Abstract
Plants release a variety of volatiles and herbivore-induced plant volatiles (HIPVs) after being damaged by herbivorous insects, which play multiple roles in the interactions with other plants and insects. Agasicles hygrophila Selman and Vogt (Coleoptera: Chrysomelidae) is a monophagous natural enemy and an effective biocontrol agent for Alternanthera philoxeroides (Mart.) Griseb. Here, we reported differences among the volatiles of A. philoxeroides by solid phase microextraction (SPME) using a gas chromatography-mass spectrometer (GC-MS). We compared the volatile emission of: (1) clean plants (CK); (2) A. philoxeroides plants with mechanical damage treatment (MD); and (3) A. philoxeroides plants infested with A. hygrophila 1st, 2nd, and 3rd larvae and female and male adults. A total of 97 volatiles were recorded, of which 5 occurred consistently in all treatments, while 61 volatiles were only observed in A. philoxeroides infested by A. hygrophila, such as trans-nerolidol, (E)-β-farnesene, and (3E,7E)-4,8,12-trimethyltrideca-1,3,7,11-tetraene (E, E-TMTT), etc. Among the 97 volatile compounds, 37 compounds belong to alkenes, 29 compounds belong to alkanes, and there were 8 esters, 8 alcohols and 6 ketones. Orthogonal partial least squares-discrimination analysis (OPLS-DA) showed that the different treatments were separated from each other, especially insect feeding from CK and MD treatments, and 19 volatiles contributed most to the separation among the treatments, with variable importance for the projection (VIP) values > 1. Our findings indicated that the alligatorweed plants could be induced to release volatiles by different stages of A. hygrophila, and the volatile compounds released differ quantitatively and qualitatively. The results from this study laid an important foundation for using volatile organic compounds (VOCs) and HIPVs of alligatorweed to improve the control effect of A. hygrophila on A. philoxeroides.
1. Introduction
Volatile organic compounds (VOCs) are the most important pathway through which plants emit signals to the environment [1]. VOCs play an important role in the vegetative relationship at plant-herbivorous insect-natural enemy [2,3,4]. In the long-term co-evolution of plants and herbivorous insects, plants gradually form an inductive defense response when they are threatened by herbivorous insects; that is, when plants are attacked by herbivorous insects, pest-induced VOCs will be released, which are called herbivore-induced plant volatiles (HIPVs) [5,6,7]. HIPV compounds can protect against herbivores directly or indirectly by enhancing the defense response, regulating insect behavior, and sending “early warning signals” to neighboring plants [8,9,10,11]. In addition, when plants are threatened by herbivorous insects, a volatile chemical signal, synomone, can be released to lure the natural enemies of herbivorous insects for indirect defense [12,13].
The sources of volatiles include mechanical damage caused by feeding insects and the induction of chemical attractants in insects. The types of plant volatiles induced by insect pests mainly include green leaf volatiles (GLVs), terpenoids, nitrogen-containing compounds, aldehydes, alcohols, ketones, ethers, and carboxylic acids [14,15]. The volatiles directly affects host selection, oviposition and courtship behavior in herbivores [16], while these behaviors of natural enemies would also affect their control effects on weeds. However, the current research on plant volatiles induced by pests mainly focuses on the relationship among crops, herbivorous pests and natural enemies of pests [17,18,19], and there have been few reports on VOC and HIPV components in invasive weeds affected by feeding of natural enemies for biological control.
Alternanthera philoxeroides (Mart.) Griseb is an important invasive weed native to South America [20]. Agasicles hygrophila Selman and Vogt (Coleoptera: Chrysomelidae) is a monophagous natural enemy and an effective biocontrol agent for A. philoxeroides. In the 1960s, A. hygrophila was first released to control A. philoxeroides in the southeastern United States, which was regarded as the first successful aquatic weed biological control program in the world [21,22]. In 1986, A. hygrophila was introduced into China, and 39 plants from 21 families were tested to confirm the specialism of A. hygrophila [23]. Specialists (i.e., monophagous and oligophagous insects) subsist on one or a few plants from the same family [24]. Studies on host specialization and adaptation have revealed that plant volatiles play important roles in host selection processes in herbivorous insects [25,26,27,28]. Li et al. demonstrated that the specialist beetle A. hygrophila adults used two common plant volatiles, (E)-4,8-dimethyl-1,3,7-nonatriene and (Z)-3-hexenol, for host discrimination [26]. However, little is known about A. philoxeroides responses to the feeding of A. hygrophila in different stages. Recognizing VOCs from A. philoxeroides will help to find new ways to manage this important invasive weed.
In the current study, we analyzed HIPVs emitted by undamaged and damaged A. philoxeroides (including mechanically damaged and A. hygrophila-infested) and identified the changes in emission profiles. Additionally, we also discussed the possible roles of these volatiles in plant defense, host localization, or natural enemy attraction. The results may help us to improve the control effect of A. hygrophila by using the volatiles of A. philoxeroides.
