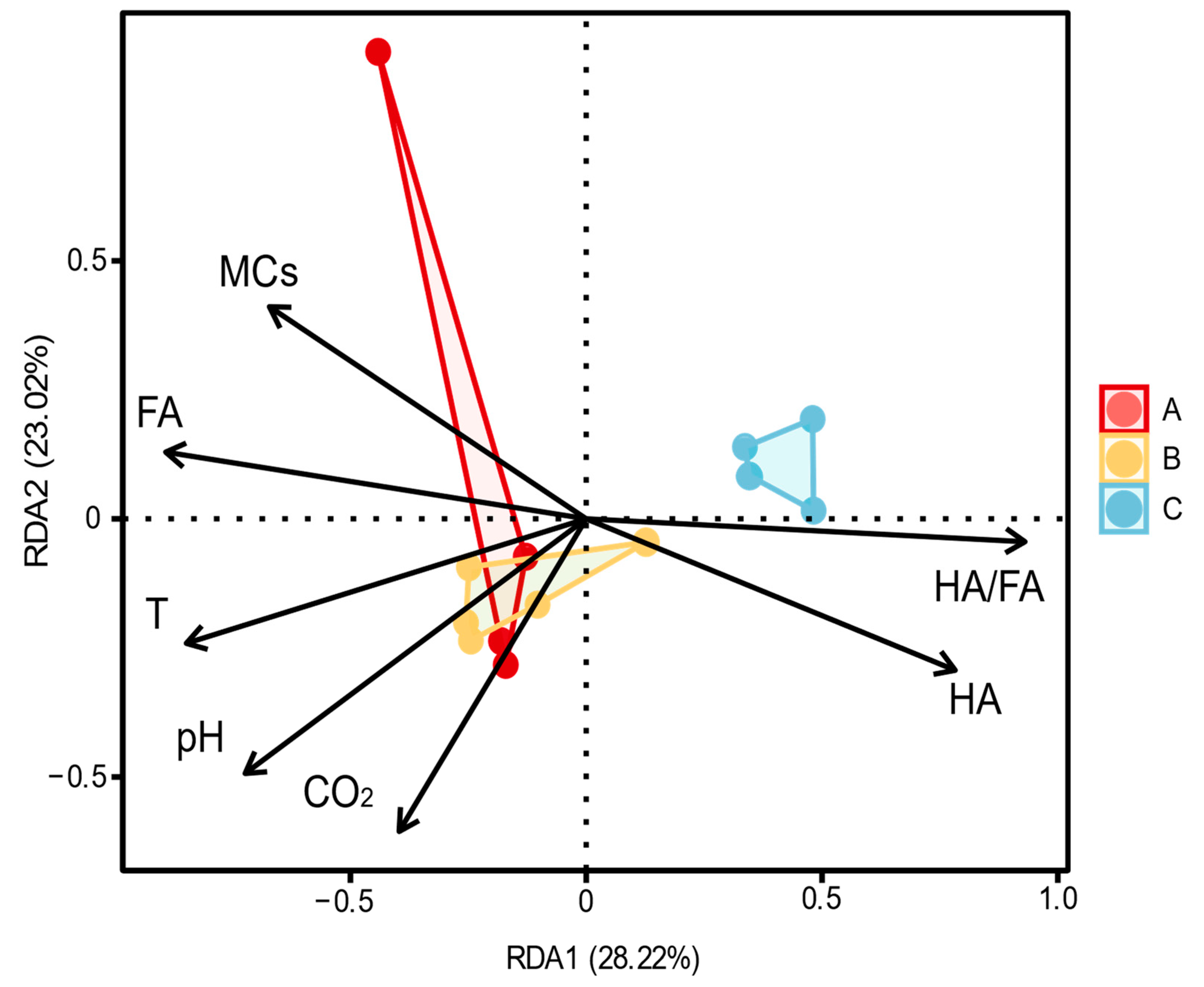
3.3. Relationship between Physicochemical Properties and Fungal Communities
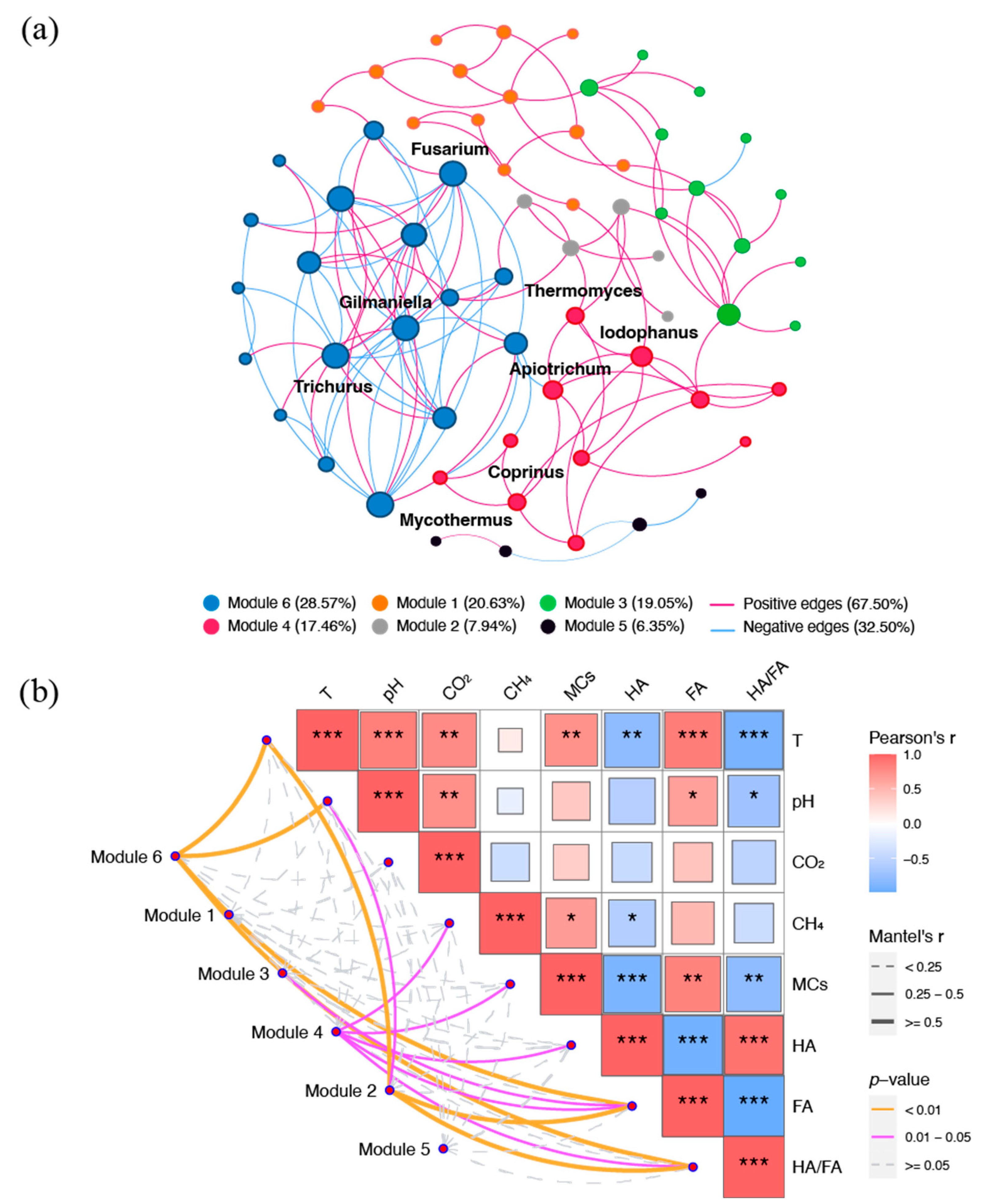
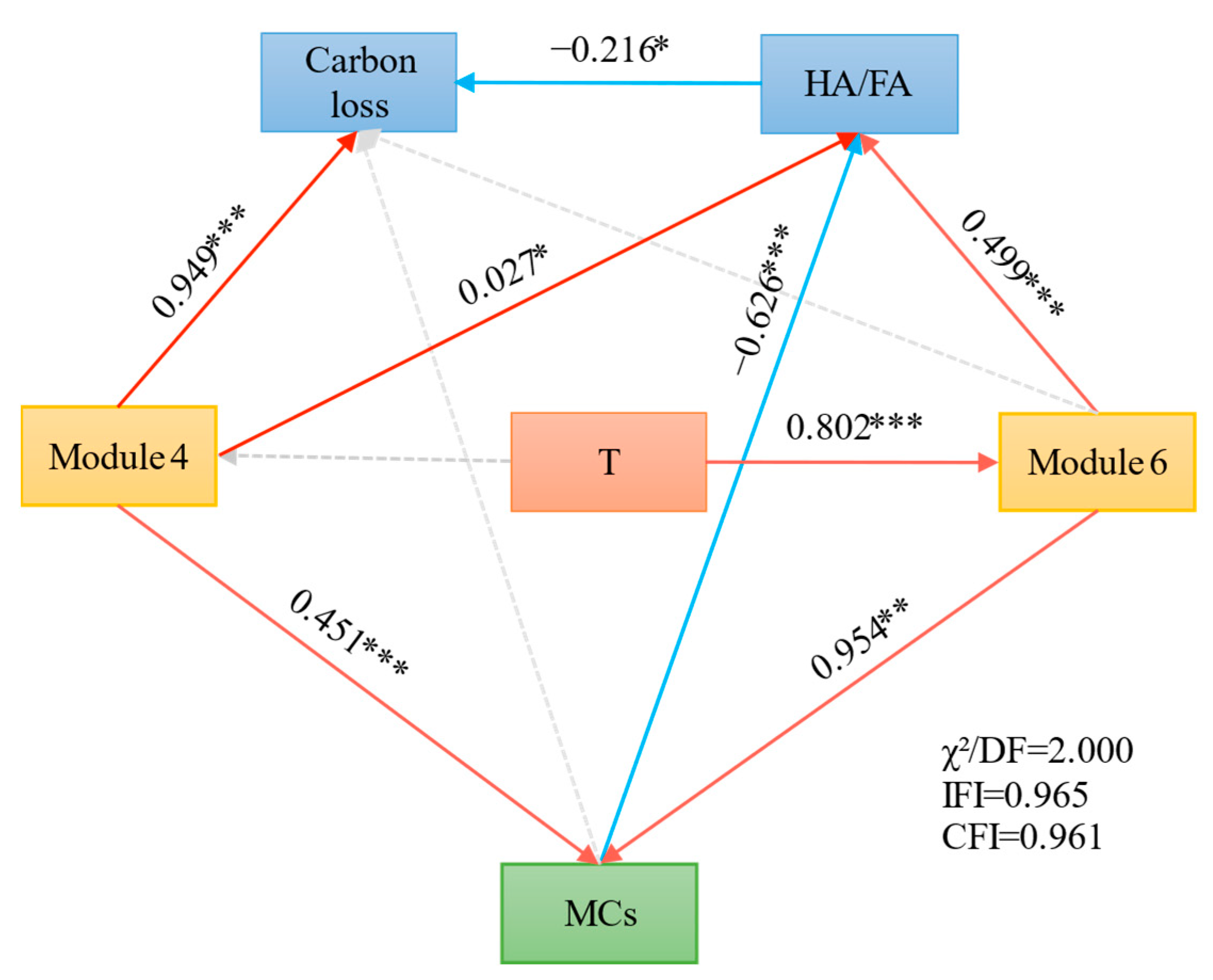
3.4. Relative Contributions of Potential Factors to Carbon Loss and Humification Process
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhou, J.; Leavitt, P.R.; Zhang, Y.; Qin, B. Anthropogenic Eutrophication of Shallow Lakes: Is It Occasional? Water Res. 2022, 221, 118728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hu, M.; Bian, B.; Yang, Z.; Yang, W.; Zhang, L. Full-Scale Thermophilic Aerobic Co-Composting of Blue-Green Algae Sludge with Livestock Faeces and Straw. Sci. Total Environ. 2021, 753, 142079. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Wang, W.; Xu, Q.; Liu, Z.; Teng, J.; Yan, H.; Liu, X. Microcystins in Water: Detection, Microbial Degradation Strategies, and Mechanisms. Int. J. Environ. Res. Public Health 2022, 19, 13175. [Google Scholar] [CrossRef] [PubMed]
- Kumar Awasthi, S.; Verma, S.; Liu, T.; Kumar Awasthi, M.; Zhang, Z.; Syed, A.; Bahkali, A.H. Regulation of Fungal Communities during Pig Manure Composting. Bioresour. Technol. 2023, 389, 129823. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sun, S.; Pei, F.; Zhang, C.; Cao, X.; Kang, J.; Wu, Z.; Ling, H.; Ge, J. Response Characteristics of Flax Retting Liquid Addition during Chicken Manure Composting: Focusing on Core Bacteria in Organic Carbon Mineralization and Humification. Bioresour. Technol. 2023, 381, 129112. [Google Scholar] [CrossRef] [PubMed]
- Bui, V.K.H.; Truong, H.B.; Hong, S.; Li, X.; Hur, J. Biotic and Abiotic Catalysts for Enhanced Humification in Composting: A Comprehensive Review. J. Clean. Prod. 2023, 402, 136832. [Google Scholar] [CrossRef]
- Hwang, H.Y.; Kim, S.H.; Kim, M.S.; Park, S.J.; Lee, C.H. Co-Composting of Chicken Manure with Organic Wastes: Characterization of Gases Emissions and Compost Quality. Appl. Biol. Chem. 2020, 63, 3. [Google Scholar] [CrossRef]
