
Influence of Aspect and Elevational Gradient on Vegetation Pattern, Tree Characteristics and Ecosystem Carbon Density in Northwestern Himalayas


 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
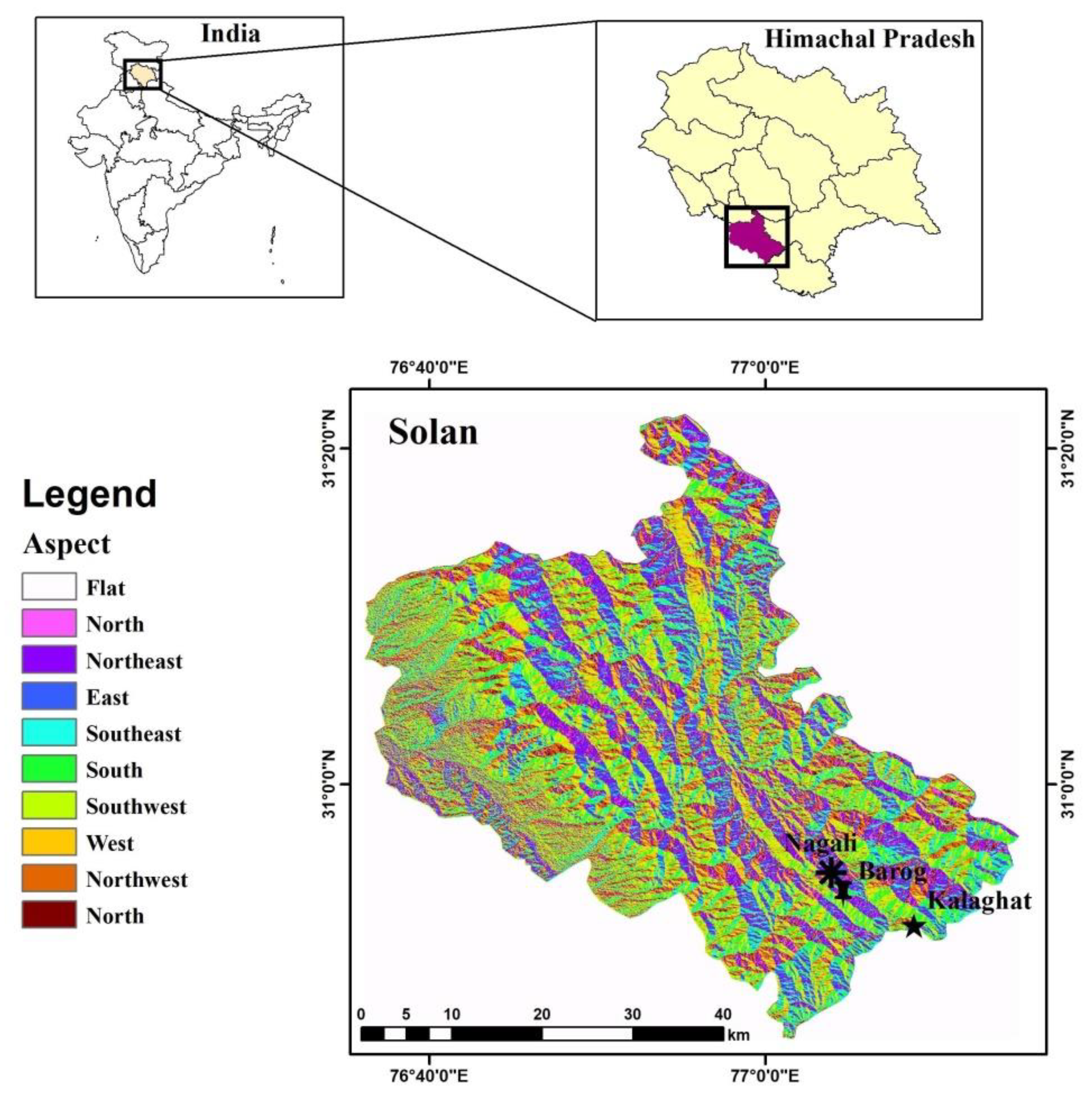
2.1. Study Area
2.2. Vegetation Sampling and Data Analysis
2.3. Biomass and Carbon
2.4. Total Carbon Density
2.5. Vegetation Carbon Density
2.6. Determination of Soil Characteristics
2.7. Ecosystem Carbon Density
2.8. Statistical Analysis
Data: dependent variables; Replication vector: vector containing replications; fact.A: vector containing levels of first factor; fact.B: vector containing levels of second factor; Multiple.comparison.test: 0 for no test, 1 for lsd test, 2 for Duncan test and 3 for HSD test
3. Results
3.1. Floristic Composition of Vegetation Species
3.2. Distribution of Vegetation Communities and Forest Types
3.3. Variation in Tree Characteristics
3.3.1. Diameter at Breast Height (DBH, cm)
3.3.2. Height (m)
3.3.3. Crown Length (m)
3.3.4. Stem Density (N ha−1)
3.3.5. Stem Volume (m3 ha−1)
3.4. Vegetation Biomass and Carbon Density
3.4.1. Above Ground-Below Ground Biomass (Mg ha−1)
3.4.2. Shrub and Herb Biomass (Mg ha−1)
3.4.3. Vegetation Biomass Density (Mg ha−1)
3.5. Soil Physico-Chemical Characteristics
4. Discussion
4.1. Floristic Diversity and Distribution of Vegetation Community
4.2. Variations in Tree Characteristics
4.3. Vegetation Biomass and Carbon Density
4.4. Soil Physico-Chemical Properties and Correlation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Price, M.F.; Georg, G.; Lalsa, A.D.; Thomas, K.; Daniel, M.; Rosalaura, R. Mountain Forests in a Changing World: Realizing Values, Addressing Challenges; Food and Agriculture Organisations of the United Nations (FAO) with the Support of the Swiss Agency for Development and Cooperation (SDC): Rome, Italy, 2011. [Google Scholar]
- FAO. The state of Food Insecurity in the World; Food and agriculture Organisation of the United Nations (FAO): Rome, Italy, 2002. [Google Scholar]
- Singh, J.S. Sustainable development of the Indian Himalayan region: Linking ecological and economic concern. Curr. Sci. 2006, 90, 784–788. [Google Scholar]
- Kumar, A.; Sharma, M.P.; Taxak, A.K. Effect of vegetation communities and altitudes on the soil organic carbon stock in Kotli Bhel-1A Catchment, India. CLEAN—Soil Air Water 2017, 45, 1600650. [Google Scholar] [CrossRef]
- Brang, P.; Schönenberger, W.; Ott, E.; Gardner, B. Forests as protection from natural hazards. In The Forests Handbook; Evans, J., Ed.; Blackwell Science: Oxford, UK, 2001; Volume 2, pp. 53–81. [Google Scholar]
- Huang, W.; Pohjonen, V.; Johansson, S.; Nashanda, M.; Katigula, M.; Luukkanen, O. Species diversity, forest structure and species composition in Tanzanian tropical forests. For. Ecol. Manag. 2003, 173, 11–24. [Google Scholar] [CrossRef]
- Bhat, J.A.; Kumar, M.; Negi, A.; Todaria, N.; Malik, Z.A.; Pala, N.A.; Kumar, A.; Shukla, G. Altitudinal gradient of Species diversity and community of woody vegetation along altitudinal gradient of the Western Himalayas. Glob. Ecol. Conserv. 2020, 24, e01302. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Zavaleta, E.S.; Eviner, V.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef]
