


Fluctuating Asymmetry of Fallow Deer Antlers Is Associated with Weapon Damage but Not Tactical Behaviour during Fights

Abstract
1. Introduction
2. Methods
2.1. Study Site and Population
2.2. Data Collection
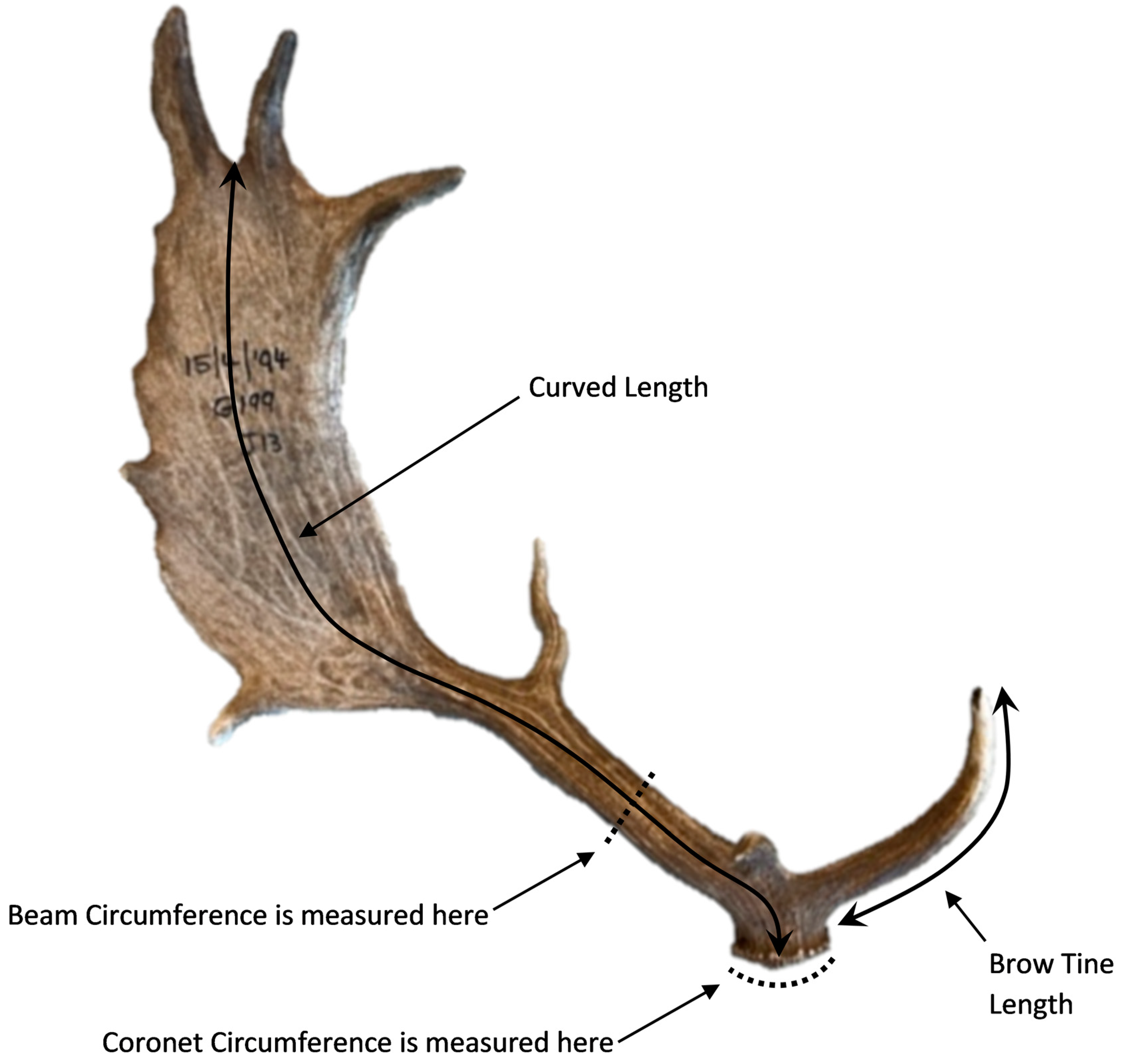
2.3. Antler Collection and Measurement
2.4. Statistical Analyses
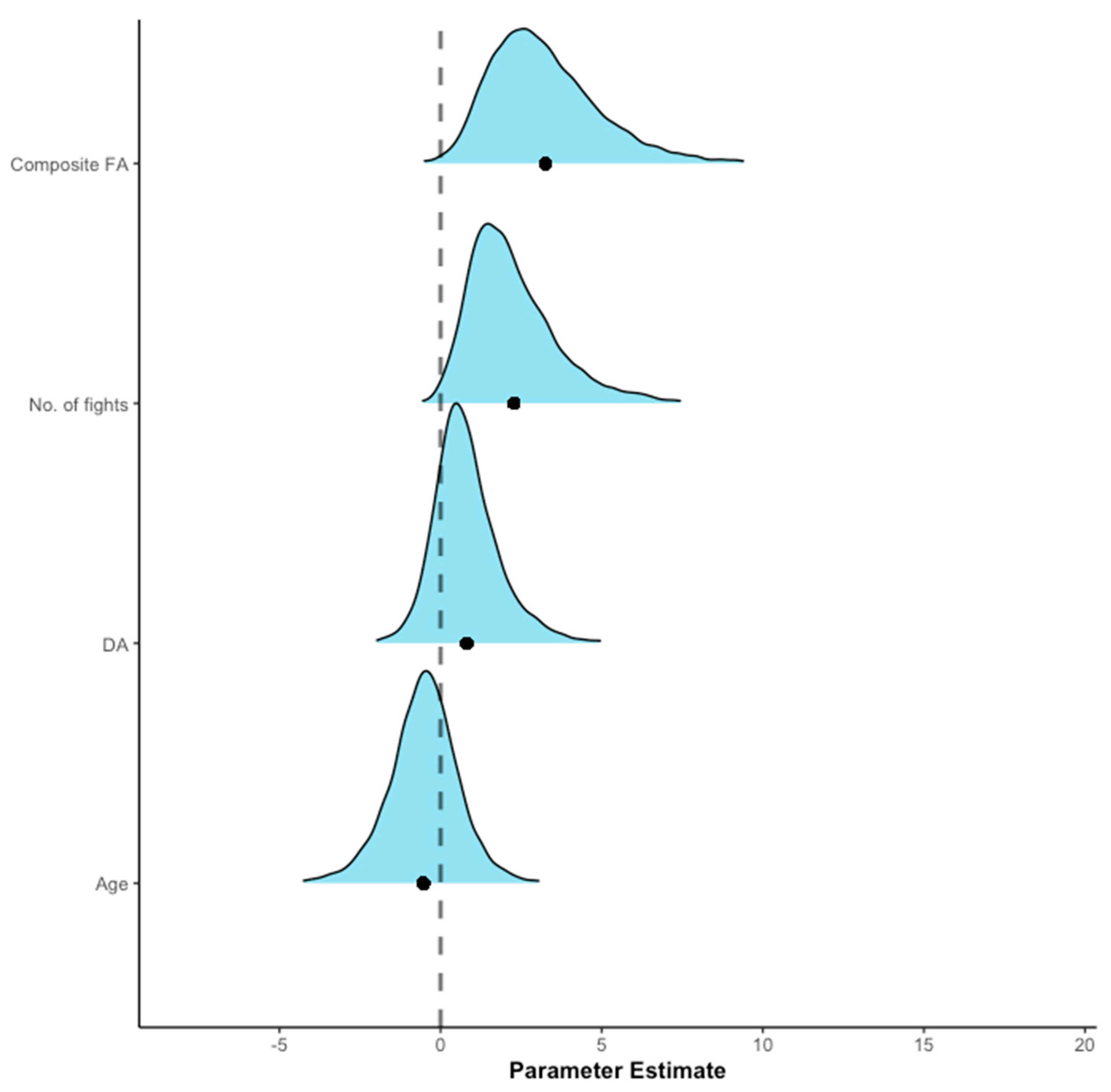
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersson, M. Sexual Selection; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Enquist, M.; Leimar, O. Evolution of fighting behaviour: Decision rules and assessment of relative strength. J. Theor. Biol. 1983, 102, 387–410. [Google Scholar] [CrossRef]
- Enquist, M.; Leimar, O.; Ljungberg, T.; Mallner, Y.; Segerdahl, N. A test of the sequential assessment game: Fighting in the cichlid fish Nanncara anomala. Anim. Behav. 1990, 40, 1–14. [Google Scholar] [CrossRef]
- Rower, S. The evolution of reliable and unreliable badges of fighting ability. Am. Zool. 1982, 22, 531–546. [Google Scholar] [CrossRef]
- Johnstone, R.A.; Norris, K. Badges of status and the cost of aggression. Behav. Ecol. Sociobiol. 1993, 32, 127–134. [Google Scholar] [CrossRef]
- Berglund, A.; Bisazza, A.; Pilastro, A. Armaments and ornaments: An evolutionary explanation of traits of dual utility. Biol. J. Linn. Soc. 1996, 58, 385–399. [Google Scholar] [CrossRef]
- Geist, V. Mountain Sheep: A Study in Behaviour and Evolution; University of Chicago Press: Chicago, IL, USA, 1971. [Google Scholar]
- Clutton-Brock, T.; Albon, S.D.; Gibson, R.M.; Guinness, F.E. The logical stag: Adaptive aspects of fighting in red deer (Cervus elaphus L.). Anim. Behav. 1979, 27, 211–225. [Google Scholar] [CrossRef]
- Barrette, C.; Vandal, D. Sparring, relative antler size, and assessment in male caribou. Behav. Ecol. Sociobiol. 1990, 26, 383–387. [Google Scholar] [CrossRef]
- Hoem, S.A.; Melis, C.; Linnell, J.D.C.; Andersen, R. Fighting behaviour in territorial male roe deer Capreolus capreolus: The effects of antler size and residence. Eur. J. Wildl. Res. 2007, 53, 1–8. [Google Scholar] [CrossRef]
- Sneddon, L.U.; Huntingford, F.A.; Taylor, A.C. Weapon size versus body size as a predictor of winning in fights between shore crabs, Carcinus maenas (L.). Behav. Ecol. Sociobiol. 1997, 41, 237–242. [Google Scholar] [CrossRef]
- Kelly, C.D. Fighting for harems: Assessment strategies during male–male contests in the sexually dimorphic Wellington tree weta. Anim. Behav. 2006, 72, 727–736. [Google Scholar] [CrossRef]
