
Investigating Immunotoxicity in Black Carp (Mylopharyngodon piceus) Fingerlings Exposed to Niclosamide

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Niclosamide Preparation and Testing Animals
2.3. Construction of the NIC Exposure Model and Sampling
2.4. Liver and Serum Biochemical Parameters
2.5. RNA Isolation and qPCR Analysis
2.6. Transcriptome Analysis
2.7. Statistical Analysis
3. Results
3.1. Immune Biochemical Parameters of NIC Exposure in Liver and Serum
3.2. Immune-Related Gene Expression in Liver after NIC Exposure
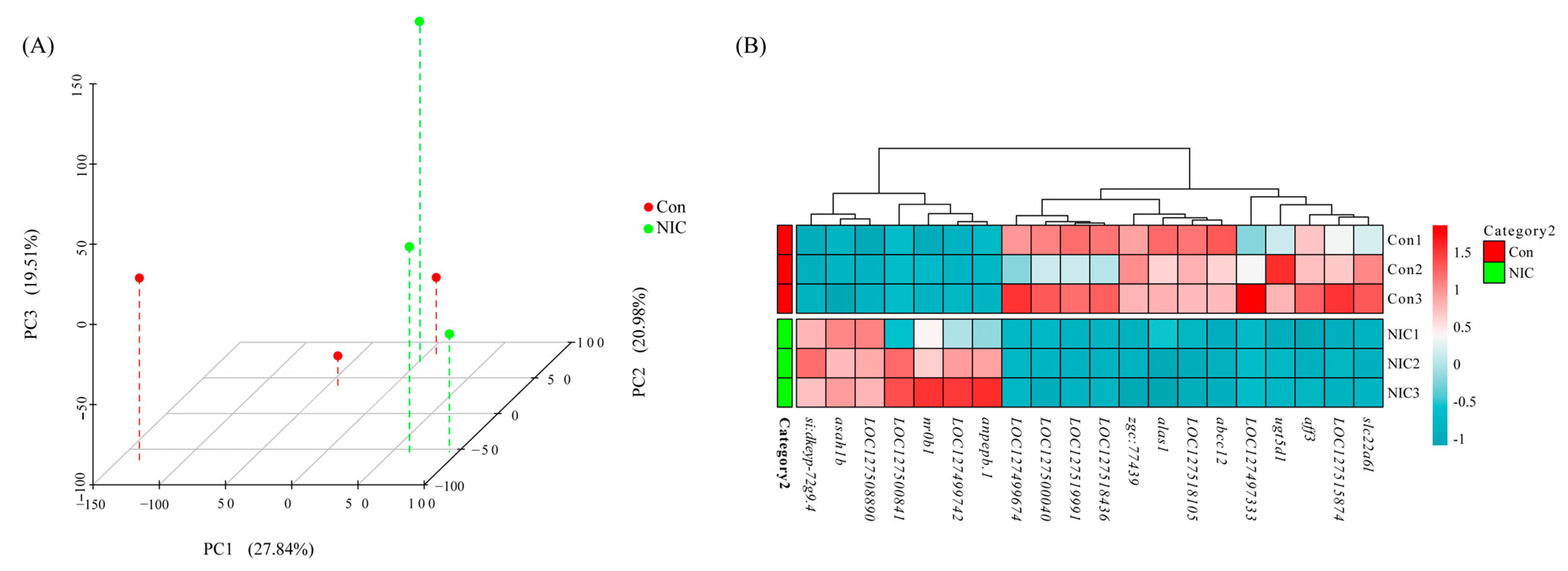
3.3. RNA-Seq Sequencing in Liver after NIC Exposure
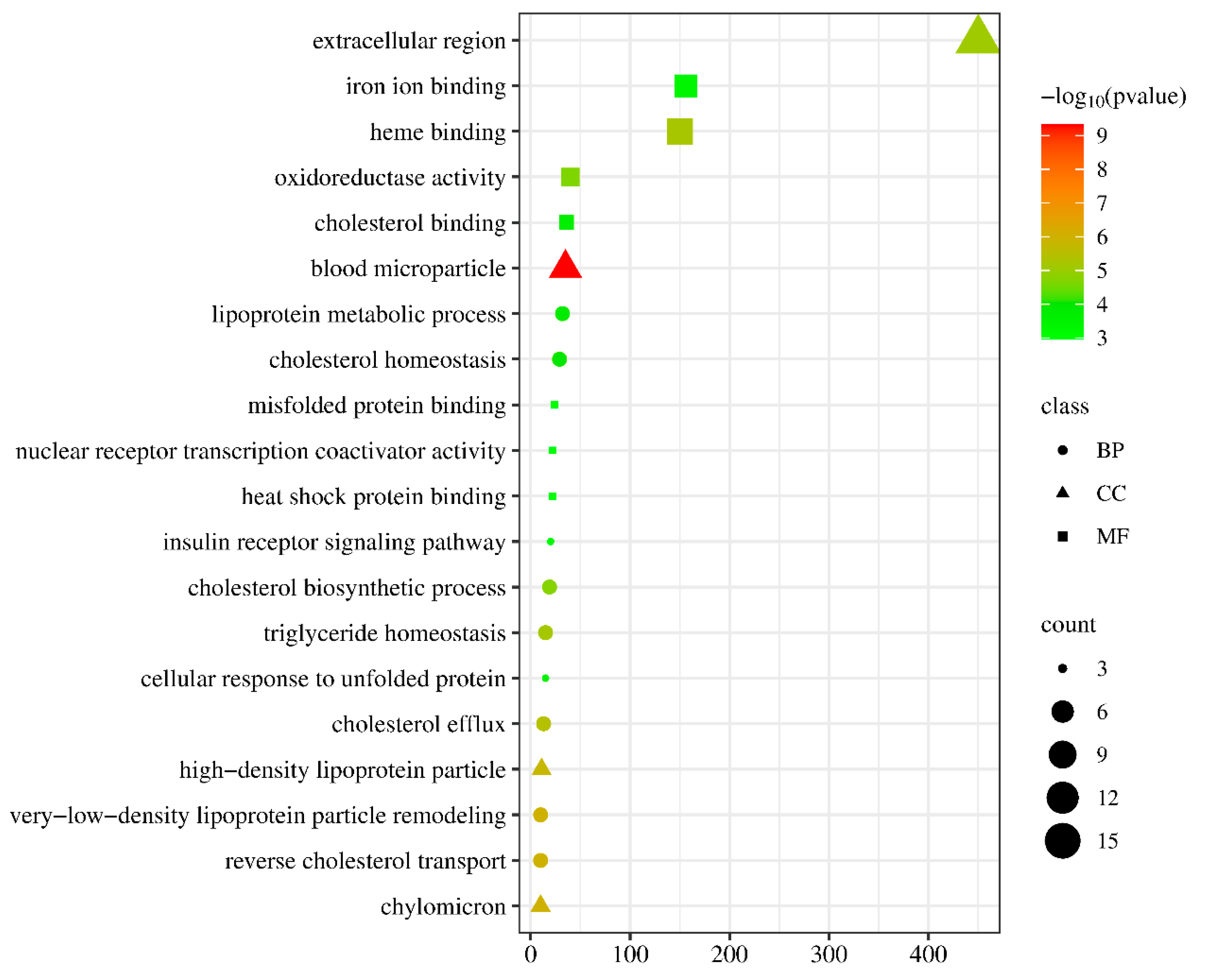
3.4. Analysis of Immune and Detoxification Process in the Liver Transcriptome after NIC Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ionescu, R.A.; Mitrovic, D.; Wilkie, M.P. Reversible disruptions to energy supply and acid-base balance in larval sea lamprey exposed to the pesticide: Niclosamide (2′,5-dichloro-4′-nitrosalicylanilide). Aquat. Toxicol. 2022, 242, 106006. [Google Scholar] [CrossRef]
- Ionescu, R.A.; Mitrovic, D.; Wilkie, M.P. Disturbances to energy metabolism in juvenile lake sturgeon (Acipenser fulvescens) following exposure to niclosamide. Ecotoxicol. Environ. Saf. 2022, 229, 112969. [Google Scholar] [CrossRef]
- Zhang, J.L.; Si, H.F.; Shang, X.F.; Zhang, X.K.; Li, B.; Zhou, X.Z.; Zhang, J.Y. New life for an old drug: In vitro and in vivo effects of the anthelmintic drug niclosamide against Toxoplasma gondii RH strain. Int. J. Parasitol.-Drug 2019, 9, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.G.; Zhen, J.H.; Quan, S.Q.; Liu, M.; Liu, L. Risk assessment for niclosamide residues in water and sediments from Nan Ji Shan Island within Poyang Lake region, China. Adv. Mater. Res. 2013, 721, 608–612. [Google Scholar] [CrossRef]
- Tudi, M.; Ruan, H.D.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef]
- Chen, X.; Yang, S.Y.; Zhu, B.; Zhang, M.; Zheng, N.; Hua, J.; Li, R.; Han, J.; Yang, L.; Zhou, B. Effects of environmentally relevant concentrations of niclosamide on lipid metabolism and steroid hormone synthesis in adult female zebrafish. Sci. Total Environ. 2024, 910, 168737. [Google Scholar] [CrossRef]
- Zhu, B.; He, W.; Yang, F.; Chen, L. High-throughput transcriptome sequencing reveals the developmental toxicity mechanisms of niclosamide in zebrafish embryo. Chemosphere 2020, 244, 125468. [Google Scholar] [CrossRef]