- Wood, E.; Sivapalan, M.; Beven, K.; Band, L. Effects of spatial variability and scale with implications to hydrologic modeling. J. Hydrol. 1988, 102, 29–47. [Google Scholar] [CrossRef]
- Dawes, W.R.; Short, D. The significance of topology for modeling the surface hydrology of fluvial landscapes. Water Resour. Res. 1994, 30, 1045–1055. [Google Scholar] [CrossRef]
- O’Loughlin, E. Saturation regions in catchments and their relations to soil and topographic properties. J. Hydrol. 1981, 53, 229–246. [Google Scholar] [CrossRef]
- Enright, N.; Miller, B.; Akhter, R. Desert vegetation and vegetation-environment relationships in Kirthar National Park, Sindh, Pakistan. J. Arid. Environ. 2005, 61, 397–418. [Google Scholar] [CrossRef]
- Brown, J.H. Mammals on mountainsides: Elevational patterns of diversity. Glob. Ecol. Biogeogr. 2001, 10, 101–109. [Google Scholar] [CrossRef]
- Kumar, A.; Pinto, M.C.; Candeias, C.; Dinis, P.A. Baseline maps of potentially toxic elements in the soils of Garhwal Himalayas, India: Assessment of their eco-environmental and human health risks. Land Degrad. Dev. 2021, 32, 3856–3869. [Google Scholar] [CrossRef]
- Holland, P.G.; Steyn, D.G. Vegetational Responses to Latitudinal Variations in Slope Angle and Aspect. J. Biogeogr. 1975, 2, 179–183. [Google Scholar] [CrossRef]
- Sharma, C.M.; Suyal, S.; Gairola, S.; Ghildiyal, S.K. Species richness and diversity along an altitudinal gradient in moist temperate forest of Garhwal Himalaya. J. Am. Sci. 2009, 5, 119–128. [Google Scholar]
- Singh, S.; Malik, Z.A.; Sharma, C.M. Tree species richness, diversity, and regeneration status in different oak (Quercus spp.) dominated forests of Garhwal Himalaya, India. J. Asia-Pac. Biodivers. 2016, 9, 293–300. [Google Scholar] [CrossRef]
- Chen, W.J.; Black, T.A.; Yang, P.C.; Barr, A.; Neumann, H.H.; Nesic, Z.; Blanken, P.D.; Novak, M.D.; Eley, J.; Ketler, R.J.; et al. Effects of climatic variability on the annual carbon sequestration by a boreal aspen forest. Glob. Chang. Biol. 1999, 5, 41–53. [Google Scholar] [CrossRef]
- Wang, H.; Saigusa, N.; Yamamoto, S.; Kondo, H.; Hirano, T.; Toriyama, A.; Fujinuma, Y. Net ecosystem CO2 exchange over a larch forest in Hokkaido, Japan. Atmos. Environ. 2004, 38, 7021–7032. [Google Scholar] [CrossRef]
- Li, S.G.; Asanuma, J.; Kotani, A.; Eugster, W.; Davaa, G.; Oyunbaatar, D.; Sugita, M. Year-round measurements of net eco-system CO2 flux over a montane larch forest in Mongolia. J. Geophys. Res. Atmos. 2005, 110. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, M.P.; Yang, T. Estimation of carbon stock for greenhouse gas emissions from hydropower reservoirs. Stoch. Environ. Res. Risk Assess. 2018, 32, 3183–3193. [Google Scholar] [CrossRef]
- Brown, J.H.; Stevens, G.C.; Kaufman, D.M. the geographic range: Size, shape, boundaries, and internal structure. Annu. Rev. Ecol. Syst. 1996, 27, 597–623. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, M. Estimation of Biomass and Soil Carbon Stock in the Hydroelectric Catchment of India and its Implementation to Climate Change. J. Sustain. For. 2020, 36, 1–16. [Google Scholar] [CrossRef]
- Sandrine, L.; Claude, N.Y.S.; Christian, W.; Françoise, F.; Sandrine, H.; Paula, R.; Stéphane, F. Estimation of carbon stocks in a beech forest (Fougères Forest—W. France): Extrapolation from the plots to the whole forest. Ann. For. Sci. 2006, 63, 139–148. [Google Scholar] [CrossRef][Green Version]