- Lailvaux, S.P.; Hathway, J.; Pomfret, J.; Knell, R.J. Horn size predicts physical performance in the beetle Euoniticellus intermedius (Coleoptera: Scarabaeidae). Funct. Ecol. 2005, 19, 632–639. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P.; Carlin, C.M.; Hayden, T.J. Is Difference in Body Weight, Antler Length, Age or Dominance Rank related to the Number of Fights between Fallow Deer (Dama dama)? Ethology 2006, 112, 258–269. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry: Measurement, analysis, patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Parsons, P.A. Fluctuating Asymmetry: An Epigenetic Measure of Stress. Biol. Rev. 1990, 65, 131–145. [Google Scholar] [CrossRef]
- Solberg, E.J.; Sæther, B.E. Fluctuating asymmetry in the antlers of moose (Alces alces): Does it signal male quality? Proc. R. Soc. B 1993, 254, 251–255. [Google Scholar]
- Møller, A.P.; Swaddle, J.P. Asymmetry, Developmental Stability, and Evolution; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Chirichella, R.; Rocca, M.; Brugnoli, A.; Mustoni, A.; Apollonio, M. Fluctuating asymmetry in Alpine chamois horns: An indicator of environmental stress. Evol. Ecol. 2020, 34, 573–587. [Google Scholar] [CrossRef]
- Folstad, I.; Arneberg, P.; Karter, A.J. Antlers and parasites. Oecologia 1996, 105, 556–558. [Google Scholar] [CrossRef]
- Pélabon, C.; van Breukelen, L. Asymmetry in antler size in roe deer (Capreolus capreolus): An index of individual and population conditions. Oecologia 1998, 116, 1–8. [Google Scholar] [CrossRef]
- Lens, L.; van Dongen, S.; Kark, S.; Matthysen, E. Fluctuating asymmetry as an indicator of fitness: Can we bridge the gap between studies? Biol. Rev. 2002, 77, 27–38. [Google Scholar] [CrossRef]
- Bartoš, L.; Bahbouh, R. Antler size and fluctuating asymmetry in red deer (Cervus elaphus) stags and probability of becoming a harem holder in rut. Biol. J. Linn. Soc. 2006, 87, 59–68. [Google Scholar] [CrossRef]
- Ciuti, S.; Apollonio, M. Do antlers honestly advertise the phenotypic quality of fallow buck (Dama dama) in a lekking population? Ethology 2011, 117, 133–144. [Google Scholar] [CrossRef]
- Kruuk, L.E.B.; Slate, J.; Pemberton, J.M.; Clutton-Brock, T.H. Fluctuating asymmetry in a secondary sexual trait: No associations with individual fitness, environmental stress or inbreeding, and no heritability. J. Evol. Biol. 2003, 16, 101–113. [Google Scholar] [CrossRef]