- Borowiec, B.G.; Birceanu, O.; Wilson, J.M.; McDonald, A.E.; Wilkie, M.P. Niclosamide is a much more potent toxicant of mitochondrial respiration than TFM in the invasive sea lamprey (Petromyzon marinus). Environ. Sci. Technol. 2022, 56, 4970–4979. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Li, X.; Ma, J.Z.; Yang, Y.; Luo, S.; Xie, X.D.; Ya, B.H.; Yang, J.; Luo, L.; Cao, L.Y. The potential endocrine disruption mechanism of anthelmintic drug niclosamide by activating estrogen receptors and estrogen-related receptors. Toxicology 2021, 457, 152805. [Google Scholar] [CrossRef]
- Wu, H.; Yuan, X.; Xie, M.; Gao, J.; Xiong, Z.; Song, R.; Xie, Z.; Ou, D. The impact of niclosamide exposure on the activity of antioxidant enzymes and the expression of glucose and lipid metabolism genes in black carp (Mylopharyngodon piceus). Genes 2023, 14, 2196. [Google Scholar] [CrossRef]
- Xiang, J.; Wu, H.; Gao, J.; Jiang, W.; Tian, X.; Xie, Z.; Zhang, T.; Feng, J.; Song, R. Niclosamide exposure disrupts antioxidant defense, histology, and the liver and gut transcriptome of Chinese soft-shelled turtle (Pelodiscus sinensis). Ecotox. Environ. Saf. 2023, 260, 115081. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, Y.; Chen, H.; Xu, L.; Wang, Q.; Feng, J. Gut–liver immune response and gut microbiota profiling reveal the pathogenic mechanisms of Vibrio harveyi in pearl gentian grouper (Epinephelus lanceolatus♂ × E. fuscoguttatus♀). Front. Immunol. 2020, 11, 607754. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Lei, L.; Sun, Y.; Shi, X.; Fu, K.; Hua, J.; Martyniuk, C.J.; Han, J.; Yang, L.; Zhou, B. Niclosamide exposure at environmentally relevant concentrations efficaciously inhibited the growth and disturbed the liver-gut axis of adult male zebrafish. Environ. Sci. Technol. 2022, 56, 11516–11526. [Google Scholar] [CrossRef]
- Segner, H.; Bailey, C.; Tafalla, C.; Bo, J. Immunotoxicity of xenobiotics in fish: A role for the aryl hydrocarbon receptor (AhR)? Int. J. Mol. Sci. 2021, 22, 9460. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Huang, Y.; Lu, Z.; Ma, Y.; Ran, X.; Yan, X.; Zhang, M.; Qiu, X.; Luo, L.; Yue, G.; et al. Sublethal effects of niclosamide on the aquatic snail Pomacea canaliculata. Ecotox. Environ. Saf. 2023, 259, 115064. [Google Scholar] [CrossRef]
