
Construction and Characterization of Immortalized Fibroblast Cell Line from Bactrian Camel


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Reagents
2.2. Isolation and Culture of Primary Camel Cells
2.3. Cell Culture and Passage
2.4. Cell Transfection and G418 Screening
2.5. Cell Viability and Proliferation Detection
2.6. Immunofluorescence Assay
2.7. Real-Time Quantitative PCR (qPCR) Assay
2.8. Western Blot Assay
2.9. Cell Cycle Detection
2.10. Karyotype Analysis
2.11. Statistical Analysis
3. Results
3.1. Morphological Characteristics of Primary Bactrian Camel Fibroblast (BCF)
3.2. Construction of Immortalized BCF Cell Line
3.3. Determination of BCF Culture Conditions
3.4. hTERT Gene Is Stably Expressed in Immortalized BCF
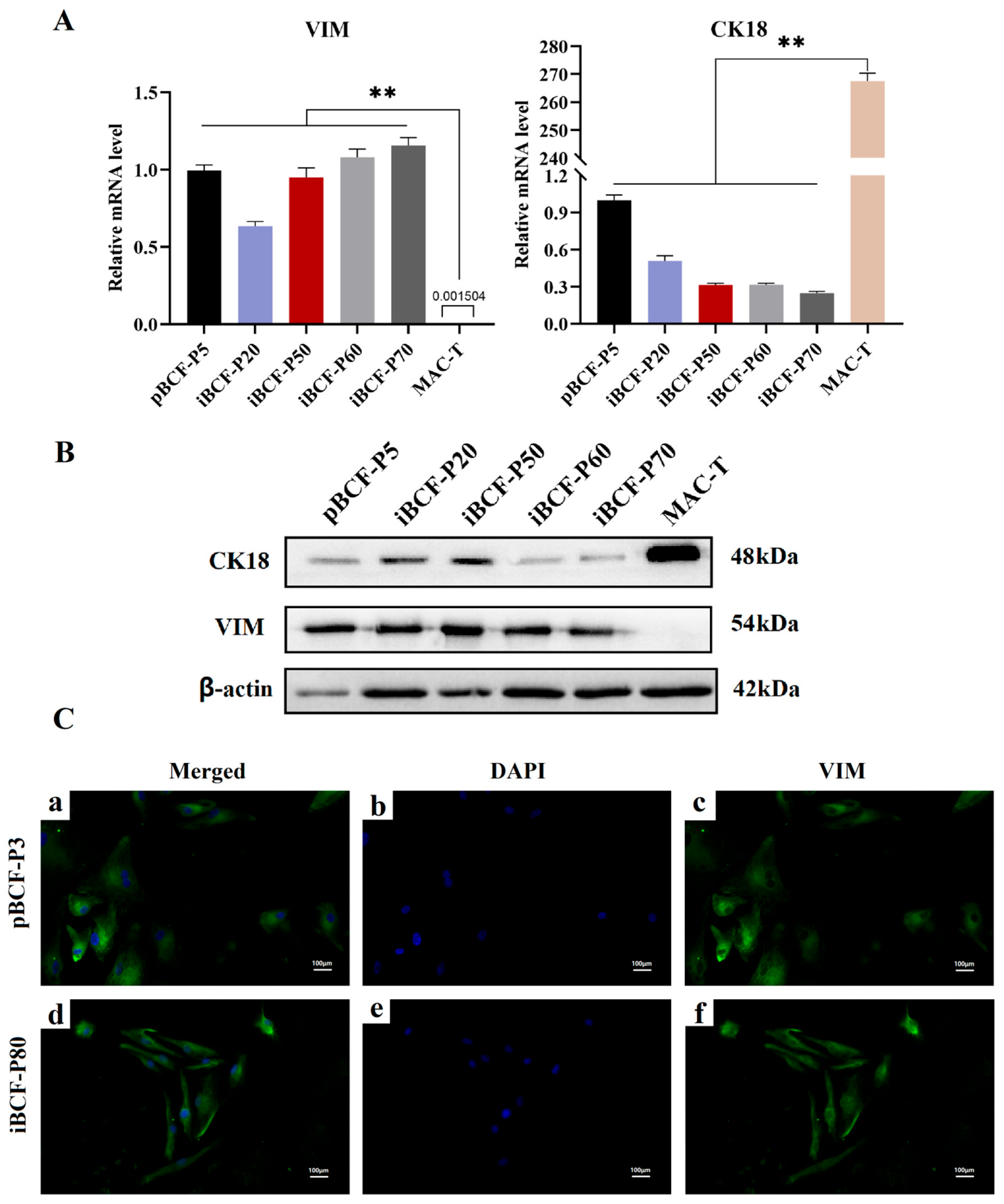
3.5. Fibroblast Marker Vimentin (VIM) Is Highly Expressed in Immortalized BCF
3.6. The Growth Characteristic of iBCF Cell Line
3.7. Karyotype Analysis of iBCF
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, Y.; Zhao, S.; Zhang, W.; Tang, H.; Yan, M.; Yong, F.; Bai, X.; Wu, X.; Zhang, Y.; Zhang, Q. Localization of FGF21 Protein and Lipid Metabolism-Related Genes in Camels. Life 2023, 13, 432. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hu, S.; Wang, M.; Zhao, B.; Yang, N.; Li, J.; Liu, M.; Zhou, J.; Bao, G.; Wu, X. Characterization and Establishment of an Immortalized Rabbit Melanocyte Cell Line Using the SV40 Large T Antigen. Int. J. Mol. Sci. 2019, 20, 4874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwack, M.H.; Yang, J.M.; Won, G.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Establishment and characterization of five immortalized human scalp dermal papilla cell lines. Biochem. Biophys. Res. Commun. 2018, 496, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Kurita, R.; Funato, K.; Abe, T.; Watanabe, Y.; Shiba, M.; Tadokoro, K.; Nakamura, Y.; Nagai, T.; Satake, M. Establishment and characterization of immortalized erythroid progenitor cell lines derived from a common cell source. Exp. Hematol. 2019, 69, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.M.; Lee, B.-H.; Chang, S.-N.; Oh, H.; Ryu, B.; Kim, U.; Park, J.-H. Establishment, characterization, and toxicological application of a spontaneous immortalized cell line from the striped field mouse, Apodemus agrarius. Vitr. Cell Dev. Biol. Anim. 2018, 54, 779–787. [Google Scholar] [CrossRef]
- Amoli, A.D.; Fazeli, S.A.S.; Aminafshar, M.; Kashan, N.E.J.; Khaledi, H. Identification of Bactrian camel cell lines using genetic markers. Vitr. Cell. Dev. Biol. Anim. 2018, 54, 265–271. [Google Scholar] [CrossRef]
- Miesen, L.; Wetzels, R.; Eymael, J.; Mooren, F.; Monge, V.V.; Berge, B.T.V.D.; Broek, M.V.D.; van der Velden, T.J.; Heuvel, L.P.V.D.; Wetzels, J.F.; et al. Establishment and characterization of a novel conditionally immortalized human parietal epithelial cell line. Exp. Cell Res. 2021, 405, 112712. [Google Scholar] [CrossRef]