2. Materials and Methods
2.1. Plant and Insects
Alternanthera philoxeroides were grown in the greenhouses of Institute of Plant Protection, Fujian Academy of Agricultural Sciences (IPP, FAAS). Agasicles hygrophila were reared in the controlled laboratory of IPP, FAAS. Here, 1st, 2nd, and 3rd instar larvae, as well as female and male adults used in the experiments, were kept in climate cabinets (26 ± 2 °C, 70 ± 5% R.H., L14: D10) in plastic cases (18 × 11.5 × 7 cm) containing A. philoxeroides plants, respectively.
2.2. Plant Treatments
To characterize the differences in plant volatiles released in response to attack by mechanical damage and A. hygrophila, we collected headspace volatiles of A. philoxeroides plants subjected to different treatments. Plants were subjected to the three following treatments: (1) clean plants (CK), i.e., without herbivory and damaged; (2) mechanically damaged plants (MD); (3) A. hygrophila-infested plants, i.e., 1st instar larvae (1L), 2nd instar larvae (2L), 3rd instar larvae (3L), female adults (Female), and male adults (Male), respectively. A. philoxeroides stems (6–8 leaves and 20–25 cm in height) were used in the experiments and were placed in separate vials (d = 2.5 cm, h = 25 cm).
Mechanically damaged plants: 24 h before the volatiles was collected, A. philoxeroides leaves were cut with carborundum, a damaged spot of about 1 × 1 cm was created on each leaf, and then they were placed in separate vials.
A. hygrophila-infested plants: After 6 h of starvation, 6 individuals of 1st instar larvae, 2nd instar larvae, 3rd instar larvae, female adults, and male adults were released into each vial for 24 h, respectively, and then the insects and their feces were removed before collecting volatiles.
Clean plants, mechanically treated plants and herbivore-infested were kept in separate controlled chambers (26 ± 2 °C, 70 ± 5% R.H., L14: D10). All the samples were prepared in triplicate.
2.3. Collection of Plant Volatiles
After 24 h of mechanical damage treatment, volatiles were collected. The above A. hygrophila-infested plant was put into a new vial and kept for 24 h, respectively. Volatiles from healthy plants, mechanically damaged plants and A. hygrophila-infested plants were collected. After 24 h, the sample in the vial was equilibrated at 40 °C for 10 min in a water bath, respectively. After equilibration, the solid phase microextraction (SPME) was exposed to the headspace of the vial for 30 min, after which the SPME was inserted into a GC-MS (Shimadzu Corporation, Kyoto, Japan) desorption for 2 min for analysis.
2.4. Analysis of Plant Volatiles by GC-MS/MS
Volatile analysis was done with reference to the method described previously with some modifications [29]. GC-MS analysis was performed on the Nexis GC-2030 (GC) coupled with a QP2020NX mass spectrometer (Shimadzu Corporation). Helium was used as the carrier gas at a flow rate of 1 mL/ min in a splitless injection and a velocity of 36.3 cm/s. Volatile compounds were separated using a Rxi-5Sil MS (30 m × 0.25 mm, 0.25 μm) under the following conditions: the starting temperature of 50 °C was held for 2 min, followed by an increase from 50 to 180 °C at a rate of 5 °C/min, and then an increase to 280 °C at a rate of 20 °C/min, where the temperature was maintained until the procedure was manually stopped (a total of 35 min). The injector temperature was 200 °C, and the interface temperature was 280 °C. The mass spectrometric detector operated in the scan mode, and the m/z range was from 35 to 550. Compound identification was performed using the data system library (NIST 17-1, NIST 17-2, NIST 17s, FFNSC 1.2).
Volatile compounds were identified initially by comparing the mass spectra of the samples with the data system library (NIST 17-1, NIST 17-2, NIST 17s, FFNSC 1.2) and the retention index (RI). The compounds with a similarity index (SI) < 80 were deleted. The peak area normalization method was used to calculate the relative contents of each volatile compound.
2.5. Statistical Analysis
Orthogonal partial least squares-discrimination analysis (OPLS-DA) of the volatile compounds was performed using soft independent modelling of class analogies (SIMCA, Version 13.0.0.0, Umetrics AB, Umea, Sweden). The variable importance in the projection (VIP) value generated from OPLS-DA is often used to quantify the contribution of each material to the classification. The larger the VIP value is, the greater the contribution of the volatile compounds to discriminate the different groups. Therefore, the VIP value is usually taken as one of the important evaluation indexes. A heat map was obtained using OriginPro 2018C (SR1 b9.5.1.195, OriginLab Corporation, Northamnton, MA, USA).
3. Results
3.1. Volatile Compounds A. philoxeroides in Leaves Infested with A. hygrophila
Ninety-seven volatile substances were identified in A. philoxeroides of different treatments (Table 1). Nineteen compounds were identified from the clean plants (CK) of A. philoxeroides, mainly (E)-4,8-dimethylnona-1,3,7-triene (DMNT), heneicosane, trans-α-bergamotene and naphthalene. There were 26 volatile compounds in mechanically treated plants (MD), mainly heneicosane, tetracosane, cis-α-bergamotene, and (+)-β-cedrene. There were 32, 32, and 50 compounds, respectively, in A. philoxeroides infested by 1st, 2nd, and 3rd larvae A. hygrophila, mainly (E)-4,8-dimethylnona-1,3,7-triene, 2-ethenyl-1,1-dimethyl-3-methylene-cyclohexane and (E)-β-farnesene. The number of volatile compounds detected in A. philoxeroides leaves infested with female adults and male adults was 41 and 40, respectively, mainly including (E)-4,8-dimethylnona-1,3,7-triene (DMNT), trans-nerolidol, and 1-tridecene.

