
Is the Cis-Element CACCC-Box a Master Regulatory Element during Cardiovascular Disease? A Bioinformatics Approach from the Perspective of the Krüppel-like Family of Transcription Factors
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
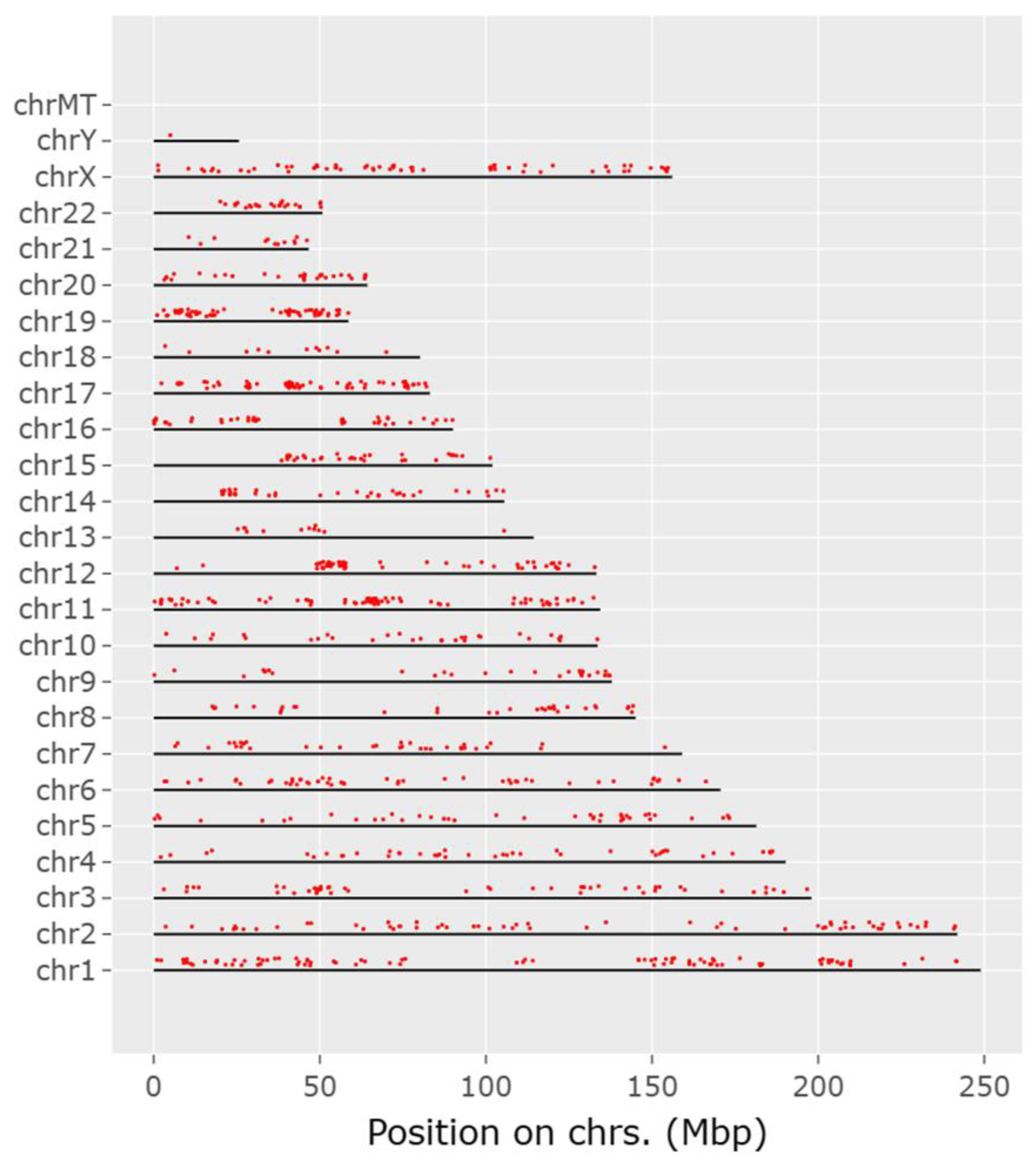
2.1. Raw data Collection and Web Scraping
2.2. Implementation with Python and Selenium
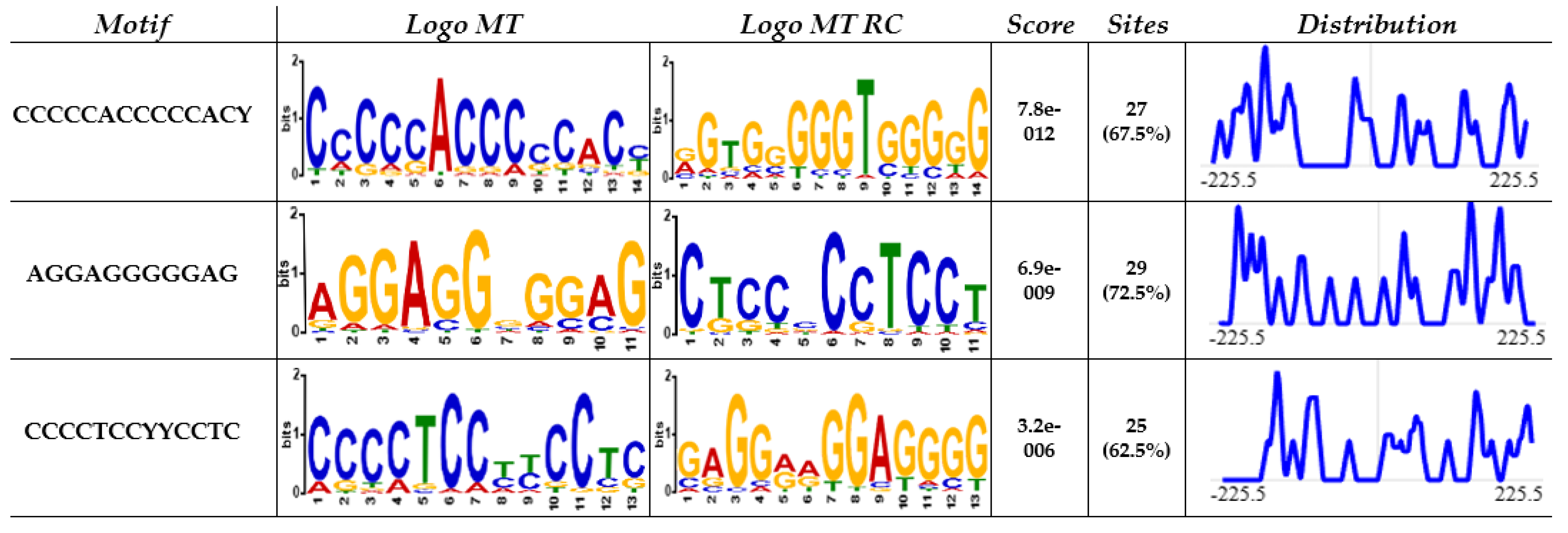
2.3. MEME
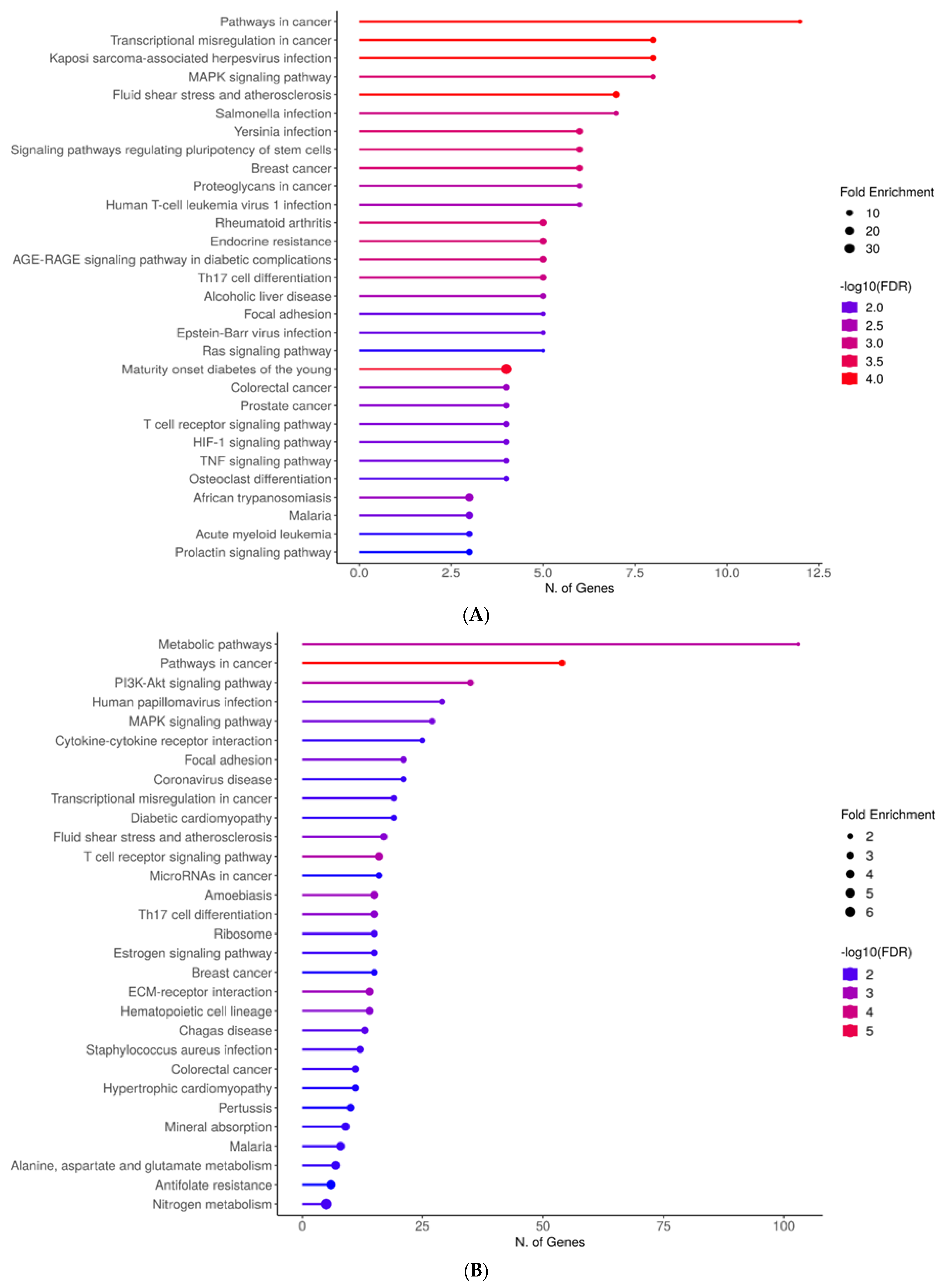
2.4. STRING
2.5. ShinyGO
2.6. Enrichr and Appyter
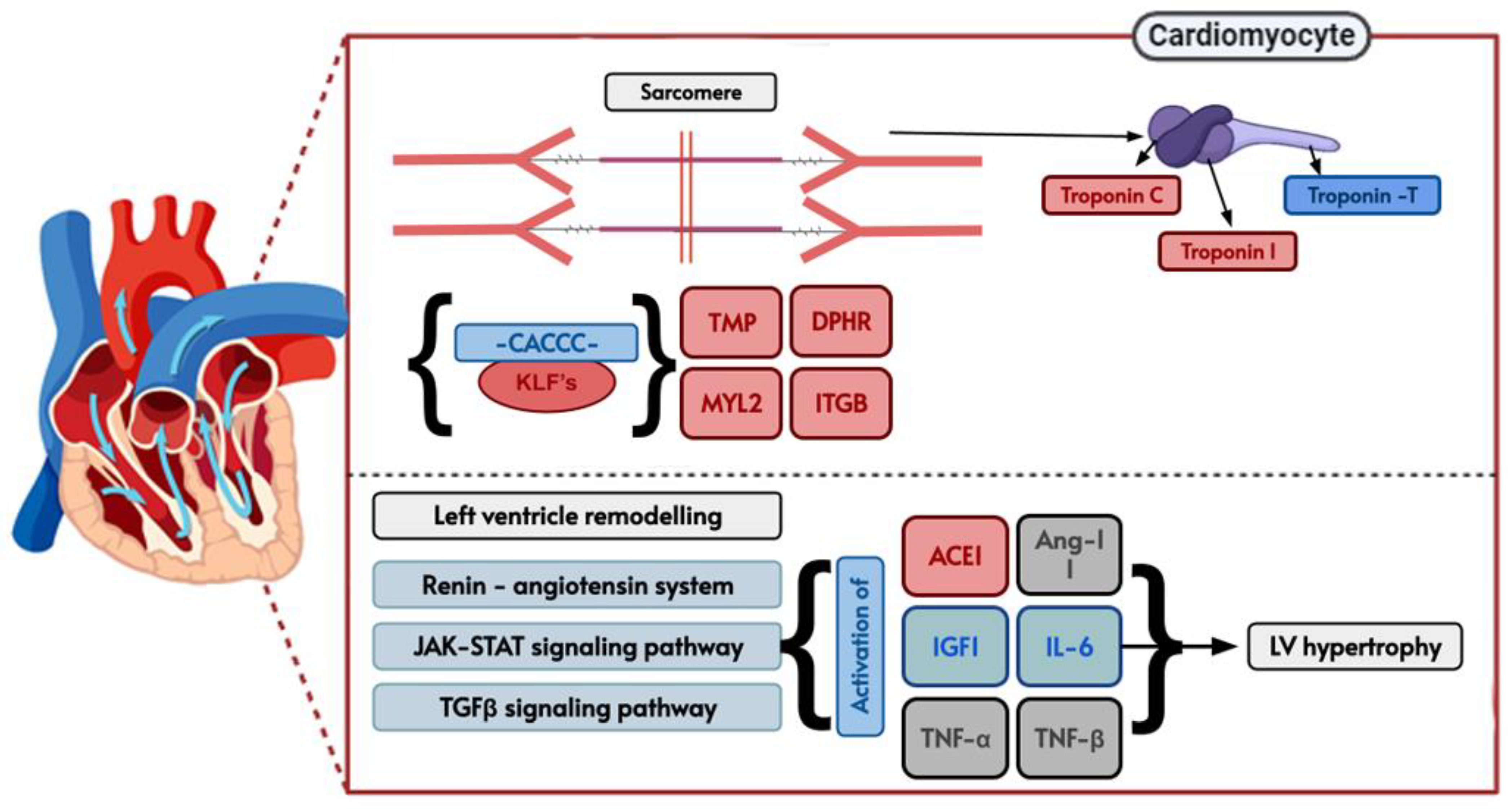
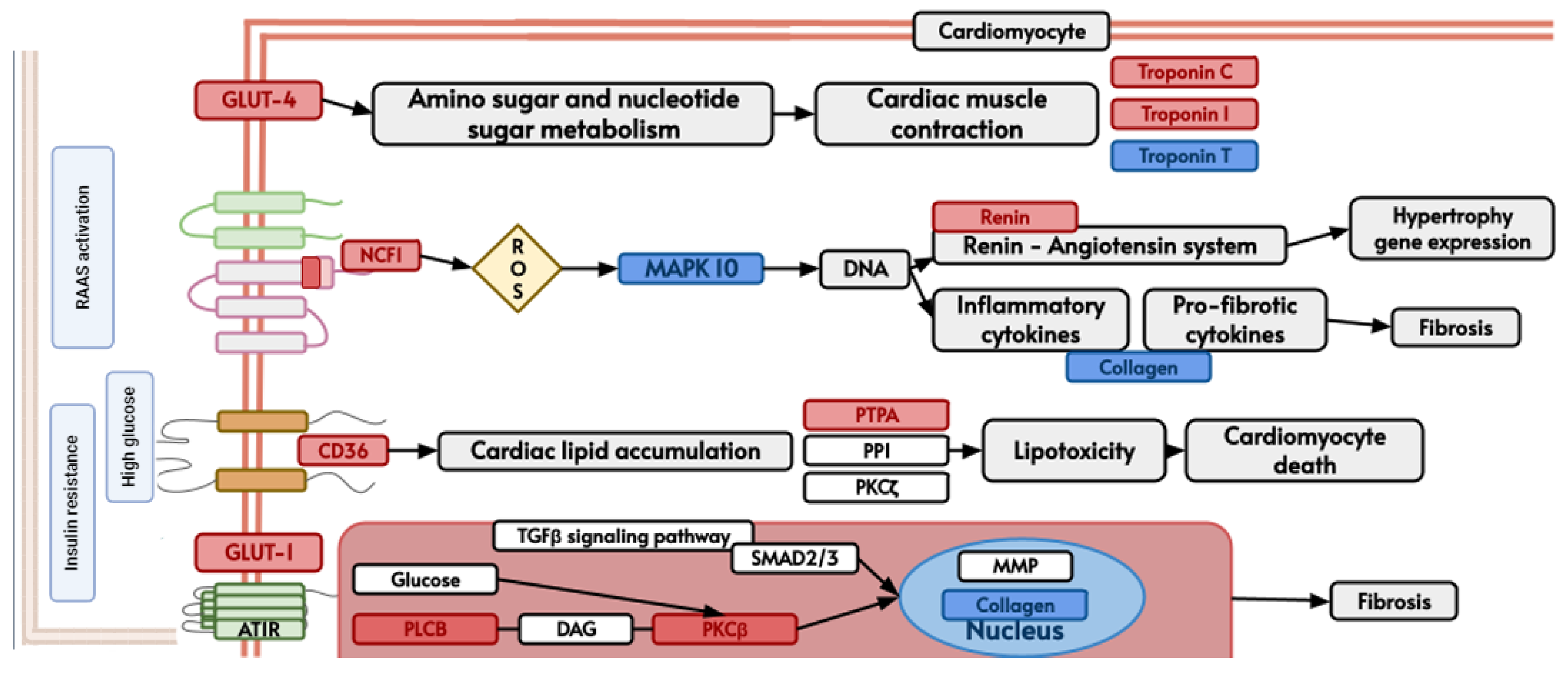
3. Results
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davie, K.; Jacobs, J.; Atkins, M.; Potier, D.; Christiaens, V.; Halder, G.; Aerts, S. Discovery of Transcription Factors and Regulatory Regions Driving in Vivo Tumor Development by ATAC-Seq and FAIRE-Seq Open Chromatin Profiling. PLoS Genet. 2015, 11, e1004994. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lei, C.; He, Q.; Pan, Z.; Xiao, D.; Tao, Y. Nuclear Functions of Mammalian MicroRNAs in Gene Regulation, Immunity and Cancer. Mol. Cancer 2018, 17, 64. [Google Scholar] [CrossRef] [PubMed]
- Frietze, S.; Farnham, P.J. Transcription Factor Effector Domains. Subcell Biochem. 2011, 52, 261–277. [Google Scholar] [PubMed]
