
Pharmacological Modulation of Rate-Dependent Depression of the Spinal H-Reflex Predicts Therapeutic Efficacy against Painful Diabetic Neuropathy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
3. Results
3.1. Spinal Pharmacology of RDD in Naïve Rats
3.2. Time Course of RDD and Spinal KCC2 Protein Expression in Diabetic Rats
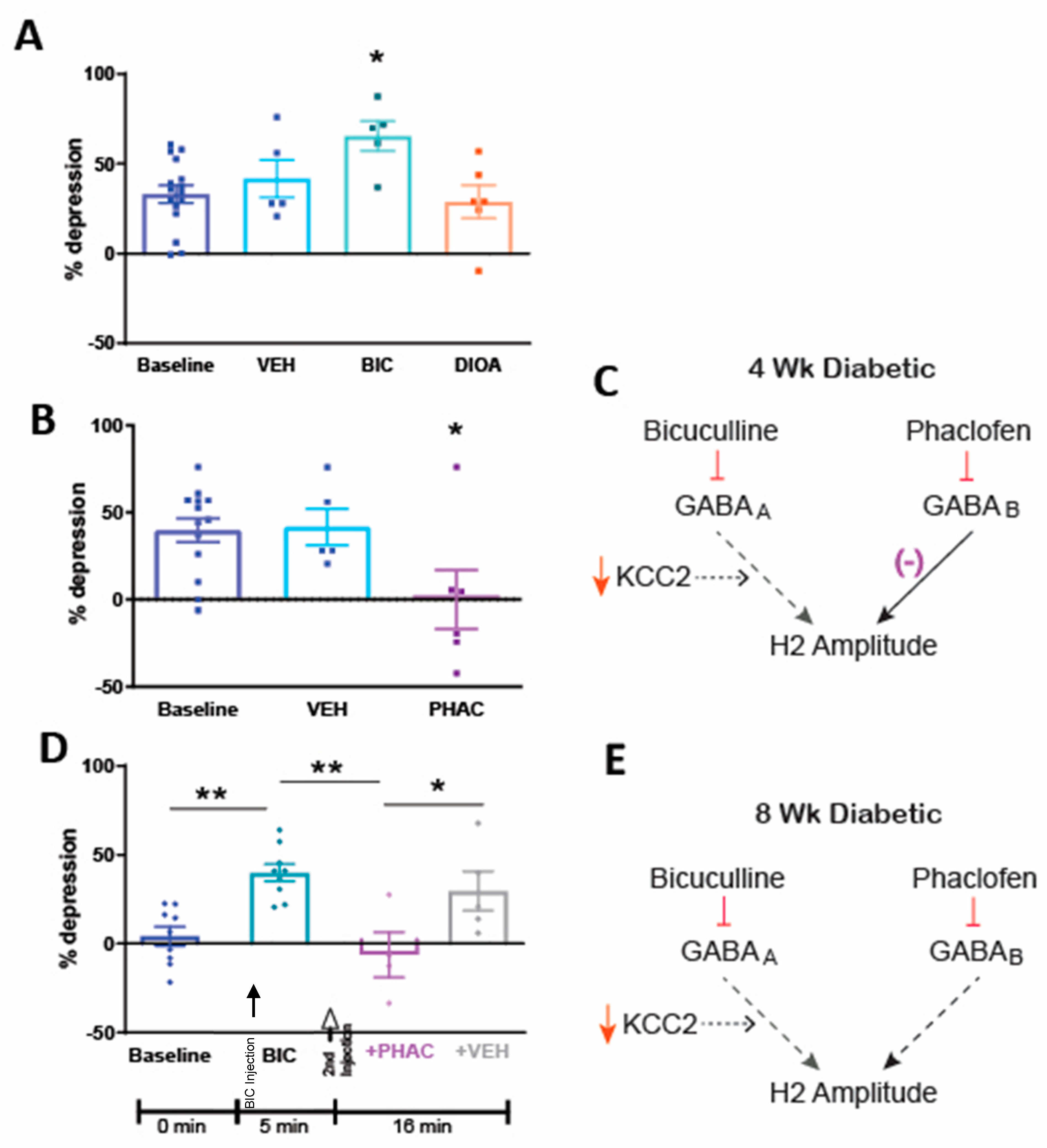
3.3. Spinal Pharmacology of RDD after 4 Weeks of Diabetes
3.4. Spinal Pharmacology of RDD after 8 Weeks of Diabetes
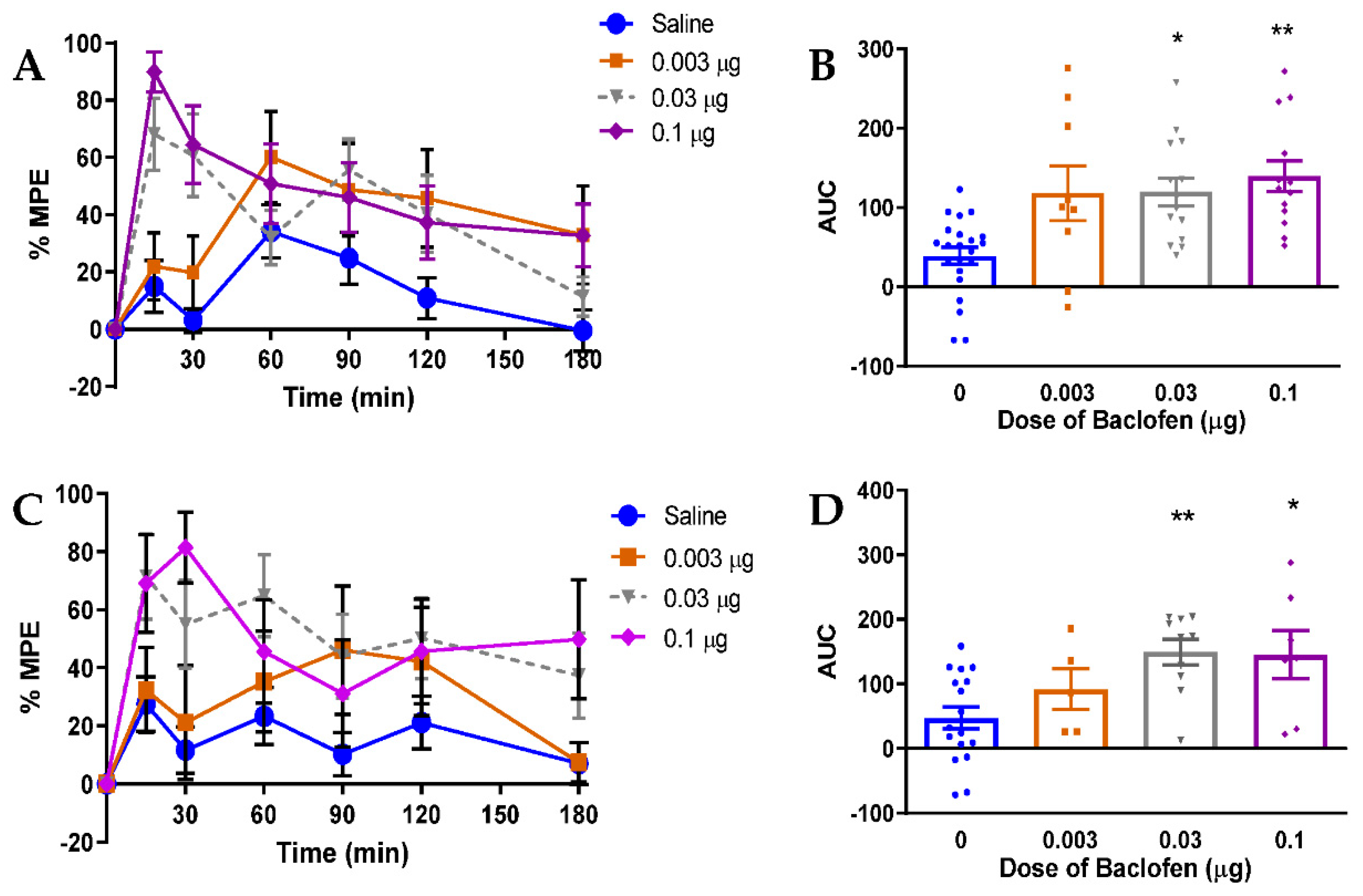
3.5. Relevance of GABAB Receptor-Mediated Inhibition to Neuropathic Pain in Rats
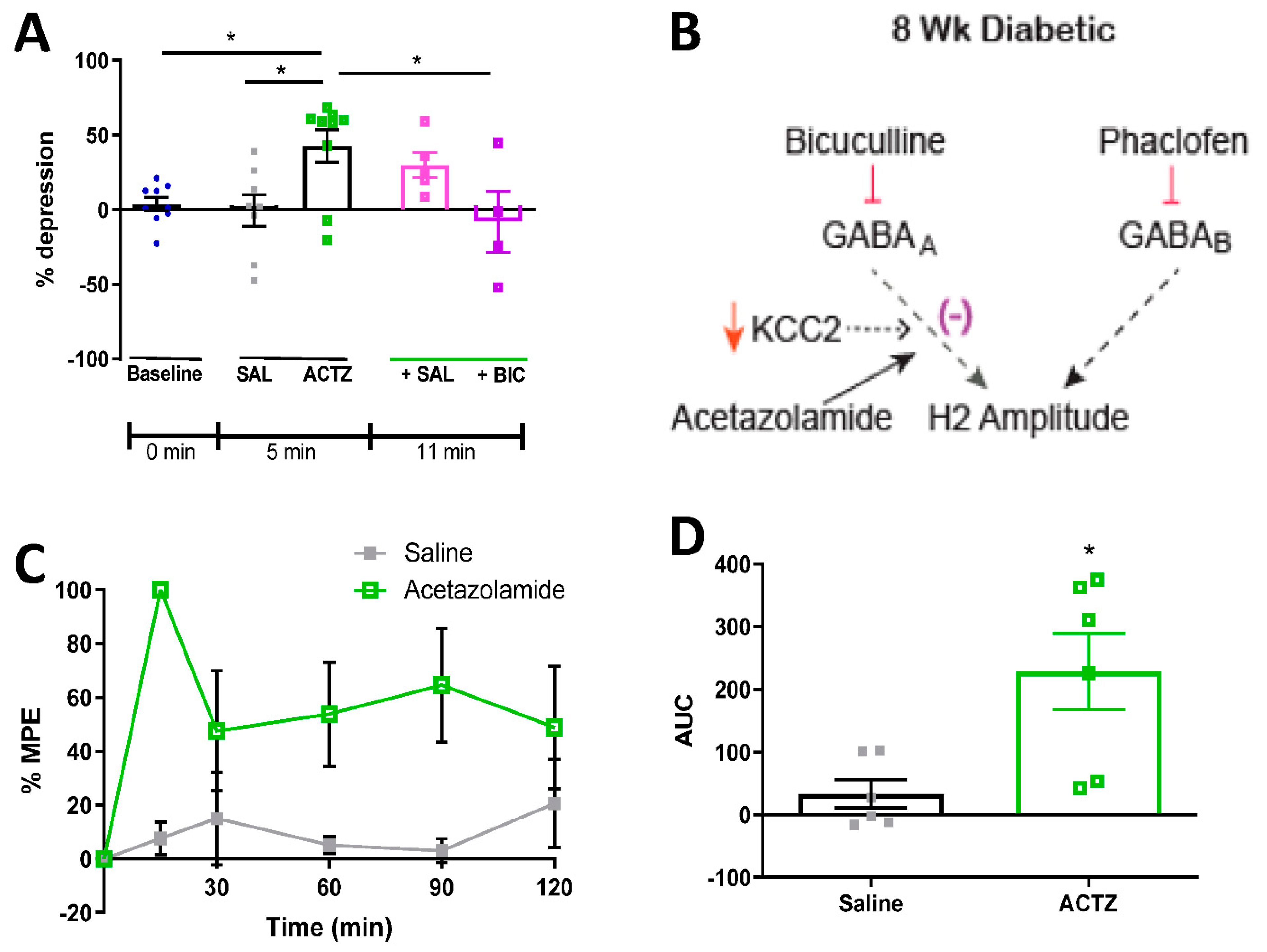
3.6. Acetazolamide Reverses RDD Deficits and Alleviates Tactile Allodynia in Diabetic Rats
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Woolf, C.J. Dissecting out mechanisms responsible for peripheral neuropathic pain: Implications for diagnosis and therapy. Life Sci. 2004, 74, 2605–2610. [Google Scholar] [CrossRef]
- Jensen, T.S.; Baron, R. Translation of symptoms and signs into mechanisms in neuropathic pain. Pain 2003, 102, 1–8. [Google Scholar] [CrossRef]
- Demant, D.T.; Lund, K.; Vollert, J.; Maier, C.; Segerdahl, M.; Finnerup, N.B.; Jensen, T.S.; Sindrup, S.H. The effect of oxcarbazepine in peripheral neuropathic pain depends on pain phenotype: A randomised, double-blind, placebo-controlled phenotype-stratified study. Pain 2014, 155, 2263–2273. [Google Scholar] [CrossRef]
- Demant, D.T.; Lund, K.; Finnerup, N.B.; Vollert, J.; Maier, C.; Segerdahl, M.S.; Jensen, T.S.; Sindrup, S.H. Pain relief with lidocaine 5% patch in localized peripheral neuropathic pain in relation to pain phenotype. Pain 2015, 156, 2234–2244. [Google Scholar] [CrossRef] [PubMed]
- Serra, J.; Duan, W.R.; Locke, C.R.; Solà, R.; Liu, W.; Nothaft, W. Effects of a T-type calcium channel blocker, ABT-639, on spontaneous activity in C-nociceptors in patients with painful diabetic neuropathy. Pain 2015, 156, 2175–2183. [Google Scholar] [CrossRef]