Table 1.
Volatile components in healthy, mechanical damaged and A. hygrophila feeding of A. philoxeroides.
Among them, 5 compounds were detected in all treatments, and these compounds were identified as α-cedrene, 4,6-dimethyl-dodecane, eicosane, 2-ethylhexyl tert-butyl-ether, and heneicosane. Furthermore, 1 and 7 compounds were only found in the clean plants and the mechanically treated plants, respectively, and 61 volatiles were only observed in A. philoxeroides infested by A. hygrophila, which were absent in CK and MD treatments, and (3E,7E)-4,8,12-trimethyltrideca-1,3,7,11-tetraene (E, E-TMTT), trans-nerolidol, and (E)-β-farnesene were detected in the treatments with A. hygrophila.
Among the 97 volatile compounds, 37 compounds (38.14%) belong to alkenes, 29 compounds (29.90%) belong to alkanes, and there were 8 esters (8.25%), 8 alcohols (8.25%) and 6 ketones (6.19%) (Figure 1). In clean plants, volatile compounds included alkenes (6 compounds), alkanes (9 compounds), esters (2 compounds), alcohols (1 compound), and others (1 compound). The profile of the volatiles in mechanical treatments was as follows: alkenes (3 compounds), alkanes (19 compounds), esters (3 compounds), and alcohols (1 compound) (Figure 1). In the A. philoxeroides infested by 1st instar (1L), 2nd instar (2L), 3rd instar (3L), females and males of A. hygrophila, volatile compounds included alkenes (11, 19, 28, 9, 18 compounds), alkanes (12, 7, 12, 19, 10 compounds), esters (2, 0, 2, 5, and 1 compounds), alcohols (4, 2, 2, 5, and 3 compounds), ketones (1, 1, 2, 1, and 5 compounds), and others (2, 3, 4, 1, and 3 compounds), respectively.