- Santoyo-Suarez, M.G.; Mares-Montemayor, J.D.; Padilla-Rivas, G.R.; Delgado-Gallegos, J.L.; Quiroz-Reyes, A.G.; Roacho-Perez, J.A.; Benitez-Chao, D.F.; Garza-Ocañas, L.; Arevalo-Martinez, G.; Garza-Treviño, E.N.; et al. The Involvement of Krüppel-like Factors in Cardiovascular Diseases. Life 2023, 13, 420. [Google Scholar] [CrossRef] [PubMed]
- Okumura, K.; Sakaguchi, G.; Takagi, S.; Naito, K.; Mimori, T.; Igarashi, H. Sp1 Family Proteins Recognize the U5 Repressive Element of the Long Terminal Repeat of Human T Cell Leukemia Virus Type I through Binding to the CACCC Core Motif. J. Biol. Chem. 1996, 271, 12944–12950. [Google Scholar] [CrossRef]
- Fraizer, G.C.; Shimamura, R.; Zhang, X.; Saunders, G.F. PAX 8 Regulates Human WT1 Transcription through a Novel DNA Binding Site. J. Biol. Chem. 1997, 272, 30678–30687. [Google Scholar] [CrossRef] [PubMed]
- Dickson, J.; Gowher, H.; Strogantsev, R.; Gaszner, M.; Hair, A.; Felsenfeld, G.; West, A.G. VEZF1 Elements Mediate Protection from DNA Methylation. PLoS Genet. 2010, 6, e1000804. [Google Scholar] [CrossRef] [PubMed]
- De Albuquerque Dias, R.; Balbinot, K.M.; da Silva Kataoka, M.S.; de Melo Alves Júnior, S.; de Jesus Viana Pinheiro, J. Expression of Stem Cell Markers SALL4, LIN28A, and KLF4 in Ameloblastoma. Diagn. Pathol. 2023, 18, 92. [Google Scholar] [CrossRef]