- Zhang, N.; Yu, Z.; Yu, G.; Wu, J. Scaling up ecosystem productivity from patch to landscape: A case study of Changbai Mountain Nature Reserve, China. Landsc. Ecol. 2007, 22, 303–315. [Google Scholar] [CrossRef]
- FAO. Climate-Smart Agriculture: Policies, Practices and Financing for Food Security, Adaptation and Mitigation; Food and Agriculture Organisation of the United Nations (FAO): Rome, Italy, 2010. [Google Scholar]
- McMahon, S.M.; Parker, G.; Miller, D.R. Evidence for a recent increase in forest growth. Proc. Natl. Acad. Sci. USA 2010, 107, 3611–3615. [Google Scholar] [CrossRef]
- Mwakisunga, B.; Majule, A.E. The influence of altitude and management on carbon stock quantities in rung we forest, southern highland of Tanzania. Open J. Ecol. 2012, 02, 214–221. [Google Scholar] [CrossRef]
- MacDicken, K.G. A Guide to Monitoring Carbon Emissions from Deforestation and Degradation in Developing Countries: An Examination of Issues Facing the Incorporation of REDD into Market-Based Climate Policies; Resource for Future: Washington, DC, USA, 1997. [Google Scholar]
- IPCC. Climate Change: A Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 1995. [Google Scholar]
- Chauhan, M.; Kumar, M.; Kumar, A. Impact of carbon stocks of Anogeissus latifolia on climate change and socio-economic development: A case study of Garhwal Himalaya, India. Water Air Soil Pollut. 2020, 231, 436. [Google Scholar] [CrossRef]
- McNulty, S.; Treasure, E.; Jennings, L.; Meriwether, D.; Harris, D.; Arndt, P. Translating national level forest service goals to local level land management: Carbon sequestration. Clim. Chang. 2018, 146, 133–144. [Google Scholar] [CrossRef]
- St-Laurent, G.P.; Hagerman, S.; Kozak, R.; Hoberg, G. Public perceptions about climate change mitigation in British Columbia’s forest sector. PLoS ONE 2018, 13, e0195999. [Google Scholar] [CrossRef]
- A Ontl, T.; Janowiak, M.K.; Swanston, C.W.; Daley, J.; Handler, S.; Cornett, M.; Hagenbuch, S.; Handrick, C.; McCarthy, L.; Patch, N. Forest Management for Carbon Sequestration and Climate Adaptation. J. For. 2020, 118, 86–101. [Google Scholar] [CrossRef]
- Sharma, C.M.; Mishra, A.K.; Tiwari, O.P.; Krishan, R.; Rana, Y.S. Effect of altitudinal gradients on forest structure and composition on ridge tops in Garhwal Himalaya. Energy Ecol. Environ. 2017, 2, 404–417. [Google Scholar] [CrossRef]
- Sharma, C.M.; Gairola, S.; Baduni, N.P.; Ghildiyal, S.K.; Suyal, S. Variation in carbon stocks on different slope aspects in seven major forest types of temperate region of Garhwal Himalaya, India. J. Biosci. 2011, 36, 701–708. [Google Scholar] [CrossRef]
- Bhardwaj, D.R.; Banday, M.; Pala, N.A.; Rajput, B.S. Variation of biomass and carbon pool with NDVI and altitude in sub-tropical forests of northwestern Himalaya. Environ. Monit. Assess. 2016, 188, 635. [Google Scholar] [CrossRef]
- Banday, M.; Bhardwaj, D.R.; Pala, N.A.; Rajput, B.S. Quantitative analysis of woody vegetation in subtropical forests of Himachal Pradesh, India. Indian For. 2017, 143, 617–629. [Google Scholar]
