
The Temporal and Spatial Dynamics of Succession in a Glacial Foreland in Southern Iceland: The Effects of Landscape Heterogeneity

Abstract
:1. Introduction
2. Methods
2.1. Location and Setting
2.2. Previous Work
2.3. Field Techniques
2.4. Data Treatment
3. Results
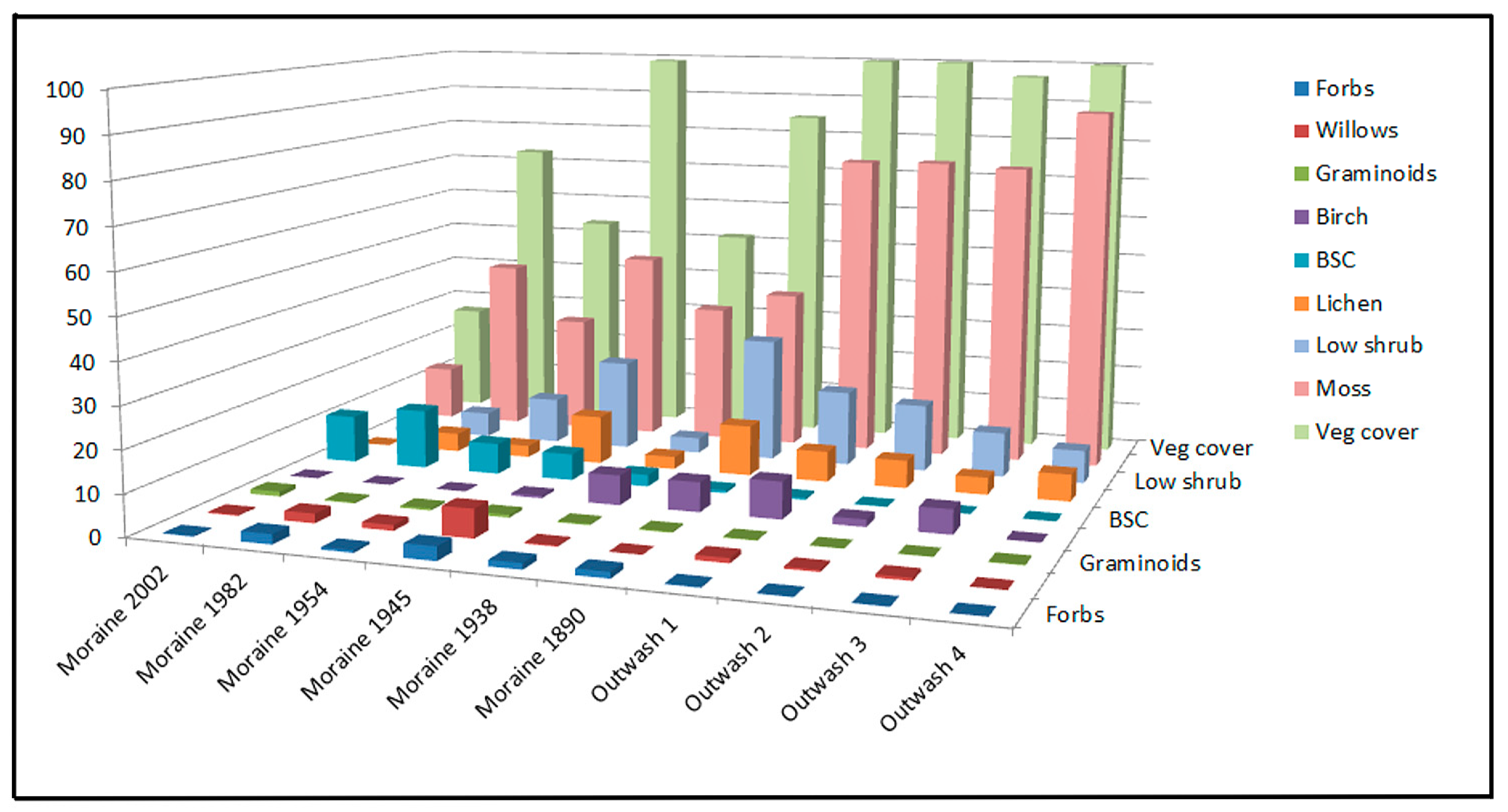
3.1. Field Observations
- Transect 1: The youngest land surface examined was the ice-ward-oriented slope of a push moraine which is proximal to the present lagoon in front of the Skaftafellsjökull (Figure 2a). The slope varied in direction from east to north-northwest, with the average facing north, and in angle from 6° to 12° (mean = 10°). This surface was estimated to have been ice free since circa 2002. This surface varies from poorly sorted gravel to (rare) boulder-sized clasts. The surface of this transect was dominated by rocks greater than 1 cm in diameter, although rock coverage varied from station to station, namely from 41.3% to 92.5% (mean = 72.1%; SD = 20.8; Table 1). Vegetative cover, including mosses, lichens, and BSCs, was also greatly variable, being from 7.8% to 58.5% and averaging 24.5% across the five stations (SD = 20.9). The vegetative component was dominated by mosses (13.3%) and BSCs (11.8%). Graminoids averaged 1.2% cover, while forbs, willows, and lichens all averaged less than 1.0% cover. Low shrub vegetation and birches were not observed in this transect.
- Transect 2: The transect line was established on the west-facing slope (away from the glacier) of the second prominent push moraine ridge away from the current lagoon (Figure 2b). The land surface at the transect stations has a general westward slope from 2° to 10° (mean = 5.6°). This surface was estimated to have been ice free since 1982. Vegetative cover dominated the surface on this transect, averaging 65.5% across all stations (SD = 17.9; Table 1). The vegetation was dominated by mosses (mean = 42.8%), which varied from 4.5% to 80.0% (SD = 33.9), and BSCs (mean = 14.5%), which also varied from a low of 1.0% to a maximum of 36.3% (SD = 17.7). Other vegetative components, specifically forbs, willows, low shrubs, and lichens were all present at levels between 1% and 10%. Graminoids were present but sparse (0.3%), and birches were absent.