- Lavallée, G.; Andelfinger, G.; Nadeau, M.; Lefebvre, C.; Nemer, G.; Horb, M.E.; Nemer, M. The Kruppel-like Transcription Factor KLF13 Is a Novel Regulator of Heart Development. EMBO J. 2006, 25, 5201–5213. [Google Scholar] [CrossRef]
- Abhinav, P.; Zhang, G.-F.; Zhao, C.-M.; Xu, Y.-J.; Wang, J.; Yang, Y.-Q. A Novel KLF13 Mutation Underlying Congenital Patent Ductus Arteriosus and Ventricular Septal Defect, as Well as Bicuspid Aortic Valve. Exp. Ther. Med. 2022, 23, 311. [Google Scholar] [CrossRef]
- Li, W.; Li, B.; Li, T.; Zhang, E.; Wang, Q.; Chen, S.; Sun, K. Identification and Analysis of KLF13 Variants in Patients with Congenital Heart Disease. BMC Med. Genet. 2020, 21, 78. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-S.; Wang, T.-M.; Qiao, X.-H.; Huang, R.-T.; Xue, S.; Dong, B.-B.; Xu, Y.-J.; Liu, X.-Y.; Yang, Y.-Q. KLF13 Loss-of-Function Variation Contributes to Familial Congenital Heart Defects. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11273–11285. [Google Scholar] [CrossRef]