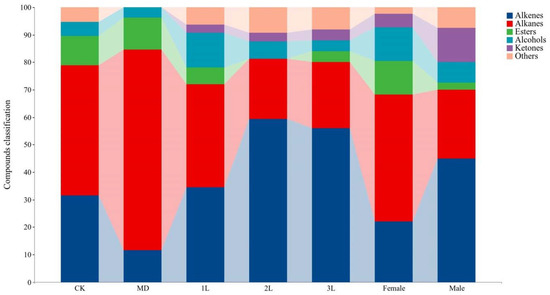
Figure 1.
Variation in volatile compounds classification of the A. philoxeroides treated with clean plants (CK), mechanical damaged (MD), infested by 1st instar (1L), 2nd instar (2L), 3rd instar (3L), females and males of A. hygrophila.
3.2. OPLS-DA Analysis of Volatile Components in Leaves
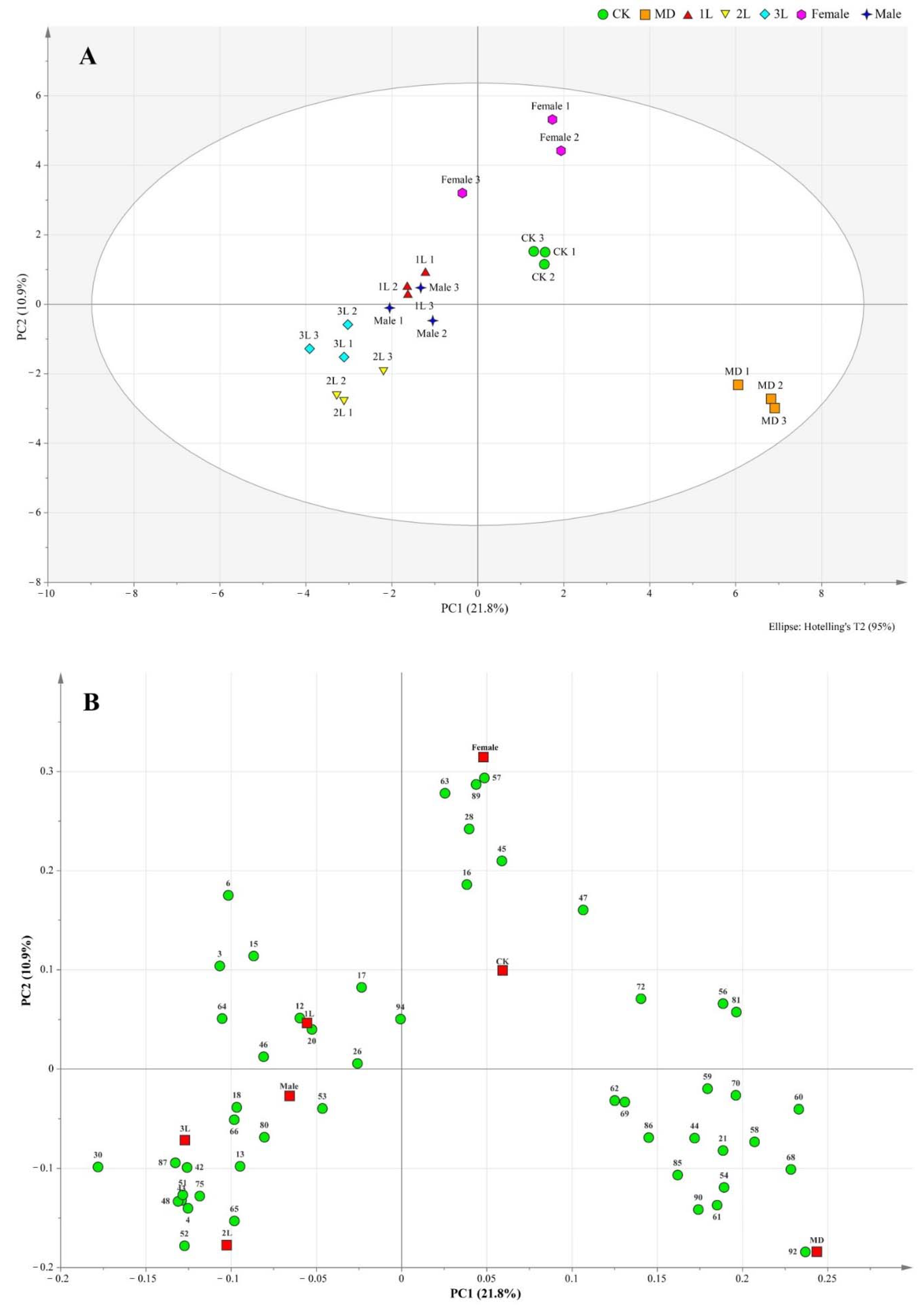
Projections to latent structures-orthogonal partial least-squares discrimination analysis (OPLS-DA) of all treatments together are presented in Figure 1, where the different treatments were separated from each other, especially insect feeding from CK and MD treatments (Figure 2A).
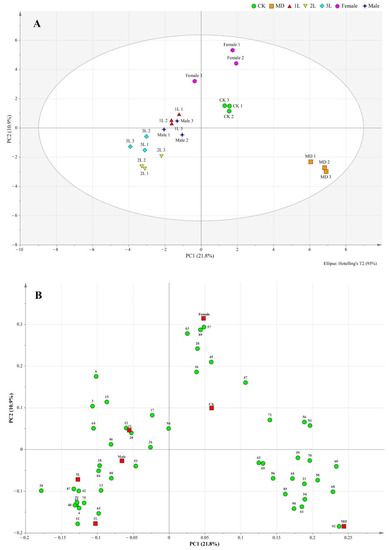
Figure 2.
Score scatter plot (A) and loading scatter plot (B) of orthogonal partial least squares-discrimination analysis (OPLS-DA) based on headspace composition of A. philoxeroides treated with clean plants (CK), mechanically damaged (MD), and infested by 1st instar (1L), 2nd instar (2L), 3rd instar (3L), females and males of A. hygrophila. For compound identity in relation to the numbering in the loading plot, please refer to Table 1.
The correlations among the contributions of these compounds with the different treatments were clearly visible from the loading scatter plot (Figure 2B).