- Shah, S.; Sharma, D.P.; Pala, N.A.; Tripathi, P.; Kumar, M. Temporal variations in carbon stock of Pinus roxburghii Sargent forests of Himachal Pradesh, India. J. Mt. Sci. 2014, 11, 959–966. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, M.; Pandey, R.; Yu, Z.; Cabral-Pinto, M. Forest soil nutrient stocks along with an altitudinal range of Uttarakhand Himalayas: An aid to Nature Based Climate Solutions. CATENA 2021, 207, 105667. [Google Scholar] [CrossRef]
- Cottam, G.; Curtis, J.T. The use of distance measures in phytosociological sampling. Ecology 1956, 35, 451–460. [Google Scholar] [CrossRef]
- Chaturvedi, A.N.; Khanna, L.S. Forest Mensuration; International Book Distributors: Dehradun, India, 1982. [Google Scholar]
- Assmann, E. The Principles of Forest Yield Study, 2nd ed.; Oxford Pregmon Press Ltd.: Oxford, UK, 1970. [Google Scholar]
- FSI. Volume Equations for Forests of India, Nepal, and Bhutan; Forest Survey of India, Ministry of Environment and Forests, Government of India: Dehradun, India, 1996. [Google Scholar]
- Brown, S.L.; Schroeder, P.; Kern, J.S. Spatial distribution of biomass in forests of the eastern USA. For. Ecol. Manag. 1999, 123, 81–90. [Google Scholar] [CrossRef]
- Rajput, S.S.; Shukla, N.K.; Gupta, V.K. Specific gravity of Indian timbers. J. Timber Dev. Assoc. India 1985, 31, 12–41. [Google Scholar]
- IPCC. Guidelines for National Greenhouse Gas Inventories; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2006; Volume 4. [Google Scholar]
- Smith, D.M. Maximum Moisture Content Method for Determining Specific Gravity of Small Wood Samples; US Department of Agriculture, Forest Service, Forest Products Laboratory: Madison, WI, USA, 1954; pp. 110–120. [Google Scholar]
- IPCC. Impacts, adaptations and mitigation of climate: Scientific-technical analyses. In Contribution of the Second Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Cairns, M.A.; Brown, S.; Helmer, E.H.; Baumgardner, G.A. Root biomass allocation in the world’s upland forests. Oecologia 1997, 111, 1–11. [Google Scholar] [CrossRef]
- Richards, L.A. Diagnosis and Improvement of Saline and Alkali Soils; USDA Handbook 60, United States Deptartment of Agriculture: Washington, DC, USA, 1954.
- Walkley, A.; Black, J.A. Estimation of soil organic carbon by chromic acid filtration method. Soil Sci. 1934, 37, 38–39. [Google Scholar]
- Rich, C.I. Elemental analysis by flame photometry. In Methods of Soil Analysis. Part 2: Chemical and Microbiological Properties; Black, C.A., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 849–864. [Google Scholar] [CrossRef]
- Subbiah, B.V.; Asija, G.L. A rapid procedure for the determination of available nitrogen in soil. Curr. Sci. 1956, 25, 259–260. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. In Method of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Summer, M.E., Eds.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Champion, H.G.; Seth, S.K. A Revised Survey of Forest Types of India; Government of India Press: New Delhi, India, 1968. [Google Scholar]
- Chawla, A.; Rajkumar, S.; Singh, K.N.; Lal, B.; Singh, R.D.; Thukral, A.K. Plant species diversity along an altitudinal gradient of Bhabha Valley in western Himalaya. J. Mt. Sci. 2008, 5, 157–177. [Google Scholar] [CrossRef]
- Körner, C. Why are there global gradients in species richness? Mountains might hold the answer. Trends Ecol. Evol. 2000, 15, 513–514. [Google Scholar] [CrossRef]