- Kyriazis, I.D.; Hoffman, M.; Gaignebet, L.; Lucchese, A.M.; Markopoulou, E.; Palioura, D.; Wang, C.; Bannister, T.D.; Christofidou-Solomidou, M.; Oka, S.I.; et al. KLF5 Is Induced by FOXO1 and Causes Oxidative Stress and Diabetic Cardiomyopathy; American Heart Association: Dallas, TX, USA, 2021; Volume 128, ISBN 1215707142. [Google Scholar]
- Gao, C.; Qian, H.; Shi, Q.; Zhang, H. MicroRNA-363-3p Serves as a Diagnostic Biomarker of Acute Myocardial Infarction and Regulates Vascular Endothelial Injury by Targeting KLF2. Cardiovasc. Diagn. Ther. 2020, 10, 421–430. [Google Scholar] [CrossRef]
- Dai, Y.; Yan, T.; Gao, Y. Silence of MiR-32-5p Promotes Endothelial Cell Viability by Targeting KLF2 and Serves as a Diagnostic Biomarker of Acute Myocardial Infarction. Diagn. Pathol. 2020, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Vinjamur, D.S.; Wade, K.J.; Mohamad, S.F.; Haar, J.L.; Sawyer, S.T.; Lloyd, J.A. Krüppel-like Transcription Factors KLF1 and KLF2 Have Unique and Coordinate Roles in Regulating Embryonic Erythroid Precursor Maturation. Haematologica 2014, 99, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Walther, O. New Study Reveals Latest Data on Global Burden of Cardiovascular Disease. Available online: https://www.acc.org/About-ACC/Press-Releases/2023/12/11/18/48/New-Study-Reveals-Latest-Data-on-Global-Burden-of-Cardiovascular-Disease (accessed on 4 March 2024).