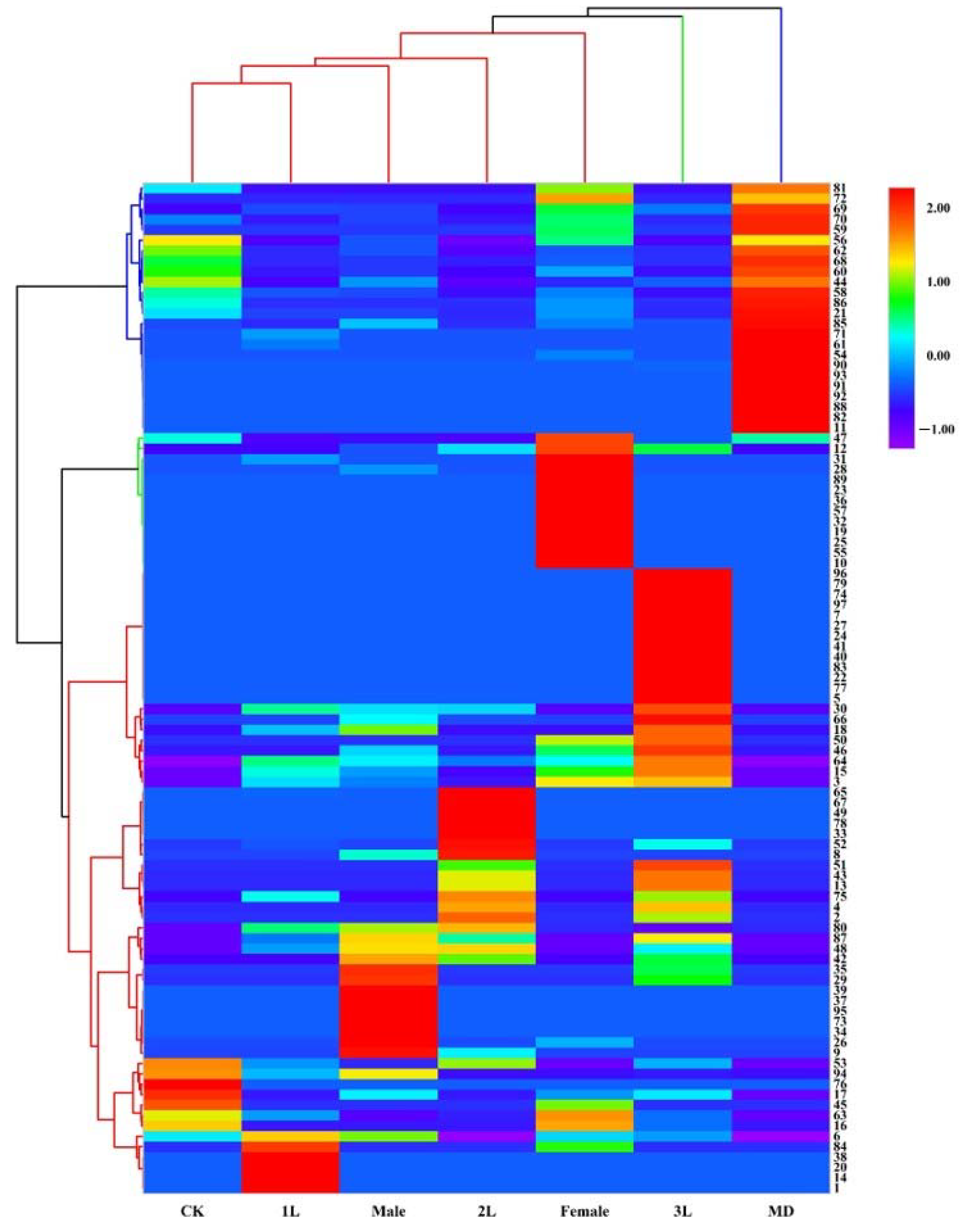
The loading plot shows the distribution of volatile components corresponding to the distribution and location of sample points in the score plot. Among all the volatiles in this study, 19 contributed the most to the separation among the treatments, with variable importance for the projection (VIP) values >1 (Table 2). These compounds included 6, 15, 17, 20, 26, 30, 42, 43, 45, 48, 51, 52, 57, 63, 65, 66, 89, 92, 94. Heat map analysis showed that the volatile compounds of A. philoxeroides were well differentiated in different treatments (Figure 3).
Table 2.
Values of variable importance in the projection (VIP) of volatile compounds for the corresponding OPLS-DA plots (VIP > 1).
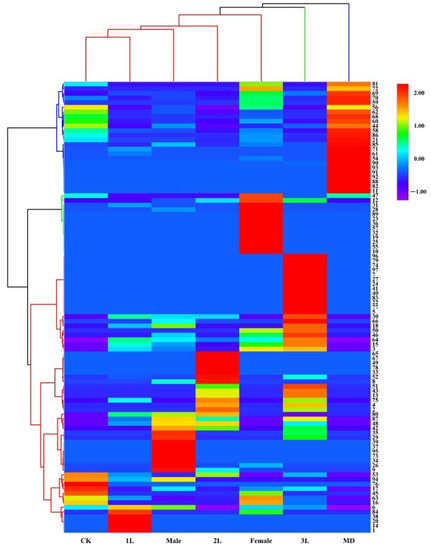
Figure 3.
Heat map analysis of volatile compounds of A. philoxeroides treated with clean plants (CK), mechanically damaged (MD), and infested by 1st instar (1L), 2nd instar (2L), 3rd instar (3L), females and males of A. hygrophila.
4. Discussion
When plants are subjected to herbivorous insect stress, they may release different quantities and species of volatiles from those released by healthy plants and even resynthesize and/or release more stressed VOCs [30,31,32]. This study found that A. philoxeroides damaged mechanically and infested with A. hygrophila released some new VOCs, such as new volatiles in the plant after mechanical damage, including 5-methyl-5-propyl-nonane, 7-epi-sesquithujene, 4-methyl-tetradecane, and tetratetracontane, and volatiles that appeared after being infested by A. hygrophila, including (+)-longifolene, E, E-TMTT, trans-nerolidol, and (E)-β-farnesene. Cui et al. reported that compared with healthy plants and mechanically damaged plants, feeding-damaged plants released unique chemicals, including eucalyptol, phytol, and β-ocimene [33]. Feeding by Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) and Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) induced quantitatively and qualitatively different HIPV blends from tomato plants [34]. This could be due to the direct or indirect defense response of host plants to regulate the behavior of herbivorous insects by adjusting the compounds of volatiles after being fed on by herbivorous insects.