- Krashevs’ka, V. Diversity and Community Structure of Testate Amoebae (Protista) in Tropical Montane Rain Forests of Southern Ecuador: Altitudinal Gradient, Aboveground Habitats and Nutrient Limitation. Doctoral Thesis, Vom Fachbereich Biologie der Technischen Universität, Darmstadt, Germany, 2008. [Google Scholar]
- Noguesbravo, D.; Araújo, M.B.; Romdal, T.S.; Rahbek, C. Scale effects and human impact on the elevational species richness gradients. Nature 2008, 453, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Sang, W. Plant diversity patterns and their relationships with soil and climatic factors along an altitudinal gradient in the middle Tianshan Mountain area, Xinjiang, China. Ecol. Res. 2009, 24, 303–314. [Google Scholar] [CrossRef]
- McCain, C.M. Elevational gradients in diversity of small mammals. Ecology 2005, 86, 366–372. [Google Scholar] [CrossRef]
- Bässler, C.; Müller, J.; Hothorn, T.; Kneib, T.; Badeck, F.-W.; Dziock, F. Estimation of the extinction risk for high-montane species as a consequence of global warming and assessment of their suitability as cross-taxon indicators. Ecol. Indic. 2010, 10, 341–352. [Google Scholar] [CrossRef]
- Monarrez-Gonzalez, J.C.; Gonzalez-Elizondo, M.S.; Marquez-Linares, M.A.; Gutierrez-Yurrita, P.J.; Perez-Verdin, G. Effect of forest management on tree diversity in temperate ecosystem forests in northern Mexico. PLoS ONE 2020, 15, e0233292. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.T.; Liu, H.Y.; Dai, J.U. Research on Mountain Ecology and Alpine Treeline; Science Press: Beijing, China, 2005. [Google Scholar]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Shen, Z.; Cui, H. Ecological characteristics of mountains and research issues of mountain ecology. Biodivers. Sci. 2004, 12, 10–19. [Google Scholar]
- Måren, I.; Karki, S.; Prajapati, C.; Yadav, R.K.; Shrestha, B.B. Facing north or south: Does slope aspect impact forest stand characteristics and soil properties in a semiarid trans-Himalayan valley? J. Arid. Environ. 2015, 121, 112–123. [Google Scholar] [CrossRef]
- Mong, C.E.; Vetaas, O.R. Establishment of Pinuswallichiana on a Himalayan glacier foreland: Stochastic distribution or safe sites? Arct. Antarct. Alp. Res. 2006, 38, 584–592. [Google Scholar] [CrossRef]
- Banday, M.; Bhardwaj, D.R.; Pala, N.A. Variation of stem density and vegetation carbon pool in subtropical forests of Northwestern Himalaya. J. Sustain. For. 2018, 37, 389–402. [Google Scholar] [CrossRef]
- Missanjo, E.; Kamanga-Thole, G.; Manda, V. Estimation of Genetic and Phenotypic Parameters for Growth Traits in a Clonal Seed Orchard of Pinus kesiya in Malawi. ISRN For. 2013, 2013, 346982. [Google Scholar] [CrossRef]
- Pala, N.A.; Negi, A.K.; Gokhale, Y.; Aziem, S.; Vikrant, K.K.; Todaria, N.P. Carbon stock estimation for tree species of Sem Mukhem sacred forest in Garhwal Himalaya, India. J. For. Res. 2013, 24, 457–460. [Google Scholar] [CrossRef]
- Iqbal, K.; Pala, N.A.; Bhat, J.A.; Negi, A.K. Regeneration status of trees around Khoh River in Garhwal Himalaya. Indian J. For. 2012, 35, 471–476. [Google Scholar]
- Iqbal, K.; Bhat, J.A.; Pala, N.A.; Hussain, A.; Negi, A.K. Carbon and biomass density of trees in Duggada area of Garhwal Himalaya, India. Indian For. 2014, 140, 18–22. [Google Scholar]