- Tong, Y.; Cheng, L.-Q.; Li, X.; Yu, X.-P.; Shu, R.-H.; Zhang, J.-H.; Meng, Q.; Qin, Q.-L.; Tang, K.; Xu, J.-X.; et al. Establishment of an immortalized cell line derived from the pupal ovary of Mythimna separata (Lepidoptera: Noctuidae) and identification of the cell source. Cell Tissue Res. 2021, 386, 661–677. [Google Scholar] [CrossRef]
- Sone, M.; Nakamura, S.; Umeda, S.; Ginya, H.; Oshima, M.; Kanashiro, M.A.; Paul, S.K.; Hashimoto, K.; Nakamura, E.; Harada, Y.; et al. Silencing of p53 and CDKN1A establishes sustainable immortalized megakaryocyte progenitor cells from human iPSCs. Stem Cell Rep. 2021, 16, 2861–2870. [Google Scholar] [CrossRef]
- Holdsworth-Carson, S.J.; Colgrave, E.M.; Donoghue, J.F.; Fung, J.N.; Churchill, M.L.; Mortlock, S.; Paiva, P.; Healey, M.; Montgomery, G.W.; Girling, J.E.; et al. Generation of immortalized human endometrial stromal cell lines with different endometriosis risk genotypes. Mol. Hum. Reprod. 2019, 25, 194–205. [Google Scholar] [CrossRef]
- Nemeth, J.; Schundner, A.; Quast, K.; Winkelmann, V.E.; Frick, M. A Novel Fibroblast Reporter Cell Line for in vitro Studies of Pulmonary Fibrosis. Front. Physiol. 2020, 11, 567675. [Google Scholar] [CrossRef]
- Hanyu, S.; Sakuma, K.; Tanaka, A. A Study on the Effect of Human Dental Pulp Stem Cell Conditioned Medium on Human Oral Squamous Cell Carcinoma Cell Lines. J. Hard Tissue Biol. 2019, 28, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Nuta, O.; Rothkamm, K.; Darroudi, F. The Role of Telomerase in Radiation-Induced Genomic Instability. Radiat. Res. 2020, 193, 451–459. [Google Scholar] [CrossRef]
- Inada, E.; Saitoh, I.; Kubota, N.; Iwase, Y.; Kiyokawa, Y.; Shibasaki, S.; Noguchi, H.; Yamasaki, Y.; Sato, M. piggyBac Transposon-Based Immortalization of Human Deciduous Tooth Dental Pulp Cells with Multipotency and Non-Tumorigenic Potential. Int. J. Mol. Sci. 2019, 20, 4904. [Google Scholar] [CrossRef] [Green Version]
- Yin, B.; Song, Q.; Chen, L.; Li, X.; Han, Y.; Wang, X.; Dai, J.; Sun, X. Establishment of an immortalized intestinal epithelial cell line from tree shrews by lentivirus-mediated hTERT gene transduction. Cytotechnology 2019, 71, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Cruz, E.L.d.S.; Loureiro, F.J.A.; Silva, A.L.d.C.d.; Ramos, R.T.; Kataoka, M.S.d.S.; Pinheiro, J.d.J.V.; Júnior, S.d.M.A. Gene expression in cell lines from human ameloblastoma immortalized using hTERT and HPV16-E6/E7. Oral Dis. 2021, 28, 2230–2238. [Google Scholar] [CrossRef]
- Evtushenko, N.A.; Beilin, A.K.; Dashinimaev, E.B.; Ziganshin, R.H.; Kosykh, A.V.; Perfilov, M.M.; Rippa, A.L.; Alpeeva, E.V.; Vasiliev, A.V.; Vorotelyak, E.A.; et al. hTERT-Driven Immortalization of RDEB Fibroblast and Keratinocyte Cell Lines Followed by Cre-Mediated Transgene Elimination. Int. J. Mol. Sci. 2021, 22, 3809. [Google Scholar] [CrossRef]