- Xie, Z.; Chen, J.; Wang, C.; Zhang, J.; Wu, Y.; Yan, X. Current Knowledge of Krüppel-like Factor 5 and Vascular Remodeling: Providing Insights for Therapeutic Strategies. J. Mol. Cell Biol. 2021, 13, 79–90. [Google Scholar] [CrossRef]
- Chang, E.; Nayak, L.; Jain, M.K. Krüppel-like Factors in Endothelial Cell Biology. Curr. Opin. Hematol. 2017, 24, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xu, S.; Liu, P.; Koroleva, M.; Zhang, S.; Si, S.; Jin, Z.G. Suberanilohydroxamic Acid as a Pharmacological Kruppel-Like Factor 2 Activator That Represses Vascular Inflammation and Atherosclerosis. J. Am. Heart Assoc. 2017, 6, e007134. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Liu, Y.; Wang, N.; Qi, Y.; Du, J. Krüppel-Like Factor 4 Transcriptionally Regulates TGF-Β1 and Contributes to Cardiac Myofibroblast Differentiation. PLoS ONE 2013, 8, e63424. [Google Scholar] [CrossRef]
- Yang, F.; Luo, L.; Zhu, Z.D.; Zhou, X.; Wang, Y.; Xue, J.; Zhang, J.; Cai, X.; Chen, Z.L.; Ma, Q.; et al. Chlorogenic Acid Inhibits Liver Fibrosis by Blocking the MiR-21-Regulated TGF-Β1/Smad7 Signaling Pathway in Vitro and in Vivo. Front. Pharmacol. 2017, 8, 929. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Liu, J.; Chen, X.; Duan, Y.; Wang, X.; Shen, Y.; Kuang, Y.; Zhuang, T.; Tomlinson, B.; et al. Endothelial Klf2-Foxp1-TGFβ Signal Mediates the Inhibitory Effects of Simvastatin on Maladaptive Cardiac Remodeling. Theranostics 2021, 11, 1609–1625. [Google Scholar] [CrossRef]
- Hoffman, M.; Palioura, D.; Kyriazis, I.D.; Cimini, M.; Badolia, R.; Rajan, S.; Gao, E.; Nikolaidis, N.; Schulze, P.C.; Goldberg, I.J.; et al. Cardiomyocyte Krüppel-Like Factor 5 Promotes De Novo Ceramide Biosynthesis and Contributes to Eccentric Remodeling in Ischemic Cardiomyopathy. Circulation 2021, 143, 1139–1156. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.K.; Wai, B.; Lang, C.C.; Levin, D.; Palmer, C.N.A.; Parry, H.M.; Velkoska, E.; Harrap, S.B.; Srivastava, P.M.; Burrell, L.M. Genetic Variation in Kruppel like Factor 15 Is Associated with Left Ventricular Hypertrophy in Patients with Type 2 Diabetes: Discovery and Replication Cohorts. EBioMedicine 2017, 18, 171–178. [Google Scholar] [CrossRef]
- Sweet, D.R.; Lam, C.; Jain, M.K. Evolutionary Protection of Krüppel-Like Factors 2 and 4 in the Development of the Mature Hemovascular System. Front. Cardiovasc. Med. 2021, 8, 645719. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.-K.; He, P.; Bialkowska, A.B.; Yang, V.W. SP and KLF Transcription Factors in Digestive Physiology and Diseases. Gastroenterology 2017, 152, 1845–1875. [Google Scholar] [CrossRef]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Cai, Y.; Dai, X.; Zhang, Q.; Dai, Z. Gene Expression of OCT4, SOX2, KLF4 and MYC (OSKM) Induced Pluripotent Stem Cells: Identification for Potential Mechanisms. Diagn. Pathol. 2015, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Shao, G. Pharmacological Analysis of Empagliflozin: Acting through the CaMKII Pathway in Type 2 Diabetes and Acute Cardiovascular Events. PLoS ONE 2022, 17, e0270152. [Google Scholar] [CrossRef] [PubMed]
- Yuce, K.; Ozkan, A.I. The Kruppel-like Factor (KLF) Family, Diseases, and Physiological Events. Gene 2024, 895, 148027. [Google Scholar] [CrossRef]
- Zheng, B.; Han, M.; Wen, J.-K. Role of Krüppel-like Factor 4 in Phenotypic Switching and Proliferation of Vascular Smooth Muscle Cells. IUBMB Life 2010, 62, 132–139. [Google Scholar] [CrossRef]
- Shankman, L.S.; Gomez, D.; Cherepanova, O.A.; Salmon, M.; Alencar, G.F.; Haskins, R.M.; Swiatlowska, P.; Newman, A.A.C.; Greene, E.S.; Straub, A.C.; et al. KLF4-Dependent Phenotypic Modulation of Smooth Muscle Cells Has a Key Role in Atherosclerotic Plaque Pathogenesis. Nat. Med. 2015, 21, 628–637. [Google Scholar] [CrossRef]