- Srivastava, R.K.; Khanduri, V.P.; Sharma, C.M.P. Structure, diversity and regeneration potential of Q. leucotrichophora domi-nant conifer mixed forest along an altitudinal gradient of Garhwal Himalaya. Indian For. 2005, 131, 1537–1553. [Google Scholar]
- Kitayama, K.; Aiba, S.-I. Ecosystem structure and productivity of tropical rain forests along altitudinal gradients with contrasting soil phosphorus pools on Mount Kinabalu, Borneo. J. Ecol. 2002, 90, 37–51. [Google Scholar] [CrossRef]
- Moser, G.; Hertel, D.; Leuschner, C. Altitudinal Change in LAI and Stand Leaf Biomass in Tropical Montane Forests: A Transect Study in Ecuador and a Pan-Tropical Meta-Analysis. Ecosystems 2007, 10, 924–935. [Google Scholar] [CrossRef]
- Leuschner, C.; Moser, G.; Bertsch, C.; Röderstein, M.; Hertel, D. Large altitudinal increase in tree root/shoot ratio in tropical mountain forests of Ecuador. Basic Appl. Ecol. 2007, 8, 219–230. [Google Scholar] [CrossRef]
- Raich, J.W.; Russell, A.E.; Kitayama, K.; Parton, W.J.; Vitousek, P.M. Temperature influences carbon accumulation in moist tropical forests. Ecology 2006, 87, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Alves, L.F.; Vieira, S.A.; Scaranello, M.A.; de Camargo, P.B.; Santos, F.; Joly, C.A.; Martinelli, L. Forest structure and live aboveground biomass variation along an elevational gradient of tropical Atlantic moist forest (Brazil). For. Ecol. Manag. 2010, 260, 679–691. [Google Scholar] [CrossRef]
- Moser, G.; Leuschner, C.; Hertel, D.; Graefe, S.; Soethe, N.; Iost, S. Elevation effects on the carbon budget of tropical mountain forests (S Ecuador): The role of the belowground compartment. Glob. Chang. Biol. 2010, 17, 2211–2226. [Google Scholar] [CrossRef]


{kind=link}
| Altitudinal Ranges (AR) (m a.s.l.) | Aspect (A) | ||||
|---|---|---|---|---|---|
| E | W | N | S | Mean | |
| DBH (cm) | |||||
| AR1 (1000–1300) | 25.00 | 19.00 | 36.30 | 23.40 | 26.00 a |
| AR2 (1000–1300) | 22.00 | 18.60 | 22.50 | 22.80 | 22.00 b |
| AR3 (1000–1300) | 26.20 | 20.00 | 26.00 | 19.40 | 20.00 b |
| Mean | 28.00 a | 21.80 b | 22.00 b | 21.16 b | |
| LSD | AR = 3.70 | A = 4.30 | AR × A = 3.90 | ||
| Height (m) | |||||
| AR1 (1000–1300) | 14.50 | 10.22 | 22.40 | 16.48 | 16.26 a |
| AR2 (1000–1300) | 13.00 | 9.30 | 11.60 | 10.00 | 12.18 b |
| AR3 (1000–1300) | 12.00 | 12.30 | 13.00 | 10.00 | 11.38 b |
| Mean | 13.90 ab | 12.40 bc | 16.30 a | 10.50 c | |
| LSD | AR = 2.70 | A = 3.20 | AR × A = NS | ||
| Crown length (m) | |||||
| AR1 (1000–1300) | 4.70 | 5.50 | 7.90 | 4.40 | 6.00 a |
| AR2 (1000–1300) | 6.00 | 4.60 | 5.30 | 3.50 | 5.00 a |
| AR3 (1000–1300) | 5.00 | 3.00 | 3.15 | 2.20 | 3.00 b |
| Mean | 5.16 | 3.70 | 5.20 | 4.40 | |
| LSD | AR = 1.90 | A = NS | AR × A = 4.50 | ||
| Stem density (N ha−1) | |||||
| AR1 (1000–1300) | 258.00 | 198.00 | 217.00 | 170.00 | 211.00 a |
| AR2 (1000–1300) | 228.00 | 175.00 | 283.00 | 165.00 | 188.00 a |
| AR3 (1000–1300) | 116.00 | 188.00 | 133.00 | 143.00 | 146.00 b |
| Mean | 200.00 ab | 154.60 c | 211.00 a | 166.00 bc | |
| LSD | AR = 37.00 | A = 43.00 | AR × A = NS | ||
| Stem volume (m3 ha−1) | |||||
| AR1 (1000–1300) | 118.83 | 40.90 | 236.50 | 74.40 | 117.68 a |
| AR2 (1000–1300) | 132.16 | 39.50 | 105.40 | 32.17 | 77.33 b |
| AR3 (1000–1300) | 101.58 | 35.30 | 40.90 | 53.00 | 57.89 b |
| Mean | 127.60 a | 53.00 b | 117.50 a | 38.60 b | |
| LSD | AR = 38.80 | A = 44.80 | AR × A = 53.23 | ||
