
Neuron–Oligodendrocyte Communication in Myelination of Cortical GABAergic Cells



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. GABAergic Neuron Properties
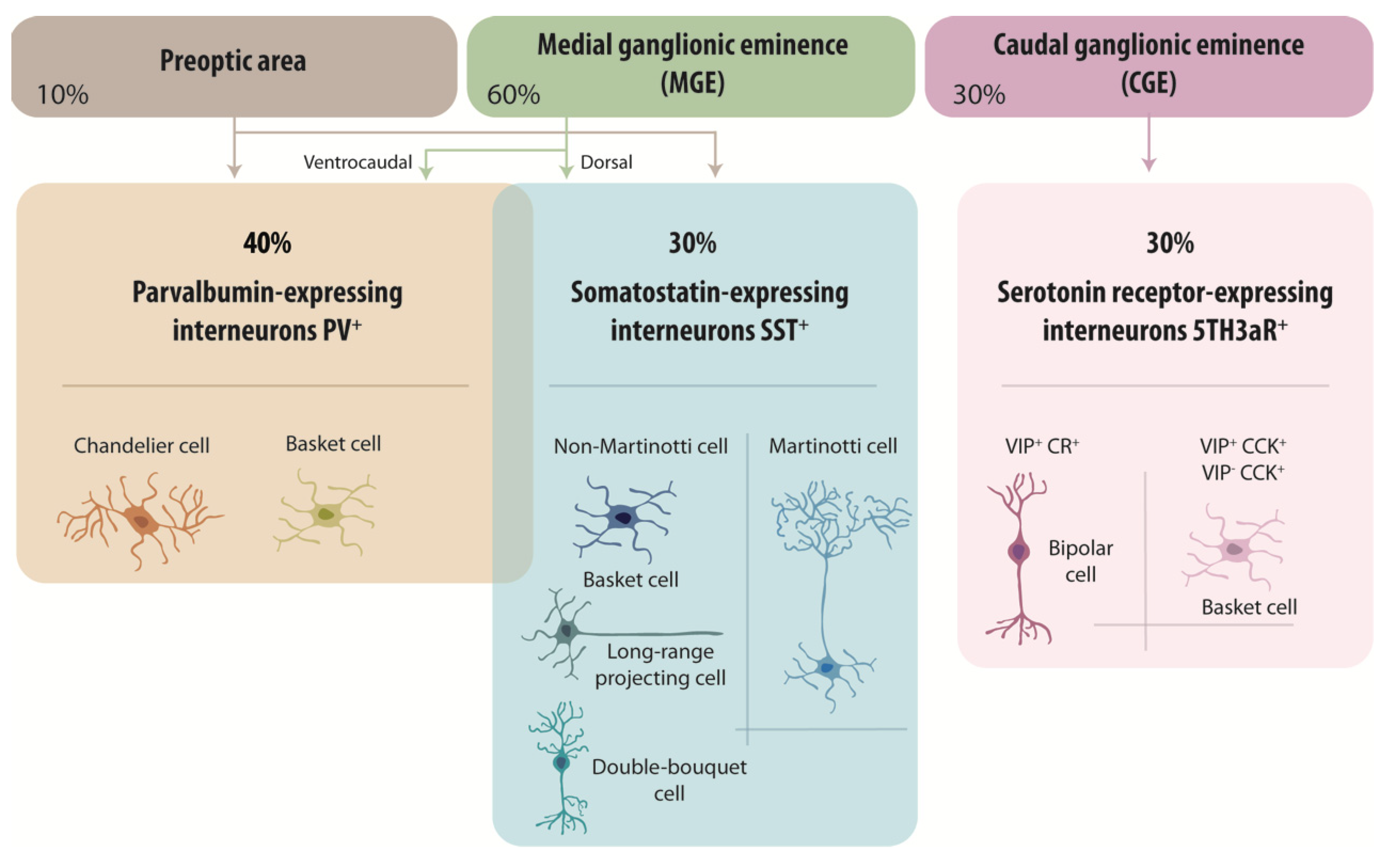
2.1. GABAergic Neuron Diversity and Origins
2.2. GABAergic Neuron Functions
3. Oligodendroglial Cells and Their Interactions with Neurons
3.1. Oligodendroglial Lineage Cells
3.2. Oligodendroglial Cell Interactions with Neurons
3.2.1. Axon Myelination
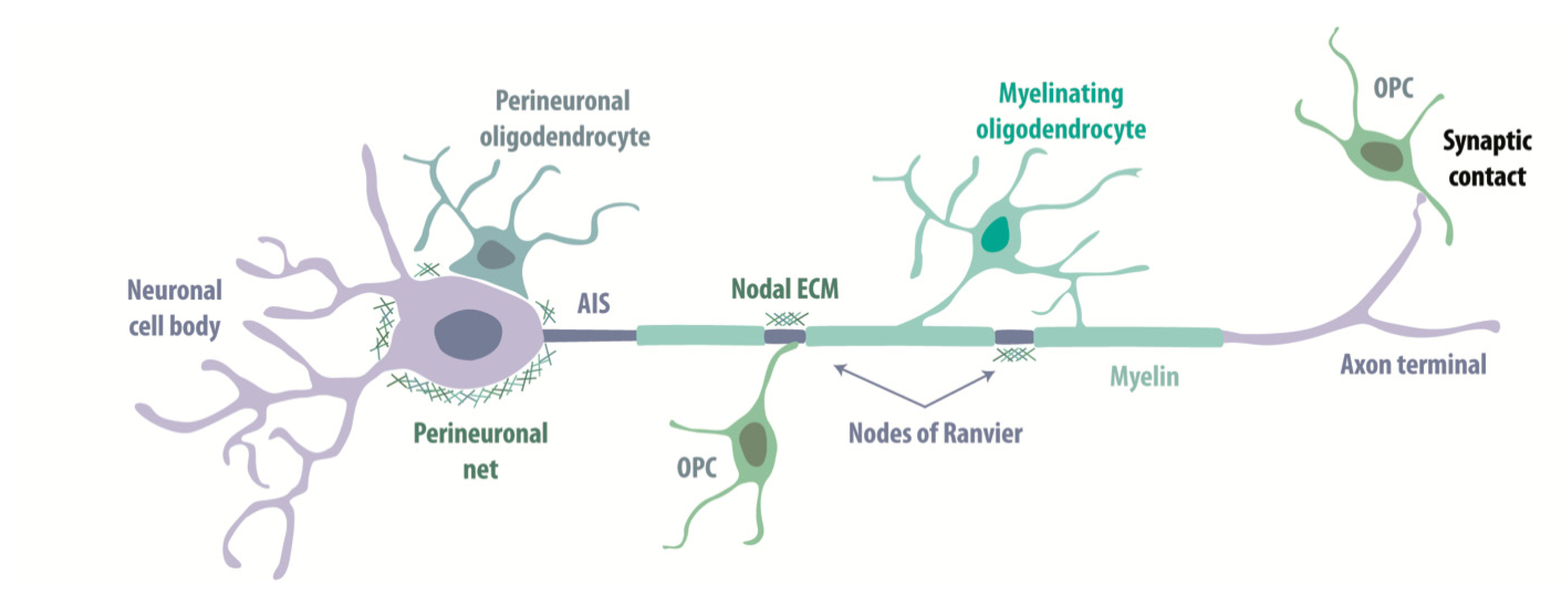
3.2.2. Perineuronal Interactions
3.2.3. Nodal Interactions
3.2.4. Perineuronal Nets
3.3. Effects of Oligodendrocyte Lineage Cells on Synapses
4. Myelination of GABAergic Neurons
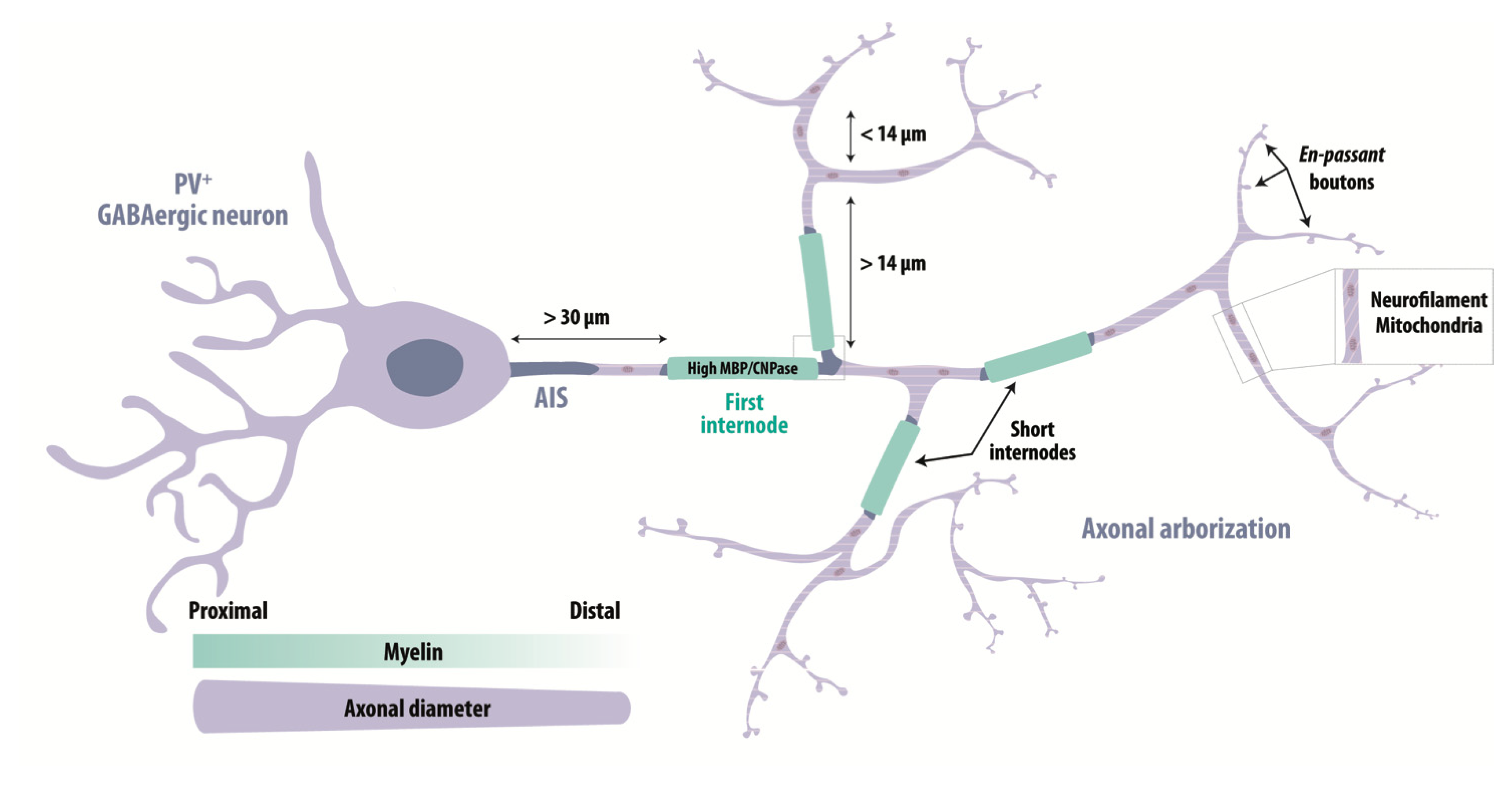
4.1. Identification and Localization of Myelinated Axons
4.2. Characteristics of Myelinated GABAergic Axons
4.3. Prenodes Are Formed before Myelination of Hippocampal GABAergic Neurons
5. Myelin, Axonal Conduction and Neural Circuit Function
5.1. Determinants of Action Potential Propagation along Myelinated Fibers
5.2. Effects of Myelination on GABAergic Neurons
5.3. Contributions of Myelination to Sensory Processing and Learning
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baldwin, K.T.; Eroglu, C. Molecular mechanisms of astrocyte-induced synaptogenesis. Curr. Opin. Neurobiol. 2017, 45, 113–120. [Google Scholar] [CrossRef]
- Li, Q.; Barres, B.A. Microglia and macrophages in brain homeostasis and disease. Nat. Rev. Immunol. 2018, 18, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Seidl, A.H. Regulation of conduction time along axons. Neuroscience 2014, 276, 126–134. [Google Scholar] [CrossRef]
- Monje, M. Myelin Plasticity and Nervous System Function. Annu. Rev. Neurosci. 2018, 41, 61–76. [Google Scholar] [CrossRef]
- Fünfschilling, U.; Supplie, L.M.; Mahad, D.; Boretius, S.; Saab, A.S.; Edgar, J.; Brinkmann, B.G.; Kassmann, C.M.; Tzvetanova, I.D.; Möbius, W.; et al. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 2012, 485, 517–521. [Google Scholar] [CrossRef]
- Lee, Y.; Morrison, B.M.; Li, Y.; Lengacher, S.; Farah, M.H.; Hoffman, P.N.; Liu, Y.; Tsingalia, A.; Jin, L.; Zhang, P.-W.; et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nat. Cell Biol. 2012, 487, 443–448. [Google Scholar] [CrossRef]
- Saab, A.S.; Tzvetavona, I.D.; Trevisiol, A.; Baltan, S.; Dibaj, P.; Kusch, K.; Möbius, W.; Goetze, B.; Jahn, H.M.; Huang, W.; et al. Oligodendroglial NMDA Receptors Regulate Glucose Import and Axonal Energy Metabolism. Neuron 2016, 91, 119–132. [Google Scholar] [CrossRef]
- Jinno, S.; Klausberger, T.; Marton, L.F.; Dalezios, Y.; Roberts, J.D.B.; Fuentealba, P.; Bushong, E.A.; Henze, D.; Buzsáki, G.; Somogyi, P. Neuronal Diversity in GABAergic Long-Range Projections from the Hippocampus. J. Neurosci. 2007, 27, 8790–8804. [Google Scholar] [CrossRef] [PubMed]
- Micheva, K.D.; Wolman, D.; Mensh, B.D.; Pax, E.; Buchanan, J.; Smith, S.J.; Bock, D.D. A large fraction of neocortical myelin ensheathes axons of local inhibitory neurons. eLife 2016, 5, e15784. [Google Scholar] [CrossRef] [PubMed]
- Micheva, K.D.; Chang, E.F.; Nana, A.L.; Seeley, W.W.; Ting, J.T.; Cobbs, C.; Lein, E.; Smith, S.J.; Weinberg, R.J.; Madison, D.V. Distinctive Structural and Molecular Features of Myelinated Inhibitory Axons in Human Neocortex. eNeuro 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Stedehouder, J.; Couey, J.J.; Brizee, D.; Hosseini, B.; Slotman, J.A.; Dirven, C.M.F.; Shpak, G.; Houtsmuller, A.B.; Kushner, S.A. Fast-spiking Parvalbumin Interneurons are Frequently Myelinated in the Cerebral Cortex of Mice and Humans. Cereb. Cortex 2017, 27, 5001–5013. [Google Scholar] [CrossRef]
- Freund, T.F.; Buzsáki, G. Interneurons of the Hippocampus. Hippocampus 1996, 6, 347–470. [Google Scholar] [CrossRef]
- Rudy, B.; Fishell, G.; Lee, S.; Hjerling-Leffler, J. Three groups of interneurons account for nearly 100% of neocortical GABAergic neurons. Dev. Neurobiol. 2011, 71, 45–61. [Google Scholar] [CrossRef]
- Hu, H.; Gan, J.; Jonas, P. Fast-spiking, parvalbumin+ GABAergic interneurons: From cellular design to microcircuit function. Science 2014, 345, 1255263. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.; Mi, D.; Llorca, A.; Marín, O. Development and functional diversification of cortical interneurons. Neuron 2018, 100, 294–313. [Google Scholar] [CrossRef]
- Fishell, G.; Kepecs, A. Interneuron Types as Attractors and Controllers. Annu. Rev. Neurosci. 2020, 43, 1–30. [Google Scholar] [CrossRef]
- The Petilla Interneuron Nomenclature Group (PING). Petilla terminology: Nomenclature of features of GABAergic interneurons of the cerebral cortex. Nat. Rev. Neurosci. 2008, 9, 557–568. [Google Scholar] [CrossRef]
- Nassar, M.; Simonnet, J.; Lofredi, R.; Cohen, I.; Savary, E.; Yanagawa, Y.; Miles, R.; Fricker, D. Diversity and overlap of parvalbumin and somatostatin expressing interneurons in mouse presubiculum. Front. Neural Circuits 2015, 9, 20. [Google Scholar] [CrossRef] [PubMed]
- Jinno, S.; Kosaka, T. Colocalization of Parvalbumin and Somatostatin-like Immunoreactivity in the Mouse Hippocampus: Quantitative Analysis with Optical Disector. J. Comp. Neurol. 2000, 428, 377–388. [Google Scholar] [CrossRef]
- Zeisel, A.; Muñoz-Manchado, A.B.; Codeluppi, S.; Lönnerberg, P.; La Manno, G.; Juréus, A.; Marques, S.; Munguba, H.; He, L.; Betsholtz, C.; et al. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 2015, 347, 1138–1142. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, A.; Hochgerner, H.; Lönnerberg, P.; Johnsson, A.; Memic, F.; van der Zwan, J.; Häring, M.; Braun, E.; Borm, L.E.; La Manno, G.; et al. Molecular Architecture of the Mouse Nervous System. Cell 2018, 174, 999–1014.e22. [Google Scholar] [CrossRef] [PubMed]
- Cembrowski, M.S.; Wang, L.; Sugino, K.; Shields, B.C.; Spruston, N. Hipposeq: A comprehensive RNA-seq database of gene expression in hippocampal principal neurons. eLife 2016, 5, e14997. [Google Scholar] [CrossRef]
- Tasic, B.; Yao, Z.; Graybuck, L.T.; Smith, K.A.; Nguyen, T.N.; Bertagnolli, D.; Goldy, J.; Garren, E.; Economo, M.N.; Viswanathan, S.; et al. Shared and distinct transcriptomic cell types across neocortical areas. Nat. Cell Biol. 2018, 563, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Gouwens, N.W.; Sorensen, S.A.; Baftizadeh, F.; Budzillo, A.; Lee, B.R.; Jarsky, T.; Alfiler, L.; Baker, K.; Barkan, E.; Berry, K.; et al. Toward an Integrated Classification of Cell Types: Morphoelectric and Transcriptomic Characterization of Individual GABAergic Cortical Neurons. SSRN Electron. J. 2020. [Google Scholar] [CrossRef]
