
Comparison of Tissue Factors in the Ontogenetic Aspects of Human Cholesteatoma



Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Samples
2.2. Immunohistochemical Analysis
2.3. Statistical Analysis
3. Results
3.1. Description of the Analyzed Tissue
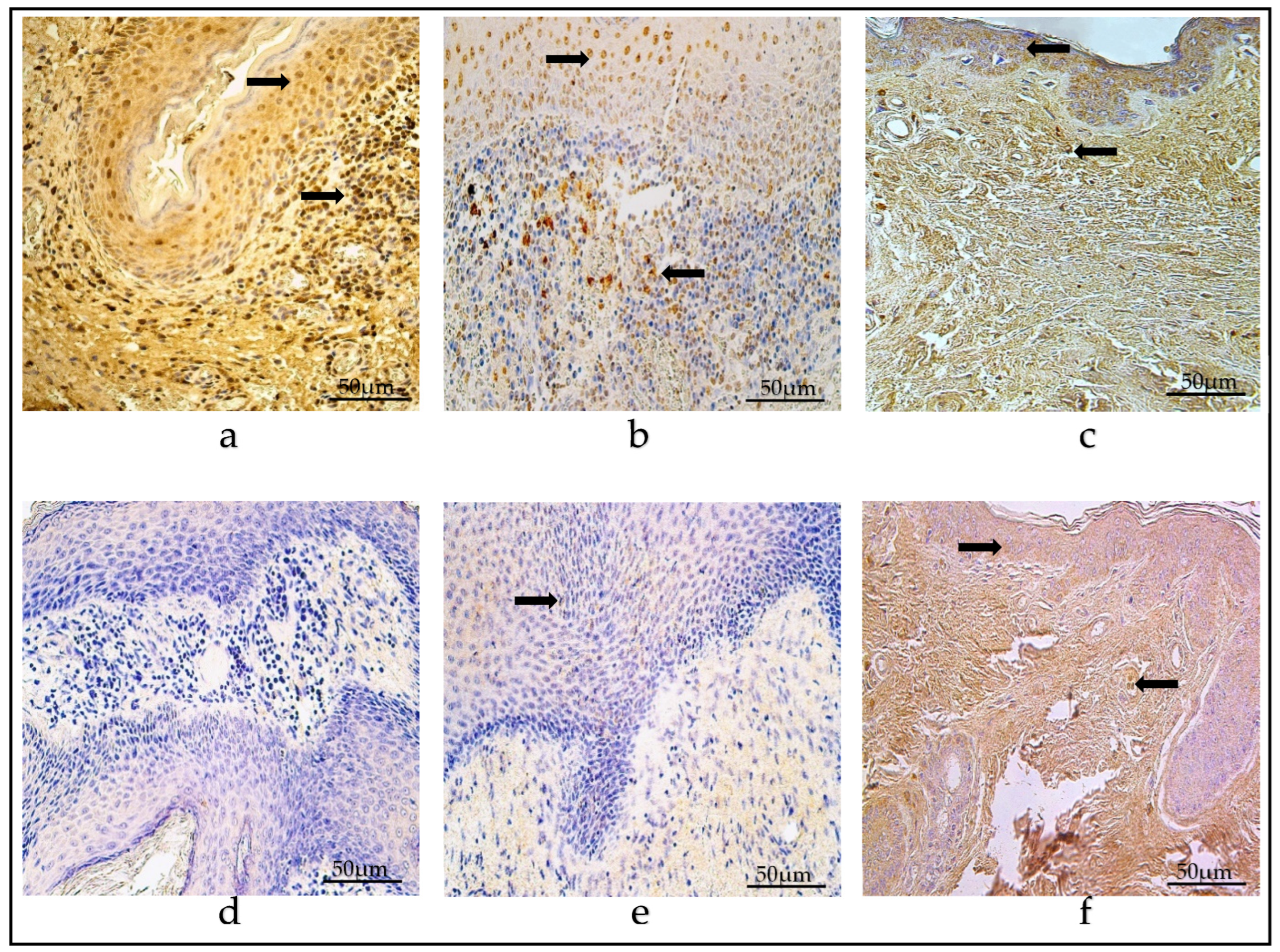
3.2. Immunohistochemistry of Defensins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | HβD-2 | HβD-4 | IL-1α | IL-10 | Ki-67 | NF-κβ | VEGF | SHH | MMP-2 | MMP-9 | TIMP-2 | TIMP-4 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | P | M | P | M | P | M | P | M | P | M | P | M | P | M | P | M | P | M | P | M | P | M | P | |
| Children | +/++ | + | 0/+-+ | 0/+-+ | +/++ | + | +/++ | + | 0/+ | 0/+-+ | ++ | + | ++ | 0/+-+ | ++ | +/++ | +/++ | + | 0/+-+ | 0/+ | + | 0/+ | ++/+++ | ++ |
| Adults | +/++ | + | + | 0/+ | +/++ | + | +/++ | + | 0/+ | 0/+ | ++ | + | +/++ | 0/+-+ | ++/+++ | +/++ | +/++ | + | 0/+ | 0/+ | + | 0/+ | ++/+++ | ++ |
| Control | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT | E | CT |
| + | 0-0/+ | + | 0/+ | 0/+-+ | 0/+-+ | +/++ | +/++ | 0-0/+ | 0-0/+ | 0/+ | 0/+-+ | ++/+++ | 0/+-+ | +/++ | 0/+-+ | + | + | + | 0/+-+ | +/++ | 0/+-+ | ++ | +/++ | |
3.3. Immunohistochemistry of Cytokines
3.4. Ki-67 Immunohistochemistry
3.5. NF-κβ Immunohistochemistry
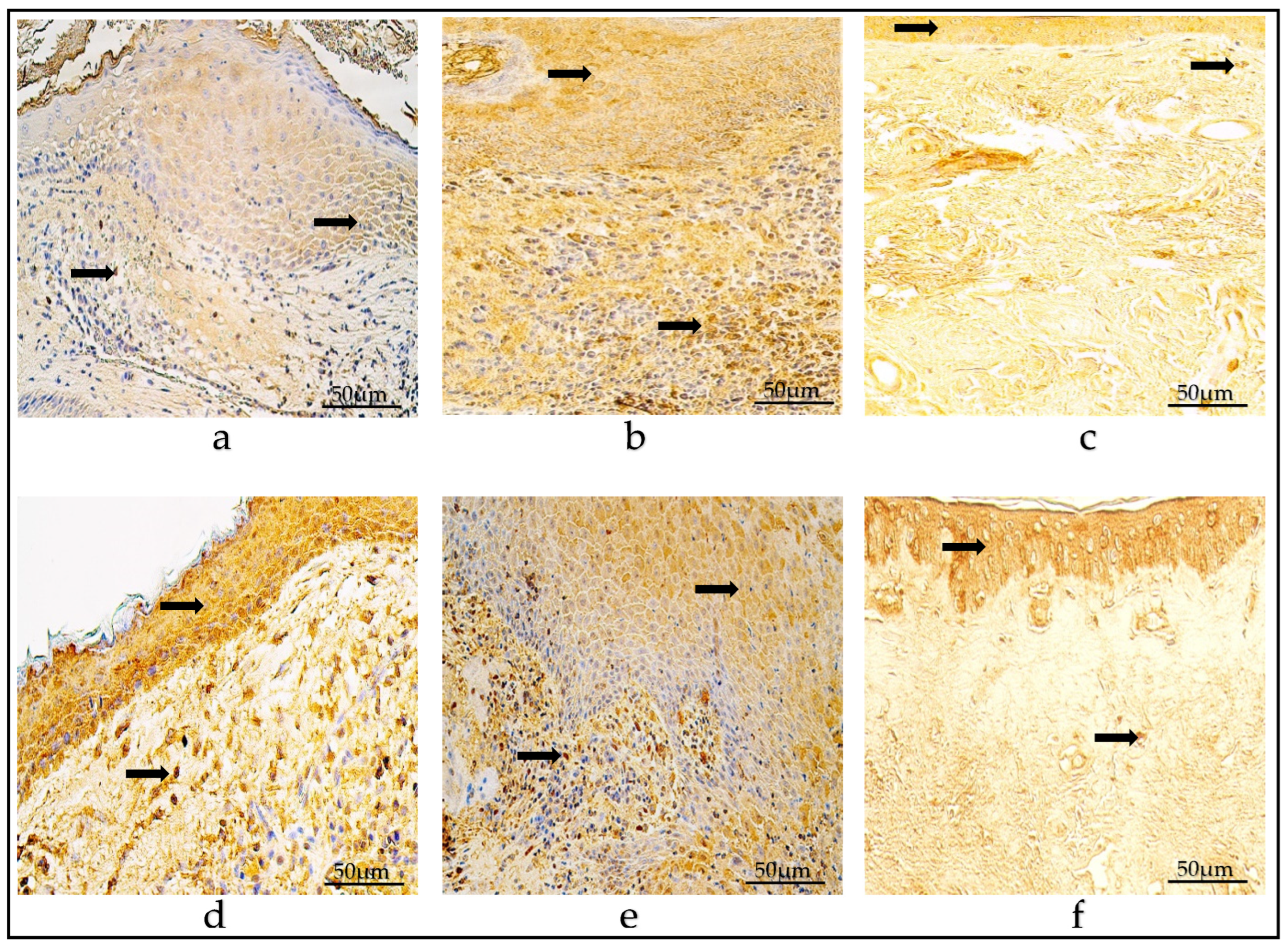
3.6. VEGF Immunohistochemistry