- Lee-Kubli, C.A.G.; Calcutt, N.A. Altered rate-dependent depression of the spinal H-reflex as an indicator of spinal disinhibition in models of neuropathic pain. Pain 2014, 155, 250–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolivalt, C.G.; Lee, C.A.; Ramos, K.M.; Calcutt, N.A. Allodynia and hyperalgesia in diabetic rats are mediated by GABA and depletion of spinal potassium-chloride co-transporters. Pain 2008, 140, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magladery, J.W. Some observations on spinal reflexes in man. Pflügers Arch. Gesamte Physiol. Menschen Tiere 1955, 261, 302–321. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.G.; Lee-Kubli, C.; Azmi, S.; Zhang, M.; Ferdousi, M.; Mixcoatl-Zecuatl, T.; Petropoulos, I.N.; Ponirakis, G.; Fineman, M.S.; Fadavi, H.; et al. Spinal disinhibition in experimental and clinical painful diabetic neuropathy. Diabetes 2017, 66, 1380–1390. [Google Scholar] [CrossRef] [Green Version]
- Mixcoatl-Zecuatl, T.; Jolivalt, C. A spinal mechanism of action for duloxetine in a rat model of painful diabetic neuropathy. Br. J. Pharmacol. 2011, 164, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, P. Beitrag zur Kenntnis der menschlichen Reflexe mit besonderer Berucksichtigung der elektrischen Erscheinungen. Arch. Anat. Physiol. 1910, 1, 223–246. [Google Scholar]
- Lloyd, D.P.C.; Wilson, V.J. Reflex depression in rhythmically active monosynaptic reflex pathways. J. Gen. Physiol. 1957, 40, 409–426. [Google Scholar] [CrossRef] [Green Version]
- Kakinohana, O.; Hefferan, M.; Nakamura, S.; Galik, J.; Tomori, Z.; Marsala, J.; Yaksh, T. Development of GABA-sensitive spasticity and rigidity in rats after transient spinal cord ischemia: A qualitative and quantitative electrophysiological and histopathological study. Neuroscience 2006, 141, 1569–1583. [Google Scholar] [CrossRef]
- Ishikawa, K.; Ott, K.; Porter, R.W.; Stuart, D. Low frequency depression of the H wave in normal and spinal man. Exp. Neurol. 1966, 15, 140–156. [Google Scholar] [CrossRef]
- Ho, S.M.; Waite, P.M. Effects of different anesthetics on the paired-pulse depression of the h reflex in adult rat. Exp. Neurol. 2002, 177, 494–502. [Google Scholar] [CrossRef] [Green Version]