- Yuste, R.; Hawrylycz, M.; Aalling, N.; Aguilar-Valles, A.; Arendt, D.; Arnedillo, R.A.; Ascoli, G.A.; Bielza, C.; Bokharaie, V.; Bergmann, T.B.; et al. A community-based transcriptomics classification and nomenclature of neocortical cell types. Nat. Neurosci. 2020, 23, 1456–1468. [Google Scholar] [CrossRef]
- Tasic, B.; Menon, V.; Nguyen, T.N.; Kim, T.K.; Jarsky, T.; Yao, Z.; Levi, B.P.; Gray, L.T.; Sorensen, S.A.; Dolbeare, T.A.; et al. Adult mouse cortical cell taxonomy revealed by single cell transcriptomics. Nat. Neurosci. 2016, 19, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Gouwens, N.W.; Sorensen, S.A.; Berg, J.; Lee, C.; Jarsky, T.; Ting, J.; Sunkin, S.M.; Feng, D.; Anastassiou, C.A.; Barkan, E.; et al. Classification of electrophysiological and morphological neuron types in the mouse visual cortex. Nat. Neurosci. 2019, 22, 1182–1195. [Google Scholar] [CrossRef]
- Paul, A.; Crow, M.; Raudales, R.; He, M.; Gillis, J.; Huang, Z.J. Transcriptional Architecture of Synaptic Communication Delineates GABAergic Neuron Identity. Cell 2017, 171, 522–539. [Google Scholar] [CrossRef] [PubMed]
- Flames, N.; Pla, R.; Gelman, D.M.; Rubenstein, J.L.R.; Puelles, L.; Marín, O. Delineation of Multiple Subpallial Progenitor Domains by the Combinatorial Expression of Transcriptional Codes. J. Neurosci. 2007, 27, 9682–9695. [Google Scholar] [CrossRef]
- Anderson, S.A.; Kaznowski, C.E.; Horn, C.; Rubenstein, J.L.; McConnell, S.K. Distinct Origins of Neocortical Projection Neurons and Interneurons In Vivo. Cereb. Cortex 2002, 12, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Hansen, D.V.; Lui, J.H.; Flandin, P.; Yoshikawa, K.; Rubenstein, J.L.; Alvarez-Buylla, A.; Kriegstein, A.R. Non-epithelial stem cells and cortical interneuron production in the human ganglionic eminences. Nat. Neurosci. 2013, 16, 1576–1587. [Google Scholar] [CrossRef]
- Ma, T.; Wang, C.; Wang, L.; Zhou, X.; Tian, M.; Zhang, Q.; Zhang, Y.; Li, J.; Liu, Z.; Cai, Y.; et al. Subcortical origins of human and monkey neocortical interneurons. Nat. Neurosci. 2013, 16, 1588–1597. [Google Scholar] [CrossRef]
- Gelman, D.M.; Martini, F.J.; Nóbrega-Pereira, S.; Pierani, A.; Kessaris, N.; Marín, O. The Embryonic Preoptic Area Is a Novel Source of Cortical GABAergic Interneurons. J. Neurosci. 2009, 29, 9380–9389. [Google Scholar] [CrossRef]
- Gelman, D.M.; Griveau, A.; Dehorter, N.; Teissier, A.; Varela, C.; Pla, R.; Pierani, A.; Marín, O. A Wide Diversity of Cortical GABAergic Interneurons Derives from the Embryonic Preoptic Area. J. Neurosci. 2011, 31, 16570–16580. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Cobos, I.; De La Cruz, E.; Rubenstein, J.L.; Anderson, S.A. Origins of Cortical Interneuron Subtypes. J. Neurosci. 2004, 24, 2612–2622. [Google Scholar] [CrossRef] [PubMed]
- Butt, S.J.; Fuccillo, M.; Nery, S.; Noctor, S.; Kriegstein, A.; Corbin, J.G.; Fishell, G. The Temporal and Spatial Origins of Cortical Interneurons Predict Their Physiological Subtype. Neuron 2005, 48, 591–604. [Google Scholar] [CrossRef] [PubMed]
- Wonders, C.P.; Anderson, S.A. The origin and specification of cortical interneurons. Nat. Rev. Neurosci. 2006, 7, 687–696. [Google Scholar] [CrossRef]
- Nery, S.; Fishell, G.; Corbin, J.G. The caudal ganglionic eminence is a source of distinct cortical and subcortical cell populations. Nat. Neurosci. 2002, 5, 1279–1287. [Google Scholar] [CrossRef]
- Sussel, L.; Marin, O.; Kimura, S.; Rubenstein, J.L. Loss of Nkx2.1 homeobox gene function results in a ventral to dorsal molecular respecification within the basal telencephalon: Evidence for a transformation of the pallidum into the striatum. Development 1999, 126, 3359–3370. [Google Scholar]
- Butt, S.J.; Sousa, V.H.; Fuccillo, M.V.; Hjerling-Leffler, J.; Miyoshi, G.; Kimura, S.; Fishell, G. The Requirement of Nkx2-1 in the Temporal Specification of Cortical Interneuron Subtypes. Neuron 2008, 59, 722–732. [Google Scholar] [CrossRef]
- Fogarty, M.; Richardson, W.D.; Kessaris, N. A subset of oligodendrocytes generated from radial glia in the dorsal spinal cord. Development 2005, 132, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Wonders, C.P.; Taylor, L.; Welagen, J.; Mbata, I.C.; Xiang, J.Z.; Anderson, S.A. A spatial bias for the origins of interneuron subgroups within the medial ganglionic eminence. Dev. Biol. 2008, 314, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Inan, M.; Welagen, J.; Anderson, S.A. Spatial and Temporal Bias in the Mitotic Origins of Somatostatin- and Parvalbumin-Expressing Interneuron Subgroups and the Chandelier Subtype in the Medial Ganglionic Eminence. Cereb. Cortex 2011, 22, 820–827. [Google Scholar] [CrossRef]
- Xu, Q.; Guo, L.; Moore, H.; Waclaw, R.R.; Campbell, K.; Anderson, S.A. Sonic Hedgehog Signaling Confers Ventral Telencephalic Progenitors with Distinct Cortical Interneuron Fates. Neuron 2010, 65, 328–340. [Google Scholar] [CrossRef]
- Brown, K.N.; Chen, S.; Han, Z.; Lu, C.-H.; Tan, X.; Zhang, X.-J.; Ding, L.; Lopez-Cruz, A.; Saur, D.; Anderson, S.A.; et al. Clonal Production and Organization of Inhibitory Interneurons in the Neocortex. Science 2011, 334, 480–486. [Google Scholar] [CrossRef]
- Harwell, C.C.; Fuentealba, L.C.; Gonzalez-Cerrillo, A.; Parker, P.R.; Gertz, C.C.; Mazzola, E.; Garcia, M.T.; Alvarez-Buylla, A.; Cepko, C.L.; Kriegstein, A.R. Wide Dispersion and Diversity of Clonally Related Inhibitory Interneurons. Neuron 2015, 87, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Jaglin, X.H.; Cobbs, L.V.; Bandler, R.C.; Streicher, C.; Cepko, C.L.; Hippenmeyer, S.; Fishell, G. Clonally Related Forebrain Interneurons Disperse Broadly across Both Functional Areas and Structural Boundaries. Neuron 2015, 87, 989–998. [Google Scholar] [CrossRef]
- Miyoshi, G.; Butt, S.J.B.; Takebayashi, H.; Fishell, G. Physiologically Distinct Temporal Cohorts of Cortical Interneurons Arise from Telencephalic Olig2-Expressing Precursors. J. Neurosci. 2007, 27, 7786–7798. [Google Scholar] [CrossRef]
- Miller, M.W. Cogeneration of retrogradely labeled corticocortical projection and GABA-immunoreactive local circuit neurons in cerebral cortex. Dev. Brain Res. 1985, 23, 187–192. [Google Scholar] [CrossRef]
- Valcanis, H.; Tan, S.-S. Layer Specification of Transplanted Interneurons in Developing Mouse Neocortex. J. Neurosci. 2003, 23, 5113–5122. [Google Scholar] [CrossRef]
- Benamer, N.; Vidal, M.; Angulo, M.C. The cerebral cortex is a substrate of multiple interactions between GABAergic interneurons and oligodendrocyte lineage cells. Neurosci. Lett. 2020, 715, 134615. [Google Scholar] [CrossRef]