3.7. SHH Immunohistochemistry
3.8. Immunohistochemistry of Tissue-Remodeling Factors
3.9. Statistical Comparison between the Groups
| Detected Factor | Kruskal–Wallis Test | p-Value | |
|---|---|---|---|
| Children | Adults | ||
| HβD-2 matrix | HβD-2 matrix | 0.587 | >0.999 |
| HβD-2 perimatrix | HβD-2 perimatrix | 0.480 | >0.999 |
| HβD-4 matrix | HβD-4 matrix | 0.868 | >0.999 |
| HβD-4 perimatrix | HβD-4 perimatrix | −0.719 | >0.999 |
| IL-1α matrix | IL-1α matrix | −0.039 | >0.999 |
| IL-1α perimatrix | IL-1α perimatrix | 0.717 | >0.999 |
| IL-10 matrix | IL-10 matrix | 0.043 | >0.999 |
| IL-10 perimatrix | IL-10 perimatrix | 0.868 | >0.999 |
| Ki-67 matrix | Ki-67 matrix | 0.886 | >0.999 |
| Ki-67 perimatrix | Ki-67 perimatrix | 0.908 | >0.999 |
| NF-κβ matrix | NF-κβ matrix | −0.079 | >0.999 |
| NF-κβ perimatrix | NF-κβ perimatrix | 0.039 | >0.999 |
| VEGF matrix | VEGF matrix | −0.694 | >0.999 |
| VEGF perimatrix | VEGF perimatrix | 0.486 | >0.999 |
| SHH matrix | SHH matrix | 0.482 | >0.999 |
| SHH perimatrix | SHH perimatrix | 0.363 | >0.999 |
| MMP-2 matrix | MMP-2 matrix | 0.415 | >0.999 |
| MMP-2 perimatrix | MMP-2 perimatrix | 0.986 | 0.972 |
| MMP-9 matrix | MMP-9 matrix | −0.365 | >0.999 |
| MMP-9 perimatrix | MMP-9 perimatrix | 1.290 | 0.591 |
| TIMP-2 matrix | TIMP-2 matrix | 0.576 | >0.999 |