- Ye, P.; Fang, L.; Song, D.; Zhang, M.; Li, R.; Awasthi, M.K.; Zhang, Z.; Xiao, R.; Chen, X. Insights into Carbon Loss Reduction during Aerobic Composting of Organic Solid Waste: A Meta-Analysis and Comprehensive Literature Review. Sci. Total Environ. 2023, 862, 160787. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-H.; Chen, X.-H.; Li, Q.-F.; Lü, H.; Mo, C.-H.; Feng, N.-X.; Xiang, L.; Zhao, H.-M.; Li, H.; Li, Y.-W.; et al. Fungal Community Enhanced Humification and Influenced by Heavy Metals in Industrial-Scale Hyperthermophilic Composting of Municipal Sludge. Bioresour. Technol. 2022, 360, 127523. [Google Scholar] [CrossRef]
- Duan, Y.; Awasthi, S.K.; Liu, T.; Chen, H.; Zhang, Z.; Wang, Q.; Ren, X.; Tu, Z.; Awasthi, M.K.; Taherzadeh, M.J. Dynamics of Fungal Diversity and Interactions with Environmental Elements in Response to Wheat Straw Biochar Amended Poultry Manure Composting. Bioresour. Technol. 2019, 274, 410–417. [Google Scholar] [CrossRef]
- Xie, G.; Kong, X.; Kang, J.; Su, N.; Fei, J.; Luo, G. Fungal Community Succession Contributes to Product Maturity during the Co-Composting of Chicken Manure and Crop Residues. Bioresour. Technol. 2021, 328, 124845. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Luo, H.; Zhao, D.; Qiao, Z.; Zheng, J.; An, M.; Huang, D. Environmental Factors and Interactions among Microorganisms Drive Microbial Community Succession during Fermentation of Nongxiangxing Daqu. Bioresour. Technol. 2022, 345, 126549. [Google Scholar] [CrossRef]
- Meng, Q.; Liu, S.; Guo, Y.; Hu, Y.; Yu, Z.; Bello, A.; Wang, Z.; Xu, W.; Xu, X. The Co-Occurrence Network Patterns and Keystone Species of Microbial Communities in Cattle Manure-Corn Straw Composting. Environ. Sci. Pollut. Res. 2022, 30, 20265–20276. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Xu, C.; Wu, F. Microbial Co-Occurrence Networks Driven by Low-Abundance Microbial Taxa during Composting Dominate Lignocellulose Degradation. Sci. Total Environ. 2022, 845, 157197. [Google Scholar] [CrossRef] [PubMed]
- Wu, X. Isolation and Characterization of a Novel Thermotolerant Alkali Lignin-Degrading Bacterium Aneurinibacillus sp. LD3 and Its Application in Food Waste Composting. Chemosphere 2022, 307, 135859. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Liu, B.; Xi, C.; Luo, X.; Yuan, X.; Wang, X.; Zhu, W.; Wang, H.; Cui, Z. Effect of Pig Manure on the Chemical Composition and Microbial Diversity during Co-Composting with Spent Mushroom Substrate and Rice Husks. Bioresour. Technol. 