- Transect 3: This transect line was located near the crest of an older push moraine located between the glacial lagoon and several prominent kettles. The location is estimated to have been ice free since circa 1954. The surface is horizontal and slopes north at a maximum angle of 15° toward the ridge which borders the foreland to the northwest. The total vegetative cover was slightly higher (mean = 51.8%) than the exposed rock and soil surface but highly variable (SD = 31.7%; Table 1). Mosses accounted for most of the vegetation, ranging from absent (at station 4) to 62.5% (mean = 29%; SD = 23.3). Low shrubs were the next most abundant group at 11.2% cover. BSCs, lichens, and willows, in decreasing order, all occurred in abundances averaging between 1% and 10%. Forbs and graminoids were present in low abundances of 0.6% and 0.1%, respectively. The non-vegetated surface comprised gravel-to-boulder-sized rocks (37.3%) and bare soil (12.2%).
- Transect 4: Transect 4 was located proximal to a prominent kettle pond on the slope facing away from the glacier of the most distal push moraine (Figure 2c). The date of exposure of this surface is estimated to be 1945. The surface at the measurement stations slopes mainly to the west at angles from 6° to 12° (mean = 9.2°). All stations in this transect were well vegetated, ranging from 70.5% to 99% (mean = 91.8%; SD = 12.2; Table 1). Mosses dominated the vegetation, ranging from 20% to 75% (mean = 47%; SD = 21.6), followed by low shrubs (mean = 22.2%; SD = 6.4%) and lichens (mean = 11.9%; SD = 4.9). Willows, BSCs, and forbs, in decreasing order, were common components of the vegetative cover (6.9%, 6.5%, and 3.3%, respectively). Birch and graminoids were minor components at less than 1% cover each.
- Transect 5: This transect was situated near the crest of a broad overridden moraine distal to the younger push moraines and intervening kettles (Figure 2d). The date of glacial retreat from this location is estimated to be 1938. The surface where the measurement stations were situated ranges from flat to sloping southeast away from the glacier at a maximum angle of 6° (mean = 3.8°). Vegetative cover was inconsistent between stations, ranging from 7.8% to 100% (mean = 51.2%; SD = 39.4; Table 1). The cover was dominated by mosses, which varied from 3% to 77% cover (mean = 34.4%; SD = 31.6). Of the remaining vegetation groups, birch was most abundant (mean = 7.2%; SD = 11.9%), followed by low shrubs (mean = 3.8%; SD = 2.2), lichens (mean = 3.1%; SD = 2.1), BSCs (mean = 2.9%; SD = 3.1), and forbs (mean = 1.5%; SD = 2.6). Graminoids and willows were minor components (each less than 1%). The nonvegetated surface was predominantly rock (mean = 53%), consisting of gravel and boulders.
- Transect 6: Transect 6 was located on the fluvial terrace of a glacial outwash channel between the broad overridden moraine (of Transect 5) and an older arcuate moraine dated to 1904 (Figure 2e). Historical imagery suggests that the area between the moraines was occupied by a glacial outwash stream in 1962 [26] and potentially more recently. At individual station locations, the surface is generally horizontal but slopes to the west a maximum of 12° at the distal margin of the terrace. The surface of the terrace was well vegetated (mean = 96.7%; SD = 5.4; Table 1) and dominated by hummocky mosses (mean = 75.1%; SD = 16.8), followed by low shrubs (mean = 18.5%; SD = 11.6). Birch was a major component (mean = 9%) but concentrated in troughs incised in the terrace and sparse to absent on the terrace flat (SD = 8.1). Lichens were common (mean = 7.5%; SD = 3.3), but willows were sparse (mean = 1.2%). BSCs, forbs, and graminoids were minor components (each less than 1%).
- Transect 7: This transect was located on fluvial outwash proximal and to the west of the oldest of the Skaftafellsjökull arcuate moraines, which dates to the most distal position of the glacier in 1890. Although the age of the moraine is well established, there are no means for dating various locations on the outwash plain in the distal foreland directly. Presumably, the age of exposure equates to the date of deposition of the outwash, which in theory could predate or postdate the formation of the terminal moraine. The surface consists of alternating broad (10–18 m wide), flat-topped bars and shallow swales (5–12 m wide), both featuring scattered protruding boulders. The surface at the stations in this transect was consistently heavily vegetated (mean = 92.3%; SD = 8.5; Table 1). Mosses dominated the vegetative cover (mean = 75.5%; SD = 21.3), with a major contribution from low shrubs (mean = 16.5%; SD = 15.3). Lichens were common although not abundant (mean = 6.8), and birch had a low abundance (mean = 1.8%). All other vegetation groups were sparse (less than 1%).
- Transect 8: This transect, located approximately 125 m to the north-northwest of Transect 7, was also situated on the outwash plain of the Skaftafellsjökull and shared a similar subdued bar and swale topography. As with Transect 7, the age of exposure of the land surface is ambiguous. The surface here was mostly vegetated, although the total vegetative cover was variable, ranging from 23.8% to 99.8% (mean = 81.8%; SD = 32.2; Table 1). Mosses again dominated the vegetation, ranging from 9.8% to 99.3% coverage (mean = 74.7%; SD = 36.7). Low shrubs were a major vegetation component (mean = 11%; SD = 9.2). Birch was common (mean = 5.9%) but primarily limited to the swales on the landscape. Lichens were common (mean = 4.2%) but not abundant. No other vegetation groups occurred at levels of 1% or above. Exposure of bare rock was variable, ranging from 0% to 75% (mean = 17.8%; SD = 32.1).
- Transect 9: This was the most distal transect of the earlier study, located on the outwash plain approximately 120 m west-northwest from Transect 8. The age of exposure of the surface is also unknown, although hypothetically, it may be older than that in the transects located closer to the oldest moraine. As for transects 7 and 8, the surface here is a series of broad, flat-topped bars and shallow swales. The surfaces at all stations in this transect were nearly fully vegetated (mean = 99.5%; SD = 0.7). The vegetative cover was predominantly moss (mean = 89.2%; SD = 15.9%; Table 1), with smaller components of low shrubs (mean = 8.1%; SD = 13.9) and lichens (mean = 6.6%; SD = 11.7). All other vegetative groups occurred at levels below 1%.