- Boulenguez, P.; Liabeuf, S.; Bos, R.; Bras, H.; Jean-Xavier, C.; Brocard, C.; Stil, A.; Darbon, P.; Cattaert, D.; Delpire, E.; et al. Down-regulation of the potassium-chloride cotransporter KCC2 contributes to spasticity after spinal cord injury. Nat. Med. 2010, 16, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Thompson, F.J.; Reier, P.J.; Lucas, C.C.; Parmer, R. Altered patterns of reflex excitability subsequent to contusion injury of the rat spinal cord. J. Neurophysiol. 1992, 68, 1473–1486. [Google Scholar] [CrossRef]
- Toda, T.; Ishida, K.; Kiyama, H.; Yamashita, T.; Lee, S. Down-regulation of KCC2 expression and phosphorylation in motoneurons, and increases the number of in primary afferent projections to motoneurons in mice with post-stroke spasticity. PLoS ONE 2014, 9, e114328. [Google Scholar] [CrossRef] [PubMed]
- Reese, N.B.; Skinner, R.D.; Mitchell, D.; Yates, C.; Barnes, C.N.; Kiser, T.S.; Garcia-Rill, E. Restoration of frequency-dependent depression of the H-reflex by passive exercise in spinal rats. Spinal Cord 2005, 44, 28–34. [Google Scholar] [CrossRef]
- Malmberg, A.B.; O’Connor, W.T.; Glennon, J.C.; Ceseña, R.; Calcutt, N.A. Impaired formalin-evoked changes of spinal amino acid levels in diabetic rats. Brain Res. 2006, 1115, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zheng, J.; Xiong, L.; Zimmermann, M.; Yang, J. Spinal cord injury-induced attenuation of GABAergic inhibition in spinal dorsal horn circuits is associated with down-regulation of the chloride transporter KCC2 in rat. J. Physiol. 2008, 586, 5701–5715. [Google Scholar] [CrossRef]
- Price, T.J.; Cervero, F.; de Koninck, Y. Role of cation-chloride-cotransporters (CCC) in pain and hyperalgesia. Curr. Top. Med. Chem. 2005, 5, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Asiedu, M.; Ossipov, M.H.; Kaila, K.; Price, T.J. Acetazolamide and midazolam act synergistically to inhibit neuropathic pain. Pain 2010, 148, 302–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaksh, T.L.; Rudy, T.A. Chronic catheterization of the spinal subarachnoid space. Physiol. Behav. 1976, 17, 1031–1036. [Google Scholar] [CrossRef]