2018, 251, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Bi, X.; Xu, Y.; Yang, Z.; Wei, T.; Xi, M.; Li, J.; Chen, L.; Li, H.; Sun, S. Variation in Community Structure and Network Characteristics of Spent Mushroom Substrate (SMS) Compost Microbiota Driven by Time and Environmental Conditions. Bioresour. Technol. 2022, 364, 127915. [Google Scholar] [CrossRef] [PubMed]
- West, S.G.; Taylor, A.B.; Wu, W. Model Fit and Model Selection in Structural Equation Modeling. Handb. Struct. Equ. Model. 2012, 1, 209–231. [Google Scholar]
- Ma, C.; Hu, B.; Wei, M.-B.; Zhao, J.-H.; Zhang, H.-Z. Influence of Matured Compost Inoculation on Sewage Sludge Composting: Enzyme Activity, Bacterial and Fungal Community Succession. Bioresour. Technol. 2019, 294, 122165. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Luo, W.; Lu, J.; Zhang, X.; Li, S.; Wu, Y.; Li, G. Effects of Digestion Time in Anaerobic Digestion on Subsequent Digestate Composting. Bioresour. Technol. 2018, 267, 117–125. [Google Scholar] [CrossRef]
- Cai, R.; Zuo, S.; Cao, X.; Xu, C. Carbon and Nitrogen Transformation Mechanism and Functional Microorganisms of Sheep Manure Composting on the Qinghai-Tibet Plateau under Different Moisture Content. J. Environ. Chem. Eng. 2023, 11, 111341. [Google Scholar] [CrossRef]
- Meng, X.; Liu, B.; Zhang, H.; Wu, J.; Yuan, X.; Cui, Z. Co-Composting of the Biogas Residues and Spent Mushroom Substrate: Physicochemical Properties and Maturity Assessment. Bioresour. Technol. 2019, 276, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, R.; Malińska, K.; Marfà, O. Nitrification within Composting: A Review. Waste Manag. 2018, 72, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shen, Y.; Ding, J.; Luo, W.; Zhou, H.; Cheng, H.; Wang, H.; Zhang, X.; Wang, J.; Xu, P.; et al. High Oil Content Inhibits Humification in Food Waste Composting by Affecting Microbial Community Succession and Organic Matter Degradation. Bioresour. Technol. 2023, 376, 128832. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Su, Y.; He, X.; Han, L.; Huang, G. Effects of Functional Membrane Coverings on Carbon and Nitrogen Evolution during Aerobic Composting: Insight into the Succession of Bacterial and Fungal Communities. Bioresour. Technol. 2023, 369, 128463. [Google Scholar] [CrossRef]