- Transect 10: This transect was the only study location which was not included in the measurements made in 2007 [12]. The transect was located on the distal side of the oldest arcuate moraine of the Skaftafellsjökull, dated to the maximum Little Ice Age glacial extent in 1890 (Figure 2f). The surface at individual stations is irregular with protruding boulders, but in general, the surface slopes from south to west at angles ranging from 10° to 18° (mean = 13°). Vegetation covered most of the surface at all stations in the transect, ranging from 54% to 95.8% (mean = 84.7%; SD = 10.9; Table 1). Mosses were common but did not dominate the surface, ranging from 11.3% to 73.8% (mean = 39.3%; SD = 22.9). Low shrubs were nearly as abundant (mean = 30.4%; SD = 15.2). Lichens were also common (mean = 12.3%; SD = 5.2). Birch and forbs were minor vegetative components (means = 3.1% and 1.5%, respectively). Other vegetative groups were insignificant.
3.2. Inter-Transect Differences
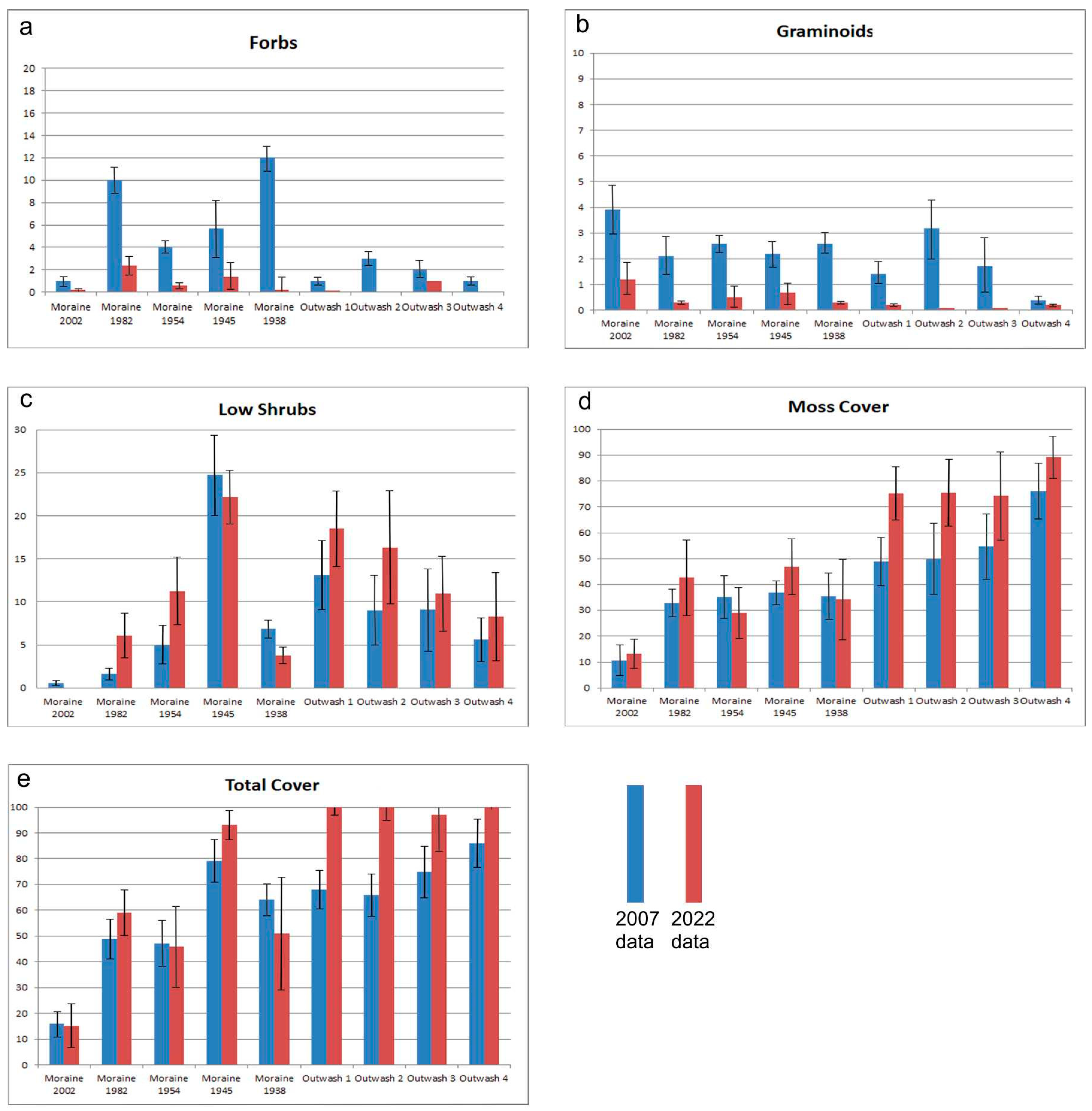
3.3. Changes from 2007 to 2022
3.3.1. Moraines
3.3.2. Outwash
4. Discussion
4.1. General Trends
4.2. Role of Substrate
4.3. Significance of Aspect
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Merzeion, B.; Cogley, J.G.; Richter, K.; Parkes, D. Attribution of global glacier mass loss to anthropogenic and natural causes. Science 2014, 345, 919–921. [Google Scholar] [CrossRef] [PubMed]
- Smiraglia, C.; Diolaiuti, G.A. (Eds.) The New Italian Glacier Inventory; Ev-K2-CNR Publ.: Bergamo, Italy, 2015. [Google Scholar]
- Hock, R.; Bliss, B.; Marzeion, B.; Giesen, R.H.; Hirabayashi, Y.; Huss, M.; Radic, V.; Slangen, A.B.A. GlacierMIP—A model intercomparison of global-scale glacier mass-balance models and projections. J. Glaciol. 2019, 65, 453–467. [Google Scholar] [CrossRef]
- Zemp, M.; Huss, M.; Thibert, E.; Eckert, N.; McNabb, R.; Huber, J.; Barandun, M.; Machguth, H.; Nussbaumer, S.U.; Gärtner-Roer, I.; et al. Global glacier mass changes and their contributions to sea-level rise from 1961 to 2016. Nature 2019, 568, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.; Malz, P.; Seehaus, T.C.; Lippl, S.; Zemp, M.; Braun, M.H. Rapid glacier retreat and downwasting throughout the European Apls in the early 21st century. Nat. Commun. 2020, 11, 3209. [Google Scholar] [CrossRef] [PubMed]
- Ficetola, G.F.; Marta, S.; Gurrieri, A.; Gobbi, M.; Ambrosini, R.; Fontaneto, D.; Zerboni, A.; Poulenard, J.; Caccianiga, M.; Thuiller, W. Dynamics of ecological communities following current retreat of glaciers. Annu. Rev. Ecol. Evol. Syst. 2021, 52, 405–426. [Google Scholar] [CrossRef]
- Matthews, J.A. The Ecology of Recently Deglaciated Terrain: A Geoecological Approach to Glacier Forelands and Primary Succession; Cambridge University Press: New York, NY, USA, 1992. [Google Scholar]
- Fickert, T.; Grüninger, F.; Damm, B. Klebelsberg revisited: Did primary succession of plants in glacier forelands a century ago differ from today? Alp. Bot. 2017, 127, 17–29. [Google Scholar] [CrossRef]
- Fickert, T. Common patterns and diverging trajectories in primary succession of plants in eastern alpine glacier forelands. Diversity 2020, 12, 191. [Google Scholar] [CrossRef]
- Bosson, J.B.; Huss, M.; Cauvy-Fraunié, S.; Clément, J.C.; Costes, G.; Fischer, M.; Poulenard, J.; Arthaud, F. Future emergence of new ecosystems caused by glacial retreat. Nature 2023, 620, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Vreeken, W.J. Principle kinds of chronosequences and their significance in soil history. J. Soil Sci. 1975, 26, 378–394. [Google Scholar] [CrossRef]
- Glausen, T.G.; Tanner, L.H. Successional trends and processes on a glacial foreland in Southern Iceland studied by repeated species counts. Ecolog. Proc. 2019, 8, 11. [Google Scholar] [CrossRef]