- De Anna, J.S.; Castro, J.M.; Darraz, L.A.; Elias, F.D.; Carcamo, J.G.; Luquet, C.M. Exposure to hydrocarbons and chlorpyrifos alters the expression of nuclear receptors and antioxidant, detoxifying, and immune response proteins in the liver of the rainbow trout, Oncorhynchus mykiss. Ecotoxicol. Environ. Saf. 2021, 208, 111394. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, K.; Xu, C.; Jiang, Y.; Shan, J.; Zhang, Z.; Cai, J. Cypermethrin induces apoptosis, autophagy and inflammation via ERS-ROS-NF-κB axis in hepatocytes of carp (Cyprinus carpio). Pestic. Biochem. Phys. 2023, 196, 105625. [Google Scholar] [CrossRef]
- Cui, J.; Liu, Y.; Hao, Z.; Liu, Y.; Qiu, M.; Kang, L.; Teng, X.; Tang, Y. Cadmium induced time-dependent kidney injury in common carp via mitochondrial pathway: Impaired mitochondrial energy metabolism and mitochondrion-dependent apoptosis. Aquat. Toxicol. 2023, 261, 106570. [Google Scholar] [CrossRef]
- Chen, Q.; Lian, X.; An, J.; Geng, N.; Zhang, H.; Challis, J.K.; Luo, Y.; Liu, Y.; Su, G.; Xie, Y.; et al. Life cycle exposure to environmentally relevant concentrations of diphenyl phosphate (DPhP) inhibits growth and energy metabolism of zebrafish in a sex-specific manner. Environ. Sci. Technol. 2021, 55, 13122–13131. [Google Scholar] [CrossRef]
- Jiang, J.; Chen, L.; Liu, X.; Wang, L.; Wu, S.; Zhao, X. Histology and multi-omic profiling reveal the mixture toxicity of tebuconazole and difenoconazole in adult zebrafish. Sci. Total Environ. 2021, 795, 148777. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Z.X.; Li, S.W.; Ding, L.; Wei, X.F.; Han, S.C.; Wang, P.; Sun, Y.C. Multi-omics profiling and biochemical assays reveal the acute toxicity of environmental related concentrations of Di-(2-ethylhexyl) phthalate (DEHP) on the gill of crucian carp (Carassius auratus). Chemosphere 2022, 307 Pt 2, 135814. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, W.; Yao, J.; Yang, M.; Guo, Y.; Deng, Z.; Mao, Q.; Liu, S.; Duan, L. Comparative proteomics suggests the mode of action of a novel molluscicide against the invasive apple snail Pomacea canaliculata, intermediate host of Angiostrongylus cantonensis. Mol. Biochem. Parasit. 2022, 247, 111431. [Google Scholar] [CrossRef] [PubMed]
- Menezes, C.A.; Montresor, L.C.; Jangola, S.T.G.; Mattos, A.C.; Domingues, A.L.C.; Júnior, A.M.; Silva, C.C.M.; Barbosa, C.S.; Mendonca, C.L.F.; Massara, C.L.; et al. FioSchisto’s expert perspective on implementing WHO guidelines for schistosomiasis control and transmission elimination in Brazil. Front. Immunol. 2023, 14, 1268998. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, F.; Ai, X.; Wang, Z.; Yang, Q.; Dong, J.; Xu, N. Residue depletion and risk assessment of niclosamide in three species of freshwater fish. Food Addit. Contam. A 2018, 35, 1497–1507. [Google Scholar] [CrossRef]