- Shindo, T.; Manabe, I.; Fukushima, Y.; Tobe, K.; Aizawa, K.; Miyamoto, S.; Kawai-Kowase, K.; Moriyama, N.; Imai, Y.; Kawakami, H.; et al. Krüppel-like Zinc-Finger Transcription Factor KLF5/BTEB2 Is a Target for Angiotensin II Signaling and an Essential Regulator of Cardiovascular Remodeling. Nat. Med. 2002, 8, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Chen, L.; Zhuang, Y.; Han, Y.; Tang, W.; Xia, F. Fibronectin 1 Inhibits the Apoptosis of Human Trophoblasts by Activating the PI3K/Akt Signaling Pathway. Int. J. Mol. Med. 2020, 46, 1908–1922. [Google Scholar] [CrossRef] [PubMed]
- Ericson, J.; Rashbass, P.; Schedl, A.; Brenner-Morton, S.; Kawakami, A.; van Heyningen, V.; Jessell, T.M.; Briscoe, J. Pax6 Controls Progenitor Cell Identity and Neuronal Fate in Response to Graded Shh Signaling. Cell 1997, 90, 169–180. [Google Scholar] [CrossRef]
- Christoforou, N.; Chellappan, M.; Adler, A.F.; Kirkton, R.D.; Wu, T.; Addis, R.C.; Bursac, N.; Leong, K.W. Transcription Factors MYOCD, SRF, Mesp1 and SMARCD3 Enhance the Cardio-Inducing Effect of GATA4, TBX5, and MEF2C during Direct Cellular Reprogramming. PLoS ONE 2013, 8, e63577. [Google Scholar] [CrossRef]
- Cornwell, J.D.; McDermott, J.C. MEF2 in Cardiac Hypertrophy in Response to Hypertension. Trends Cardiovasc. Med. 2022, 33, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Schwartz, R.J. Recruitment of the Tinman Homolog Nkx-2.5 by Serum Response Factor Activates Cardiac Alpha-Actin Gene Transcription. Mol. Cell Biol. 1996, 16, 6372–6384. [Google Scholar] [CrossRef]
- Dahlgren, A.; Zethelius, B.; Jensevik, K.; Syvänen, A.-C.; Berne, C. Variants of the TCF7L2 Gene Are Associated with Beta Cell Dysfunction and Confer an Increased Risk of Type 2 Diabetes Mellitus in the ULSAM Cohort of Swedish Elderly Men. Diabetologia 2007, 50, 1852–1857. [Google Scholar] [CrossRef]
- Aggarwal, A.; Costa, M.J.; Rivero-Gutiérrez, B.; Ji, L.; Morgan, S.L.; Feldman, B.J. The Circadian Clock Regulates Adipogenesis by a Per3 Crosstalk Pathway to Klf15. Cell Rep. 2017, 21, 2367–2375. [Google Scholar] [CrossRef]
- Zhang, L.; Prosdocimo, D.A.; Bai, X.; Fu, C.; Zhang, R.; Campbell, F.; Liao, X.; Coller, J.; Jain, M.K. KLF15 Establishes the Landscape of Diurnal Expression in the Heart. Cell Rep. 2015, 13, 2368–2375. [Google Scholar] [CrossRef]
- Butzner, M.; Leslie, D.L.; Cuffee, Y.; Hollenbeak, C.S.; Sciamanna, C.; Abraham, T. Stable Rates of Obstructive Hypertrophic Cardiomyopathy in a Contemporary Era. Front. Cardiovasc. Med. 2022, 8, 765876. [Google Scholar] [CrossRef] [PubMed]
- Dandamudi, S.; Slusser, J.; Mahoney, D.W.; Redfield, M.M.; Rodeheffer, R.J.; Chen, H.H. The Prevalence of Diabetic Cardiomyopathy: A Population-Based Study in Olmsted County, Minnesota. J. Card. Fail. 2014, 20, 304–309. [Google Scholar] [CrossRef]
- Grarup, N.; Andersen, G.; Krarup, N.T.; Albrechtsen, A.; Schmitz, O.; Jørgensen, T.; Borch-Johnsen, K.; Hansen, T.; Pedersen, O. Association Testing of Novel Type 2 Diabetes Risk Alleles in the JAZF1, CDC123/CAMK1D, TSPAN8, THADA, ADAMTS9, and NOTCH2 Loci with Insulin Release, Insulin Sensitivity, and Obesity in a Population-Based Sample of 4516 Glucose-Tolerant Middle-Aged Danes. Diabetes 2008, 57, 2534–2540. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, A.; Ladenvall, C.; Ahluwalia, T.S.; Kravic, J.; Krus, U.; Taneera, J.; Isomaa, B.; Tuomi, T.; Renström, E.; Groop, L.; et al. Effects of Common Genetic Variants Associated With Type 2 Diabetes and Glycemic Traits on α- and β-Cell Function and Insulin Action in Humans. Diabetes 2013, 62, 2978–2983. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, G.; QiLi, M.; Liang, H.; Li, T.; E, X.Q.; Feng, Y.; Zhang, Y.; Liu, X.; Qian, M.; et al. Aspirin Reduces Cardiac Interstitial Fibrosis by Inhibiting Erk1/2-Serpine2 and P-Akt Signalling Pathways. Cell. Physiol. Biochem. 2018, 45, 1955–1965. [Google Scholar] [CrossRef]