- Smidt, E.; Meissl, K.; Schmutzer, M.; Hinterstoisser, B. Co-Composting of Lignin to Build up Humic Substances—Strategies in Waste Management to Improve Compost Quality. Ind. Crops Prod. 2008, 27, 196–201. [Google Scholar] [CrossRef]
- Chen, L.; Chen, Y.; Li, Y.; Liu, Y.; Jiang, H.; Li, H.; Yuan, Y.; Chen, Y.; Zou, B. Improving the Humification by Additives during Composting: A Review. Waste Manag. 2023, 158, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, Y.; Wang, R.; Lu, Q.; Wu, J.; Zhang, D.; Nie, Z.; Wei, Z. Effect of the Addition of Exogenous Precursors on Humic Substance Formation during Composting. Waste Manag. 2018, 79, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Selvam, A.; Wong, J.W.C. Evaluation of Humic Substances during Co-Composting of Food Waste, Sawdust and Chinese Medicinal Herbal Residues. Bioresour. Technol. 2014, 168, 229–234. [Google Scholar] [CrossRef]
- Zhu, N.; Zhu, Y.; Kan, Z.; Li, B.; Cao, Y.; Jin, H. Effects of Two-Stage Microbial Inoculation on Organic Carbon Turnover and Fungal Community Succession during Co-Composting of Cattle Manure and Rice Straw. Bioresour. Technol. 2021, 341, 125842. [Google Scholar] [CrossRef]
- Chen, Z.; Fu, Q.; Wen, Q.; Wu, Y.; Bao, H.; Guo, J. Microbial Community Competition Rather than High-Temperature Predominates ARGs Elimination in Swine Manure Composting. J. Hazard. Mater. 2022, 423, 127149. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, M.K.; Pandey, A.K.; Khan, J.; Bundela, P.S.; Wong, J.W.C.; Selvam, A. Evaluation of Thermophilic Fungal Consortium for Organic Municipal Solid Waste Composting. Bioresour. Technol. 2014, 168, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Su, C.; Mo, T.; Liao, L.; Zhu, F.; Chen, Y.; Chen, M. Effect of Excess Sludge and Food Waste Feeding Ratio on the Nutrient Fractions, and Bacterial and Fungal Community during Aerobic Co-Composting. Bioresour. Technol. 2021, 320, 124339. [Google Scholar] [CrossRef] [PubMed]
- Bilgrami, A.L.; Khan, A. Biopesticidal Potentials of Predaceous and Parasitic Fungi. In Plant Nematode Biopesticides; Elsevier: Amsterdam, The Netherlands, 2022; pp. 141–163. ISBN 978-0-12-823006-0. [Google Scholar]
