
Diversity of Macrophytes and Macroinvertebrates in Different Types of Standing Waters in the Drava Field




Abstract
1. Introduction
2. Materials and Methods
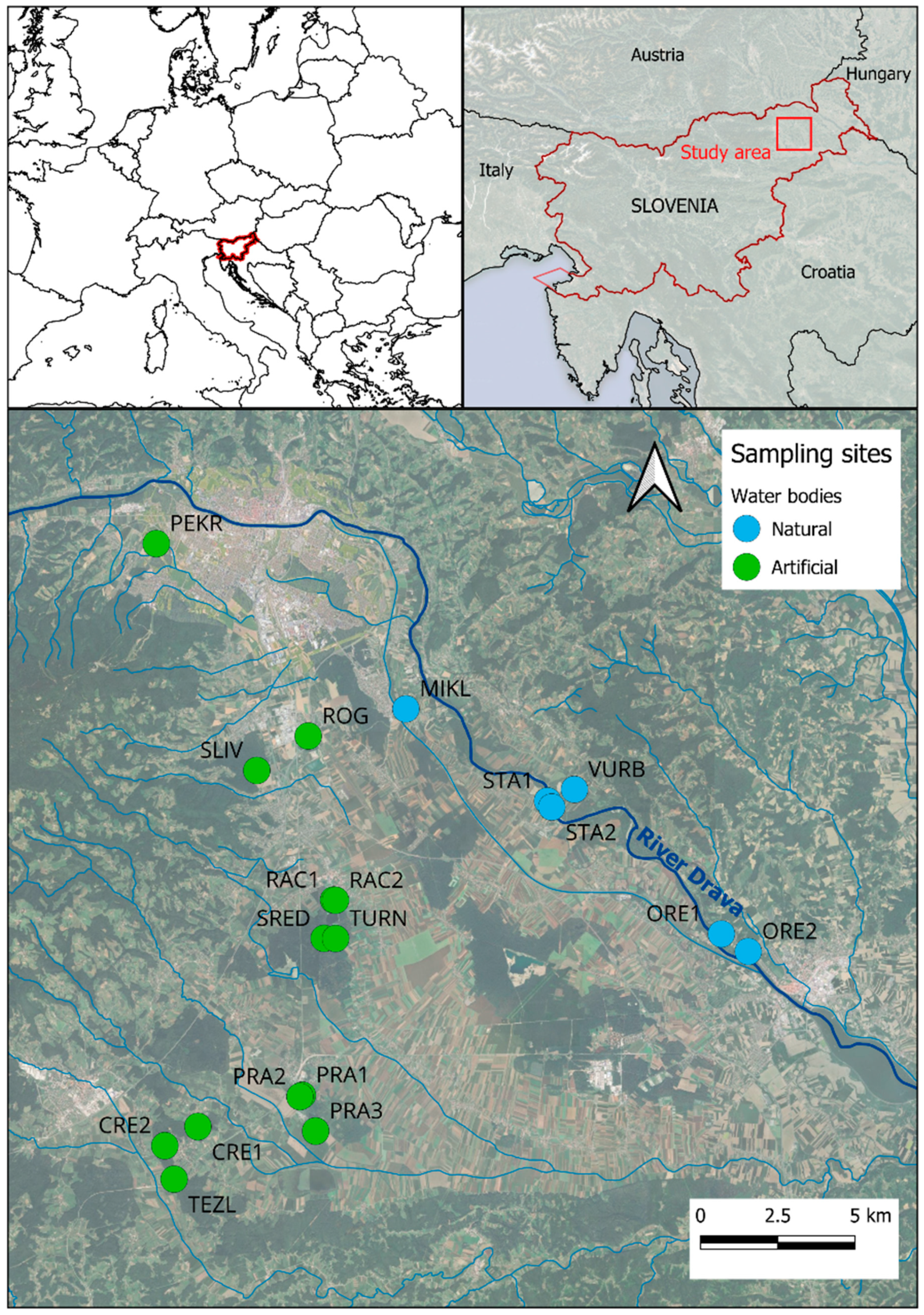
2.1. Study Area
2.2. Field Surveys

2.3. Data Analyses
3. Results
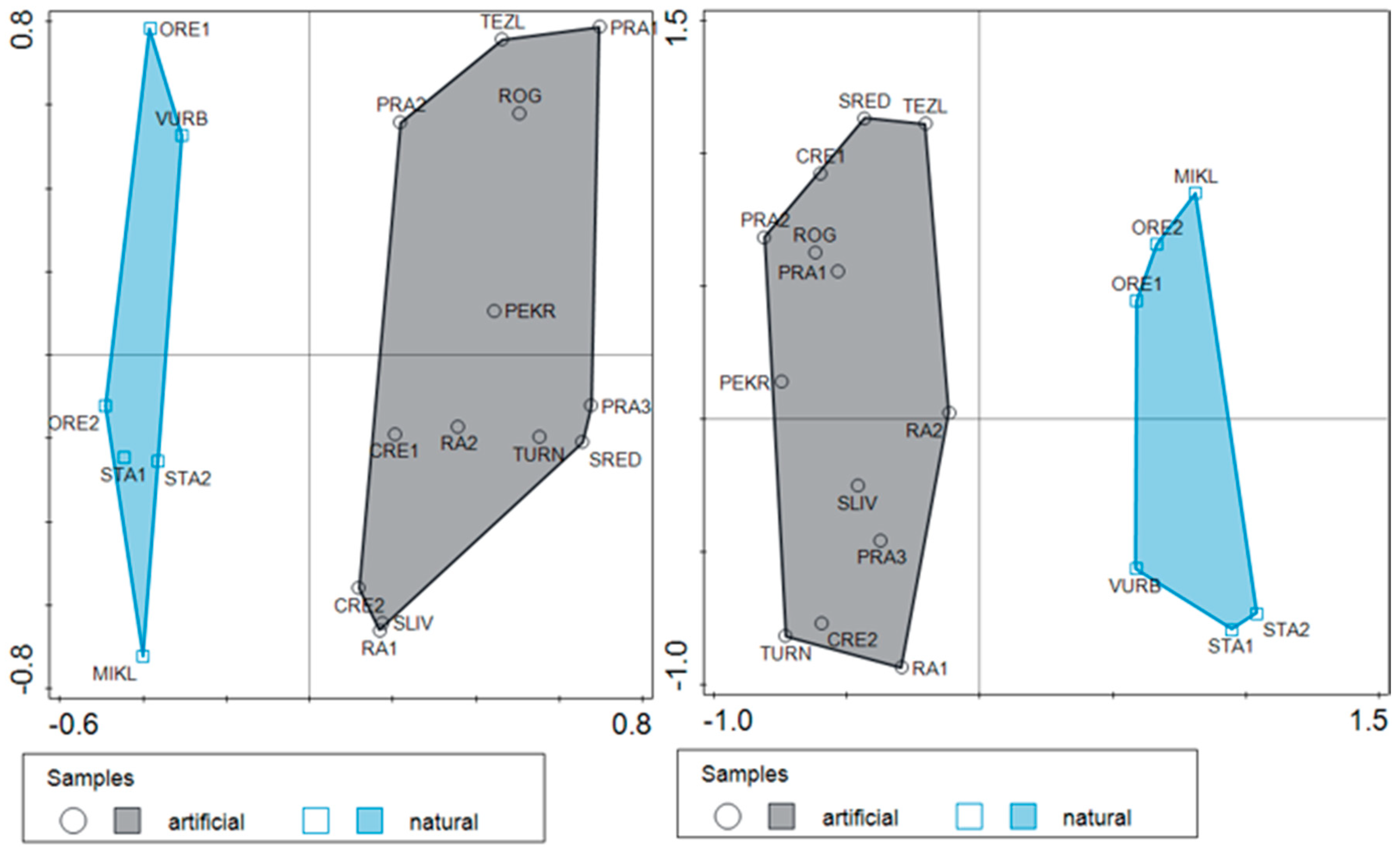
3.1. Diversity Patterns in Natural and Artificial Ponds
3.2. Differentiation in the Taxonomic Composition of Communities in the Artificial and Natural Ponds
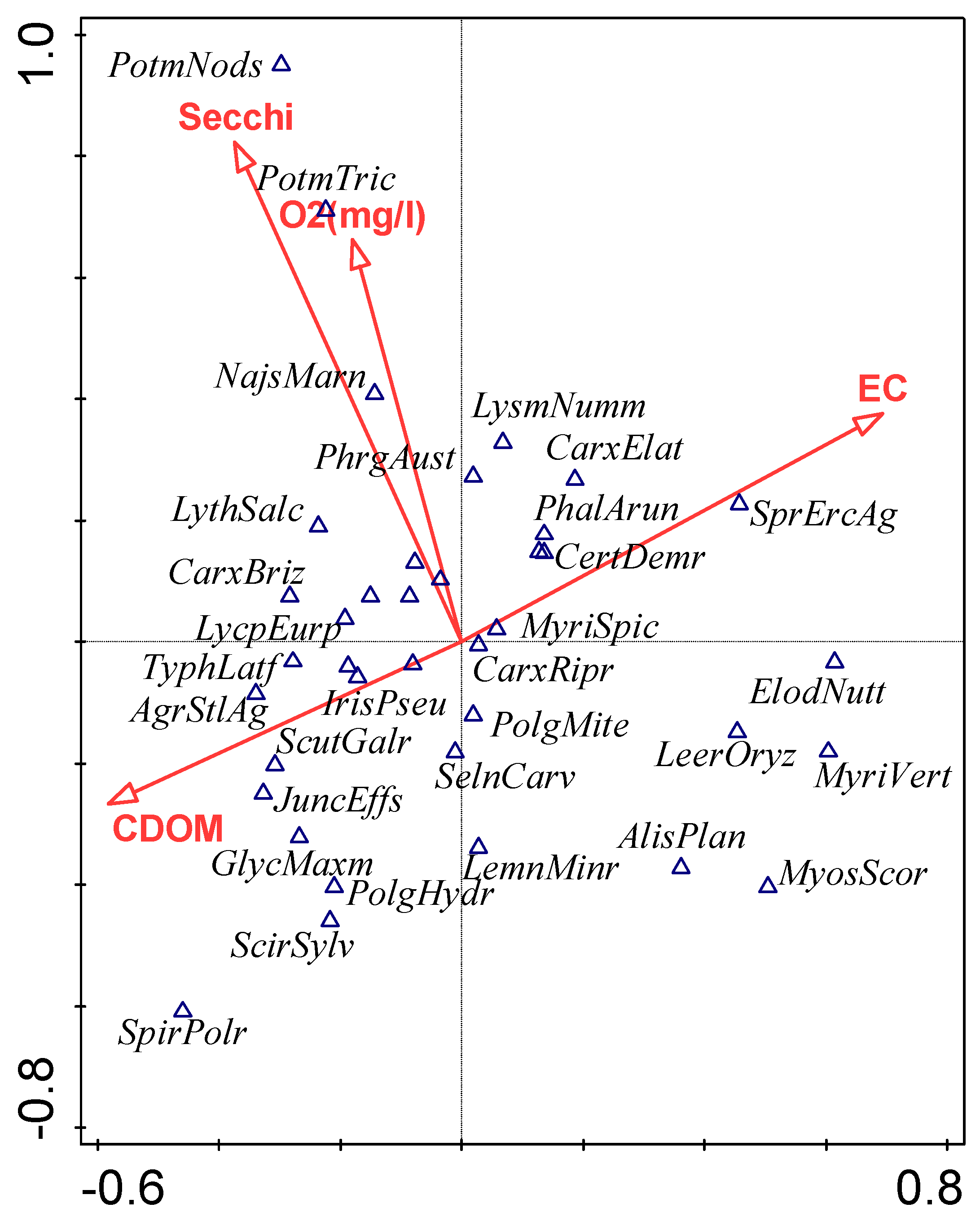
3.3. Influence of Environmental Factors on Macrophyte Community Composition
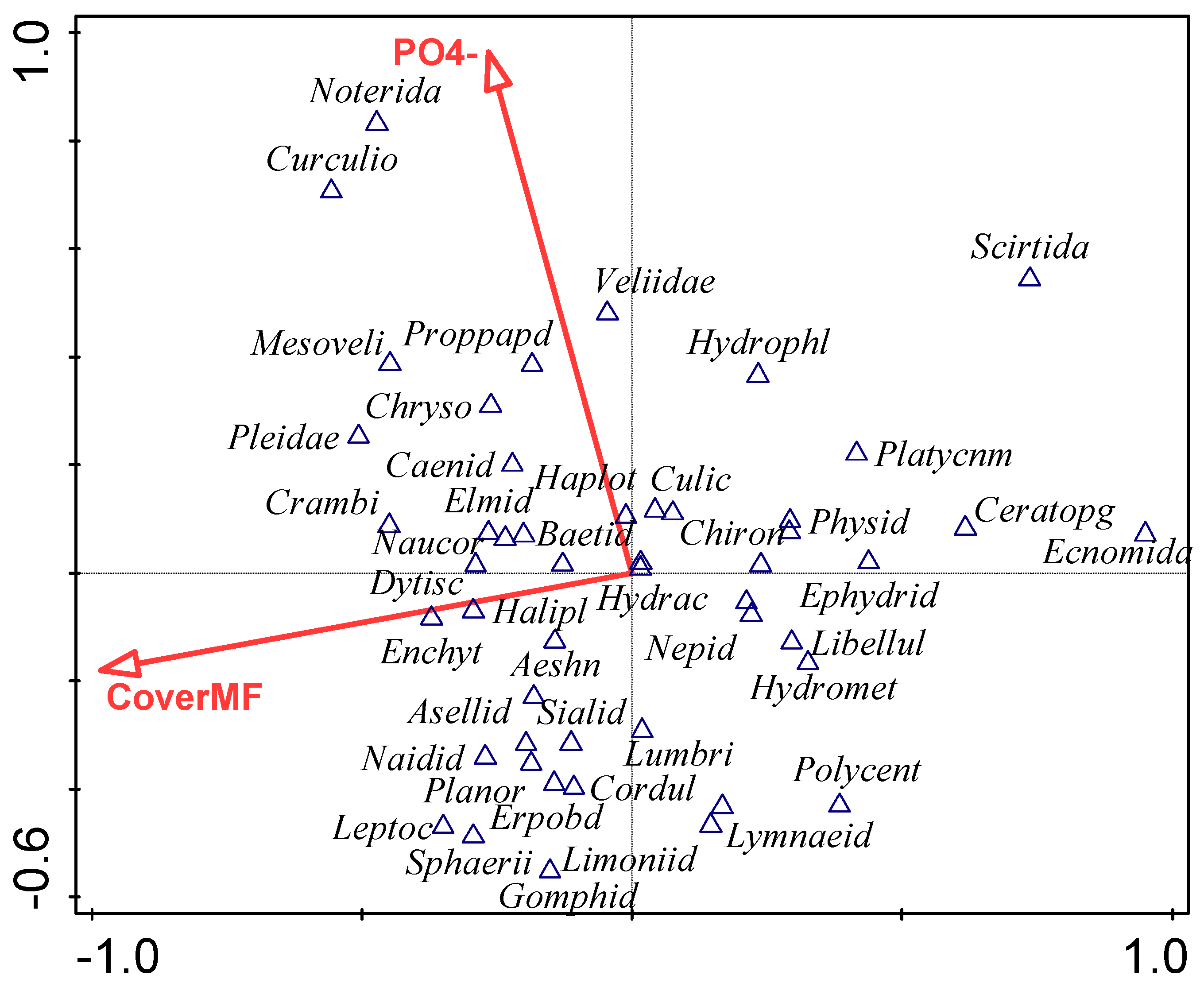
3.4. Influence of Environmental Factors on Macroinvertebrate Community
3.5. Influence of Environmental Factors on the Diversity of Macroinvertebrate Community
3.6. Influence of Macrophyte Taxa on the Macroinvertebrate Communities
4. Discussion
4.1. Diversity Patterns in Natural and Artificial Ponds
4.2. Different Taxonomic Composition of Communities in the Artificial and Natural Ponds
4.3. Influence of Environmental Factors on Macrophyte Community Composition
4.4. Influence of Environmental Factors on Macroinvertebrate Community Composition
4.5. Influence of Environmental Factors on the Diversity of Macroinvertebrate Community
4.6. Influence of Macrophyte Taxa on the Macroinvertebrate Communities
4.7. Monitoring and Management
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, P.; Whitfield, M.; Biggs, J.; Bray, S.; Fox, G.; Nicolet, P.; Sear, D. Comparative biodiversity of rivers, streams, ditches and ponds in an agricultural landscape in Southern England. Biol. Conserv. 2004, 115, 329–341. [Google Scholar] [CrossRef]
- Hassal, C.; Hollinshead, J.; Hull, A. Environmental correlates of plant and invertebrate species richness in ponds. Biodivers. Conserv. 2011, 20, 3189–3222. [Google Scholar] [CrossRef]
- De Meester, L.; Declerck, S.; Stoks, R.; Louette, G.; Van De Meutter, F.; De Bie, T.; Michels, E.; Brendonck, L. Ponds and pools as model systems in conservation biology, ecology and evolutionary biology. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 715–725. [Google Scholar] [CrossRef]
- Davidson, N.C.; Finlayson, C.M. Extent, regional distribution and changes in area of different classes of wetland. Mar. Freshw. Res. 2018, 69, 1525–1533. [Google Scholar] [CrossRef]