- Jiao, Y.; Tao, Y.; Yang, Y.; Diogene, T.; Yu, H.; He, Z.; Han, W.; Chen, Z.; Wu, P.; Zhang, Y. Monobutyl phthalate (MBP) can dysregulate the antioxidant system and induce apoptosis of zebrafish liver. Environ. Pollut. 2020, 257, 113517. [Google Scholar] [CrossRef]
- Ullah, S.; Li, Z.; Hasan, Z.; Khan, S.U.; Fahad, S. Malathion induced oxidative stress leads to histopathological and biochemical toxicity in the liver of rohu (Labeo rohita, Hamilton) at acute concentration. Ecotoxicol. Environ. Saf. 2018, 161, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, J.; Kong, X.; Pei, C.; Zhao, X.; Li, L. Molecular characterization of polymeric immunoglobulin receptor and expression response to Aeromonas hydrophila challenge in Carassius auratus. Fish Shellfish Immun. 2017, 70, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Lv, X.; Zhang, Y.; Xin, Q.; Zou, Y.; Li, X. Tartrazine exposure results in histological damage, oxidative stress, immune disorders and gut microbiota dysbiosis in juvenile crucian carp (Carassius carassius). Aquat. Toxicol. 2021, 241, 105998. [Google Scholar] [CrossRef]
- Chen, J.; Liu, N.; Li, B.; Zhang, H.; Zhao, Y.; Cao, X. The effects of fipronil exposure on oxidative stress, non-specific immunity, autophagy, and apoptosis in the common car. Environ. Sci. Pollut. Res. 2021, 28, 27799–27810. [Google Scholar] [CrossRef]
- Huo, D.; Sun, L.; Ru, X.; Zhang, L.; Lin, C.; Liu, S.; Xin, X.; Yang, H. Impact of hypoxia stress on the physiological responses of sea cucumber Apostichopus japonicus: Respiration, digestion, immunity and oxidative damage. PeerJ 2018, 6, e4651. [Google Scholar] [CrossRef]
- Bols, N.C.; Brubacher, J.L.; Ganassin, R.C.; Lee, L.E.J. Ecotoxicology and innate immunity in fish. Dev. Comp. Immunol. 2001, 25, 853–873. [Google Scholar] [CrossRef] [PubMed]
- Twahirwa, I.; Wu, C.; Ye, J.; Zhou, Q. The effect of dietary fish meal replacement with blood meal on growth performance, metabolic activities, antioxidant and innate immune responses of fingerlings black carp, Mylopharyngodon piceus. Aquac. Res. 2020, 52, 702–714. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Ding, Q.; Teame, T.; Yang, Y.; Ran, C.; Zhang, Z.; Zhou, Z. Research advances in the structure, function, and regulation of the gill barrier in teleost fish. Water Biol. Secur. 2023, 2, 100139. [Google Scholar] [CrossRef]