- Liestøl, O. The glaciers in the Kongsfjorden area, Spitsbergen. Nor. Geogr. Tidsskr. 1988, 42, 231–238. [Google Scholar] [CrossRef]
- Dowdeswell, J.A. Glaciers in the high Arctic and recent environmental change. Phil. Trans. Royal Soc. London Ser. A 1995, 352, 321–334. [Google Scholar]
- Liengen, T.; Olsen, R.A. Seasonal and site-specific variations in nitrogen fixation in a high arctic area, Ny-Ålesund, Spitsbergen. Can. J. Microbiol. 1997, 43, 759–769. [Google Scholar] [CrossRef]
- Lefauconnier, B.; Hagen, J.O.; Orbaeck, J.B.; Melvold, K.; Isaksson, E. Glacier balance trends in the Kongsfjorden area, western Spitsbergen, Svalbard, in relation to the climate. Polar Res. 1999, 18, 307–313. [Google Scholar] [CrossRef]
- Liengen, T. Environmental factors influencing the nitrogen fixation activity of free-living cyanobacteria from a high arctic area, Spitsbergen. Can. J. Microbiol. 1999, 45, 573–581. [Google Scholar] [CrossRef]
- Hodkinson, I.D.; Coulson, S.J.; Webb, N.R. Community assembly along proglacial chronosequences in the high Arctic: Vegetation and soil development in north-West Svalbard. J. Ecol. 2003, 91, 651–663. [Google Scholar] [CrossRef]
- Szymanski, W.; Maciejowski, W.; Ostafin, K.; Ziaja, W.; Sobucki, M. Impact of parent material, vegetation cover, and site wetness on variability of soil properties in proglacial areas of small glaciers along the northeastern coast of Sørkappland (SE Spitsbergen). Catena 2019, 183, 104209. [Google Scholar] [CrossRef]
- Wietrzyk-Pełka, P.; Rola, K.; Szymański, W.; Węgrzyn, M.H. Organic carbon accumulation in the glacier forelands with regard to variability of environmental conditions in different ecogenesis stages of High Arctic ecosystems. Sci. Total Environ. 2020, 717, 135151. [Google Scholar] [CrossRef]
- Jones, G.A.; Henry, G.H.R. Primary plant succession on recently deglaciated terrain in the Canadian high Arctic. J. Biogeogr. 2003, 30, 277–296. [Google Scholar] [CrossRef]
- Sigurðsson, O. Glacier variations in Iceland 1930–1960, 1960–1990 og 2003–2004. Jökull 2005, 55, 163–170. [Google Scholar] [CrossRef]
- Vilmundardóttir, O.K.; Gísladóttir, G.; Lal, R. Soil carbon accretion along an age chronosequence formed by the retreat of the Skaftafellsjökull glacier, SE-Iceland. Geomorphology 2015, 228, 124–133. [Google Scholar] [CrossRef]
- Evans, D.J.A.; Ewertowski, M.; Orton, C. Skaftafellsjöokull, Iceland: Glacial geomorphology recording glacier recession since the Little Ice Age. J. Maps 2017, 13, 358–368. [Google Scholar] [CrossRef]
- Chandler, B.M.P.; Evans, D.J.A.; Roberts, D.H. Recent retreat at a temperate Icelandic glacier in the context of the last ~80 years of climate change in the North Atlantic region. Arktos 2016, 2, 24. [Google Scholar] [CrossRef]
- Perrson, Å. The vegetation at the margin of the receding glacier Skaftafellsjökull, south eastern Iceland. Bot. Not. 1964, 117, 323–354. [Google Scholar]
- Hannesdóttir, H.; Björnsson, H.; Pálsson, F.; Aðalgeirsdóttir, G.; Guðmundsson, S. Area, volume and mass changes of southeast Vatnajökull ice cap, Iceland, from the Little Ice Age maximum in the late 19th century to 2010. Cryosph. Discuss. 2014, 8, 4681–4735. [Google Scholar]
- Huggett, R.J. Soil chronosequences, soil development, and soil evolution: A critical review. Catena 1998, 32, 155–172. [Google Scholar] [CrossRef]
- Vilmundardóttir, O.; Gísladóttir, G.; Lal, R. Early stage development of selected soil properties along the proglacial moraines of Skaftafellsjökull glacier, SE-Iceland. Catena 2014, 121, 142–150. [Google Scholar] [CrossRef]
- Lindröth, C.H. Skaftafell, Iceland: A living glacial refugium. Oikos Suppl. 1965, 6, 1–142. [Google Scholar]
- Hoinkes, H. Beitrígezurkennte des gletscherwindes. Arch. Fur Meteorol. Geophys. Und Bioklimatol. Ser. B 1954, 6, 36–53. [Google Scholar] [CrossRef]
- Geiger, R. The Climate Near the Ground; Harvard University Press: Cambridge, MA, USA, 1971. [Google Scholar]