- Gugjoo, M.B.; Amarpal Fazili, M.R.; Shah, R.A.; Sharma, G.T. Goat mesenchymal stem cell basic research and potential applications. Small Rumin. Res. 2020, 183, 106045. [Google Scholar] [CrossRef]
- Parfett, C.; Denhardt, D.T. Changes in Gene Expression During Senescence/Immortalization of Mouse Embryo Fibroblasts. In Growth Control during Cell Aging; Routledge: London, UK, 2020. [Google Scholar]
- Zhi, M.; Zhang, J.; Tang, Q.; Yu, D.; Gao, S.; Gao, D.; Liu, P.; Guo, J.; Hai, T.; Gao, J.; et al. Generation and characterization of stable pig pregastrulation epiblast stem cell lines. Cell Res. 2022, 32, 383–400. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, Y.; Wang, Z.; Luo, X.; Zhang, H.; Du, Q.; Chang, L.; Zhao, X.; Tong, D. Establishment and characterization of a telomerase immortalized porcine luteal cells. Theriogenology 2017, 94, 105–113. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, X.; Zhao, S.; Zhang, Y.; Ma, H.; Yang, Z.; Yang, W.; Zhao, C.; Wang, L.; Zhang, Q. Melatonin receptor depletion suppressed hCG-induced testosterone expression in mouse Leydig cells. Cell. Mol. Biol. Lett. 2019, 24, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.-Q.; Gao, Y.; Zhang, Y.; Yang, X.-P.; Yang, Z. cAMP/PKA/CREB signaling pathway-mediated effects of melatonin receptor genes on clock gene expression in Bactrian camel ovarian granulosa cells. Domest. Anim. Endocrinol. 2021, 76, 106609. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhang, S.; Long, C.; An, Z.; Xing, X.; Wen, F.; Bao, S. Mycoplasmas bovis P48 induces apoptosis in EBL cells via an endoplasmic reticulum stress-dependent signaling pathway. Veter-Microbiol. 2021, 255, 109013. [Google Scholar] [CrossRef]
- Rinkevich, Y.; Walmsley, G.G.; Hu, M.S.; Maan, Z.N.; Newman, A.M.; Drukker, M.; Januszyk, M.; Krampitz, G.W.; Gurtner, G.C.; Lorenz, H.P.; et al. Identification and isolation of a dermal lineage with intrinsic fibrogenic potential. Science 2015, 348, aaa2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lendahl, U.; Muhl, L.; Betsholtz, C. Identification, discrimination and heterogeneity of fibroblasts. Nat. Commun. 2022, 13, 3409. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, L.; Lin, P.; Jiang, T.; Wang, N.; Zhao, F.; Chen, H.; Tang, K.; Zhou, D.; Wang, A.; et al. An immortalized steroidogenic goat granulosa cell line as a model system to study the effect of the endoplasmic reticulum (ER)-stress response on steroidogenesis. J. Reprod. Dev. 2017, 63, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.-Y.; Yang, W.; Lee, E.-J.; Han, G.H.; Cho, H.; Chay, D.B.; Kim, J.-H. Establishment of five immortalized human ovarian surface epithelial cell lines via SV40 T antigen or HPV E6/E7 expression. PLoS ONE 2018, 13, e0205297. [Google Scholar] [CrossRef]