- Calcutt, N.A.; Chaplan, S.R. Spinal pharmacology of tactile allodynia in diabetic rats. Br. J. Pharmacol. 1997, 122, 1478–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaplan, S.; Bach, F.; Pogrel, J.; Chung, J.; Yaksh, T. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Bowery, N.; Hudson, A.; Price, G. GABAA and GABAB receptor site distribution in the rat central nervous system. Neuroscience 1987, 20, 365–383. [Google Scholar] [CrossRef]
- Malan, T.P.; Mata, H.P.; Porreca, F. Spinal GABA(A) and GABA(B) receptor pharmacology in a rat model of neuropathic pain. Anesthesiology 2002, 96, 1161–1167. [Google Scholar] [CrossRef]
- Asiedu, M.N.; Mejia, G.L.; Hübner, C.A.; Kaila, K.; Price, T.J. Inhibition of carbonic anhydrase augments GABAA receptor-mediated analgesia via a spinal mechanism of action. J. Pain 2014, 15, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y.; Prescott, S.A. Chloride dysregulation and inhibitory receptor blockade yield equivalent disinhibition of spinal neurons yet are differentially reversed by carbonic anhydrase blockade. Pain 2015, 156, 2431–2437. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T. Carbonic anhydrase inhibition and the management of neuropathic pain. Expert Rev. Neurother. 2016, 16, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Hernández, M.; Corleto, J.A.; Barragán-Iglesias, P.; González-Ramírez, R.; Pineda-Farias, J.B.; Felix, R.; Calcutt, N.A.; Delgado-Lezama, R.; Marsala, M.; Granados-Soto, V. The α5 subunit containing GABAA receptors contribute to chronic pain. Pain 2016, 157, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Stuart, G.J.; Redman, S.J. The role of GABAA and GABAB receptors in presynaptic inhibition of Ia EPSPs in cat spinal motoneurones. J. Physiol. 1992, 447, 675–692. [Google Scholar] [CrossRef] [PubMed]