| TIMP-2 perimatrix | TIMP-2 perimatrix | 0.958 | >0.999 |
| TIMP-4 matrix | TIMP-4 matrix | −0.205 | >0.999 |
| TIMP-4 perimatrix | TIMP-4 perimatrix | 0.159 | >0.999 |
| Detected Factor | Kruskal–Wallis Test | p-Value |
|---|---|---|
| Adult HβD-2 P vs. Control HβD-2 CT | 2.815 | 0.015 |
| Children HβD-2 P vs. Control HβD-2 CT | 2.498 | 0.038 |
| Children HβD-4 M vs. Control HβD-4 E | −2.132 | 0.099 * |
| Adult Ki-67 M vs. Control Ki-67 E | 3.697 | 0.001 |
| Children Ki-67 M vs. Control Ki-67 E | 3.110 | 0.006 |
| Adult Ki-67 P vs. Control Ki-67 CT | 2.577 | 0.030 |
| Adult NF-κβ M vs. Control NF-κβ E | 2.864 | 0.013 |
| Children NF-κβ M vs. Control NF-κβ E | 2.915 | 0.011 |
| Adult VEGF M vs. Control VEGF E | −2.146 | 0.096 * |
| Adult SHH P vs. Control SHH CT | 3.146 | 0.005 |
| Children SHH P vs. Control SHH CT | 2.906 | 0.011 |
| Children TIMP-2 M vs. Control TIMP-2 E | -2.207 | 0.082 * |
3.10. Correlations between Tissue Factors in Patient Groups
| Factor 1 | Factor 2 | Spearman’s Correlation Coefficient; p Value | |
|---|---|---|---|
| MMP-2 matrix | MMP-2 perimatrix | Children r = 0.803; p = 0.000 | Adult r = 0.574; p = 0.003 |
| MMP-2 matrix | TIMP-2 matrix | Children r = 0.622; p = 0.001 | Adult r = 0.484; p = 0.014 |
| MMP-2 matrix | SHH matrix | Children r = 0.786; p = 0.000 | Adult r = 0.719; p = 0.000 |