- Rallu, M.; Corbin, J.G.; Fishell, G. Parsing the prosencephalon. Nat. Rev. Neurosci. 2002, 3, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Tsoa, R.W.; Coskun, V.; Ho, C.K.; De Vellis, J.; Sun, Y.E. Spatiotemporally different origins of NG2 progenitors produce cortical interneurons versus glia in the mammalian forebrain. Proc. Natl. Acad. Sci. USA 2014, 111, 7444–7449. [Google Scholar] [CrossRef] [PubMed]
- Kessaris, N.; Fogarty, M.J.; Iannarelli, P.; Grist, M.; Wegner, M.; Richardson, W.D. Competing waves of oligodendrocytes in the forebrain and postnatal elimination of an embryonic lineage. Nat. Neurosci. 2005, 9, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Southwell, D.G.; Paredes, M.F.; Galvao, R.P.; Jones, D.L.; Froemke, R.C.; Sebe, J.Y.; Alfaro-Cervello, C.; Tang, Y.; Verdugo, J.M.G.; Rubenstein, J.L.; et al. Intrinsically determined cell death of developing cortical interneurons. Nat. Cell Biol. 2012, 491, 109–113. [Google Scholar] [CrossRef]
- Minocha, S.; Valloton, D.; Brunet, I.; Eichmann, A.; Hornung, J.-P.; Lebrand, C. NG2 glia are required for vessel network formation during embryonic development. eLife 2015, 4, e09102. [Google Scholar] [CrossRef]
- Orduz, D.; Benamer, N.; Ortolani, D.; Coppola, E.; Vigier, L.; Pierani, A.; Angulo, M.C. Developmental cell death regulates lineage-related interneuron-oligodendroglia functional clusters and oligodendrocyte homeostasis. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Wong, M. Too Much Inhibition Leads to Excitation in Absence Epilepsy. Epilepsy Curr. 2010, 10, 131–133. [Google Scholar] [CrossRef]
- Selten, M.; Van Bokhoven, H.; Kasri, N.N. Inhibitory control of the excitatory/inhibitory balance in psychiatric disorders. F1000Research 2018, 7, 23. [Google Scholar] [CrossRef]
- Tremblay, R.; Lee, S.; Rudy, B. GABAergic interneurons in the neocortex: From cellular properties to circuits. Neuron 2016, 91, 260–292. [Google Scholar] [CrossRef]
- Gulyas, A.I.; Megías, M.; Emri, Z.; Freund, T.F.; Megías, M. Total Number and Ratio of Excitatory and Inhibitory Synapses Converging onto Single Interneurons of Different Types in the CA1 Area of the Rat Hippocampus. J. Neurosci. 1999, 19, 10082–10097. [Google Scholar] [CrossRef]
- Nörenberg, A.; Hu, H.; Vida, I.; Bartos, M.; Jonas, P. Distinct nonuniform cable properties optimize rapid and efficient activation of fast-spiking GABAergic interneurons. Proc. Natl. Acad. Sci. USA 2009, 107, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Kubota, Y.; Karube, F.; Nomura, M.; Gulledge, A.T.; Mochizuki, A.; Schertel, A.; Kawaguchi, Y. Conserved properties of dendritic trees in four cortical interneuron subtypes. Sci. Rep. 2011, 1, 89. [Google Scholar] [CrossRef] [PubMed]
- Tukker, J.J.; Lasztóczi, B.; Katona, L.; Roberts, J.D.B.; Pissadaki, E.K.; Dalezios, Y.; Márton, L.; Zhang, L.; Klausberger, T.; Somogyi, P. Distinct Dendritic Arborization and In Vivo Firing Patterns of Parvalbumin-Expressing Basket Cells in the Hippocampal Area CA. J. Neuroscience 2013, 33, 6809–6825. [Google Scholar] [CrossRef] [PubMed]
- Bezaire, M.J.; Soltesz, I. Quantitative assessment of CA1 local circuits: Knowledge base for interneuron-pyramidal cell connectivity. Hippocampus 2013, 23, 751–785. [Google Scholar] [CrossRef]
- Meeks, J.P.; Mennerick, S. Action Potential Initiation and Propagation in CA3 Pyramidal Axons. J. Neurophysiol. 2007, 97, 3460–3472. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Hieber, C.; Jonas, P.; Bischofberger, J. Action potential initiation and propagation in hippocampal mossy fibre axons. J. Physiol. 2008, 586, 1849–1857. [Google Scholar] [CrossRef]
- Hu, H.; Jonas, P. A supercritical density of Na+ channels ensures fast signaling in GABAergic interneuron axons. Nat. Neurosci. 2014, 17, 686–693. [Google Scholar] [CrossRef]
- Fricker, D.; Miles, R. EPSP Amplification and the Precision of Spike Timing in Hippocampal Neurons. Neuron 2000, 28, 559–569. [Google Scholar] [CrossRef]
- Pouille, F. Enforcement of Temporal Fidelity in Pyramidal Cells by Somatic Feed-Forward Inhibition. Science 2001, 293, 1159–1163. [Google Scholar] [CrossRef]
- Buzsàki, G.; Eidelberg, E. Commissural projection to the dentate gyrus of the rat: Evidence for feed-forward inhibition. Brain Res. 1981, 230, 346–350. [Google Scholar] [CrossRef]
- Miles, R. Synaptic excitation of inhibitory cells by single CA3 hippocampal pyramidal cells of the guinea-pig in vitro. J. Physiol. 1990, 428, 61–77. [Google Scholar] [CrossRef]
- Pouille, F.; Scanziani, M. Routing of spike series by dynamic circuits in the hippocampus. Nat. Cell Biol. 2004, 429, 717–723. [Google Scholar] [CrossRef]
- Kassab, R.; Alexandre, F. Pattern separation in the hippocampus: Distinct circuits under different conditions. Brain Struct. Funct. 2018, 223, 2785–2808. [Google Scholar] [CrossRef]
- Senzai, Y. Function of local circuits in the hippocampal dentate gyrus-CA3 system. Neurosci. Res. 2019, 140, 43–52. [Google Scholar] [CrossRef]
- Pernía-Andrade, A.J.; Jonas, P. Theta-Gamma-Modulated Synaptic Currents in Hippocampal Granule Cells In Vivo Define a Mechanism for Network Oscillations. Neuron 2014, 81, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Riedemann, T. Diversity and Function of Somatostatin-Expressing Interneurons in the Cerebral Cortex. Int. J. Mol. Sci. 2019, 20, 2952. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Toledo-Rodriguez, M.; Gupta, A.; Wu, C.; Silberberg, G.; Luo, J.; Markram, H. Anatomical, physiological and molecular properties of Martinotti cells in the somatosensory cortex of the juvenile rat. J. Physiol. 2004, 561, 65–90. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Hu, H.; Berrebi, A.S.; Mathers, P.H.; Agmon, A. Distinct Subtypes of Somatostatin-Containing Neocortical Interneurons Revealed in Transgenic Mice. J. Neurosci. 2006, 26, 5069–5082. [Google Scholar] [CrossRef]
- McGarry, L.M.; Packer, A.M.; Fino, E.; Nikolenko, V.; Sippy, T.; Yuste, R. Quantitative classification of somatostatin-positive neocortical interneurons identifies three interneuron subtypes. Front. Neural Circuits 2010, 4, 12. [Google Scholar] [CrossRef]
- Riedemann, T.; Straub, T.; Sutor, B. Two types of somatostatin-expressing GABAergic interneurons in the superficial layers of the mouse cingulate cortex. PLoS ONE 2018, 13, e0200567. [Google Scholar] [CrossRef]
- Simonnet, J.; Nassar, M.; Stella, F.; Cohen, I.; Mathon, B.; Boccara, C.N.; Miles, R.; Fricker, D. Activity dependent feedback inhibition may maintain head direction signals in mouse presubiculum. Nat. Commun. 2017, 8, 16032. [Google Scholar] [CrossRef] [PubMed]