- Reynaud, S.; Raveton, M.; Ravanel, P. Interactions between immune and biotransformation systems in fish: A review. Aquat. Toxicol. 2008, 87, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Regoli, F.; Giuliani, M.E.; Benedetti, M.; Arukwe, A. Molecular and biochemical biomarkers in environmental monitoring: A comparison of biotransformation and antioxidant defense systems in multiple tissues. Aquat. Toxicol. 2011, 105 (Suppl. S3), 56–66. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, M.P.; Hubert, T.D.; Boogaard, M.A.; Birceanu, O. Control of invasive sea lampreys using the piscicides TFM and niclosamide: Toxicology, successes & future prospects. Aquat. Toxicol. 2019, 211, 235–252. [Google Scholar] [PubMed]
- Lawrence, M.J.; Grayson, P.; Jeffrey, J.D.; Docker, M.F.; Garroway, C.J.; Wilson, J.M.; Manzon, R.G.; Wilkie, M.P.; Jeffries, K.M. Transcriptomic impacts and potential routes of detoxification in a lampricide-tolerant teleost exposed to TFM and niclosamide. Comp. Biochem. Physiol. Part D Genom. Proteom. 2023, 46, 101074. [Google Scholar] [CrossRef]
- Lawrence, M.J.; Mitrovic, D.; Foubister, D.; Bragg, L.M.; Sutherby, J.; Docker, M.F.; Servos, M.R.; Wilkie, M.P.; Jeffries, K.M. Contrasting physiological responses between invasive sea lamprey and non-target bluegill in response to acute lampricide exposure. Aquat. Toxicol. 2021, 237, 105848. [Google Scholar] [CrossRef] [PubMed]
- Schults, M.A.; Sanen, K.; Godschalk, R.W.; Theys, J.; Schooten, F.J.V.; Chiu, R.K. Hypoxia diminishes the detoxification of the environmental mutagen benzo [a] pyrene. Mutagenesis 2014, 29, 481–487. [Google Scholar] [CrossRef]
- Chen, J.; Cai, B.; Tian, C.; Jiang, D.; Shi, H.; Huang, Y.; Zhu, C.; Li, G.; Deng, S. RNA sequencing (RNA-Seq) analysis reveals liver lipid metabolism divergent adaptive response to low- and high-salinity stress in spotted scat (Scatophagus argus). Animals 2023, 13, 1503. [Google Scholar] [CrossRef]
- Sudip, B.; Krishnendu, S.; Parames, C.S. Cytochrome P450s: Mechanisms and biological implications in drug metabolism and its interaction with oxidative stress. Curr. Drug Metab. 2014, 15, 719–742. [Google Scholar]
- Lawrence, M.J.; Grayson, P.; Jeffrey, J.D.; Docker, M.F.; Garroway, C.J.; Wilson, J.M.; Manzon, R.G.; Wilkie, M.P.; Jeffries, K.M. Variation in the transcriptome response and detoxification gene diversity drives pesticide tolerance in fishes. Environ. Sci. Technol. 2022, 56, 12137–12147. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Abbreviation | Primer Sequences (5′-3′) | Annealing Temperature (°C) |
|---|---|---|---|
| Proinflammatory cytokines | |||