- Bubíková, K.; Hrivnák, R. Artificial ponds in Central Europe do not fall behind the natural ponds in terms of macrophyte diversity. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 8. [Google Scholar] [CrossRef]
- Soomets, E.; Rannap, R.; Lõhmus, A. Patterns of assemblage structure indicate a broader conservation potential of focal amphibians for pond management. PLoS ONE 2016, 11, e0160012. [Google Scholar]
- Hill, M.J.; Biggs, J.; Thornhill, I.; Briers, R.A.; Ledger, M.; Gledhill, D.G.; Wood, P.J.; Hassall, C. Community heterogeneity of aquatic macroinvertebrates in urban ponds at a multi-city scale. Landsc. Ecol. 2018, 33, 389–405. [Google Scholar] [CrossRef]
- Cereghino, R.; Ruggiero, A.; Marty, P.; Angelibert, S. Biodiversity and distribution patterns of freshwater invertebrates in farm ponds of a south-western French agricultural landscape. Hydrobiologia 2008, 597, 43–51. [Google Scholar] [CrossRef]
- Davies, B.R.; Biggs, J.; Williams, P.J.; Lee, J.T.; Thompson, S. A comparison of the catchment sizes of rivers, streams, ponds, ditches and lakes: Implications for protecting aquatic biodiversity in an agricultural landscape. Hydrobiologia 2008, 597, 7–17. [Google Scholar] [CrossRef]
- Zelnik, I.; Gregorič, N.; Tratnik, A. Diversity of macroinvertebrates positively correlates with diversity of macrophytes in karst ponds. Ecol. Engin. 2018, 117, 96–103. [Google Scholar] [CrossRef]
- Ruhi, A.; Boix, D.; Sala, J.; Gascon, S.; Quintana, X.D. Spatial and temporal patterns of pioneer macrofauna in recently created ponds: Taxonomic and functional approaches. Hydrobiologia 2009, 634, 137–151. [Google Scholar] [CrossRef]
- Reyne, M.; Nolan, M.; McGuiggan, H.; Aubry, A.; Emmerson, M.; Marnell, F.; Reid, N. Artificial agri-environment scheme ponds do not replicate natural environments despite higher aquatic and terrestrial invertebrate richness and abundance. J. Appl. Ecol. 2020, 58, 304–314. [Google Scholar] [CrossRef]
- Zelnik, I.; Potisek, M.; Gaberščik, A. Environmental conditions and macrophytes of Karst ponds. Pol. J. Environ. Stud. 2012, 21, 1911–1920. [Google Scholar]
- Svitok, M.; Hrivnák, R.; Kochjarová, J.; Oťaheľová, H.; Paľove-Balang, P. Environmental thresholds and predictors of macrophyte species richness in aquatic habitats in central Europe. Folia Geobot. 2015, 51, 227–238. [Google Scholar] [CrossRef]