| Stem biomass (Mg ha−1) | |||||
| AR1 (1000–1300) | 120.50 | 36.58 | 72.75 | 21.58 | 82.85 |
| AR2 (1000–1300) | 58.92 | 16.25 | 132.16 | 18.66 | 56.50 |
| AR3 (1000–1300) | 20.08 | 35.50 | 63.91 | 21.91 | 35.35 |
| Mean | 66.50 a | 29.44 b | 89.61 a | 20.72 b | |
| LSD | AR = NS | A = 30.73 | AR × A = 53.23 | ||
| Altitudinal Ranges (AR) (m a.s.l.) | Aspect (A) | ||||
|---|---|---|---|---|---|
| E | W | N | S | Mean | |
| Above ground biomass (t ha−1) | |||||
| AR1 (1000–1300) | 222.05 | 69.88 | 130.61 | 42.49 | 116.25 a |
| AR2 (1000–1300) | 112.44 | 31.14 | 252.31 | 43.36 | 109.81 a |
| AR3 (1000–1300) | 38.38 | 67.87 | 115.75 | 33.07 | 63.76 b |
| Mean | 124.28 a | 56.29 b | 166.22 a | 39.64 b | |
| lsd | AR = 40.00 | A = 55.25 | AR × A = 95.693 | ||
| Below ground biomass (t ha−1) | |||||
| AR1 (1000–1300) | 47.88 | 14.67 | 41.65 | 9.39 | 28.40 |
| AR2 (1000–1300) | 29.77 | 6.54 | 90.31 | 7.6 | 33.56 |
| AR3 (1000–1300) | 8.06 | 22.96 | 32.24 | 6.94 | 17.5 |
| Mean | 28.57 b | 14.72 bc | 54.73 a | 7.98 c | |
| lsd | AR = NS | A = 18.74 | AR × A = 32.46 | ||
| Total tree biomass (t ha−1) | |||||
| AR1 (1000–1300) | 315.65 | 84.56 | 172.26 | 51.88 | 156.08 a |
| AR2 (1000–1300) | 142.21 | 37.68 | 342.62 | 53.09 | 143.90 a |
| AR3 (1000–1300) | 46.44 | 90.83 | 147.99 | 40.01 | 81.32 b |
| Mean | 168.09 a | 71.02 b | 220.95 a | 48.32 b | |
| lsd | AR = 58.69 | A = 67.77 | AR × A = 117.388 | ||
| Shrub biomass (Mg ha−1) | |||||
| AR1 (1000–1300) | 1.84 | 1.75 | 1.85 | 2.37 | 1.95 a |
| AR2 (1000–1300) | 1.60 | 1.76 | 1.56 | 1.79 | 1.67 ab |
| AR3 (1000–1300) | 1.40 | 1.33 | 1.38 | 1.72 | 1.45 b |
| Mean | 1.61 | 1.62 | 1.59 | 1.96 | |
| lsd | AR = 0.303 | A = NS | AR × A = NS | ||
| Herb biomass (Mg ha−1) | |||||
| AR1 (1000–1300) | 2.93 | 3.27 | 3.10 | 3.92 | 3.30 |
| AR2 (1000–1300) | 3.24 | 2.77 | 2.80 | 3.94 | 3.19 |
| AR3 (1000–1300) | 2.55 | 2.46 | 2.22 | 3.49 | 2.68 |
| Mean | 2.90 b | 2.83 b | 2.71 b | 3.78 a | |
| lsd | AR = NS | A = 0.63 | AR × A = NS | ||
| Vegetation biomass density (Mg ha−1) | |||||
| AR1 (1000–1300) | 320.41 | 89.57 | 177.22 | 57.97 | 161.29 a |
| AR2 (1000–1300) | 146.82 | 42.11 | 347.33 | 58.66 | 148.73 a |
| AR3 (1000–1300) | 50.39 | 94.63 | 151.86 | 45.16 | 85.50 b |
| Mean | 172.62 a | 75.46 b | 225.27 a | 53.93 b | |
| lsd | AR = 58.78 | A = 67.85 | AR × A = 117.52 | ||
| Altitudinal Ranges (AR) (m a.s.l.) | Aspect (A) | ||||
|---|---|---|---|---|---|
| E | W | N | S | Mean | |
| Vegetation carbon density (Mg ha−1) | |||||
| AR1 (1000–1300) | 160.21 | 44.79 | 88.61 | 28.98 | 80.64 a |
| AR2 (1000–1300) | 73.41 | 21.05 | 173.67 | 29.33 | 74.36 ab |
| AR3 (1000–1300) | 25.20 | 47.31 | 75.93 | 22.58 | 42.75 b |
| Mean | 86.27 a | 37.71 b | 112.73 a | 26.96 b | |
| LSD | AR = 33.92 | A = 29.38 | AR × A = 58.76 | ||
| Soil carbon density (Mg ha−1) | |||||
| AR1 (1000–1300) | 43.23 | 40.83 | 33.83 | 39.77 | 39.42 a |
| AR2 (1000–1300) | 40.93 | 36.90 | 36.67 | 37.50 | 38.00 a |
| AR3 (1000–1300) | 25.53 | 28.20 | 31.43 | 29.40 | 28.64 b |
| Mean | 36.56 | 35.31 | 33.97 | 35.55 | |
| LSD | AR = 36.56 | A = NS | AR × A = 5.94 | ||
| Soil humus carbon density (Mg ha−1) | |||||
| AR1 (1000–1300) | 1.57 | 0.73 | 1.23 | 1.60 | 1.28 b |
| AR2 (1000–1300) | 3.10 | 1.33 | 1.73 | 2.57 | 2.18 a |
| AR3 (1000–1300) | 2.17 | 1.27 | 1.17 | 1.80 | 1.60 b |