- Tanner, L.H.; Walker, A.E.; Nivison, M.; Smith, D.L. Changes in soil composition and floral coverage on a glacial foreland chronosequence in southern Iceland. Open J. Soil Sci. 2013, 3, 191–198. [Google Scholar] [CrossRef]
- Synan, H.E.; Melfi, M.A.; Tanner, L.H. Spatial and temporal dynamics of growth of woody plant species (birch and willows) on the foreland of a retreating glacier in southern Iceland. Ecol. Proc. 2021, 10, 13. [Google Scholar] [CrossRef]
- Elven, R.; Ryvarden, L. Dispersal and primary establishment of vegetation. In Fennoscandian Tundra Ecosystems, Part 1: Plants and Microorganisms; Wielgolaski, F.E., Ed.; Springer: Berlin/Heidelberg, Germany, 1975; pp. 82–85. [Google Scholar]
- Tichit, P.; Brickle, P.; Newton, R.J.; Convey, P.; Dawson, W. Introduced species infiltrate recent stages of succession after glacial retreat on sub-Antarctic South Georgia. NeoBiota 2024, 92, 85–110. [Google Scholar] [CrossRef]
- Kjær, U.; Olsen, S.L.; Klanderud, K. Shift from facilitative to neutral interactions by the cushion plant Silene acaulis along a primary succession gradient. J. Veg. Sci. 2018, 29, 42–51. [Google Scholar] [CrossRef]
- Tanner, L.H.; Kikukawa, G.; Weits, K. Albedo on a glacial foreland at ground level and landscape scale driven by vegetation-substrate patterns. Eur. J. Environ. Sci. 2023, 13, 71. [Google Scholar] [CrossRef]
- Armesto, J.J.; Martínez, J.A. Relations between vegetation structure and slope aspect in the Mediterranean region of Chile. J. Ecol. 1978, 66, 881–889. [Google Scholar] [CrossRef]
- Walker, L.R.; Shiels, A.B.; Bellingham, P.J.; Sparrow, A.D.; Fetcher, N.; Landau, F.H.; Lodge, D.J. Changes in abiotic influences on seed plants and ferns during 18 years of primary succession on Puerto Rican landslides. J. Ecol. 2013, 101, 650–661. [Google Scholar] [CrossRef]
- Marler, T.E.; del Moral, R. Increasing topographic influence on vegetation structure during primary succession. Plant Ecol. 2018, 219, 1009–1020. [Google Scholar] [CrossRef]
- Aström, M.; Dynesius, M.; Hylander, K.; Nilsson, C. Slope aspect modifies community responses to clear-cutting in Boreal forests. Ecology 2007, 88, 749–758. [Google Scholar] [CrossRef]
- Matthews, J.A.; Whittaker, R.J. Vegetation succession on the Storbreen glacier foreland, Jotunheimen, Norway: A review. Arct. Alp. Res. 1987, 19, 385–395. [Google Scholar] [CrossRef]
- Foskett, J.I.J. The Nature and Significance of Microtopographic Effects on Vegetation Succession in Selected Glacier Forelands, Jotenheimen and Jostedalen, Norway. Ph.D. Thesis, University of Greenwich, London, UK, 1998. [Google Scholar]
- Garobotti, I.A.; Pissolito, C.I.; Villalba, R. Spatiotemporal pattern of primary succession in relation to meso-topographic gradients on recently deglaciated terrains in the Patagonian Andes. Arct. Antarc. Alp. Res. 2011, 43, 555–567. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transect 1 (−) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 0.6 | 0.1 | 0.1 | 0.1 | 0.1 | 0.2 | 0.2 |
| Willows | 1.3 | 0.0 | 0.0 | 0.1 | 0.0 | 0.3 | 0.6 |
| Moss | 36.3 | 7.8 | 2.1 | 16.3 | 4.0 | 13.3 | 14.0 |
| Lichen | 0.0 | 0.0 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Low shrub | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| BSC | 23.8 | 3.8 | 13.8 | 15.0 | 1.8 | 11.6 | 9.0 |
| Graminoids | 4.0 | 0.1 | 0.1 | 1.0 | 0.6 | 1.2 | 1.6 |
| Birch | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Soil | 0.3 | 0.0 | 15.5 | 1.5 | 0.3 | 3.5 | 6.7 |
| Rock cover | 41.3 | 90.3 | 68.8 | 67.5 | 92.5 | 72.1 | 20.8 |
| Veg. cover | 58.5 | 9.8 | 16.3 | 30.3 | 7.8 | 24.5 | 20.9 |
| Transect 2 (+) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 2.8 | 0.8 | 1.8 | 6.0 | 0.8 | 2.4 | 2.2 |
| Willows | 0.8 | 1.8 | 4.0 | 3.8 | 1.3 | 2.3 | 1.5 |
| Moss | 4.5 | 16.8 | 37.5 | 80.0 | 75.0 | 42.8 | 33.9 |