- Hübner, C.A.; Stein, V.; Hermans-Borgmeyer, I.; Meyer, T.; Ballanyi, K.; Jentsch, T.J. Disruption of KCC2 reveals an essential role of K-Cl cotransport already in early synaptic inhibition. Neuron 2001, 30, 515–524. [Google Scholar] [CrossRef] [Green Version]
- Kahle, K.T.; Rinehart, J.; Lifton, R.P. Phosphoregulation of the Na–K–2Cl and K–Cl cotransporters by the WNK kinases. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2010, 1802, 1150–1158. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.H.C.; Walker, J.A.; Williams, J.R.; Goodier, R.J.; Payne, J.A.; Moss, S.J. Direct protein kinase c-dependent phosphorylation regulates the cell surface stability and activity of the potassium chloride cotransporter KCC2. J. Biol. Chem. 2007, 282, 29777–29784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wake, H.; Watanabe, M.; Moorhouse, A.J.; Kanematsu, T.; Horibe, S.; Matsukawa, N.; Asai, K.; Ojika, K.; Hirata, M.; Nabekura, J. Early changes in KCC2 phosphorylation in response to neuronal stress result in functional downregulation. J. Neurosci. 2007, 27, 1642–1650. [Google Scholar] [CrossRef] [Green Version]
- Rivera, C.; Voipio, J.; Thomas-Crusells, J.; Li, H.; Emri, Z.; Sipilä, S.; Payne, J.A.; Minichiello, L.; Saarma, M.; Kaila, K. Mechanism of activity-dependent downregulation of the neuron-specific K-Cl cotransporter KCC2. J. Neurosci. 2004, 24, 4683–4691. [Google Scholar] [CrossRef] [Green Version]
- Drew, G.M.; Siddall, P.J.; Duggan, A.W. Mechanical allodynia following contusion injury of the rat spinal cord is associated with loss of GABAergic inhibition in the dorsal horn. Pain 2004, 109, 379–388. [Google Scholar] [CrossRef]
- Vikman, K.S.; Duggan, A.W.; Siddall, P.J. Interferon-γ induced disruption of GABAergic inhibition in the spinal dorsal horn in vivo. Pain 2007, 133, 18–28. [Google Scholar] [CrossRef]
- Huang, Y.-J.; Grau, J.W. Ionic plasticity and pain: The loss of descending serotonergic fibers after spinal cord injury transforms how GABA affects pain. Exp. Neurol. 2018, 306, 105–116. [Google Scholar] [CrossRef]