- Pélabon, C.; Joly, P. What, if anything, does visual asymmetry in fallow deer antlers reveal? Anim. Behav. 2000, 59, 193–199. [Google Scholar] [CrossRef]
- Chapman, D.; Chapman, N. Fallow Deer: Their History, Distribution, and Biology; Coch-y-Bonddu Books: Machynlleth, UK, 1997. [Google Scholar]
- Emlen, D.J. The Evolution of Animal Weapons. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 387–413. [Google Scholar] [CrossRef]
- Moen, R.; Pastor, J. Simulating antler growth and energy, nitrogen, calcium and phosphorus metabolism in caribou. Rangifer 1998, 18, 85–97. [Google Scholar] [CrossRef]
- Moen, R.A.; Pastor, J.; Cohen, J. Antler growth and extinction of Irish elk. Evol. Ecol. Res. 1999, 1, 235–249. [Google Scholar]
- Landete-Castillejos, T.; Estevez, J.A.; Ceacero, F.; Garcia, A.J.; Gallego, L. A review of factors effecting antler composition and mechanics. Front. Biosci. 2012, E4, 2328–2339. [Google Scholar] [CrossRef]
- Landete-Castillejos, T.; Estevez, J.; Martínez, A.; Ceacero, F.; Garcia, A.; Gallego, L. Does chemical composition of antler bone reflect the physiological effort made to grow it? Bone 2007, 40, 1095–1102. [Google Scholar] [CrossRef]
- Bartoš, L.; Perner, V.; Losos, S. Red deer stags rank position, body weight and antler growth. Acta Thériol. 1988, 33, 209–217. [Google Scholar] [CrossRef]
- Bartoš, L.; Losos, S. Response of antler growth to changing rank of fallow deer buck during the velvet period. Can. J. Zoöl. 1997, 75, 1934–1939. [Google Scholar] [CrossRef]
- Alvarez, F. Risks of fighting in relation to age and territory holding in fallow deer. Can. J. Zool. 1993, 71, 376–383. [Google Scholar] [CrossRef]
- Alvarez, F. Functional directional asymmetry in fallow deer (Dama dama) antlers. J. Zool. 1995, 236, 563–569. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P.; Carlin, C.M.; Hayden, T.J. Does Lateral Presentation of the Palmate Antlers During Fights by Fallow Deer (Dama dama L.) Signify Dominance or Submission? Ethology 2002, 108, 389–401. [Google Scholar] [CrossRef]
- Jennings, D.J. Contest behaviour varies in relation to reproductive opportunities and reproductive success in the fallow deer. Anim. Behav. 2020, 163, 95–103. [Google Scholar] [CrossRef]
- Bartoš, L. Relationships between Behaviour and Antler Cycle Timing in Red Deer. Ethology 1986, 71, 305–314. [Google Scholar] [CrossRef]