Some volatiles also exist in healthy plants themselves, but the release amount increases significantly after being fed on by insects, which is also called HIPVs [35,36]. The increased HIPVs emitted from cotton plants damaged by Agrotis segetum (Lepidoptera: Noctuidae) larvae mainly consisted of linalool, β-caryophyllene, humulene, tetradecane, and hexadecane. [37]. In this study, it was found that the plants fed on by A. hygrophila released more (E)-4, 8-dimethylnona-1,3,7-triene (DMNT), α-farnesene, amd 8-hexyl-pentadecane (Table 1). These HIPVs play important roles in host plants’ resistance to herbivorous insect feeding [32]. The OPLS-DA analysis identified that 19 major volatile compounds played major roles in discriminating the plants fed by A. hygrophila, mechanically damage plants and healthy plants, such as (3E)-4, 8-dimethyl-1,3, 7-nontriene, copaene, caryophyllene, trans-nerolidol, cedarol and β-cedarene. These volatile compounds are closely related to the defense of host plants [38], regulation of insect behavior [18,39] and attraction of insect enemies [4,35].
Host localization and recognition by herbivorous insects depend on specific volatiles released by host plants [18,40]. Jacobi et al. found that Dichelops furcatus (Hemiptera: Pentatomidae) could distinguish hosts according to volatiles released by different varieties of the same hosts, and linalool was the main clue for their localization [16]. Li et al. demonstrated that the accurate localization of A. hygrophila was based on DMNT emitted by A. philoxeroides, and the effect of this compound on host selection preference could be effectively exploited in biological control [28]. In our study, the relative contents of DMNT emitted from A. philoxeroides infested by A. hygrophila were significantly higher than that of the clean plants. It is indicated that DMNT is an important clue in the localization and recognition of host plants of A. hygrophila.
Plant volatiles not only affect the behavior of herbivorous insects but also attract the natural enemies of herbivorous insects, especially the HIPVs [4]. The HIPVs released by Tibraca limbativentris Stål (Heteroptera: Pentatomidae) and Glyphepomis spinosa Campos et Grazia (Heteroptera: Pentatomidae) feeding on rice are attractive to the natural enemy of the rice pest, Telenomus podisi Ashmead (Hymenoptera: Platygastridae) [41]. Our results showed that trans-nerolidol, cis-3-hexenyl benzoate, and α-farnesene all appeared after being fed on by A. hygrophila. Numerous studies showed that terpenoids and GLVs, such as trans-nerolidol, cis-3-hexenoacetate, trans-β-farnesene and jasmonone, play major roles in the attraction of natural enemies [35,42]. However, no studies have reported the natural enemies of A. hygrophila, which may be further studied on this basis.
The emitting of volatiles by insects feeding on host plants is a complex process. An understanding of volatiles of A. philoxeroides infested by different stages A. hygrophila is needed for developing alligatorweed management strategies based on semiochemicals. The results of this study showed that new volatile compounds, such as (+)-longifolene, E, E-TMTT, trans-nerolidol, and (E)-β-farnesene, were produced in A. philoxeroides after being fed on by A. hygrophila. At the same time, the contents of some volatiles increased, such as (E)-4, 8-dimethylnona-1,3,7-triene (DMNT), α-farnesene, and so on. Among them, DMNT is an important substance for the host plant localization and recognition of A. hygrophila, while trans-nerolidol and (E)-β-farnesene play important roles in natural enemy attraction. In the following studies, attention should be paid to the application of VOCs and HIPVs in the population collection and field population migration and monitoring of A. hygrophila, so as to improve the prevention and biocontrol of A. hygrophila on A. philoxeroides.