- Wang, X.-L.; Zhang, H.-M.; Chen, S.-R.; Pan, H.-L. Altered synaptic input and GABABreceptor function in spinal superficial dorsal horn neurons in rats with diabetic neuropathy. J. Physiol. 2007, 579, 849–861. [Google Scholar] [CrossRef]
- Wang, X.-L.; Zhang, Q.; Zhang, Y.-Z.; Liu, Y.-T.; Dong, R.; Wang, Q.-J.; Guo, Y.-X. Downregulation of GABAB receptors in the spinal cord dorsal horn in diabetic neuropathy. Neurosci. Lett. 2011, 490, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Floeter, M.K.; Kohn, A.F. H-reflexes of different sizes exhibit differential sensitivity to low frequency depression. Electroencephalogr. Clin. Neurophysiol. Electromyogr. Mot. Control 1997, 105, 470–475. [Google Scholar] [CrossRef]
- Liu, P.; Guo, W.-Y.; Zhao, X.-N.; Bai, H.-P.; Wang, Q.; Wang, X.-L.; Zhang, Y.-Z. Intrathecal baclofen, a GABAB receptor agonist, inhibits the expression of p-CREB and NR2B in the spinal dorsal horn in rats with diabetic neuropathic pain. Can. J. Physiol. Pharmacol. 2014, 92, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Malcangio, M.; Tomlinson, D.R. A pharmacologic analysis of mechanical hyperalgesia in streptozotocin/diabetic rats. Pain 1998, 76, 151–157. [Google Scholar] [CrossRef]
- Fox, A.; Eastwood, C.; Gentry, C.; Manning, D.; Urban, L. Critical evaluation of the streptozotocin model of painful diabetic neuropathy in the rat. Pain 1999, 81, 307–316. [Google Scholar] [CrossRef]
- Liu, P.; Yuan, H.-B.; Zhao, S.; Liu, F.-F.; Jiang, Y.-Q.; Guo, Y.-X.; Wang, X.-L. Activation of GABAB receptor suppresses diabetic neuropathic pain through toll-like receptor 4 signaling pathway in the spinal dorsal horn. Mediat. Inflamm. 2018, 2018, 6016272. [Google Scholar] [CrossRef] [Green Version]
- Angeli, A.; Mannelli, L.D.C.; Ghelardini, C.; Peat, T.S.; Bartolucci, G.; Menicatti, M.; Carta, F.; Supuran, C.T. Benzensulfonamides bearing spyrohydantoin moieties act as potent inhibitors of human carbonic anhydrases II and VII and show neuropathic pain attenuating effects. Eur. J. Med. Chem. 2019, 177, 188–197. [Google Scholar] [CrossRef]