- Gärtner, U.; Härtig, W.; Brauer, K.; Brückner, G.; Arendt, T. Immunofluorescence and immunoelectron microscopic evidence for differences in myelination of GABAergic and cholinergic septohippocampal fibres. Int. J. Dev. Neurosci. 2001, 19, 347–352. [Google Scholar] [CrossRef]
- Caputi, A.; Melzer, S.; Michael, M.; Monyer, H. The long and short of GABAergic neurons. Curr. Opin. Neurobiol. 2013, 23, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Melzer, S.; Monyer, H. Diversity and function of corticopetal and corticofugal GABAergic projection neurons. Nat. Rev. Neurosci. 2020, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hormuzdi, S.G.; Pais, I.; Lebeau, F.E.; Towers, S.K.; Rozov, A.; Buhl, E.H.; Whittington, M.A.; Monyer, H. Impaired Electrical Signaling Disrupts Gamma Frequency Oscillations in Connexin 36-Deficient Mice. Neuron 2001, 31, 487–495. [Google Scholar] [CrossRef]
- Traub, R.D.; Kopell, N.; Bibbig, A.; Buhl, E.H.; Lebeau, F.E.N.; Whittington, M.A. Gap Junctions between Interneuron Dendrites Can Enhance Synchrony of Gamma Oscillations in Distributed Networks. J. Neurosci. 2001, 21, 9478–9486. [Google Scholar] [CrossRef]
- Cardin, J.A.; Carlén, M.; Meletis, K.; Knoblich, U.; Zhang, F.; Deisseroth, K.; Tsai, L.-H.; Moore, C.I. Driving Fast-Spiking Cells Induces Gamma Rhythm and Controls Sensory Responses. Nature 2009, 459, 663–667. [Google Scholar] [CrossRef]
- Sohal, V.S.; Zhang, F.; Yizhar, O.; Deisseroth, K. Parvalbumin neurons and gamma rhythms enhance cortical circuit performance. Nat. Cell Biol. 2009, 459, 698–702. [Google Scholar] [CrossRef]
- Antonoudiou, P.; Tan, Y.L.; Kontou, G.; Upton, A.L.; Mann, E.O. Parvalbumin and Somatostatin Interneurons Contribute to the Generation of Hippocampal Gamma Oscillations. J. Neurosci. 2020, 40, 7668–7687. [Google Scholar] [CrossRef]
- Howard, M.W.; Rizzuto, D.S.; Caplan, J.B.; Madsen, J.R.; Lisman, J.; Aschenbrenner-Scheibe, R.; Schulze-Bonhage, A.; Kahana, M.J. Gamma Oscillations Correlate with Working Memory Load in Humans. Cereb. Cortex 2003, 13, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Senkowski, D.; Gallinat, J. Dysfunctional Prefrontal Gamma-Band Oscillations Reflect Working Memory and Other Cognitive Deficits in Schizophrenia. Biol. Psychiatry 2015, 77, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hjerling-Leffler, J.; Zagha, E.; Fishell, G.; Rudy, B. The Largest Group of Superficial Neocortical GABAergic Interneurons Expresses Ionotropic Serotonin Receptors. J. Neurosci. 2010, 30, 16796–16808. [Google Scholar] [CrossRef] [PubMed]
- Tyan, L.; Chamberland, S.; Magnin, E.; Camiré, O.; Francavilla, R.; David, L.S.; Deisseroth, K.; Topolnik, L. Dendritic Inhibition Provided by Interneuron-Specific Cells Controls the Firing Rate and Timing of the Hippocampal Feedback Inhibitory Circuitry. J. Neurosci. 2014, 34, 4534–4547. [Google Scholar] [CrossRef]
- Jiang, X.; Shen, S.; Cadwell, C.R.; Berens, P.; Sinz, F.; Ecker, A.S.; Patel, S.C.; Tolias, A.S. Principles of connectivity among morphologically defined cell types in adult neocortex. Science 2015, 350, aac9462. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Rickmann, M.; Hafner, G.; Staiger, J.F. Subcellular Targeting of VIP Boutons in Mouse Barrel Cortex is Layer-Dependent and not Restricted to Interneurons. Cereb. Cortex 2017, 27, 5353–5368. [Google Scholar] [CrossRef]
- Porter, J.T.; Cauli, B.; Staiger, J.F.; Lambolez, B.; Rossier, J.; Audinat, E. Properties of bipolar VIPergic interneurons and their excitation by pyramidal neurons in the rat neocortex. Eur. J. Neurosci. 1998, 10, 3617–3628. [Google Scholar] [CrossRef]
- Bergles, D.E.; Richardson, W.D. Oligodendrocyte Development and Plasticity. Cold Spring Harb. Perspect. Biol. 2016, 8, a020453. [Google Scholar] [CrossRef]
- Zuchero, J.B.; Barres, B.A. Intrinsic and extrinsic control of oligodendrocyte development. Curr. Opin. Neurobiol. 2013, 23, 914–920. [Google Scholar] [CrossRef]
- Marques, S.; Zeisel, A.; Codeluppi, S.; Van Bruggen, D.; Falcão, A.M.; Xiao, L.; Li, H.; Häring, M.; Hochgerner, H.; Romanov, R.A.; et al. Oligodendrocyte heterogeneity in the mouse juvenile and adult central nervous system. Science 2016, 352, 1326–1329. [Google Scholar] [CrossRef]
- Levine, J.M.; Reynolds, R.; Fawcett, J.W. The oligodendrocyte precursor cell in health and disease. Trends Neurosci. 2001, 24, 39–47. [Google Scholar] [CrossRef]
- Dimou, L.; Götz, M. Glial Cells as Progenitors and Stem Cells: New Roles in the Healthy and Diseased Brain. Physiol. Rev. 2014, 94, 709–737. [Google Scholar] [CrossRef]
- Hughes, E.G.; Kang, S.H.; Fukaya, M.; Bergles, D.E. Oligodendrocyte progenitors balance growth with self-repulsion to achieve homeostasis in the adult brain. Nat. Neurosci. 2013, 16, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.R.L. NG2-expressing glial progenitor cells: An abundant and widespread population of cycling cells in the adult rat CNS. Mol. Cell. Neurosci. 2003, 24, 476–488. [Google Scholar] [CrossRef]
- Nishiyama, A.; Lin, X.H.; Giese, N.; Heldin, C.H.; Stallcup, W.B. Co-Localization of NG2 Proteoglycan and PDGF Alpha-Receptor on O2A Progenitor Cells in the Developing Rat Brain. J. Neurosci. Res. 1996, 43, 299–314. [Google Scholar] [CrossRef]
- Bansal, R.; Pfeiffer, S.E. Reversible inhibition of oligodendrocyte progenitor differentiation by a monoclonal antibody against surface galactolipids. Proc. Natl. Acad. Sci. USA 1989, 86, 6181–6185. [Google Scholar] [CrossRef]
- Hardy, R.J.; Friedrich, J.V.L. Progressive Remodeling of the Oligodendrocyte Process Arbor during Myelinogenesis. Dev. Neurosci. 1996, 18, 243–254. [Google Scholar] [CrossRef]
- Baumann, N.; Pham-Dinh, D. Biology of Oligodendrocyte and Myelin in the Mammalian Central Nervous System. Physiol. Rev. 2001, 81, 871–927. [Google Scholar] [CrossRef]
- Solly, S.K.; Thomas, J.L.; Monge, M.; Demerens, C.; Lubetzki, C.; Gardinier, M.V.; Matthieu, J.M.; Zalc, B. Myelin/Oligodendrocyte Glycoprotein (MOG) Expression Is Associated with Myelin Deposition. Glia 1996, 18, 39–48. [Google Scholar] [CrossRef]
- Pesheva, P.; Spiess, E.; Schachner, M. J1-160 and J1-180 are oligodendrocyte-secreted nonpermissive substrates for cell adhesion. J. Cell Biol. 1989, 109, 1765–1778. [Google Scholar] [CrossRef]
- Bartsch, U.; Bartsch, S.; Dörries, U.; Schachner, M. Immunohistological Localization of Tenascin in the Developing and Lesioned Adult Mouse Optic Nerve. Eur. J. Neurosci. 1992, 4, 338–352. [Google Scholar] [CrossRef]