Author Contributions
Conceptualization, J.-W.F.; Data curation, M.-Z.S., J.-Y.L., L.F. and H.W.; Funding acquisition, M.-Z.S.; Investigation, M.-Z.S., J.-Y.L. and Y.-T.C.; Supervision, J.-W.F.; Writing—original draft, M.-Z.S., J.-Y.L. and Y.-T.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (31901951), National Key Research and Development Program of China (2021YFC2600400), Natural Science Foundation of Fujian Province (2020J011356), Project of Fujian Academy of Agricultural Sciences (GJYS2019002, YC2021006, YC2021017, CXTD2021002), and the “5511” Collaborative Innovation Project (XTCXGC2021017, XTCXGC2021011, XTCXGC2021020).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Min Jiang for their assistance with the investigation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Snoeren, T.A.; De Jong, P.W.; Dicke, M. Ecogenomic approach to the role of herbivore-induced plant volatiles in community ecology. J. Ecol. 2007, 95, 17–26. [Google Scholar] [CrossRef]
- Mathews, C.R.; Brown, M.W.; Bottrell, D.G. Leaf extrafloral nectaries enhance biological control of a key economic pest, Grapholita molesta (Lepidoptera: Tortricidae), in peach (Rosales: Rosaceae). Environ. Entomol. 2007, 36, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Heil, M. Herbivore-Induced Plant Volatiles: Targets, Perception and Unanswered Questions. New Phytol. 2014, 204, 297–306. [Google Scholar] [CrossRef]
- Joo, Y.; Goldberg, J.K.; Chrétien, L.T.; Kim, S.G.; Baldwin, I.T.; Schuman, M.C. The circadian clock contributes to diurnal patterns of plant indirect defense in nature. J. Integr. Plant Biol. 2019, 61, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Felton, G.W. Priming of antiherbivore defensive responses in plants. Insect Sci. 2013, 20, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Ul Hassan, M.N.; Zainal, Z.; Ismail, I. Green leaf volatiles: Biosynthesis, biological functions and their applications in biotechnology. Plant Biotechnol. J. 2015, 13, 727–739. [Google Scholar] [CrossRef]
- Takabayashi, J.; Shiojiri, K. Multifunctionality of herbivory-induced plant volatiles in chemical communication in tritrophic interactions. Curr. Opin. Insect Sci. 2019, 32, 110–117. [Google Scholar] [CrossRef]
- Zhang, P.; Zhao, C.; Ye, Z.; Yu, X. Trade-off between defense priming by herbivore-induced plant volatiles and constitutive defense in tomato. Pest Manag. Sci. 2020, 76, 1893–1901. [Google Scholar] [CrossRef]
- Sugimoto, K.; Matsuia, K.; Iijimac, Y.; Akakabe, Y.; Muramoto, S.; Ozawa, R.; Uefune, M.; Sasaki, R.; Alamgir, K.; Akitake, S.; et al. Intake and transformation to a glycoside of (Z) -3-hexenol from infested neighbors reveals a mode of plant odor reception and defense. Proc. Natl. Acad. Sci. USA 2014, 111, 7144–7149. [Google Scholar] [CrossRef]
- Wenda-Piesik, A.; Piesik, D.; Nowak, A.; Wawrzyniak, M. Tribolium confusum responses to blends of cereal kernels and plant volatiles. J. Appl. Entomol. 2016, 140, 558–563. [Google Scholar] [CrossRef]
- Piesik, D.; Wenda-Piesik, A. Sitophilus granarius responses to blends of five groups of cereal kernels and one group of plant volatiles. J. Stored Prod. Res. 2015, 62, 36–39. [Google Scholar] [CrossRef]
- Hare, J.D. Ecological role of volatiles produced by plants in response to damage by herbivorous insects. Annu. Rev. Entomol. 2011, 56, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Wang, C. The ethological significance and olfactory detection of herbivore-induced plant volatiles in interactions of plants, herbivorous insects, and parasitoids. Arthropod Plant Inte. 2019, 13, 161–179. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef]
- Fincheira, P.; Quiroz, A.; Tortella, G.; Diez, M.C.; Rubilar, O. Current advances in plant-microbe communication via volatile organic compounds as an innovative strategy to improve plant growth. Microbiol. Res. 2021, 247, 126726. [Google Scholar] [CrossRef]
- Yang, W.; Ji, Z.; An, Y.; Xie, Q.; Zhao, Y.; Shen, Y. Plant volatile organic compounds (VOCs): A review. Chin. J. Ecol. 2008, 27, 1386–1392. [Google Scholar]
- Aráoz, M.C.; Jacobi, V.G.; Fernandez, P.; Albarracin, E.L.; Virla, E.G.; Hill, J.G.; Catalan, C.A.N. Volatiles mediate host-selection in the corn hoppers Dalbulus maidis (Hemiptera: Cicadellidae) and Peregrinus maidis (Hemiptera: Delphacidae). B Entomol. Res. 2019, 109, 633–642. [Google Scholar] [CrossRef]
- Jacobi, V.G.; Fernandez, P.C.; Barriga, L.G.; Almeida-Trapp, M.; Mithöfer, A.; Zavala, J.A. Plant volatiles guide the new pest Dichelops furcatus to feed on corn seedlings. Pest Manag. Sci. 2021, 77, 2444–2453. [Google Scholar] [CrossRef]
- Su, J.; Cai, Z.; Qiao, F.; Miao, L.; Yin, S.; Zheng, P. Numbers of natural enemies of corn pests attracted by plant volatile lures. Chin. J. Appl. Entomol. 2020, 57, 196. [Google Scholar]
- Krug, P.; Sosa, A.J. Mother knows best: Plant polyploidy affects feeding and oviposition preference of the alligator weed biological control agent, Agasicles hygrophila. BioControl 2019, 64, 623–632. [Google Scholar] [CrossRef]
- Coulson, J.R. Biological Control of Alligatorweed, 1959–1972: A Review and Evaluation; United States Department of Agriculture and United States Department of the Army: Washington, DC, USA, 1977.