| Tumor necrosis factor α | TNF-α | Forward: ACGCTGCCCTTACCGAGGGT | 60 |
| Reverse: AGGGCCACAGCCAGAAGAGC | |||
| Interferon α | IFN-α | Forward: ATGGCTCGGCCGATACAGGA | 60 |
| Reverse: TGGCATCCATGAGGCGGATGA | |||
| Interleukin 1β | IL-1β | Forward: TCACTGGGAGGTGGTTCA | 59 |
| Reverse: GAGCTGGTTTAATGGTTGTT | |||
| Interleukin 6 | IL-6 | Forward: TGCCGGTCAAATCCGCATGGA | 60 |
| Reverse: CCCGGTGTCCACCCTTCCTCT | |||
| Interleukin 11 | IL-11 | Forward: CAGTACCAAAGTTGACGGATAT | 60 |
| Reverse: CGGGAGTAGGTGGGAGTGA | |||
| Interleukin 8 | IL-8 | Forward: CCTCACGGCGCGGGTTACAA | 60 |
| Reverse: CCGCCGCAGGTTGTCAGGTG | |||
| Innate defense molecules | |||
| Complement component 3 | C3 | Forward: CAAGTGGCTGGTTCTCAA | 56 |
| Reverse: ATGGCAATCACAATAAAGG | |||
| Complement component 9 | C9 | Forward: TCAGAAATCACGACGACCAA | 56 |
| Reverse: AGGGCATCCACATCATCCAA | |||
| Hepcidin | HEPC | Forward: GCAGCCGTTCCATTCGT | 56.5 |
| Reverse: GCCAGGGGATTTGTTTGT | |||
| Liver-expressed antimicrobial peptide 2 | LEAP | Forward: AAACCTCACGGTGCCTACT | 63 |
| Reverse: CTCCTGCATATTCCTGTCG | |||
| Natural resistance-associated macrophage protein | NRAMP | Forward: TCTGGTCTGGCGCTGTCT | 63 |
| Reverse: AACTCTGGCTGTTCGTCGTAG | |||
| Housekeeping gene | |||
| Beta-actin | β-actin | Forward: CCAGCAGATGTGGATTAGCA | 56 |
| Reverse: CAGTTTGAGTCGGCGTGA | |||
| Tissue | Parameters | Niclosamide Concentrations (μg/L) | ||
|---|---|---|---|---|
| Control | 10 μg/L NIC | 50 μg/L NIC | ||
| Liver | LYZ (U/g) | 1857.51 ± 139.73 c | 2587.39 ± 128.77 b | 3056.71 ± 107.07 a |
| POD (U/g) | 7505.66 ± 136.01 a | 7920.33 ± 276.36 a | 6600.33 ± 330.31 b | |
| C3 (μg/mL) | 73.03 ± 2.42 a | 82.38 ± 6.09 a | 83.62 ± 0.99 a | |
| C4 (μg/mL) | 41.29 ± 1.74 b | 47.21 ± 1.89 a | 52.03 ± 0.99 a | |
| ALP (ng/g) | 573.13 ± 34.96 b | 747.11 ± 41.13 a | 673.06 ± 28.55 ab | |
| IgM (ng/g) | 12,538.63 ± 1500.87 a | 12,040.37 ± 805.83 a | 14,046.03 ± 578.79 a | |
| IgT (ng/g) | 171.83 ± 7.91 b | 201.65 ± 2.65 a | 153.56 ± 7.83 b | |
| Serum | LYZ (U/g) | 858.40 ± 38.72 b | 961.33 ± 33.16 ab | 1073.10 ± 52.39 a |
| POD (U/g) | 491.67 ± 37.03 a | 563. 67 ± 16.35 a | 563.67 ± 35.12 a | |
| C3 (μg/mL) | 523.78 ± 16.03 b | 610.21 ± 37.82 a | 668.59 ± 25.88 a | |
| C4 (μg/mL) | 34.30 ± 0.99 b | 42.47 ± 3.19 ab | 46.31 ± 3.73 a | |
| ALP (ng/g) | 582.74 ± 39.93 b | 697.35 ± 60.24 ab | 807.87 ± 43.82 a | |
| IgM (ng/g) | 9579.19 ± 495.56 a | 12,154.30 ± 1191.52 a | 12,496.03 ± 1316.72 a | |
| IgT (ng/g) | 142.09 ± 8.30 b | 165.63 ± 9.91 ab | 190.20 ± 21.66 a | |