- Misteli, B.; Pannard, A.; Labat, F.; Fosso, L.K.; Baso, N.C.; Harpenslager, S.F.; Motitsoe, S.N.; Thiebaut, G.; Piscart, C. How Invasive Macrophytes Affect Macroinvertebrate Assemblages and Sampling Efficiency: Results from a Multinational Survey. Limnologica 2022, 96, 125998. [Google Scholar] [CrossRef]
- Germ, M. Biologija Vodnih Rastlin: Učbenik; Samozaložba: Ljubljana, Slovenia, 2013; 72p. [Google Scholar]
- Walker, P.D.; Wijnhoven, S.; van der Velde, G. Macrophyte presence and growth form influence macroinvertebrate community structure. Aquat. Bot. 2013, 104, 80–87. [Google Scholar] [CrossRef]
- Orozco-González, C.E.; Ocasio-Torres, M.E. Aquatic Macroinvertebrates as Bioindicators of Water Quality: A Study of an Ecosystem Regulation Service in a Tropical River. Ecologies 2023, 4, 209–228. [Google Scholar] [CrossRef]
- Kagalou, I.; Economidis, G.; Leonardos, I.; Papaloukas, C. Assessment of a Mediterranean shallow lentic ecosystem (Lake Pamvotis, Greece) using benthic community diversity: Response to environmental parameters. Limnologica 2006, 36, 269–278. [Google Scholar] [CrossRef]
- Bazzanti, M.; Coccia, C.; Dowgiallo, M.G. Microdistribution of macroinvertebrates in a temporary pond of Central Italy: Taxonomic and functional analyses. Limnologica 2010, 40, 291–299. [Google Scholar] [CrossRef]
- Moss, B.; Stephen, D.; Alvarez, C.; Becares, E.; van de Bund, W.; Collings, S.E.; van Donk, E.; de Eyto, E.; Feldmann, T.; Fernandez-Alaez, C.; et al. The determination of ecological status in shallow lakes—A tested system (ECOFRAME) for implementation of the European Water Framework Directive. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, 507–549. [Google Scholar] [CrossRef]
- Trigal, C.; Garcia-Criado, F.; Fernandez-Alaez, C. Towards a multimetric index for ecological assessment of Mediterranean flatland ponds: The use of macroinvertebrates as bioindicators. Hydrobiologia 2009, 618, 109–123. [Google Scholar] [CrossRef]
- Sand–Jensen, K.; Riis, T.; Vestergaard, O.; Larsen, S.E. Macrophyte decline in Danish lakes and streams over the past 100 years. J. Ecol. 2000, 88, 1030–1040. [Google Scholar] [CrossRef]
- Stendera, S.; Adrian, R.; Bonada, N.; Canedo–Arguelles, M.; Hugueny, B.; Januschke, K.; Pletterbauer, F.; Hering, D. Drivers and stressors of freshwater biodiversity patterns across different ecosystems and scales: A review. Hydrobiologia 2012, 696, 1–28. [Google Scholar] [CrossRef]