- Yuan, J.; Zhou, J.-Y.; Li, X.; Dai, C.-C. The Primary Mechanism of Endophytic Fungus Gilmaniella sp. AL12 Promotion of Plant Growth and Sesquiterpenoid Accumulation in Atractylodes Lancea. Plant Cell Tissue Organ Cult. PCTOC 2016, 125, 571–584. [Google Scholar] [CrossRef]
- Ma, S.; Shen, Y.; Ding, J.; Cheng, H.; Zhou, H.; Ge, M.; Wang, J.; Cheng, Q.; Zhang, D.; Zhang, Y.; et al. Effects of Biochar and Volcanic Rock Addition on Humification and Microbial Community during Aerobic Composting of Cow Manure. Bioresour. Technol. 2024, 391, 129973. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ma, L.; Zhao, B.; Li, Y.; Chen, Y.; Deng, Y.; Wang, Y. Humification Process Enhancement through Relative Abundance Promotion of Talaromyces and Coprinopsis by Inoculated Phanerochaete Chrysosporium during the Secondary Fermentation of Composting. Environ. Sci. Pollut. Res. 2022, 30, 9060–9065. [Google Scholar] [CrossRef] [PubMed]
- Basotra, N.; Kaur, B.; Di Falco, M.; Tsang, A.; Chadha, B.S. Mycothermus Thermophilus (Syn. Scytalidium Thermophilum): Repertoire of a Diverse Array of Efficient Cellulases and Hemicellulases in the Secretome Revealed. Bioresour. Technol. 2016, 222, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Sharma, S. Composting of a Crop Residue through Treatment with Microorganisms and Subsequent Vermicomposting. Bioresour. Technol. 2002, 85, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-P.; Wang, L.; Sun, Z.-Y.; Wang, S.-T.; Shen, C.-H.; Tang, Y.-Q.; Kida, K. Biochar Addition Reduces Nitrogen Loss and Accelerates Composting Process by Affecting the Core Microbial Community during Distilled Grain Waste Composting. Bioresour. Technol. 2021, 337, 125492. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, K.; Wang, J.; Wang, Z.; Pan, R.; Wang, Q.; Li, S.; Kumar, S.; Zhang, Z.; Li, R. Potential of Biochar Integrated Manganese Sulfate for Promoting Pig Manure Compost Humification and Its Biological Mechanism. Bioresour. Technol. 2022, 357, 127350. [Google Scholar] [CrossRef]
- Deng, L.; Zhao, M.; Bi, R.; Bello, A.; Uzoamaka Egbeagu, U.; Zhang, J.; Li, S.; Chen, Y.; Han, Y.; Sun, Y.; et al. Insight into the Influence of Biochar on Nitrification Based on Multi-Level and Multi-Aspect Analyses of Ammonia-Oxidizing Microorganisms during Cattle Manure Composting. Bioresour. Technol. 2021, 339, 125515. [Google Scholar] [CrossRef] [PubMed]