| Mean | 2.28 a | 1.11 c | 1.38 bc | 1.99 ab | |
| LSD | AR = 0.55 | A = 0.64 | AR × A = NS | ||
| Leaf litter carbon (Mg ha−1) | |||||
| AR1 (1000–1300) | 2.22 | 2.02 | 1.25 | 1.92 | 1.85 b |
| AR2 (1000–1300) | 2.17 | 1.94 | 2.06 | 2.11 | 2.07 a |
| AR3 (1000–1300) | 1.98 | 1.78 | 1.71 | 1.95 | 1.86 b |
| Mean | 2.12 a | 1.91 c | 1.67 d | 1.99 a | |
| LSD | AR = 0.02 | A = 0.051 | AR × A = 0.11 | ||
| Ecosystem carbon density (Mg ha−1) | |||||
| AR1 (1000–1300) | 207.23 | 88.37 | 125.73 | 71.58 | 123.20 a |
| AR2 (1000–1300) | 119.61 | 61.22 | 214.13 | 70.66 | 116.40 a |
| AR3 (1000–1300) | 54.88 | 78.56 | 110.52 | 55.20 | 74.78 b |
| Mean | 127.20 a | 76.11 b | 149.90 a | 65.80 b | |
| LSD | AR = 29.60 | A = 34.18 | AR × A = 59.21 | ||
| Treatment | Parameters | ||||||
|---|---|---|---|---|---|---|---|
| pH | Bulk Density (g cm−3) | Organic Carbon (g kg−1) | Available N (kg ha−1) | Available P (kg ha−1) | Available K (kg ha−1) | EC (dS m−2) | |
| Altitudinal range (AR) (m a.s.l.) | |||||||
| AR1 (1000–1300 m a.s.l.) | 6.09 b | 0.86 c | 22.8 a | 413.22 a | 19.78 a | 242.37 a | 0.31 a |
| AR2 (1300–1600 m a.s.l.) | 6.31 a | 0.90 bc | 21.2 b | 355.68 b | 16.70 b | 223.38 b | 0.30 b |
| AR3 (1600–1900 m a.s.l.) | 5.98 c | 0.94 a | 15.3 c | 327.11 c | 13.42 c | 209.20 c | 0.29 c |
| LSD (E) | 0.09 | 0.051 | 0. 89 | 21.23 | 1.33 | 6.87 | 0.009 |
| Aspect (A) | |||||||
| E (Eastern) | 5.98 c | 0.87 bc | 19.9 b | 347.28 b | 15.87 b | 229.63 a | 0.29 b |
| W (Western) | 5.99 c | 0.92 ab | 19.3 bc | 330.86 c | 18.43 a | 213.54 b | 0.29 b |
| N (Northern) | 6.18 b | 0.85 c | 21.2 a | 480.80 a | 18.62 a | 195.79 c | 0.31 a |
| S (Southern) | 6.34 a | 0.96 a | 18.6 c | 304.39 d | 13.60 c | 200.98 c | 0.30 ab |
| LSD (A) | 0.11 | 0.059 | 1.03 | 24.51 | 1.54 | 7.93 | 0.01 |
| Soil layers (cm) | |||||||
| L1 (0–20 cm) | 5.95 b | 0.79 a | 21.0 a | 380.19 a | 18.25 a | 235.89 a | 0.31 |
| L2 (20–40 cm) | 6.30 a | 1.01 b | 18.5 b | 350.48 b | 15.01 b | 214.08 b | 0.30 |
| LSD (L) | 0.07 | 0.04 | 0. 73 | 17.33 | 1.09 | 5.61 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhardwaj, D.R.; Tahiry, H.; Sharma, P.; Pala, N.A.; Kumar, D.; Kumar, A.; Bharti. Influence of Aspect and Elevational Gradient on Vegetation Pattern, Tree Characteristics and Ecosystem Carbon Density in Northwestern Himalayas. Land 2021, 10, 1109. https://doi.org/10.3390/land10111109
Bhardwaj DR, Tahiry H, Sharma P, Pala NA, Kumar D, Kumar A, Bharti. Influence of Aspect and Elevational Gradient on Vegetation Pattern, Tree Characteristics and Ecosystem Carbon Density in Northwestern Himalayas. Land. 2021; 10(11):1109. https://doi.org/10.3390/land10111109
Chicago/Turabian StyleBhardwaj, D. R., Habibullah Tahiry, Prashant Sharma, Nazir A. Pala, Dhirender Kumar, Amit Kumar, and Bharti. 2021. "Influence of Aspect and Elevational Gradient on Vegetation Pattern, Tree Characteristics and Ecosystem Carbon Density in Northwestern Himalayas" Land 10, no. 11: 1109. https://doi.org/10.3390/land10111109
APA StyleBhardwaj, D. R., Tahiry, H., Sharma, P., Pala, N. A., Kumar, D., Kumar, A., & Bharti. (2021). Influence of Aspect and Elevational Gradient on Vegetation Pattern, Tree Characteristics and Ecosystem Carbon Density in Northwestern Himalayas. Land, 10(11), 1109. https://doi.org/10.3390/land10111109