- Della Bella, V.; Bazzanti, M.; Chiarotti, F. Macroinvertebrate diversity and conservation status of Mediterranean ponds in Italy: Water permanence and mesohabitat influence. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 583–600. [Google Scholar] [CrossRef]
- Burdett, A.S.; Watts, R.J. Modifying living space: An experimental study of the influences of vegetation on aquatic invertebrate community structure. Hydrobiologia 2009, 618, 161–173. [Google Scholar] [CrossRef]
- Milesi, S.V.; Dolédec, S.; Melo, A.S. Substrate heterogeneity influences the trait composition of stream insect communities: An experimental in situ study. Freshw. Sci. 2016, 35, 1321–1329. [Google Scholar] [CrossRef]
- Rennie, M.D.; Jackson, L.J. The Influence of Habitat Complexity on Littoral Invertebrate Distributions: Patterns Differ in Shallow Prairie Lakes with and without Fish; NRC Research Press: Ottawa, ON, Canada, 2005; pp. 2088–2099. [Google Scholar]
- Environment Agency of Slovenia. Atlas Okolja. Available online: https://gis.arso.gov.si/atlasokolja/default.aspx (accessed on 15 December 2022).
- Svetina, J.; Prestor, J.; Kopač, I. Hidrogeologija Stanja Izvirov v Stražunskem Gozdu in Določitev Napajalnega Zaledja Potoka/Kanala; Geological Institute of Slovenia: Maribor, Slovenia, 2020. [Google Scholar]
- Oertli, B.; Auderset Joye, D.; Castella, E.; Juge, R.; Lehmann, A.; Lachavanne, J.-B. PLOCH: A standardized method for sampling and assessing the biodiversity in ponds. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 665–679. [Google Scholar] [CrossRef]
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertebres d’eau Douce: Systématique, Biologie, Écologie; CNRS: Paris, France, 2010; 607p. [Google Scholar]
- R Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics sfotware package for education and data analysis. Paleontol. Electr. 2001, 4, 1–9. [Google Scholar]
- Braak, C.T.; Verdonschot, P.F.M. Canonical correspondence analysis and relate multivariate methods in aquatic ecology. Aquatic Sci. 2005, 57, 255–289. [Google Scholar] [CrossRef]
- Smilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data using CANOCO 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Gee, J.H.R.; Smith, B.D.; Lee, K.M.; Griffiths, S.W. The ecological basis of freshwater pond management for biodiversity. Aquat. Conserv. Mar. Freshw. Ecosyst. 1997, 7, 91–104. [Google Scholar] [CrossRef]
- Akasaka, M.; Takamura, N.; Mitsuhashi, T.; Kadono, Y. Effects of land use on aquatic macrophyte diversity and water quality of ponds. Freshw. Biol. 2010, 55, 902–922. [Google Scholar] [CrossRef]