- Bello, A.; Wang, B.; Zhao, Y.; Yang, W.; Ogundeji, A.; Deng, L.; Egbeagu, U.U.; Yu, S.; Zhao, L.; Li, D.; et al. Composted Biochar Affects Structural Dynamics, Function and Co-Occurrence Network Patterns of Fungi Community. Sci. Total Environ. 2021, 775, 145672. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.E.J. Modularity and Community Structure in Networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef]
- Zhou, N.; Zhang, Y.; Liu, F.; Cai, L. Halophilic and Thermotolerant Gymnoascus Species from Several Special Environments, China. Mycologia 2016, 108, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Awasthi, M.K.; Zhang, Z.; Syed, A.; Bahkali, A.H.; Sindhu, R.; Verma, M. Evaluation of Fungal Dynamics during Sheep Manure Composting Employing Peach Shell Biochar. Bioresour. Technol. 2023, 386, 129559. [Google Scholar] [CrossRef] [PubMed]
- Da Rosa-Garzon, N.G.; Laure, H.J.; Rosa, J.C.; Cabral, H. Valorization of Agricultural Residues Using Myceliophthora Thermophila as a Platform for Production of Lignocellulolytic Enzymes for Cellulose Saccharification. Biomass Bioenergy 2022, 161, 106452. [Google Scholar] [CrossRef]
- Wang, J.; Jiao, M.; Zhan, X.; Hu, C.; Zhang, Z. Humification and Fungal Community Succession during Pig Manure Composting: Membrane Covering and Mature Compost Addition. Bioresour. Technol. 2024, 393, 130030. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Jiang, Y.-Q.; Jiang, G.-M.; Ou, J.-Y.; Zeng, L.-T.; Zhang, H.-H.; Chen, D.-Q.; Jiang, Y.-T. Molecular Identification and Biological Characteristic Analysis of an Apiotrichum Mycotoxinivorans (Formerly Trichosporon Mycotoxinivorans) Strain Isolated from Sputum Specimens of a Pediatric Patient with Pneumonia. J. Mycol. Médicale 2019, 29, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Tuomela, M. Biodegradation of Lignin in a Compost Environment: A Review. Bioresour. Technol. 2000, 72, 169–183. [Google Scholar] [CrossRef]
- Zhuo, R.; Fan, F. A Comprehensive Insight into the Application of White Rot Fungi and Their Lignocellulolytic Enzymes in the Removal of Organic Pollutants. Sci. Total Environ. 2021, 778, 146132. [Google Scholar] [CrossRef]