- Johnson, H.E.; Bleich, V.C.; Krausman, P.R.; Koprowski, J.L. Effects of antler breakage on mating behavior (Cervus elephus nannodes). Eur. J. Wildl. Res. 2007, 53, 9–15. [Google Scholar] [CrossRef]
- Lincoln, G.A. The role of antlers in the behaviour of red deer. J. Exp. Zoöl. 1972, 182, 233–249. [Google Scholar] [CrossRef]
- Lincoln, G.A.; Youngson, R.W.; Short, R.V. The social and sexual behaviour of the red deer stag. J. Reprod. Fertil. Suppl. 1970, 11, 71–103. [Google Scholar]
- Jennings, D.J.; Boys, R.J.; Gammell, M.P. Weapon damage is associated with contest dynamics but not mating success in fallow deer (Dama dama). Biol. Lett. 2017, 13, 20170565. [Google Scholar] [CrossRef] [PubMed]
- Munoz, N.E.; Zink, A.G. Asymmetric Forceps Increase Fighting Success among Males of Similar size in the Maritime Earwig. Ethology 2012, 118, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, L.U.; Swaddle, J.P. Asymmetry and fighting performance in the shore crab Carcinus maenas. Anim. Behav. 1999, 58, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, P.; Matyjasiak, P. Chaffinch (Fringilla Coelebs) Epaulette Display Depends on the Degree of Exposure But Not Symmetry of Intruder’s Epaulettes. Behaviour 1997, 134, 1115–1121. [Google Scholar] [CrossRef]
- Payne, R.J. Gradually escalating fights and displays: The cumulative assessment model. Anim. Behav. 1998, 56, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Payne, R.J.; Pagel, M. Escalation and Time Costs in Displays of Endurance. J. Theor. Biol. 1996, 183, 185–193. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P.; Carlin, C.M.; Hayden, T.J. Win, lose or draw: A comparison of fight structure based on fight conclusion in the fallow deer. Behaviour 2005, 142, 423–439. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P.; Payne, R.J.H.; Hayden, T.J. An Investigation of Assessment Games During Fallow Deer Fights. Ethology 2005, 111, 511–525. [Google Scholar] [CrossRef]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef]
- Haldane, J.B.S. The measurement of variation. Evolution 1955, 9, 484. [Google Scholar] [CrossRef]
- Palmer, A.R. Fluctuating asymmetry analyses: A primer. In Developmental Instability: Its Origins and Evolutionary Implications; Markow, T.A., Ed.; Kluwer: Dordrecht, The Netherlands, 1994; pp. 335–364. [Google Scholar]
- Ditchkoff, S.S.; de Freese, R.L. Assessing fluctuating asymmetry of white-tailed deer antlers in a three-dimensional context. J. Mammal. 2010, 91, 27–37. [Google Scholar] [CrossRef]