- Lukács, B.A.; Tóthmérész, B.; Borics, G.; Várbíró, G.; Juhász, P.; Kiss, B.; Müller, Z.; G-Tóth, L.; Eros, T. Macrophyte diversity of lakes in the Pannon Ecoregion (Hungary). Limnologica 2015, 53, 74–83. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Portielje, R.; Lammens, E.H.H.R.; Noordhuis, R.; van den Berg, M.S.; Joosse, W.; Meijer, M.L. Resilience of Alternative Stable States during the Recovery of Shallow Lakes from Eutrophication Lake Veluwe as a Case Study. Ecosystems 2007, 10, 4–16. [Google Scholar] [CrossRef]
- Francová, K.; Šumberová, K.; Janauer, G.A.; Adámek, Z. Effects of fish farming on macrophytes in temperate carp ponds. Aquacult. Internat. 2019, 27, 413–436. [Google Scholar] [CrossRef]
- Caraco, N.; Cole, J.; Findlay, S.; Wigand, C. Vascular Plants as Engineers of Oxygen in Aquatic Systems. BioScience 2006, 56, 219–225. [Google Scholar] [CrossRef]
- Papastergiadou, E.; Babalonas, D. The relationships between hydrochemical environmental factors and the aquatic macrophytic vegetation in stagnant and slow flowing waters II. Evaluation of plant associations indicative value. Archiv. Hydrobiol. 1993, 90, 475–491. [Google Scholar]
- Heino, J. Lentic macroinvertebrate assemblage structure along gradients in spatial heterogeneity, habitat size and water chemistry. Hydrobiologia 2000, 418, 229–242. [Google Scholar] [CrossRef]
- Boukal, D.S.; Boukal, M.; Fifaček, M.; Hajek, J.; Klečka, J.; Skalicky, S.; Štasny, J.; Travniček, D. Catalogue of water beetles of the Czech Republic. Klapalekiana 2007, 43, 1–289. [Google Scholar]
- Angélibert, S.; Marty, P.; Céréghino, R.; Giani, N. Seasonal variations in the physical and chemical characteristics of ponds: Implications for biodiversity conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2004, 14, 439–456. [Google Scholar] [CrossRef]
- Watts, J.C. The effect of organic matter on sedimentary phosphorus release in Australian reservoir. Hydrobiologia 2000, 431, 13–25. [Google Scholar] [CrossRef]
- Rezende, R.; Santos, A.M.; Henke-Oliveira, C.; Gonçalves, J.F., Jr. Effect of spatial and environmental factors on a benthic macroinvertebrate community. Zoologia 2014, 31, 426–434. [Google Scholar] [CrossRef]
- Stefanidis, K.; Kostara, K.; Papastergiadou, E. Implications of Human Activities, Land Use Changes and Climate Variability in Mediterranean Lakes of Greece. Water 2016, 8, 483. [Google Scholar] [CrossRef]