- Piao, M.; Li, A.; Du, H.; Sun, Y.; Du, H.; Teng, H. A Review of Additives Use in Straw Composting. Environ. Sci. Pollut. Res. 2023, 30, 57253–57270. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xu, Y.; Zhang, J.; Bello, A.; Li, X.; Liu, W.; Egbeagu, U.U.; Zhao, L.; Han, Y.; Cheng, L.; et al. Investigation of Underlying Links between Nitrogen Transformation and Microorganisms’ Network Modularity in the Novel Static Aerobic Composting of Dairy Manure by “Stepwise Verification Interaction Analysis. Sci. Total Environ. 2023, 883, 163674. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Awasthi, M.K.; Liu, T.; Awasthi, S.K.; Syed, A.; Bahkali, A.H.; Verma, M.; Zhang, Z. Influence of Biochar on Succession of Fungal Communities during Food Waste Composting. Bioresour. Technol. 2023, 385, 129437. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zheng, J.; Jiang, C.; Zhang, F.; Wei, L.; Zhu, L. Hydrochloric Acid-Modified Algal Biochar for the Removal of Microcystis Aeruginosa: Coagulation Performance and Mechanism. J. Environ. Chem. Eng. 2022, 10, 108903. [Google Scholar] [CrossRef]
- Han, S.; Li, J.; Zhou, Q.; Liu, G.; Wang, T. Harmless Disposal and Resource Utilization of Wastes from the Lake in China: Dewatering, Composting and Safety Evaluation of Fertilizer. Algal Res. 2019, 43, 101623. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Zhang, S.; Zhou, J.; Cong, H.; Feng, S.; Sun, F. Relative Contribution of Fungal Communities to Carbon Loss and Humification Process in Algal Sludge Aerobic Composting. Water 2024, 16, 1084. https://doi.org/10.3390/w16081084
Wu H, Zhang S, Zhou J, Cong H, Feng S, Sun F. Relative Contribution of Fungal Communities to Carbon Loss and Humification Process in Algal Sludge Aerobic Composting. Water. 2024; 16(8):1084. https://doi.org/10.3390/w16081084
Chicago/Turabian StyleWu, Hainan, Sen Zhang, Jiahui Zhou, Haibing Cong, Shaoyuan Feng, and Feng Sun. 2024. "Relative Contribution of Fungal Communities to Carbon Loss and Humification Process in Algal Sludge Aerobic Composting" Water 16, no. 8: 1084. https://doi.org/10.3390/w16081084
APA StyleWu, H., Zhang, S., Zhou, J., Cong, H., Feng, S., & Sun, F. (2024). Relative Contribution of Fungal Communities to Carbon Loss and Humification Process in Algal Sludge Aerobic Composting. Water, 16(8), 1084. https://doi.org/10.3390/w16081084