- Dubessy, A.; Mazuir, E.; Rappeneau, Q.; Ou, S.; Ghanem, C.A.; Piquand, K.; Aigrot, M.; Thétiot, M.; Desmazières, A.; Chan, E.; et al. Role of a Contactin multi-molecular complex secreted by oligodendrocytes in nodal protein clustering in the CNS. Glia 2019, 67, 2248–2263. [Google Scholar] [CrossRef]
- Bekku, Y.; Rauch, U.; Ninomiya, Y.; Oohashi, T. Brevican distinctively assembles extracellular components at the large diameter nodes of Ranvier in the CNS. J. Neurochem. 2009, 108, 1266–1276. [Google Scholar] [CrossRef]
- Asher, R.A.; Morgenstern, D.A.; Shearer, M.C.; Adcock, K.H.; Pesheva, P.; Fawcett, J.W. Versican Is Upregulated in CNS Injury and Is a Product of Oligodendrocyte Lineage Cells. J. Neurosci. 2002, 22, 2225–2236. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, U.; Pesheva, P.; Raff, M.; Schachner, M. Expression of janusin (J1-160/180) in the retina and optic nerve of the developing and adult mouse. Glia 1993, 9, 57–69. [Google Scholar] [CrossRef]
- Cicanic, M.; Syková, E.; Vargová, L. Bral1: “Superglue” for the extracellular matrix in the brain white matter. Int. J. Biochem. Cell Biol. 2012, 44, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y. Lecticans: Organizers of the brain extracellular matrix. Cell. Mol. Life Sci. 2000, 57, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Bekku, Y.; Vargová, L.; Goto, Y.; Vorisek, I.; Dmytrenko, L.; Narasaki, M.; Ohtsuka, A.; Fässler, R.; Ninomiya, Y.; Syková, E.; et al. Bral1: Its Role in Diffusion Barrier Formation and Conduction Velocity in the CNS. J. Neurosci. 2010, 30, 3113–3123. [Google Scholar] [CrossRef]
- Dityatev, A.; Schachner, M. Extracellular matrix molecules and synaptic plasticity. Nat. Rev. Neurosci. 2003, 4, 456–468. [Google Scholar] [CrossRef]
- Fawcett, J.W.; Oohashi, T.; Pizzorusso, T. The roles of perineuronal nets and the perinodal extracellular matrix in neuronal function. Nat. Rev. Neurosci. 2019, 20, 451–465. [Google Scholar] [CrossRef]
- Frischknecht, R.; Chang, K.-J.; Rasband, M.N.; Seidenbecher, C.I. Neural ECM Molecules in Axonal and Synaptic Homeostatic Plasticity; Elsevier: Amsterdam, The Netherlands, 2014; Volume 214, pp. 81–100. [Google Scholar]
- Sakry, D.; Trotter, J. The role of the NG2 proteoglycan in OPC and CNS network function. Brain Res. 2016, 1638, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Habermacher, C.; Angulo, M.C.; Benamer, N. Glutamate versus GABA in neuron–oligodendroglia communication. Glia 2019, 67, 2092–2106. [Google Scholar] [CrossRef] [PubMed]
- Sherman, D.L.; Brophy, P.J. Mechanisms of axon ensheathment and myelin growth. Nat. Rev. Neurosci. 2005, 6, 683–690. [Google Scholar] [CrossRef]
- Stadelmann, C.; Timmler, S.; Barrantes-Freer, A.; Simons, M. Myelin in the Central Nervous System: Structure, Function, and Pathology. Physiol. Rev. 2019, 99, 1381–1431. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, C.; Yamasaki, M.; Uchigashima, M.; Konno, K.; Yanagawa, Y.; Watanabe, M. Cytochemical and cytological properties of perineuronal oligodendrocytes in the mouse cortex. Eur. J. Neurosci. 2010, 32, 1326–1336. [Google Scholar] [CrossRef] [PubMed]
- Battefeld, A.; Klooster, J.; Kole, M.H.P. Myelinating satellite oligodendrocytes are integrated in a glial syncytium constraining neuronal high-frequency activity. Nat. Commun. 2016, 7, 11298. [Google Scholar] [CrossRef]
- Dours-Zimmermann, M.T.; Maurer, K.; Rauch, U.; Stoffel, W.; Fässler, R.; Zimmermann, D.R. Versican V2 Assembles the Extracellular Matrix Surrounding the Nodes of Ranvier in the CNS. J. Neurosci. 2009, 29, 7731–7742. [Google Scholar] [CrossRef] [PubMed]
- Susuki, K.; Chang, K.-J.; Zollinger, D.R.; Liu, Y.; Ogawa, Y.; Eshed-Eisenbach, Y.; Dours-Zimmermann, M.T.; Oses-Prieto, J.A.; Burlingame, A.L.; Seidenbecher, C.I.; et al. Three Mechanisms Assemble Central Nervous System Nodes of Ranvier. Neuron 2013, 78, 469–482. [Google Scholar] [CrossRef]
- Weber, P.; Bartsch, U.; Rasband, M.N.; Czaniera, R.; Lang, Y.; Bluethmann, H.; Margolis, R.U.; Levinson, S.R.; Shrager, P.; Montag, D.; et al. Mice Deficient for Tenascin-R Display Alterations of the Extracellular Matrix and Decreased Axonal Conduction Velocities in the CNS. J. Neurosci. 1999, 19, 4245–4262. [Google Scholar] [CrossRef] [PubMed]
- Desmazieres, A.; Zonta, B.; Zhang, A.; Wu, L.-M.N.; Sherman, D.L.; Brophy, P.J. Differential stability of PNS and CNS nodal complexes when neuronal neurofascin is lost. J. Neurosci. 2014, 34, 5083–5088. [Google Scholar] [CrossRef]
- Freeman, S.A.; Desmazières, A.; Fricker, D.; Lubetzki, C.; Sol-Foulon, N. Mechanisms of sodium channel clustering and its influence on axonal impulse conduction. Cell. Mol. Life Sci. 2016, 73, 723–735. [Google Scholar] [CrossRef]
- Hildebrand, C. Ultrastructural and light-microscopic studies of the nodal region in large myelinated fibres of the adult feline spinal cord white matter. Acta Physiol. Scand. 1971, 82, 43–79. [Google Scholar] [CrossRef]
- Waxman, S.G.; Swadlow, H.A. Ultrastructure of visual callosal axons in the rabbit. Exp. Neurol. 1976, 53, 115–127. [Google Scholar] [CrossRef]
- Waxman, S.G.; Black, J.A. Freeze-fracture ultrastructure of the perinodal astrocyte and associated glial junctions. Brain Res. 1984, 308, 77–87. [Google Scholar] [CrossRef]
- Ffrench-Constant, C.; Miller, R.H.; Kruse, J.; Schachner, M.; Raff, M.C. Molecular specialization of astrocyte processes at nodes of Ranvier in rat optic nerve. J. Cell Biol. 1986, 102, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Serwanski, D.R.; Jukkola, P.; Nishiyama, A. Heterogeneity of astrocyte and NG2 cell insertion at the node of ranvier. J. Comp. Neurol. 2016, 525, 535–552. [Google Scholar] [CrossRef]
- Ronzano, R.; Roux, T.; Thetiot, M.; Aigrot, M.S.; Richard, L.; Lejeune, F.X.; Mazuir, E.; Vallat, J.M.; Lubetzki, C.; Desmazières, A. Microglia-Neuron Communication at Nodes of Ranvier Depends on Neuronal Activity through Potassium Release and Contributes to Myelin Repair. bioRxiv 2020. [Google Scholar] [CrossRef]
- Pizzorusso, T.; Medini, P.; Berardi, N.; Chierzi, S.; Fawcett, J.W.; Maffei, L. Reactivation of Ocular Dominance Plasticity in the Adult Visual Cortex. Science 2002, 298, 1248–1251. [Google Scholar] [CrossRef]
- Berardi, N.; Pizzorusso, T.; Maffei, L. Extracellular Matrix and Visual Cortical Plasticity. Neuron 2004, 44, 905–908. [Google Scholar] [CrossRef]
- Hensch, T.K. Critical period plasticity in local cortical circuits. Nat. Rev. Neurosci. 2005, 6, 877–888. [Google Scholar] [CrossRef]