- Remsburg, A. Relative Influence of Prior Life Stages and Habitat Variables on Dragonfly (Odonata: Gomphidae) Densities among Lake Sites. Diversity 2011, 3, 200–216. [Google Scholar] [CrossRef]
- Kolar, V.; Boukal, D. Habitat preferences of the endangered diving beetle Graphoderus bilineatus: Implications for conservation management. Insect Conserv. Divers. 2020, 13, 480–494. [Google Scholar] [CrossRef]
- Hill, M.J.; Greaves, H.M.; Sayer, C.D.; Hassall, C.; Milin, M.; Milner, V.S.; Marazzi, L.; Hall, R.; Harper, L.R.; Thornhill, I.; et al. Pond ecology and conservation: Research priorities and knowledge gaps. Ecosphere 2021, 12, e03853. [Google Scholar] [CrossRef]
- Tasker, S.J.L.; Foggo, A.; Bilton, D.T. Quantifying the ecological impacts of alien aquatic macrophytes: A global meta-analysis of effects on fish, macroinvertebrate and macrophyte assemblages. Freshw. Biol. 2022, 67, 1847–1860. [Google Scholar] [CrossRef]
- Zelnik, I.; Germ, M.; Kuhar, U.; Gaberščik, A. Waterbodies in the Floodplain of the Drava River Host Species-Rich Macrophyte Communities despite Elodea Invasions. Diversity 2022, 14, 870. [Google Scholar] [CrossRef]
- Grime, J. Competitive exclusion in herbaceous vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pond Type | Natural | Artificial | |||||
|---|---|---|---|---|---|---|---|
| Average | SE | p | Average | SE | |||
| EC (µS/cm) | 418 | ± | 51 | 0.006 | 207 | ± | 24 |
| Temperature of water (°C) | 19.2 | ± | 1.0 | 0.002 | 24.2 | ± | 0.6 |
| Saturation with O2 [%] | 59 | ± | 13 | n.s. | 77 | ± | 13 |
| [O2] (mg/L) | 5.4 | ± | 1.2 | n.s. | 6.5 | ± | 1.0 |
| [NO3−] (mg/L) | 1.4 | ± | 0.9 | n.s. | 0.3 | ± | 0.2 |
| [PO43−] (mg/L) | 41 | ± | 12 | 0.017 | 97 | ± | 18 |
| BGA-PC (Cells/mL) | 6323 | ± | 3441 | n.s. | 22,341 | ± | 9114 |
| [CDOM] (µg/L) | 38 | ± | 20 | n.s. | 73 | ± | 9 |
| Secchi depth (cm) | 31 | ± | 9 | n.s. | 42 | ± | 6 |
| Canopies above water [%] | 38 | ± | 11 | n.s. | 31 | ± | 9 |
| No. of HT adjacent to pond | 1.8 | ± | 0.3 | n.s. | 1.7 | ± | 0.2 |
| No. of HT within 50 m | 5.0 | ± | 0.3 | n.s. | 4.4 | ± | 0.3 |
| Clay, sand (<0.2 cm) [%] | 56 | ± | 12 | 0.056 | 86 | ± | 6 |
| Gravel (0.2–2 cm) [%] | 13 | ± | 4 | n.s. | 10 | ± | 5 |
| Pebbles (2–6 cm) [%] | 13 | ± | 4 | n.s. | 3 | ± | 3 |
| CPOM [%] | 42 | ± | 10 | n.s. | 23 | ± | 4 |