- Lee, K.H.; Nam, H.; Jeong, D.E.; Kim, S.S.; Song, H.J.; Pyeon, H.J.; Kang, K.; Hong, S.-C.; Nam, D.-H.; Joo, K.M. Sensitive Tumorigenic Potential Evaluation of Adult Human Multipotent Neural Cells Immortalized by hTERT Gene Transduction. PLoS ONE 2016, 11, e0158639. [Google Scholar] [CrossRef] [Green Version]
- Furihata, T.; Ito, R.; Kamiichi, A.; Saito, K.; Chiba, K. Establishment and characterization of a new conditionally immortalized human astrocyte cell line. J. Neurochem. 2016, 136, 92–105. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, J.; Liu, S. Establishment and Characterization of a Telomerase-Immortalized Sheep Trophoblast Cell Line. BioMed Res. Int. 2016, 2016, 5808575. [Google Scholar] [CrossRef] [Green Version]
- Balmus, G.; Trifonov, V.A.; Biltueva, L.S.; O’Brien, P.C.M.; Alkalaeva, E.S.; Fu, B.; Skidmore, J.A.; Allen, T.; Graphodatsky, A.S.; Yang, F.; et al. Cross-species chromosome painting among camel, cattle, pig and human: Further insights into the putative Cetartiodactyla ancestral karyotype. Chromosome Res. Int. J. Mol. Supramol. Evol. Asp. Chromosome Biol. 2007, 15, 499–515. [Google Scholar] [CrossRef]
- Baumgartner, A.; Hartshorne, C.F.; Polyzos, A.A.; Weier, H.-U.G.; Weier, J.F.; O’brien, B. Full Karyotype Interphase Cell Analysis. J. Histochem. Cytochem. 2018, 66, 595–606. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | pBCF | iBCF |
|---|---|---|
| Proliferative capacity | Weaker | Stronger |
| Passage number | About 10 passages | Infinite generations |
| Growth rate | Slow, 3–4 d | Fast, 2–3 d |
| Training difficulty | Difficult | Easy |
| The doubling time | Longer | Shorter |
| Optimal growth conditions (temp, CO2, media) | 37 °C, 5% CO2 humidity incubator, 10% FBS, 4.5 g/L glucose DMEM | 37 °C, 5% CO2 humidity incubator, 10% FBS, 4.5 g/L glucose DMEM |
| Biological characteristics | Approaching in vivo cell biology | Approaching in vivo cell biology |
| Genetic integrity | Unchanged | Unchanged |
| Karyotype | 2n = 74 | 2n = 74 |
| Malignant transformations or clonality | Negative | Negative |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, M.; Yong, F.; Ji, W.; Zhang, L.; Zhao, S.; Gao, Y. Construction and Characterization of Immortalized Fibroblast Cell Line from Bactrian Camel. Life 2023, 13, 1337. https://doi.org/10.3390/life13061337
Yan M, Yong F, Ji W, Zhang L, Zhao S, Gao Y. Construction and Characterization of Immortalized Fibroblast Cell Line from Bactrian Camel. Life. 2023; 13(6):1337. https://doi.org/10.3390/life13061337
Chicago/Turabian StyleYan, Meilin, Fang Yong, Wangye Ji, Lili Zhang, Shuqin Zhao, and Yuan Gao. 2023. "Construction and Characterization of Immortalized Fibroblast Cell Line from Bactrian Camel" Life 13, no. 6: 1337. https://doi.org/10.3390/life13061337