| Features | Gene Name | log2FoldChange | p-adj | Functions |
|---|---|---|---|---|
| LOC127517132 | uncharacterized | 2.083422335 | <0.001 | Immune response |
| LOC127518174 | tumor necrosis factor receptor superfamily member 14-like | 1.626307483 | 0.014 | |
| LOC127520149 | HERV-H LTR-associating protein 2 | 2.799184351 | 0.001 | |
| LOC127500841 | MHC class I antigen | 8.785602796 | <0.001 | |
| LOC127508899 | MHC class II antigen | −2.729012249 | 0.042 | |
| LOC127500693 | uncharacterized | −2.966246836 | <0.001 | |
| LOC127496265 | cytochrome P450 2G1-like | −1.208747206 | <0.001 | Detoxification |
| LOC127506642 | cytochrome P450 2F2-like | −1.160051571 | 0.002 | |
| LOC127510152 | cytochrome P450 family 2 subfamily K | −1.959354958 | <0.001 | |
| LOC127518105 | cytochrome P450 3A30 | −1.727033364 | <0.001 | |
| LOC127500040 | cytochrome P450 1A1 | −1.698724375 | <0.001 | |
| LOC127523202 | cytochrome P450 4F3 | −1.222647819 | <0.001 | |
| LOC127505748 | cytochrome b5 | −1.014338096 | <0.001 | |
| LOC127518436 | UDP-glucuronosyltransferase 1–6 | −1.515533776 | <0.001 | |
| ugt5d1 | UDP glucuronosyltransferase 5 family, polypeptide D1 | −1.917067215 | <0.001 | |
| slc35g1 | solute carrier family 35 member G1 | −1.071229676 | <0.001 | |
| slc35f2 | solute carrier family 35, member F1/2 | 2.707230679 | 0.013 | |
| abcc12 | ATP-binding cassette, sub-family C (CFTR/MRP), member 12 | −1.436705268 | <0.001 | |
| abcc2 | ATP-binding cassette, sub-family C (CFTR/MRP), member 2 | −1.036890355 | <0.001 | |
| LOC127509173 | hemoglobin cathodic subunit alpha-like | 1.608493192 | 0.003 | |
| bmf2 | BCL2 modifying factor 2 | 1.657943074 | <0.001 | Apoptotic process |
| ddit4 | DNA-damage-inducible transcript 4 | −2.753519746 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Yuan, X.; Tian, X.; Gao, J.; Xie, M.; Xie, Z.; Song, R.; Ou, D. Investigating Immunotoxicity in Black Carp (Mylopharyngodon piceus) Fingerlings Exposed to Niclosamide. Life 2024, 14, 544. https://doi.org/10.3390/life14050544
Wu H, Yuan X, Tian X, Gao J, Xie M, Xie Z, Song R, Ou D. Investigating Immunotoxicity in Black Carp (Mylopharyngodon piceus) Fingerlings Exposed to Niclosamide. Life. 2024; 14(5):544. https://doi.org/10.3390/life14050544
Chicago/Turabian StyleWu, Hao, Xiping Yuan, Xing Tian, Jinwei Gao, Min Xie, Zhonggui Xie, Rui Song, and Dongsheng Ou. 2024. "Investigating Immunotoxicity in Black Carp (Mylopharyngodon piceus) Fingerlings Exposed to Niclosamide" Life 14, no. 5: 544. https://doi.org/10.3390/life14050544