| Lichen | 0.1 | 2.3 | 13.0 | 4.8 | 3.0 | 4.6 | 5.3 |
| Low shrub | 0.1 | 3.0 | 12.0 | 1.0 | 14.3 | 6.1 | 6.6 |
| BSC | 31.3 | 36.3 | 2.5 | 1.3 | 1.0 | 14.5 | 17.7 |
| Graminoids | 0.8 | 0.1 | 0.3 | 0.1 | 0.1 | 0.3 | 0.3 |
| Birch | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Soil | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Rock cover | 60.0 | 40.0 | 27.5 | 11.8 | 12.0 | 30.3 | 20.4 |
| Veg. cover | 40.5 | 58.5 | 75.0 | 65.3 | 88.0 | 65.5 | 17.9 |
| Transect 3 (−) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 0.6 | 1.0 | 1.1 | 0.0 | 0.1 | 0.6 | 0.5 |
| Willows | 1.3 | 0.1 | 3.0 | 0.0 | 2.5 | 1.4 | 1.4 |
| Moss | 27.5 | 17.5 | 37.5 | 0.0 | 62.5 | 29.0 | 23.3 |
| Lichen | 4.3 | 0.5 | 4.0 | 0.1 | 5.8 | 2.9 | 2.5 |
| Low shrub | 20.0 | 13.8 | 4.5 | 0.0 | 17.5 | 11.2 | 8.6 |
| BSC | 2.3 | 22.5 | 6.5 | 0.0 | 6.8 | 7.6 | 8.8 |
| Graminoids | 0.1 | 2.0 | 0.1 | 0.1 | 0.1 | 0.5 | 0.8 |
| Birch | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Soil | 7.8 | 3.8 | 1.3 | 47.5 | 0.5 | 12.2 | 20.0 |
| Rock cover | 38.8 | 40.0 | 42.5 | 52.5 | 12.5 | 37.3 | 14.9 |
| Veg. cover | 55.0 | 59.5 | 57.3 | 0.3 | 87.0 | 51.8 | 31.7 |
| Transect 4 (+) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 2.3 | 0.1 | 1.8 | 6.0 | 6.3 | 3.3 | 2.7 |
| Willows | 5.0 | 13.0 | 0.1 | 9.3 | 7.0 | 6.9 | 4.8 |
| Moss | 48.8 | 58.8 | 20.0 | 32.5 | 75.0 | 47.0 | 21.6 |
| Lichen | 16.3 | 15.0 | 8.0 | 5.3 | 15.0 | 11.9 | 4.9 |
| Low shrub | 29.3 | 16.3 | 28.8 | 16.8 | 20.0 | 22.2 | 6.4 |
| BSC | 0.0 | 0.0 | 5.5 | 23.8 | 3.3 | 6.5 | 9.9 |
| Graminoids | 0.1 | 0.6 | 0.6 | 2.0 | 0.1 | 0.7 | 0.8 |
| Birch | 0.0 | 0.0 | 0.0 | 2.8 | 0.0 | 0.6 | 1.2 |
| Soil | 1.0 | 0.1 | 20.0 | 1.3 | 0.0 | 4.5 | 8.7 |
| Rock cover | 1.3 | 0.1 | 9.5 | 6.3 | 1.0 | 3.6 | 4.1 |
| Veg. cover | 97.5 | 99.5 | 70.5 | 92.5 | 99.0 | 91.8 | 12.2 |
| Transect 5 (+) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 6.0 | 0.1 | 0.1 | 1.0 | 0.1 | 1.5 | 2.6 |
| Willows | 0.8 | 0.1 | 0.0 | 0.0 | 0.8 | 0.3 | 0.4 |
| Moss | 57.5 | 23.0 | 77.0 | 3.0 | 11.3 | 34.4 | 31.6 |
| Lichen | 5.3 | 3.0 | 4.5 | 1.1 | 1.8 | 3.1 | 2.1 |
| Low shrub | 6.8 | 3.8 | 3.3 | 0.8 | 4.3 | 3.8 | 2.2 |
| BSC | 2.8 | 2.8 | 0.1 | 0.8 | 8.0 | 2.9 | 3.1 |
| Graminoids | 0.5 | 0.1 | 0.1 | 0.6 | 0.3 | 0.3 | 0.2 |
| Birch | 3.3 | 1.3 | 28.3 | 0.1 | 3.0 | 7.2 | 11.9 |
| Soil | 11.3 | 0.0 | 0.0 | 6.5 | 0.0 | 3.6 | 5.2 |
| Rock cover | 9.5 | 72.5 | 17.3 | 90.8 | 75.0 | 53.0 | 36.9 |
| Veg. cover | 84.8 | 37.0 | 100.0 | 7.8 | 26.3 | 51.2 | 39.4 |
| Transect 6 (0) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 0.1 | 0.3 | 0.3 | 0.3 | 0.1 | 0.2 | 0.1 |
| Willows | 2.5 | 0.0 | 0.1 | 3.3 | 0.1 | 1.2 | 1.6 |
| Moss | 73.3 | 82.5 | 48.3 | 77.5 | 93.8 | 75.1 | 16.8 |
| Lichen | 10.8 | 8.8 | 6.0 | 9.3 | 2.5 | 7.5 | 3.3 |
| Low shrub | 16.5 | 22.5 | 35.0 | 15.0 | 3.3 | 18.5 | 11.6 |
| BSC | 0.0 | 0.0 | 0.1 | 0.0 | 2.3 | 0.5 | 1.0 |
| Graminoids | 0.1 | 0.1 | 0.6 | 0.3 | 0.1 | 0.2 | 0.2 |
| Birch | 0.6 | 3.8 | 0.0 | 14.3 | 17.5 | 7.2 | 8.1 |
| Soil | 0.3 | 0.1 | 12.8 | 0.0 | 0.8 | 2.8 | 5.6 |
| Rock cover | 0.0 | 0.0 | 0.3 | 0.3 | 1.8 | 0.5 | 0.7 |
| Veg. cover | 99.3 | 99.8 | 87.3 | 99.8 | 97.3 | 96.7 | 5.4 |
| Transect 7 (0) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 0.1 | 0.1 | 0.1 | 0.0 | 0.1 | 0.1 | 0.0 |
| Willows | 0.0 | 0.0 | 0.1 | 2.5 | 0.0 | 0.5 | 1.1 |
| Moss | 79.8 | 93.0 | 87.0 | 38.8 | 78.8 | 75.5 | 21.3 |
| Lichen | 1.0 | 3.3 | 2.3 | 21.3 | 6.0 | 6.8 | 8.2 |
| Low shrub | 0.3 | 8.8 | 15.8 | 41.3 | 16.3 | 16.5 | 15.3 |
| BSC | 0.0 | 0.0 | 0.0 | 0.0 | 0.3 | 0.1 | 0.1 |
| Graminoids | 0.1 | 0.1 | 0.0 | 0.0 | 0.1 | 0.1 | 0.1 |
| Birch | 0.0 | 0.0 | 4.0 | 1.3 | 3.5 | 1.8 | 1.9 |
| Soil | 0.0 | 0.0 | 0.3 | 0.3 | 0.3 | 0.2 | 0.1 |
| Rock cover | 20.3 | 4.0 | 2.0 | 0.0 | 13.8 | 8.0 | 8.6 |
| Veg. cover | 79.0 | 97.3 | 97.8 | 99.8 | 87.5 | 92.3 | 8.5 |