- Bozdag, M.; Poli, G.; Angeli, A.; Lucarini, E.; Tuccinardi, T.; Mannelli, L.D.C.; Selleri, S.; Ghelardini, C.; Winum, J.-Y.; Carta, F.; et al. N-aryl-N′-ureido-O-sulfamates: Potent and selective inhibitors of the human Carbonic Anhydrase VII isoform with neuropathic pain relieving properties. Bioorg. Chem. 2019, 89, 103033. [Google Scholar] [CrossRef] [PubMed]
- Carta, F.; Mannelli, L.D.C.; Pinard, M.; Ghelardini, C.; Scozzafava, A.; McKenna, R.; Supuran, C.T. A class of sulfonamide carbonic anhydrase inhibitors with neuropathic pain modulating effects. Bioorg. Med. Chem. 2015, 23, 1828–1840. [Google Scholar] [CrossRef] [PubMed]
- Vali, Y.K.; Gundla, R.; Singh, O.V.; Tamboli, Y.; Manelli, L.D.C.; Ghelardini, C.; Al-Tamimi, A.-M.S.; Carta, F.; Angeli, A.; Supuran, C.T. Spirocyclic sulfonamides with carbonic anhydrase inhibitory and anti-neuropathic pain activity. Bioorg. Chem. 2019, 92, 103210. [Google Scholar] [CrossRef] [PubMed]
- Potenzieri, A.; Riva, B.; Rigolio, R.; Chiorazzi, A.; Pozzi, E.; Ballarini, E.; Cavaletti, G.; Genazzani, A.A. Oxaliplatin-induced neuropathy occurs through impairment of haemoglobin proton buffering and is reversed by carbonic anhydrase inhibitors. Pain 2020, 161, 405–415. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee-Kubli, C.A.; Zhou, X.; Jolivalt, C.G.; Calcutt, N.A. Pharmacological Modulation of Rate-Dependent Depression of the Spinal H-Reflex Predicts Therapeutic Efficacy against Painful Diabetic Neuropathy. Diagnostics 2021, 11, 283. https://doi.org/10.3390/diagnostics11020283
Lee-Kubli CA, Zhou X, Jolivalt CG, Calcutt NA. Pharmacological Modulation of Rate-Dependent Depression of the Spinal H-Reflex Predicts Therapeutic Efficacy against Painful Diabetic Neuropathy. Diagnostics. 2021; 11(2):283. https://doi.org/10.3390/diagnostics11020283
Chicago/Turabian StyleLee-Kubli, Corinne A., XiaJun Zhou, Corinne G. Jolivalt, and Nigel A. Calcutt. 2021. "Pharmacological Modulation of Rate-Dependent Depression of the Spinal H-Reflex Predicts Therapeutic Efficacy against Painful Diabetic Neuropathy" Diagnostics 11, no. 2: 283. https://doi.org/10.3390/diagnostics11020283