| FPOM [%] | 75 | ± | 4 | 0.013 | 48 | ± | 9 |
| Filamentous algae [%] | 8.3 | ± | 2.8 | n.s. | 9.2 | ± | 2.2 |
| Width of riparian zone (1–3) | 3.0 | ± | 0.0 | 0.000 | 1.8 | ± | 0.2 |
| Water surface (ha) | 0.70 | ± | 0.13 | n.s. | 3.0 | ± | 1.6 |
| Land use intensity | 2.8 | ± | 0.4 | n.s. | 3.1 | ± | 0.4 |
| TAXA | CRE1 | CRE2 | MIKL | ORE1 | ORE2 | PEKR | PRA1 | PRA2 | PRA3 | RAC1 | RAC2 | ROG | SLIV | SRED | STA1 | STA2 | TEZL | TURN | VURB |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Typha latifolia | 0 | 60 | 0 | 10 | 0 | 1 | 0 | 0 | 60 | 65 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 20 | 30 |
| Phragmites australis | 20 | 0 | 5 | 25 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 35 | 0 | 40 | 0 | 0 | 3 | 0 | 0 |
| Myriophyllum spicatum | 0 | 0 | 5 | 0 | 0 | 1 | 2 | 0 | 5 | 5 | 35 | 5 | 10 | 0 | 15 | 15 | 0 | 10 | 15 |
| Ceratophyllum demersum | 0 | 0 | 0 | 20 | 15 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 30 |
| No. of macrophytes: | 6 | 14 | 14 | 9 | 14 | 12 | 11 | 8 | 6 | 9 | 20 | 9 | 19 | 9 | 9 | 10 | 6 | 6 | 9 |
| Ephemeroptera | 5 | 2 | 424 | 45 | 56 | 9 | 1 | 2 | 2 | 396 | 386 | 4 | 40 | 17 | 83 | 271 | 15 | 39 | 149 |
| Diptera | 23 | 4 | 48 | 60 | 45 | 262 | 36 | 47 | 130 | 99 | 18 | 48 | 2 | 5 | 72 | 150 | 83 | 15 | 62 |
| Isopoda | 0 | 0 | 1 | 5 | 26 | 0 | 0 | 2 | 0 | 0 | 0 | 3 | 0 | 0 | 776 | 317 | 0 | 0 | 51 |
| Odonata | 2 | 1 | 10 | 370 | 196 | 48 | 12 | 7 | 4 | 28 | 46 | 38 | 7 | 1 | 109 | 61 | 13 | 25 | 107 |
| No. of higher MI taxa: | 8 | 6 | 12 | 12 | 13 | 8 | 9 | 10 | 6 | 7 | 8 | 10 | 8 | 7 | 11 | 12 | 9 | 8 | 10 |
| Baetidae | 4 | 2 | 417 | 32 | 53 | 8 | 1 | 1 | 2 | 393 | 357 | 2 | 35 | 14 | 82 | 270 | 15 | 4 | 146 |
| Asellidae | 0 | 0 | 1 | 5 | 26 | 0 | 0 | 2 | 0 | 0 | 0 | 3 | 0 | 0 | 776 | 317 | 0 | 0 | 51 |
| Chironomidae | 14 | 4 | 41 | 21 | 2 | 172 | 33 | 38 | 130 | 16 | 4 | 16 | 1 | 5 | 60 | 107 | 79 | 11 | 45 |
| Coenagrionidae | 2 | 0 | 10 | 319 | 191 | 46 | 10 | 1 | 4 | 24 | 44 | 34 | 7 | 1 | 105 | 59 | 12 | 25 | 98 |
| No. of MI families: | 13 | 10 | 26 | 34 | 29 | 23 | 15 | 19 | 10 | 15 | 20 | 22 | 15 | 10 | 30 | 34 | 17 | 20 | 17 |
| No. of individuals: | 52 | 18 | 1356 | 731 | 638 | 402 | 75 | 92 | 153 | 598 | 564 | 232 | 95 | 163 | 1195 | 1104 | 217 | 233 | 498 |
| Pond Type | Natural | Artificial | |||||
|---|---|---|---|---|---|---|---|
| Average | SE | p | Average | SE | |||