- Intengan, H.D.; Schiffrin, E.L. Vascular Remodeling in Hypertension. Hypertension 2001, 38, 581–587. [Google Scholar] [CrossRef]
- Brady, M. Sp1 and Sp3 Transcription Factors Are Required for Trans-Activation of the Human SERCA2 Promoter in Cardiomyocytes. Cardiovasc. Res. 2003, 60, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Nishio, Y.; Dong, Y.; Paris, M.; O’Keefe, R.J.; Schwarz, E.M.; Drissi, H. Runx2-Mediated Regulation of the Zinc Finger Osterix/Sp7 Gene. Gene 2006, 372, 62–70. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, B.; Zhou, P.; Zhang, R.-N.; He, M.; Yang, Z.; Wen, J.-K. Vascular Calcification Is Coupled with Phenotypic Conversion of Vascular Smooth Muscle Cells through Klf5-Mediated Transactivation of the Runx2 Promoter. Biosci. Rep. 2014, 34, e00148. [Google Scholar] [CrossRef]
- Petsophonsakul, P.; Furmanik, M.; Forsythe, R.; Dweck, M.; Schurink, G.W.; Natour, E.; Reutelingsperger, C.; Jacobs, M.; Mees, B.; Schurgers, L. Role of Vascular Smooth Muscle Cell Phenotypic Switching and Calcification in Aortic Aneurysm Formation. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1351–1368. [Google Scholar] [CrossRef]
- Lee, C.-T.; Ng, H.-Y.; Kuo, W.-H.; Tain, Y.-L.; Leung, F.-F.; Lee, Y.-T. The Role of TRPM7 in Vascular Calcification: Comparison between Phosphate and Uremic Toxin. Life Sci. 2020, 260, 118280. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Feng, W.; Su, X.; Luo, D.; Li, Z.; Zhou, Y.; Zhu, Y.; Zhang, M.; Chen, J.; Liu, B.; et al. SIRT6 Protects Vascular Smooth Muscle Cells from Osteogenic Transdifferentiation via Runx2 in Chronic Kidney Disease. J. Clin. Investig. 2022, 132, e150051. [Google Scholar] [CrossRef] [PubMed]
- Erez, N.; Truitt, M.; Olson, P.; Hanahan, D. Cancer-Associated Fibroblasts Are Activated in Incipient Neoplasia to Orchestrate Tumor-Promoting Inflammation in an NF-ΚB-Dependent Manner. Cancer Cell 2010, 17, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Paavola, J.; Alakoski, T.; Ulvila, J.; Kilpiö, T.; Sirén, J.; Perttunen, S.; Narumanchi, S.; Wang, H.; Lin, R.; Porvari, K.; et al. Vezf1 Regulates Cardiac Structure and Contractile Function. EBioMedicine 2020, 51, 102608. [Google Scholar] [CrossRef] [PubMed]
- Chiplunkar, A.R.; Lung, T.K.; Alhashem, Y.; Koppenhaver, B.A.; Salloum, F.N.; Kukreja, R.C.; Haar, J.L.; Lloyd, J.A. Krüppel-Like Factor 2 Is Required for Normal Mouse Cardiac Development. PLoS ONE 2013, 8, e54891. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Lu, H.; Liang, W.; Hu, W.; Zhang, J.; Chen, Y.E. Krüppel-like Factors and Vascular Wall Homeostasis. J. Mol. Cell Biol. 2017, 9, 352–363. [Google Scholar] [CrossRef]
- Kotlyarov, S.; Kotlyarova, A. Participation of Krüppel-like Factors in Atherogenesis. Metabolites 2023, 13, 448. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family Member | KLF | Direct Interaction |
|---|---|---|
| 1 | KLF-3 | NKX2-8, NKX2-2, LLGL2 |
| 1 | KLF-8 | NKX2-8, NKX2-2 |
| 1 | KLF-12 | TFAP2A, NKX2-2, DNASE1 |
| 2 | KLF-1 | SPI1, HBZ, NKX2-8, TET2, CD44, HBB, HBE1, HBG2, GATA2, AHSP EPB42, UBC |
| 2 | KLF-2 | GZMB, TNNT2, DUSP1, CCR7, LEF1, ZAP70, CXCR4, TEK, HAS2, CD44, FOXP3, FOS, FBXW7, ICAM1, GATA2, TRPV4, CD8A, TBX3 |
| 2 | KLF-4 | SALL4, COL1A1, SPI1, TNNT2, SOX9, MYOD1, TBX3, EED, EPAS1, EPCAM, LEF1, CDH2, NODAL, GRHL3, T, IGF1, FOS, HAS2, TBX5, TUBB3, NKX2-5, DLK1, MEF2C, GATA2, FN1, PAX9, SOX15, LEFTY2, NES, TCF7L2, FLNA, DKK1, RARG, KLF-6, KDM2B, TET2, ESR1, GFAP, CXCR4, IL6, MYH11, CD24, CD44, TCF4, PAX6, PBX1 |
| 2 | KLF-5 | SOX9, TBX3, TP63, NCOR1, FBXW7, NODAL, UCP3, GATA2, MKX, SUMO1, ESR1 |
| 2 | KLF-6 | PPP1R15A, FOS, DUSP1, KLF-4, ESR1, CYP2E1, NAT1, ANGPT1 |