| Transect 8 (0) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 0.1 | 0.0 | 0.1 | 0.1 | 0.0 | 0.1 | 0.1 |
| Willows | 3.3 | 0.1 | 0.0 | 0.3 | 0.0 | 0.7 | 1.4 |
| Moss | 99.3 | 93.0 | 84.0 | 87.3 | 9.8 | 74.7 | 36.7 |
| Lichen | 2.8 | 0.1 | 4.8 | 2.8 | 10.5 | 4.2 | 4.7 |
| Low shrub | 0.0 | 18.3 | 21.0 | 12.5 | 3.3 | 11.0 | 9.2 |
| BSC | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Graminoids | 0.1 | 0.1 | 0.1 | 0.1 | 0.0 | 0.1 | 0.0 |
| Birch | 0.0 | 12.5 | 13.0 | 3.8 | 0.1 | 5.9 | 6.5 |
| Soil | 0.3 | 0.0 | 0.3 | 0.0 | 0.0 | 0.1 | 0.1 |
| Rock cover | 0.0 | 4.0 | 7.0 | 3.0 | 75.0 | 17.8 | 32.1 |
| Veg. cover | 99.8 | 95.5 | 92.8 | 97.0 | 23.8 | 81.8 | 32.2 |
| Transect 9 (0) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 0.1 | 0.0 | 0.1 | 0.0 | 0.0 | 0.0 | 0.1 |
| Willows | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Moss | 96.3 | 91.8 | 98.5 | 61.3 | 98.3 | 89.2 | 15.9 |
| Lichen | 3.0 | 1.6 | 0.8 | 27.5 | 0.1 | 6.6 | 11.7 |
| Low shrub | 1.5 | 6.3 | 0.0 | 32.5 | 0.0 | 8.1 | 13.9 |
| BSC | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Graminoids | 0.1 | 0.8 | 0.1 | 0.1 | 0.1 | 0.2 | 0.3 |
| Birch | 0.0 | 0.0 | 0.3 | 0.0 | 0.0 | 0.1 | 0.1 |
| Soil | 0.0 | 0.0 | 0.0 | 1.3 | 0.0 | 0.3 | 0.6 |
| Rock cover | 0.0 | 0.0 | 0.0 | 0.0 | 1.3 | 0.3 | 0.6 |
| Veg. cover | 100.0 | 100.0 | 100.0 | 98.8 | 98.8 | 99.5 | 0.7 |
| Transect 10 (+) | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Mean | SD |
| Forbs | 6.3 | 0.0 | 0.0 | 0.1 | 1.0 | 1.5 | 2.7 |
| Willows | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Moss | 35.0 | 45.0 | 11.3 | 31.3 | 73.8 | 39.3 | 22.9 |
| Lichen | 15.5 | 15.8 | 9.4 | 12.6 | 8.4 | 12.3 | 5.2 |
| Low shrub | 35.5 | 31.3 | 52.5 | 17.5 | 15.0 | 30.4 | 15.2 |
| BSC | 0.0 | 0.3 | 0.0 | 1.0 | 2.3 | 0.7 | 1.0 |
| Graminoids | 0.3 | 0.3 | 0.3 | 0.3 | 0.1 | 0.3 | 0.1 |
| Birch | 0.0 | 0.3 | 15.0 | 0.3 | 0.1 | 3.1 | 6.7 |
| Soil | 1.8 | 2.3 | 13.3 | 0.6 | 0.5 | 3.7 | 5.4 |
| Rock cover | 7.3 | 2.5 | 0.0 | 58.8 | 15.0 | 16.7 | 24.2 |
| Veg. cover | 95.0 | 95.8 | 91.8 | 54.0 | 87.0 | 84.7 | 17.5 |
| Transects | BCIij |
|---|---|
| 1 vs. 2 | 0.485 |
| 2 vs. 3 | 0.239 |
| 3 vs. 4 | 0.314 |
| 4 vs. 5 | 0.383 |
| 5 vs. 10 | 0.383 |
| 5 vs. 6 | 0.389 |
| 6 vs. 7 | 0.149 |
| 7 vs. 8 | 0.064 |
| 8 vs. 9 | 0.128 |
| 9 vs. 10 | 0.467 |
| Transect 1 | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 |
| Moss | 1.0 | 1.0 | 1.0 | 22.5 | 26.9 |
| Graminoids | 3.5 | 3.5 | 4.8 | 6.9 | 1.0 |
| Willows | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Low shrubs | 2.0 | 0.0 | 1.0 | 0.0 | 0.0 |
| Forbs | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Birch | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Veg. cover | 7.5 | 5.5 | 7.8 | 30.4 | 28.9 |
| Transect 2 | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 |
| Moss | 37.5 | 51.0 | 19.0 | 23.0 | 33.0 |
| Graminoids | 3.5 | 0.0 | 1.0 | 4.4 | 2.5 |
| Willows | 3.3 | 1.0 | 1.0 | 5.6 | 1.3 |
| Low shrubs | 0.0 | 0.0 | 0.0 | 0.0 | 3.0 |
| Forbs | 4.5 | 4.8 | 4.5 | 14.3 | 4.2 |
| Birch | 0.0 | 1.0 | 0.0 | 0.0 | 0.0 |
| Veg. cover | 65.6 | 58.8 | 26.5 | 48.3 | 46.0 |
| Transect 3 | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 |
| Moss | 21.3 | 12.0 | 46.0 | 30.0 | 66.0 |
| Graminoids | 2.3 | 2.5 | 1.9 | 2.3 | 3.8 |
| Willows | 1.0 | 0.0 | 3.1 | 0.0 | 0.0 |
| Low shrubs | 11.9 | 1.3 | 3.1 | 0.0 | 0.0 |
| Forbs | 2.9 | 4.5 | 8.9 | 2.3 | 4.0 |
| Birch | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Veg. cover | 40.3 | 22.8 | 61.9 | 36.6 | 75.5 |
| Transect 4 | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 |
| Moss | 42.5 | 41.0 | 41.0 | 44.0 | 16.0 |
| Graminoids | 2.5 | 1.9 | 1.4 | 1.4 | 4.0 |
| Willows | 23.8 | 7.5 | 6.4 | 6.9 | 3.0 |
| Low shrubs | 36.3 | 6.1 | 13.0 | 36.3 | 25.1 |
| Forbs | 1.0 | 4.5 | 10.9 | 14.6 | 11.8 |
| Birch | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Veg. cover | 100.0 | 61.0 | 72.7 | 95.3 | 61.9 |