| Number of MF_taxa | 10.8 | ± | 1.0 | n.s. | 10.4 | ± | 1.3 |
| Total cover of all MF [%] | 108 | ± | 2 | 0.006 | 69 | ± | 12 |
| MF_Dominance index [%] | 20.0 | ± | 1.4 | 0.005 | 33.0 | ± | 4.0 |
| S-W DI_MF | 1.87 | ± | 0.06 | 0.040 | 1.57 | ± | 0.1 |
| Margalef DI_MF | 2.10 | ± | 0.22 | n.s. | 2.35 | ± | 0.3 |
| Simpson DI_MF | 0.80 | ± | 0.0 | 0.005 | 0.67 | ± | 0.0 |
| Evenness_MF | 0.61 | ± | 0.03 | n.s. | 0.52 | ± | 0.0 |
| Total abundance of MI | 890 | ± | 150 | 0.005 | 220 | ± | 51 |
| Number of MI_fam | 28.3 | ± | 2.6 | 0.003 | 16.1 | ± | 1.3 |
| S-W DI_MI fam | 1.89 | ± | 0.14 | n.s. | 1.80 | ± | 0.1 |
| Margalef DI_MI_fam | 4.06 | ± | 0.35 | 0.031 | 3.02 | ± | 0.2 |
| Number of_MI_h.taxa | 11.7 | ± | 0.4 | <0.001 | 8.0 | ± | 0.4 |
| S-W DI_MI_h.taxa | 1.60 | ± | 0.07 | n.s. | 1.40 | ± | 0.1 |
| Margalef DI_MI_h.taxa | 1.59 | ± | 0.06 | n.s. | 1.44 | ± | 0.1 |
| TaxonIndex | Number of MF_taxa | Margalef_MF | S-W DI_MF | MF_Dominance Index |
|---|---|---|---|---|
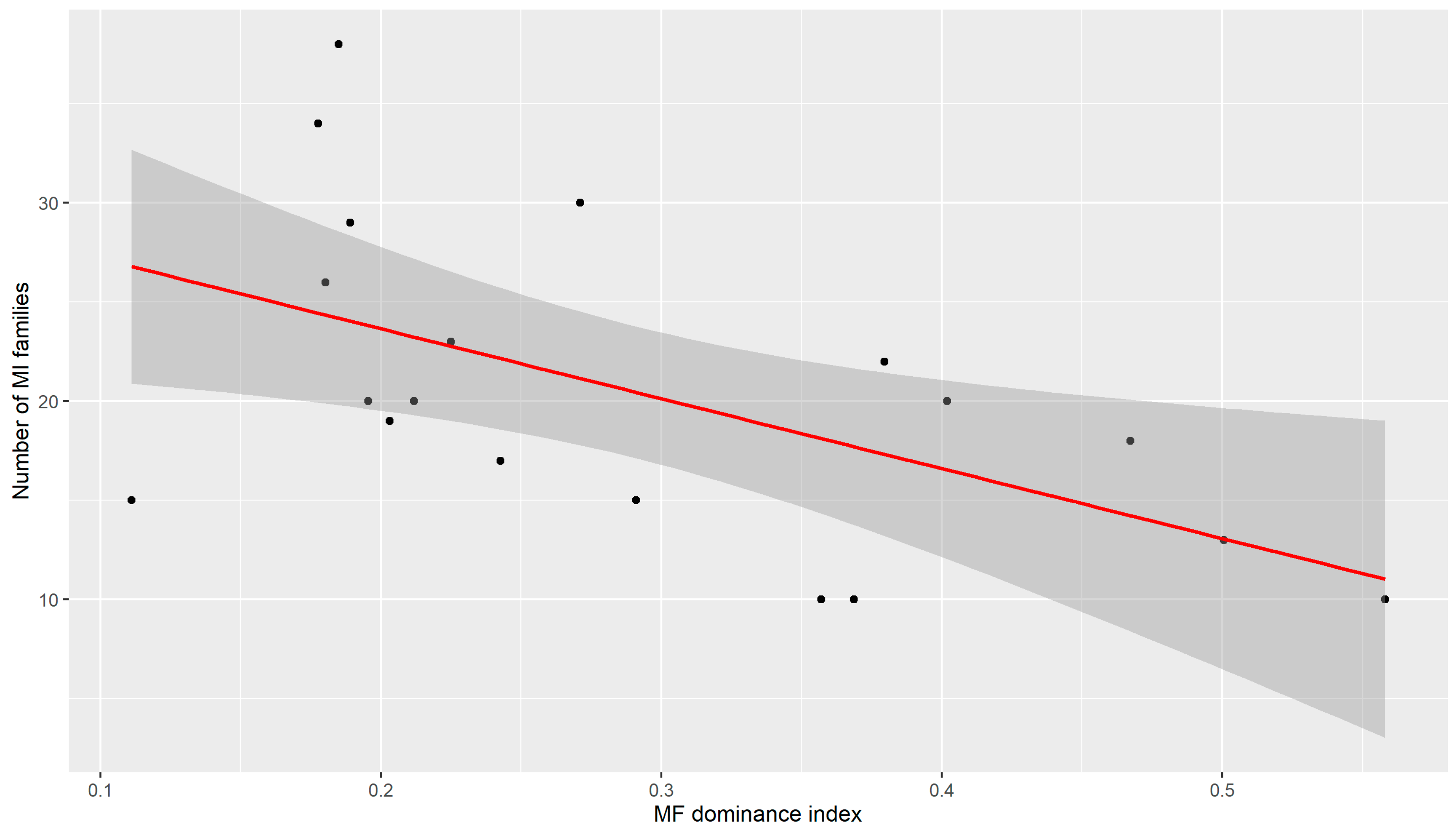
| Number of MI_fam | n.s. | n.s. | 0.564 * | −0.613 ** |
| Total abundance of MI | n.s. | n.s. | n.s. | −0.460 * |
| Number of_MI_h.taxa | n.s. | n.s. | 0.538 * | −0.666 ** |
| Margalef DI_MI fam | n.s. | n.s. | n.s. | −0.505 * |
| S-W DI_MI fam | n.s. | n.s. | n.s. | n.s. |
| Dominance of MI_fam. | n.s. | n.s. | n.s. | n.s. |
| Margalef DI_MI_h.taxa | n.s. | n.s. | n.s. | n.s. |
| S-W DI_MI_h.taxa | n.s. | n.s. | n.s. | n.s. |
| Dominance of MI h.taxa | n.s. | n.s. | n.s. | n.s. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germ, M.; Tertinek, Ž.; Zelnik, I. Diversity of Macrophytes and Macroinvertebrates in Different Types of Standing Waters in the Drava Field. Water 2024, 16, 1130. https://doi.org/10.3390/w16081130
Germ M, Tertinek Ž, Zelnik I. Diversity of Macrophytes and Macroinvertebrates in Different Types of Standing Waters in the Drava Field. Water. 2024; 16(8):1130. https://doi.org/10.3390/w16081130
Chicago/Turabian StyleGerm, Mateja, Žiga Tertinek, and Igor Zelnik. 2024. "Diversity of Macrophytes and Macroinvertebrates in Different Types of Standing Waters in the Drava Field" Water 16, no. 8: 1130. https://doi.org/10.3390/w16081130
APA StyleGerm, M., Tertinek, Ž., & Zelnik, I. (2024). Diversity of Macrophytes and Macroinvertebrates in Different Types of Standing Waters in the Drava Field. Water, 16(8), 1130. https://doi.org/10.3390/w16081130