- Faini, G.; Aguirre, A.; Landi, S.; Lamers, D.; Pizzorusso, T.; Ratto, G.M.; Deleuze, C.; Bacci, A. Perineuronal nets control visual input via thalamic recruitment of cortical PV interneurons. eLife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.C.; Lensjø, K.K.; Lepperød, M.E.; Dragly, S.-A.; Sutterud, H.; Blackstad, J.S.; Fyhn, M.; Hafting, T. Perineuronal nets stabilize the grid cell network. Nat. Commun. 2021, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Franke, A.; Kaplan, M.R.; Pfieger, F.W.; Barres, B.A. Characterization of the signaling interactions that promote the survival and growth of developing retinal ganglion cells in culture. Neuron 1995, 15, 805–819. [Google Scholar] [CrossRef]
- Pfrieger, F.W. Synaptic Efficacy Enhanced by Glial Cells in Vitro. Science 1997, 277, 1684–1687. [Google Scholar] [CrossRef] [PubMed]
- Nägler, K.; Mauch, D.H.; Pfrieger, F.W. Glia-derived signals induce synapse formation in neurones of the rat central nervous system. J. Physiol. 2001, 533, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Ullian, E.M.; Sapperstein, S.K.; Christopherson, K.S.; Barres, B.A. Control of Synapse Number by Glia. Science 2001, 291, 657–661. [Google Scholar] [CrossRef]
- Bergles, D.E.; Roberts, J.D.B.; Somogyi, P.; Jahr, C.E. Glutamatergic synapses on oligodendrocyte precursor cells in the hippocampus. Nat. Cell Biol. 2000, 405, 187–191. [Google Scholar] [CrossRef]
- Lin, S.-C.; Bergles, D.E. Synaptic signaling between GABAergic interneurons and oligodendrocyte precursor cells in the hippocampus. Nat. Neurosci. 2003, 7, 24–32. [Google Scholar] [CrossRef]
- Sakry, D.; Neitz, A.; Singh, J.; Frischknecht, R.; Marongiu, D.; Binamé, F.; Perera, S.S.; Endres, K.; Lutz, B.; Radyushkin, K.; et al. Oligodendrocyte Precursor Cells Modulate the Neuronal Network by Activity-Dependent Ectodomain Cleavage of Glial NG. PLoS Biol. 2014, 12, e1001993. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Hozumi, Y.; Kaneko, K.; Fujii, S. Modulatory Effects of Perineuronal Oligodendrocytes on Neuronal Activity in the Rat Hippocampus. Neurochem. Res. 2017, 43, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Gould, E.; Xu, J.; Kim, E.J.; Kim, J.H. Oligodendrocytes regulate presynaptic properties and neurotransmission through BDNF signaling in the mouse brainstem. eLife 2019, 8, e42156. [Google Scholar] [CrossRef]
- Frühbeis, C.; Fröhlich, D.; Kuo, W.P.; Amphornrat, J.; Thilemann, S.; Saab, A.S.; Kirchhoff, F.; Möbius, W.; Goebbels, S.; Nave, K.-A.; et al. Neurotransmitter-Triggered Transfer of Exosomes Mediates Oligodendrocyte–Neuron Communication. PLoS Biol. 2013, 11, e1001604. [Google Scholar] [CrossRef]
- Du, Y.; Dreyfus, C.F. Oligodendrocytes as providers of growth factors. J. Neurosci. Res. 2002, 68, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, I.; Hassinger, L.; Paskevich, P.A.; Shine, H.D.; Nixon, R.A. Oligodendroglia Regulate the Regional Expansion of Axon Caliber and Local Accumulation of Neurofilaments during Development Independently of Myelin Formation. J. Neurosci. 1996, 16, 5095–5105. [Google Scholar] [CrossRef]
- Birey, F.; Kloc, M.; Chavali, M.; Hussein, I.; Wilson, M.; Christoffel, D.J.; Chen, T.; Frohman, M.A.; Robinson, J.K.; Russo, S.J.; et al. Genetic and Stress-Induced Loss of NG2 Glia Triggers Emergence of Depressive-like Behaviors through Reduced Secretion of FGF. Neuron 2015, 88, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Xin, W.; Mironova, Y.A.; Shen, H.; Marino, R.A.; Waisman, A.; Lamers, W.H.; Bergles, D.E.; Bonci, A. Oligodendrocytes Support Neuronal Glutamatergic Transmission via Expression of Glutamine Synthetase. Cell Rep. 2019, 27, 2262–2271. [Google Scholar] [CrossRef] [PubMed]
- Berghuis, P.; Dobszay, M.B.; Sousa, K.M.; Schulte, G.; Mager, P.P.; Härtig, W.; Görcs, T.J.; Zilberter, Y.; Ernfors, P.; Harkany, T. Brain-derived neurotrophic factor controls functional differentiation and microcircuit formation of selectively isolated fast-spiking GABAergic interneurons. Eur. J. Neurosci. 2004, 20, 1290–1306. [Google Scholar] [CrossRef] [PubMed]
- Turko, P.; Groberman, K.; Kaiser, T.; Yanagawa, Y.; Vida, I. Primary Cell Culture of Purified GABAergic or Glutamatergic Neurons Established through Fluorescence-activated Cell Sorting. J. Vis. Exp. 2019, e58974. [Google Scholar] [CrossRef]
- Uematsu, M.; Hirai, Y.; Karube, F.; Ebihara, S.; Kato, M.; Abe, K.; Obata, K.; Yoshida, S.; Hirabayashi, M.; Yanagawa, Y.; et al. Quantitative Chemical Composition of Cortical GABAergic Neurons Revealed in Transgenic Venus-Expressing Rats. Cereb. Cortex 2008, 18, 315–330. [Google Scholar] [CrossRef]
- Turko, P.; Groberman, K.; Browa, F.; Cobb, S.; Vida, I. Differential Dependence of GABAergic and Glutamatergic Neurons on Glia for the Establishment of Synaptic Transmission. Cereb. Cortex 2019, 29, 1230–1243. [Google Scholar] [CrossRef]
- Mazuir, E.; Richevaux, L.; Nassar, M.; Robil, N.; de la Grange, P.; Lubetzki, C.; Fricker, D.; Sol-Foulon, N. Oligodendrocyte Secreted Factors Shape Hippocampal GABAergic Neuron Transcriptome and Physiology. bioRxiv 2020. [Google Scholar] [CrossRef]
- Mazuir, E.; Dubessy, A.-L.; Wallon, L.; Aigrot, M.-S.; Lubetzki, C.; Sol-Foulon, N. Generation of Oligodendrocytes and Oligodendrocyte-Conditioned Medium for Co-Culture Experiments. J. Vis. Exp. 2020, 156, e60912. [Google Scholar] [CrossRef] [PubMed]
- Le Magueresse, C.; Monyer, H. GABAergic Interneurons Shape the Functional Maturation of the Cortex. Neuron 2013, 77, 388–405. [Google Scholar] [CrossRef]
- Tomassy, G.S.; Berger, D.R.; Chen, H.-H.; Kasthuri, N.; Hayworth, K.J.; Vercelli, A.; Seung, H.S.; Lichtman, J.W.; Arlotta, P. Distinct Profiles of Myelin Distribution Along Single Axons of Pyramidal Neurons in the Neocortex. Science 2014, 344, 319–324. [Google Scholar] [CrossRef]
- Somogyi, P.; Kisvárday, Z.; Martin, K.; Whitteridge, D. Synaptic connections of morphologically identified and physiologically characterized large basket cells in the striate cortex of cat. Neuroscience 1983, 10, 261–294. [Google Scholar] [CrossRef]
- Somogyi, P.; Freund, T.; Wu, J.-Y.; Smith, A.; Smith, D. The section-Golgi impregnation procedure. Immunocytochemical demonstration of glutamate decarboxylase in Golgi-impregnated neurons and in their afferent synaptic boutons in the visual cortex of the cat. Neuroscience 1983, 9, 475–490. [Google Scholar] [CrossRef]
- Stedehouder, J.; Brizee, D.; Shpak, G.; Kushner, S.A. Activity-Dependent Myelination of Parvalbumin Interneurons Mediated by Axonal Morphological Plasticity. J. Neurosci. 2018, 38, 3631–3642. [Google Scholar] [CrossRef]