| Transect 5 | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 |
| Moss | 18.8 | 41.0 | 60.0 | 40.0 | 16.0 |
| Graminoids | 2.0 | 2.5 | 4.4 | 2.2 | 1.7 |
| Willows | 6.3 | 0.5 | 0.0 | 1.0 | 9.4 |
| Low shrubs | 9.8 | 4.8 | 2.8 | 6.0 | 11.3 |
| Forbs | 12.3 | 8.0 | 7.1 | 12.3 | 10.3 |
| Birch | 0.0 | 19.0 | 0.0 | 0.0 | 0.0 |
| Veg. cover | 58.8 | 75.8 | 74.3 | 61.5 | 48.7 |
| Transect 6 | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 |
| Moss | 35.0 | 61.0 | 51.0 | 29.0 | 68.0 |
| Graminoids | 1.0 | 2.6 | 1.4 | 1.9 | 0.0 |
| Willows | 0.0 | 6.3 | 1.9 | 1.9 | 0.0 |
| Low shrubs | 26.9 | 2.9 | 2.2 | 8.5 | 28.4 |
| Forbs | 3.0 | 3.1 | 0.0 | 65.6 | 74.4 |
| Birch | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Veg. cover | 37.3 | 78.5 | 15.6 | 61.0 | 86.3 |
| Transect 7 | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 |
| Moss | 32.5 | 51.0 | 68.0 | 93.0 | 3.8 |
| Graminoids | 0.5 | 8.3 | 2.0 | 3.1 | 2.0 |
| Willows | 0.0 | 0.0 | 1.0 | 0.0 | 0.0 |
| Low shrubs | 0.0 | 3.3 | 2.4 | 1.0 | 32.0 |
| Forbs | 3.0 | 2.6 | 1.0 | 5.7 | 4.3 |
| Birch | 1.0 | 1.0 | 0.0 | 1.9 | 0.0 |
| Veg. cover | 37.0 | 68.2 | 75.4 | 100.0 | 43.1 |
| Transect 8 | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 |
| Moss | 87.5 | 60.0 | 68.0 | 45.0 | 13.0 |
| Graminoids | 6.9 | 0.0 | 1.0 | 0.5 | 0.0 |
| Willows | 1.3 | 0.0 | 0.0 | 1.0 | 0.0 |
| Low shrubs | 0.5 | 1.9 | 0.6 | 28.0 | 11.0 |
| Forbs | 2.0 | 1.0 | 6.8 | 1.0 | 1.0 |
| Birch | 0.0 | 5.6 | 19.0 | 11.0 | 0.0 |
| Veg. cover | 98.1 | 71.0 | 95.2 | 88.5 | 25.0 |
| Transect 9 | Station 1 | Station 2 | Station 3 | Station 4 | Station 5 |
| Moss | 90.0 | 81.0 | 72.0 | 93.0 | 44.0 |
| Graminoids | 0.0 | 0.0 | 1.0 | 0.0 | 1.0 |
| Willows | 1.0 | 0.0 | 0.0 | 0.0 | 7.5 |
| Low shrubs | 2.5 | 18.0 | 0.0 | 3.8 | 1.9 |
| Forbs | 0.0 | 0.0 | 1.0 | 0.0 | 1.0 |
| Birch | 6.3 | 1.0 | 0.0 | 0.5 | 0.0 |
| Veg. cover | 100.0 | 100.0 | 74.0 | 97.3 | 56.4 |
| Transects | BCIij | p (t-Test) |
|---|---|---|
| 1 | 0.37 | 0.906 |
| 2 | 0.24 | 0.607 |
| 3 | 0.22 | 0.668 |
| 4 | 0.3 | 0.991 |
| 5 | 0.34 | 0.340 |
| 6 | 0.43 | 0.045 |
| 7 | 0.25 | 0.218 |
| 8 | 0.16 | 0.189 |
| 9 | 0.11 | 0.430 |
| Groups | p (t-Test) |
|---|---|
| Forbs | 0.016 |
| Graminoids | 0.001 |
| Birch | 0.286 |
| Willows | 0.091 |
| Low shrubs | 0.096 |
| Moss | 0.018 |
| Veg. cover | 0.015 |
| Group | Forbs | Graminoid | Birch | Willows | Shrubs | Mosses | Veg. Cover |
|---|---|---|---|---|---|---|---|
| Transect 1 | 0.001 | 0.033 | NA | 0.335 | 0.208 | 0.782 | 0.884 |
| Transect 2 | 0.024 | 0.061 | NA | 0.954 | 0.099 | 0.622 | 0.587 |
| Transect 3 | 0.019 | 0.015 | NA | 0.316 | 0.075 | 0.41 | 0.057 |
| Transect 4 | 0.048 | 0.109 | 0.374 | 0.591 | 0.868 | 0.509 | 0.838 |
| Transect 5 | <0.001 | 0.015 | 0.667 | 0.053 | 0.082 | 0.955 | 0.994 |
| Transect 6 | 0.049 | 0.058 | 0.117 | 0.624 | 0.674 | 0.039 | 0.504 |
| Transect 7 | 0.016 | 0.08 | 0.433 | 0.606 | 0.396 | 0.304 | 0.064 |
| Transect 8 | 0.103 | 0.29 | 0.653 | 0.57 | 0.721 | 0.67 | 0.37 |
| Transect 9 | 0.211 | 0.651 | 0.286 | 0.31 | 0.7 | 0.399 | 0.263 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanner, L.; Kikukawa, G.; Weits, K. The Temporal and Spatial Dynamics of Succession in a Glacial Foreland in Southern Iceland: The Effects of Landscape Heterogeneity. Land 2024, 13, 1055. https://doi.org/10.3390/land13071055
Tanner L, Kikukawa G, Weits K. The Temporal and Spatial Dynamics of Succession in a Glacial Foreland in Southern Iceland: The Effects of Landscape Heterogeneity. Land. 2024; 13(7):1055. https://doi.org/10.3390/land13071055
Chicago/Turabian StyleTanner, Lawrence, Genevieve Kikukawa, and Kaylyn Weits. 2024. "The Temporal and Spatial Dynamics of Succession in a Glacial Foreland in Southern Iceland: The Effects of Landscape Heterogeneity" Land 13, no. 7: 1055. https://doi.org/10.3390/land13071055