| 2 | KLF-7 | PROKR2, GABPB1, TBX3, LEF1, NFIA, NKX2-8 |
| 3 | KLF-9 | NKX2-8, THRA, RB1, MAPK10, TFAP2A, ACTG2 |
| 3 | KLF-10 | NKX2-8, SMAD7, NKX2-2, FOS, FOXP3 |
| 3 | KLF-11 | HNF1A, NKX2-2, FOXP3, |
| 3 | KLF-13 | HBB, HBE1, TBX5 |
| 3 | KLF-14 | HHEX, TCF7L2, FHL2, CAMK1D, JAZF1 |
| 3 | KLF-16 | - |
| - | KLF-15 | NR3C1, NFIA, ARNTL, GATA2, MKX, NKX2-8 |
| - | KLF-17 | TG, NODAL, CD44, TBX3, SLC5A5 |
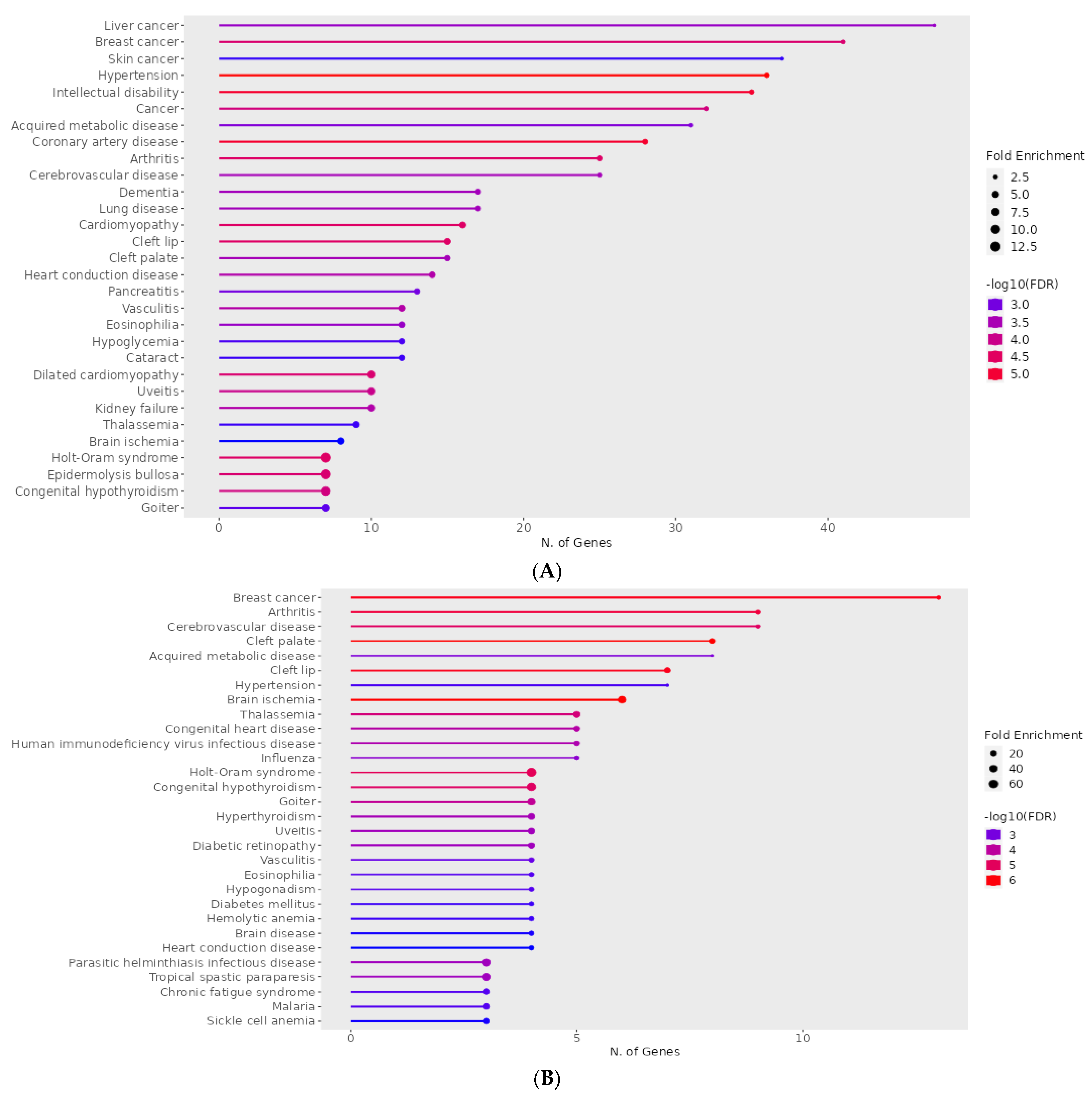
| (A) Disease | q-Value | p-Value | Genes |
|---|---|---|---|
| Holt–Oram syndrome | 2.91 × 10−6 | 0.001117367 | TFAP2B, SALL4, GJA5, TBX6, TBX5, NKX2-5, TBX3 |
| Hypertension | 6.45 × 10−6 | 0.00148591 | ENPEP, NCF1, CUL3, NPR3, KLK4, UMOD, ADM, CACNA1D, DBH, AAAS, NR3C1, ICAM1, ADD2, GJA5, NPY, TNNI3, GGT1, DUOX2, KL, ACE, ANGPT1, CYP4F2, CAV1, ATP6AP2, MTHFR, IGF1, FOS, RHOA, SELP, IL6, TH, NEDD4, SCNN1B, CYP11B1, TRPV4, REN, ANG |
| Dilated cardiomyopathy | 8.98 × 10−6 | 0.001549478 | BAG3, LAMA4, TNNC1, TNNT2, ANKRD1, LDB3, TNNI3, DSG2, CRYAB, VCL |
| Coronary artery disease | 9.42 × 10−6 | 0.001549478 | SLC22A4, KLK4, ADM, TNFRSF11B, LDB3, AAAS, MPO, CX3CL1, THBS4, ICAM1, XIRP1, TNNI3, CD36, GGT1, LDLR, SCAI, ACE, ANGPT1, APOA2, MTHFR, APOC3, IGF1, SELP, FABP3, IL6, TNNT1, TNNT2, REN, CD68 |
| Cerebrovascular disease | 0.0015946 | 0.04946049 | NTRK2, PRKCH, ACE, ANGPT1, CYP4F2, F11, KLK4, MTHFR, CXCR4, ADM, IGF1, FOS, MPO, RHOA, ICAM1, GFAP, SELP, IL6, TNNT1, TNNT2, REN, TNNI3, CD68, NES |
| Cardiomyopathy | 1.33 × 10−5 | 0.001700831 | TNNC1, LAMA4, PSEN1, TBX5, MTO1, JPH2, BAG3, GJA5, MYL2, TNNT2, DSG2, TNNI3, CTNNA3, FPGT-TNNI3K, CRYAB, VCL |
| Heart conduction disease | 0.000268757 | 0.013461245 | CAV1, IL5RA, DPT, CXCR4, MTHFR, TBX5, SLC4A4, DKK1, ARHGAP24, SFRP2, NFIA, CLDN14, TRPM7, BMPR1B |
| (B) Disease | q-value | p-value | Genes |
| Holt–Oram syndrome | 3.30 × 10−7 | 2.16 × 10−5 | SALL4, TBX5, NKX2-5, TBX3 |
| Hypertension | 0.00045705 | 0.006255711 | NR3C1, ICAM1, ANGPT1, IGF1, FOS, IL6, TRPV4 |
| Diabetes mellitus | 0.000426195 | 0.006255711 | UCP3, HNF1A, FOXP3 |
| Diabetic retinopathy | 3.12 × 10−5 | 0.000720179 | IL6, ANGPT1, IGF1, ICAM1 |
| Cerebrovascular disease | 1.25 × 10−6 | 6.14 × 10−5 | IL6, ANGPT1, TNNT2, CXCR4, FOS, IGF1, NES, GFAP, ICAM1 |
| Congenital heart disease | 1.14 × 10−5 | 0.000405807 | MEF2C, NKX2-5, TBX5, LEFTY2, NODAL |
| Heart conduction disease | 0.000629384 | 0.007274938 | CXCR4, TBX5, DKK1, NFIA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Loredo, J.A.; Santoyo-Suarez, M.G.; Rodríguez-Nuñez, O.; Benitez Chao, D.F.; Garza-Treviño, E.N.; Zapata-Morin, P.A.; Padilla-Rivas, G.R.; Islas, J.F. Is the Cis-Element CACCC-Box a Master Regulatory Element during Cardiovascular Disease? A Bioinformatics Approach from the Perspective of the Krüppel-like Family of Transcription Factors. Life 2024, 14, 493. https://doi.org/10.3390/life14040493
García-Loredo JA, Santoyo-Suarez MG, Rodríguez-Nuñez O, Benitez Chao DF, Garza-Treviño EN, Zapata-Morin PA, Padilla-Rivas GR, Islas JF. Is the Cis-Element CACCC-Box a Master Regulatory Element during Cardiovascular Disease? A Bioinformatics Approach from the Perspective of the Krüppel-like Family of Transcription Factors. Life. 2024; 14(4):493. https://doi.org/10.3390/life14040493
Chicago/Turabian StyleGarcía-Loredo, Juan Andrés, Michelle G. Santoyo-Suarez, Oscar Rodríguez-Nuñez, Diego Francisco Benitez Chao, Elsa N. Garza-Treviño, Patricio Adrián Zapata-Morin, Gerardo R. Padilla-Rivas, and Jose Francisco Islas. 2024. "Is the Cis-Element CACCC-Box a Master Regulatory Element during Cardiovascular Disease? A Bioinformatics Approach from the Perspective of the Krüppel-like Family of Transcription Factors" Life 14, no. 4: 493. https://doi.org/10.3390/life14040493