- Stedehouder, J.; Brizee, D.; Slotman, J.A.; Pascual-Garcia, M.; Leyrer, M.L.; Bouwen, B.L.; Dirven, C.M.; Gao, Z.; Berson, D.M.; Houtsmuller, A.B.; et al. Local axonal morphology guides the topography of interneuron myelination in mouse and human neocortex. eLife 2019, 8, e48615. [Google Scholar] [CrossRef]
- Zonouzi, M.; Berger, D.; Jokhi, V.; Kedaigle, A.; Lichtman, J.; Arlotta, P. Individual Oligodendrocytes Show Bias for Inhibitory Axons in the Neocortex. Cell Rep. 2019, 27, 2799–2808. [Google Scholar] [CrossRef]
- Benamer, N.; Vidal, M.; Balia, M.; Angulo, M.C. Myelination of parvalbumin interneurons shapes the function of cortical sensory inhibitory circuits. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Micheva, K.D.; Kiraly, M.; Perez, M.M.; Madison, D.V. Conduction Velocity along the Local Axons of Parvalbumin Interneurons Correlates with the Degree of Axonal Myelination. 2020. Available online: https://www.biorxiv.org/content/10.1101/2020.10.10.334656v1 (accessed on 25 November 2020).
- Schmidt, H.; Gour, A.; Straehle, J.; Boergens, K.M.; Brecht, M.; Helmstaedter, M. Axonal synapse sorting in medial entorhinal cortex. Nat. Cell Biol. 2017, 549, 469–475. [Google Scholar] [CrossRef]
- De Felipe, J.; Alonso-Nanclares, L.; Arellano, J.I. Microstructure of the neocortex: Comparative aspects. J. Neurocytol. 2002, 31, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Leach, M.K.; Redmond, S.A.; Chong, S.C.; Mellon, S.H.; Tuck, S.J.; Feng, Z.-Q.; Corey, J.M.; Chan, J.R. A culture system to study oligodendrocyte myelination processes using engineered nanofibers. Nat. Methods 2012, 9, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Mayoral, S.R.; Etxeberria, A.; Shen, Y.-A.A.; Chan, J.R. Initiation of CNS Myelination in the Optic Nerve Is Dependent on Axon Caliber. Cell Rep. 2018, 25, 544–550.e3. [Google Scholar] [CrossRef] [PubMed]
- Bechler, M.E.; Byrne, L.; Ffrench-Constant, C. CNS Myelin Sheath Lengths Are an Intrinsic Property of Oligodendrocytes. Curr. Biol. 2015, 25, 2411–2416. [Google Scholar] [CrossRef]
- Goebbels, S.; Wieser, G.L.; Pieper, A.; Spitzer, S.; Weege, B.; Yan, K.; Edgar, J.M.; Yagensky, O.; Wichert, S.P.; Agarwal, A.; et al. A neuronal PI(3,4,5)P3-dependent program of oligodendrocyte precursor recruitment and myelination. Nat. Neurosci. 2016, 20, 10–15. [Google Scholar] [CrossRef]
- Yang, S.M.; Michel, K.; Jokhi, V.; Nedivi, E.; Arlotta, P. Neuron class-specific responses govern adaptive myelin remodeling in the neocortex. Science 2020, 370, 6523. [Google Scholar] [CrossRef]
- Lubetzki, C.; Sol-Foulon, N.; Desmazières, A. Nodes of Ranvier during development and repair in the CNS. Nat. Rev. Neurol. 2020, 16, 426–439. [Google Scholar] [CrossRef] [PubMed]
- Rasband, M.N.; Peles, E. Mechanisms of node of Ranvier assembly. Nat. Rev. Neurosci. 2021, 22, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.R.; Meyer-Franke, A.; Lambert, S.; Bennett, V.; Duncan, I.D.; Levinson, S.R.; Barres, B.A. Induction of sodium channel clustering by oligodendrocytes. Nat. Cell Biol. 1997, 386, 724–728. [Google Scholar] [CrossRef]
- Kaplan, M.R.; Cho, M.H.; Ullian, E.M.; Isom, L.L.; Levinson, S.R.; Barres, B.A. Differential Control of Clustering of the Sodium Channels Na(v)1.2 and Na(v)1.6 at Developing CNS Nodes of Ranvier. Neuron 2001, 30, 105–119. [Google Scholar] [CrossRef]
- Freeman, S.A.; Desmazières, A.; Simonnet, J.; Gatta, M.; Pfeiffer, F.; Aigrot, M.S.; Rappeneau, Q.; Guerreiro, S.; Michel, P.P.; Yanagawa, Y.; et al. Acceleration of conduction velocity linked to clustering of nodal components precedes myelination. Proc. Natl. Acad. Sci. USA 2015, 112, E321–E328. [Google Scholar] [CrossRef]
- Thetiot, M.; Freeman, S.A.; Roux, T.; Dubessy, A.; Aigrot, M.; Rappeneau, Q.; Lejeune, F.; Tailleur, J.; Sol-Foulon, N.; Lubetzki, C.; et al. An alternative mechanism of early nodal clustering and myelination onset in GABAergic neurons of the central nervous system. Glia 2020, 68, 1891–1909. [Google Scholar] [CrossRef] [PubMed]
- Debanne, D.; Campanac, E.; Bialowas, A.; Carlier, E.; Alcaraz, G. Axon Physiology. Physiol. Rev. 2011, 91, 555–602. [Google Scholar] [CrossRef] [PubMed]
- Huxley, A.F.; Stämpeli, R. Evidence for saltatory conduction in peripheral myelinated nerve fibres. J. Physiol. 1949, 108, 315–339. [Google Scholar] [CrossRef]
- Tasaki, I.; Frank, K. Measurement of the Action Potential of Myelinated Nerve Fiber. Am. J. Physiol. Content 1955, 182, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Castelfranco, A.M.; Hartline, D.K. Evolution of rapid nerve conduction. Brain Res. 2016, 1641, 11–33. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.C.; Popovic, M.A.; Klooster, J.; Weil, M.-T.; Möbius, W.; Nave, K.-A.; Kole, M.H. Saltatory Conduction along Myelinated Axons Involves a Periaxonal Nanocircuit. Cell 2020, 180, 311–322.e15. [Google Scholar] [CrossRef]
- Rushton, W.A.H. A theory of the effects of fibre size in medullated nerve. J. Physiol. 1951, 115, 101–122. [Google Scholar] [CrossRef]
- Hursh, J.B. Conduction Velocity and Diameter of Nerve Fibers. Am. J. Physiol. Content 1939, 127, 131–139. [Google Scholar] [CrossRef]
- Brill, M.H.; Waxman, S.G.; Moore, J.W.; Joyner, R.W. Conduction velocity and spike configuration in myelinated fibres: Computed dependence on internode distance. J. Neurol. Neurosurg. Psychiatry 1977, 40, 769–774. [Google Scholar] [CrossRef]
- Waxman, S.G. Integrative Properties and Design Principles of Axons. Int. Rev. Neurobiol. 1975, 18, 1–40. [Google Scholar] [CrossRef]
- Ford, M.C.; Alexandrova, O.; Cossell, L.; Stange-Marten, A.; Sinclair, J.; Kopp-Scheinpflug, C.; Pecka, M.; Attwell, D.; Grothe, B. Tuning of Ranvier node and internode properties in myelinated axons to adjust action potential timing. Nat. Commun. 2015, 6, 8073. [Google Scholar] [CrossRef] [PubMed]
- Arancibia-Cárcamo, I.L.; Ford, M.C.; Cossell, L.; Ishida, K.; Tohyama, K.; Attwell, D. Node of Ranvier length as a potential regulator of myelinated axon conduction speed. eLife 2017, 6, e23329. [Google Scholar] [CrossRef]
- Cullen, C.L.; Pepper, R.E.; Clutterbuck, M.T.; Pitman, K.A.; Oorschot, V.; Auderset, L.; Tang, A.D.; Ramm, G.; Emery, B.; Rodger, J.; et al. Periaxonal and nodal plasticities modulate action potential conduction in the adult mouse brain. Cell Rep. 2021, 34, 108641. [Google Scholar] [CrossRef] [PubMed]