- Tsuboi, M.M.; Kopperud, B.T.; Syrowatka, C.; Grabowski, M.; Voje, K.; Pélabon, C.; Hansen, T.F. Measuring complex morphological traits with 3D photogrammetry: A case study with deer antlers. Evol. Biol. 2020, 47, 175–186. [Google Scholar] [CrossRef]
- Leung, B.; Forbes, M.R.; Houle, D. Fluctuating asymmetry as a bioindicator of stress: Comparing efficacy of analyses involving multiple traits. Am. Nat. 2000, 155, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Gelman, A.; Jakulin, A.; Pittau, M.G.; Su, Y.-S. A weakly informative default prior distribution for logistic and other regression models. Ann. Appl. Stat. 2008, 2, 1360–1383. [Google Scholar] [CrossRef]
- Kruschke, J.K. Doing Bayesian Data Analysis; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Gelman, A.; Carlin, J.; Stern, H.; Rubin, D. Bayesian Data Analysis; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Jennings, D.J.; Boys, R.J.; Gammell, M.P. Suffering third-party intervention during fighting is associated with reduced mating success in the fallow deer. Anim. Behav. 2018, 139, 1–8. [Google Scholar] [CrossRef]
- Bridger, D.; Bonner, S.J.; Briffa, M. Individual quality and personality: Bolder males are less fecund in the hermit crab Pagurus bernhardus. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142492. [Google Scholar] [CrossRef] [PubMed]
- Leamy, L.J.; Klingenberg, C.P. The Genetics and Evolution of Fluctuating Asymmetry. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 1–21. [Google Scholar] [CrossRef]
- Polak, M. (Ed.) Developmental Instability: Causes and Consequences; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Møller, A.P.; Pomiankowski, A. Fluctuating asymmetry and sexual selection. Genetica 1993, 89, 267–279. [Google Scholar] [CrossRef]
- Møller, A.P. Fluctuating asymmetry in male sexual ornaments may reliably reveal male quality. Anim. Behav. 1990, 40, 1185–1187. [Google Scholar] [CrossRef]
- Mateos, C.; Alarcos, S.; Carranza, J.; Sánchez-Prieto, C.B.; Valencia, J. Fluctuating asymmetry of read deer antlers negatively relates to individual condition and proximity to prime age. Anim. Behav. 2008, 75, 1629–1640. [Google Scholar] [CrossRef]
- Putman, R.J.; Sullivan, M.S. Fluctuating asymmetry in antlers of fallow deer (Dama dama): The relative roles of environmental stress and sexual selection. Biol. J. Linn. Soc. 2000, 70, 27–36. [Google Scholar] [CrossRef]