- Buckingham, G.R. Biological control of alligatorweed, Alternanthera philoxeroides, the world’s first aquatic weed success story. Castanea 1996, 61, 232–243. [Google Scholar]
- Wu, Z.; Cai, Y.; Guo, Z.; Wang, T. Host specific tests for Agasicles hygrophila (Coleoptera: Chrysomelidae), a biological control agent of alligatorweed. J. East China 1994, 3, 98–100. [Google Scholar]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects–finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef]
- Rasmann, S.; Agrawal, A.A. Evolution of specialization: A phylogenetic study of host range in the red milkweed beetle (Tetraopes tetraophthalmus). Am. Nat. 2011, 177, 728–737. [Google Scholar] [CrossRef]
- Loreto, F.; Dicke, M.; Schnitzler, J.P.; Turlings, T.C. Plant volatiles and the environment. Plant Cell Environ. 2014, 37, 1905–1908. [Google Scholar] [CrossRef]
- Li, N.; Li, S.; Ge, J.; Schuman, M.C.; Wei, J.N.; Ma, R.Y. Manipulating two olfactory cues causes a biological control beetle to shift to non-target plant species. J. Ecol. 2017, 105, 1534–1546. [Google Scholar] [CrossRef]
- Duan, W.; Sun, P.; Chen, L.; Gao, S.; Shao, W.; Li, J. Comparative analysis of fruit volatiles and related gene expression between the wild strawberry Fragaria pentaphylla and cultivated Fragaria×ananassa. Eur. Food Res. Technol. 2018, 244, 57–72. [Google Scholar] [CrossRef]
- Dicke, M.; Van Loon, J.J.; Soler, R. Chemical complexity of volatiles from plants induced by multiple attack. Nat. Chem. Biol. 2009, 5, 317–324. [Google Scholar] [CrossRef]
- Douma, J.C.; Ganzeveld, L.N.; Unsicker, S.B.; Boeckler, G.A.; Dicke, M. What makes a volatile organic compound a reliable indicator of insect herbivory? Plant Cell Environ. 2019, 42, 3308–3325. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, F.; Chen, S.; Guan, Z.; Jiang, J.; Fang, W.; Chen, F. Effects of aphid herbivory on volatile organic compounds of Artemisia annua and Chrysanthemum morifolium. Biochem. Syst. Ecol. 2015, 60, 225–233. [Google Scholar] [CrossRef]
- Cui, S.; Ma, C.; Zhang, Y.; Bai, Q.; Chen, H.; Zang, L.; Zhou, Z. Effects of herbivory by the leaf beetle Ophraella communa on volatile compounds of the invasive common ragweed Ambrosia artemisiifolia. J. Environ. Entomol. 2021, 43, 1023–1033. [Google Scholar]
- Silva, D.B.; Weldegergis, B.T.; Van Loon, J.J.; Bueno, V.H. Qualitative and quantitative differences in herbivore-induced plant volatile blends from tomato plants infested by either Tuta absoluta or Bemisia tabaci. J. Chem. Ecol. 2017, 43, 53–65. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.C.; Unsicker, S.B.; Gershenzon, J. The specificity of herbivore-induced plant volatiles in attracting herbivore enemies. Trends Plant Sci. 2012, 17, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, L.; Martínez-Solís, M.; Frattini, A.; Ye, M.; Collado, M.C.; Turlings, T.C.; Erb, M.; Herrero, S. Can herbivore-induced volatiles protect plants by increasing the herbivores’ susceptibility to natural pathogens? Appl. Environ. Microb. 2019, 85, e01468-18. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, T.; Xia, S.; Xiao, H.; Lu, Y. Cotton plant volatiles induced by larval feeding of Agrotis segetum (Lepidoptera: Noctuidae) deter oviposition of conspecific females. Acta Entomol. Sinica 2022, 65, 304–311. [Google Scholar]
- Shivaramu, S.; Jayanthi, P.D.K.; Kempraj, V.; Anjinappa, R.; Nandagopal, B.; Chakravarty, A.K. What signals do herbivore-induced plant volatiles provide conspecific herbivores? Arthropod Plant Interact. 2017, 11, 815–823. [Google Scholar] [CrossRef]
- Kong, W.; Wang, Y.; Guo, Y.; Chai, X.; Li, J.; Ma, R. Behavioral effects of different attractants on adult male and female oriental fruit moths, Grapholita molesta. Pest Manag. Sci. 2020, 76, 3225–3235. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, Q.; Erb, M.; Turlings, T.C.; Ge, L.; Hu, L.; Li, J.; Han, X.; Zhang, T.; Lu, J. Specific herbivore-induced volatiles defend plants and determine insect community composition in the field. Ecol. Lett. 2012, 15, 1130–1139. [Google Scholar] [CrossRef]
- Ulhoa, L.A.; Barrigossi, J.A.F.; Borges, M.; Laumann, R.A.; Blassioli-Moraes, M.C. Differential induction of volatiles in rice plants by two stink bug species influence behaviour of conspecifics and their natural enemy Telenomus podisi. Entomol. Exp. Appl. 2020, 168, 76–90. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Urbaneja-Bernat, P.; Jaques, J.A.; Flors, V.; Urbaneja, A. Defensive plant responses induced by Nesidiocoris tenuis (Hemiptera: Miridae) on tomato plants. J. Pest Sci. 2015, 88, 543–554. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).