- Bonetto, G.; Hivert, B.; Goutebroze, L.; Karagogeos, D.; Crépel, V.; Faivre-Sarrailh, C. Selective Axonal Expression of the Kv1 Channel Complex in Pre-myelinated GABAergic Hippocampal Neurons. Front. Cell. Neurosci. 2019, 13, 222. [Google Scholar] [CrossRef] [PubMed]
- Chiu, S.Y.; Ritchie, J.M. On the physiological role of internodal potassium channels and the security of conduction in myelinated nerve fibres. Proc. R. Soc. London. Ser. B Biol. Sci. 1984, 220, 415–422. [Google Scholar] [CrossRef]
- Vabnick, I.; Trimmer, J.S.; Schwarz, T.L.; Levinson, S.R.; Risal, D.; Shrager, P. Dynamic Potassium Channel Distributions during Axonal Development Prevent Aberrant Firing Patterns. J. Neurosci. 1999, 19, 747–758. [Google Scholar] [CrossRef]
- Jonas, P.; Bischofberger, J.; Fricker, D.; Miles, R. Interneuron Diversity series: Fast in, fast out – temporal and spatial signal processing in hippocampal interneurons. Trends Neurosci. 2004, 27, 30–40. [Google Scholar] [CrossRef]
- Hu, H.; Roth, F.C.; Vandael, D.; Jonas, P. Complementary Tuning of Na+ and K+ Channel Gating Underlies Fast and Energy-Efficient Action Potentials in GABAergic Interneuron Axons. Neuron 2018, 98, 156–165.e6. [Google Scholar] [CrossRef]
- Brohawn, S.G.; Wang, W.; Handler, A.; Campbell, E.B.; Schwarz, J.R.; MacKinnon, R. The mechanosensitive ion channel TRAAK is localized to the mammalian node of Ranvier. eLife 2019, 8, e50403. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Ling, J.; Tonomura, S.; Noguchi, K.; Matalon, S.; Gu, J.G. TREK-1 and TRAAK Are Principal K+ Channels at the Nodes of Ranvier for Rapid Action Potential Conduction on Mammalian Myelinated Afferent Nerves. Neuron 2019, 104, 960–971.e7. [Google Scholar] [CrossRef]
- Devaux, J.J.; Kleopa, K.A.; Cooper, E.C.; Scherer, S.S. KCNQ2 Is a Nodal K+ Channel. J. Neurosci. 2004, 24, 1236–1244. [Google Scholar] [CrossRef]
- Okaty, B.W.; Miller, M.N.; Sugino, K.; Hempel, C.M.; Nelson, S.B. Transcriptional and Electrophysiological Maturation of Neocortical Fast-Spiking GABAergic Interneurons. J. Neurosci. 2009, 29, 7040–7052. [Google Scholar] [CrossRef] [PubMed]
- Rosenbluth, J. Central myelin in the mouse mutant shiverer. J. Comp. Neurol. 1980, 194, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Fields, R.D.; Bukalo, O. Myelin makes memories. Nat. Neurosci. 2020, 23, 469–470. [Google Scholar] [CrossRef] [PubMed]
- Carr, C.E.; Soares, D.; Parameshwaran, S.; Perney, T. Evolution and development of time coding systems. Curr. Opin. Neurobiol. 2001, 11, 727–733. [Google Scholar] [CrossRef]
- Seidl, A.H.; Rubel, E.W.; Harris, D.M. Mechanisms for Adjusting Interaural Time Differences to Achieve Binaural Coincidence Detection. J. Neurosci. 2010, 30, 70–80. [Google Scholar] [CrossRef]
- Seidl, A.H.; Rubel, E.W. Systematic and differential myelination of axon collaterals in the mammalian auditory brainstem. Glia 2016, 64, 487–494. [Google Scholar] [CrossRef]
- Stange-Marten, A.; Nabel, A.L.; Sinclair, J.L.; Fischl, M.; Alexandrova, O.; Wohlfrom, H.; Kopp-Scheinpflug, C.; Pecka, M.; Grothe, B. Input timing for spatial processing is precisely tuned via constant synaptic delays and myelination patterns in the auditory brainstem. Proc. Natl. Acad. Sci. USA 2017, 114, E4851–E4858. [Google Scholar] [CrossRef]
- Borst, J.G.G.; Van Hoeve, J.S. The Calyx of Held Synapse: From Model Synapse to Auditory Relay. Annu. Rev. Physiol. 2012, 74, 199–224. [Google Scholar] [CrossRef]
- Kim, J.H.; Renden, R.; Von Gersdorff, H. Dysmyelination of Auditory Afferent Axons Increases the Jitter of Action Potential Timing during High-Frequency Firing. J. Neurosci. 2013, 33, 9402–9407. [Google Scholar] [CrossRef]
- Kim, S.E.; Turkington, K.; Kushmerick, C.; Kim, J.H. Central dysmyelination reduces the temporal fidelity of synaptic transmission and the reliability of postsynaptic firing during high-frequency stimulation. J. Neurophysiol. 2013, 110, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Meschkat, M.; Ruhwedel, T.; Trevisiol, A.; Tzvetanova, I.D.; Battefeld, A.; Kusch, K.; Kole, M.H.P.; Strenzke, N.; Möbius, W.; et al. A role of oligodendrocytes in information processing. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Bechler, M.E.; Swire, M.; Ffrench-Constant, C. Intrinsic and adaptive myelination-A sequential mechanism for smart wiring in the brain. Dev. Neurobiol. 2017, 78, 68–79. [Google Scholar] [CrossRef]
- Ronzano, R.; Thetiot, M.; Lubetzki, C.; Desmazieres, A. Myelin Plasticity and Repair: Neuro-Glial Choir Sets the Tuning. Front. Cell. Neurosci. 2020, 14, 42. [Google Scholar] [CrossRef] [PubMed]
- Xin, W.; Chan, J.R. Myelin plasticity: Sculpting circuits in learning and memory. Nat. Rev. Neurosci. 2020, 21, 682–694. [Google Scholar] [CrossRef]
- Hill, R.A.; Li, A.M.; Grutzendler, J. Lifelong cortical myelin plasticity and age-related degeneration in the live mammalian brain. Nat. Neurosci. 2018, 21, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Auer, F.; Vagionitis, S.; Czopka, T. Evidence for Myelin Sheath Remodeling in the CNS Revealed by In Vivo Imaging. Curr. Biol. 2018, 28, 549–559.e3. [Google Scholar] [CrossRef]
- Stedehouder, J.; Kushner, S.A. Myelination of parvalbumin interneurons: A parsimonious locus of pathophysiological convergence in schizophrenia. Mol. Psychiatry 2017, 22, 4–12. [Google Scholar] [CrossRef]
- Maas, D.A.; Eijsink, V.D.; Spoelder, M.; Van Hulten, J.A.; De Weerd, P.; Homberg, J.R.; Vallès, A.; Nait-Oumesmar, B.; Martens, G.J.M. Interneuron hypomyelination is associated with cognitive inflexibility in a rat model of schizophrenia. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zoupi, L.; Booker, S.A.; Eigel, D.; Werner, C.; Kind, P.C.; Spires-Jones, T.L.; Newland, B.; Williams, A.C. Selective vulnerability of inhibitory networks in multiple sclerosis. Acta Neuropathol. 2021, 141, 415–429. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazuir, E.; Fricker, D.; Sol-Foulon, N. Neuron–Oligodendrocyte Communication in Myelination of Cortical GABAergic Cells. Life 2021, 11, 216. https://doi.org/10.3390/life11030216
Mazuir E, Fricker D, Sol-Foulon N. Neuron–Oligodendrocyte Communication in Myelination of Cortical GABAergic Cells. Life. 2021; 11(3):216. https://doi.org/10.3390/life11030216
Chicago/Turabian StyleMazuir, Elisa, Desdemona Fricker, and Nathalie Sol-Foulon. 2021. "Neuron–Oligodendrocyte Communication in Myelination of Cortical GABAergic Cells" Life 11, no. 3: 216. https://doi.org/10.3390/life11030216
APA StyleMazuir, E., Fricker, D., & Sol-Foulon, N. (2021). Neuron–Oligodendrocyte Communication in Myelination of Cortical GABAergic Cells. Life, 11(3), 216. https://doi.org/10.3390/life11030216