- Ditchkoff, S.S.; Lochmiller, R.L.; Masters, R.E.; Starry, W.R.; Leslie, D.M., Jr. Does fluctuating asymmetry of antlers in white-tailed deer (Odocoileus virginianus) follow patters predicted for sexually selected traits? Proc. R. Soc. Biol. Sci. B 2001, 268, 891–898. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H. The Functions of Antlers. Behaviour 1982, 79, 108–124. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P. Bystander fallow deer engage in third-party behaviour based on similarities in contestant resource-holding potential. Anim. Behav. 2022, 191, 33–41. [Google Scholar] [CrossRef]
- Hayden, T.J.; Lynch, J.M.; O’Corry-Crowe, G. Antler growth and morphology in a feral sika deer (Cervus nippon) population in Killarney, Ireland. J. Zoöl. 1994, 232, 21–35. [Google Scholar] [CrossRef]
- Briffa, M.; Sneddon, L.U. Physiological constraints on contest behaviour. Funct. Ecol. 2007, 21, 627–637. [Google Scholar] [CrossRef]
- Stewart, K.M.; Bowyer, R.T.; Kie, J.G.; Gawaway, W.C. Antler size relative to body mass in moose: Tradeoffs associated with reproduction. Alces 2000, 36, 77–83. [Google Scholar]
- Vanpé, C.; Gaillard, J.-M.; Kjellander, P.; Mysterud, A.; Magnien, P.; Delorme, D.; van Laere, G.; Klein, F.; Liberg, O.; Hewison, A.J.M. Antler size provides an honest signal of male phenotypic quality in roe deer. Am. Nat. 2007, 169, 481–493. [Google Scholar] [CrossRef]
- Arnott, G.; Elwood, R.W. Information gathering and decision making about resource value in animal contests. Anim. Behav. 2008, 76, 529–542. [Google Scholar] [CrossRef]
- Hardy, I.C.W.; Briffa, M. Animal Contests; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Clutton-Brock, T.; Albon, S.D. The Roaring of Red Deer and the Evolution of Honest Advertisement. Behaviour 1979, 69, 145–170. [Google Scholar] [CrossRef]
- Briffa, M.; Elwood, R.W.; Dick, J.T.A. Analysis of repeated signals during shell fights in the hermit crab Pagurus bernhardus. Proc. R. Soc. Biol. Sci. B 1998, 265, 1467–1474. [Google Scholar] [CrossRef]
- Green, P.A.; Patek, S.N. Mutual assessment during ritualized fighting in mantis shrimp (Stomatopoda). Proc. Biol. Sci. 2018, 285, 20172542. [Google Scholar] [CrossRef]
- Thornhill, R. Fluctuating asymmetry and the mating system of the Japanese scorpionfly, Panorpa japonica. Anim. Behav. 1992, 44, 867–879. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P.; Carlin, C.M.; Hayden, T.J. Effect of body weight, antler length, resource value and experience on fight duration and intensity in fallow deer. Anim. Behav. 2004, 68, 213–221. [Google Scholar] [CrossRef]
- Møller, A.P.; Thornhill, R. A meta-analysis of the heritability of developmental stability. Am. Nat. 1997, 151, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. Patterns of fluctuating asymmetry in weapons: Evidence for reliable signalling of quality in beetle horns and bird spurs. Proc. R. Soc. B Biol. Sci. 1992, 248, 199–206. [Google Scholar] [CrossRef]
- Tomkins, J.L.; Simmons, L. Fluctuating asymmetry and sexual selection: Paradigm shifts, publication bias, and observer expectation. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: New York, NY, USA, 2003; pp. 231–261. [Google Scholar]
- Simmons, L.W.; Tomkins, J.L.; Kotiaho, J.S.; Hunt, J. Fluctuating paradigm. Proc. R. Soc. B 1999, 266, 593–595. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter | Mean | s.d. | 95% CI | %Px |
|---|---|---|---|---|
| Intercept | 20.76 | 2.11 | 16.55, 24.95 | |
| Duration | −1.13 | 2.18 | −5.39, 3.18 | 70.19 |
| Backward pushes | −0.15 | 2.87 | −5.41, 5.78 | 47.90 |
| Jump clashes | −0.66 | 2.81 | −6.21, 4.83 | 59.39 |
| Retreats | −1.99 | 2.20 | −6.23, 2.39 | 81.66 |
| Parameter | Mean | s.d. | 95% CI | %Px |
|---|---|---|---|---|
| Intercept | 1.76 | 0.84 | 0.11, 3.43 | |
| Duration | −0.85 | 0.92 | −2.68, 0.94 | 81.93 |
| Backward pushes | −0.76 | 1.23 | −3.14, 1.67 | 74.20 |
| Jump clashes | 0.76 | 1.19 | −3.15, 1.60 | 75.15 |
| Retreats | 0.98 | 0.89 | −0.76, 2.74 | 13.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jennings, D.J.; Gammell, M.P. Fluctuating Asymmetry of Fallow Deer Antlers Is Associated with Weapon Damage but Not Tactical Behaviour during Fights. Symmetry 2023, 15, 829. https://doi.org/10.3390/sym15040829
Jennings DJ, Gammell MP. Fluctuating Asymmetry of Fallow Deer Antlers Is Associated with Weapon Damage but Not Tactical Behaviour during Fights. Symmetry. 2023; 15(4):829. https://doi.org/10.3390/sym15040829
Chicago/Turabian StyleJennings, Dómhnall J., and Martin P. Gammell. 2023. "Fluctuating Asymmetry of Fallow Deer Antlers Is Associated with Weapon Damage but Not Tactical Behaviour during Fights" Symmetry 15, no. 4: 829. https://doi.org/10.3390/sym15040829
APA StyleJennings, D. J., & Gammell, M. P. (2023). Fluctuating Asymmetry of Fallow Deer Antlers Is Associated with Weapon Damage but Not Tactical Behaviour during Fights. Symmetry, 15(4), 829. https://doi.org/10.3390/sym15040829