| MMP-2 matrix | NF-κβ matrix | Children r = 0.677; p = 0.000 | Adult r = 0.399; p = 0.048 |
| MMP-2 perimatrix | SHH matrix | Children r = 0.786; p = 0.000 | Adult r = 0.453; p = 0.023 |
| MMP-2 perimatrix | SHH perimatrix | Children r = 0.653; p = 0.000 | Adult r = 0.460; p = 0.021 |
| MMP-9 matrix | IL-1α matrix | Children r = 0.549; p = 0.004 | Adult r = 0.426; p = 0.034 |
| MMP-9 matrix | IL-10 matrix | Children r = 0.418; p = 0.038 | Adult r = 0.458; p = 0.021 |
| MMP-9 perimatrix | TIMP-4 perimatrix | Children r = 0.490; p = 0.013 | Adult r = 0.664; p = 0.000 |
| MMP-9 perimatrix | IL-1α matrix | Children r = 0.642; p = 0.001 | Adult r = 0.435; p = 0.030 |
| MMP-9 perimatrix | IL-1α perimatrix | Children r = 0.714; p = 0.000 | Adult r = 0.608; p = 0.001 |
| MMP-9 perimatrix | IL-10 perimatrix | Children r = 0.468; p = 0.018 | Adult r = 0.601; p = 0.001 |
| MMP-9 perimatrix | NF-κβ perimatrix | Children r = 0.614; p = 0.001 | Adult r = 0.790; p = 0.000 |
| MMP-9 perimatrix | Ki-67 perimatrix | Children r = 0.624; p = 0.001 | Adult r = 0.677; p = 0.000 |
| MMP-9 perimatrix | HβD-2 matrix | Children r = 0.487; p = 0.014 | Adult r = 0.464; p = 0.019 |
| TIMP-2 matrix | TIMP-2 perimatrix | Children r = 0.685; p = 0.000 | Adult r = 0.676; p = 0.000 |
| TIMP-2 matrix | SHH matrix | Children r = 0.537; p = 0.006 | Adult r = 0.478; p = 0.016 |
| TIMP-2 matrix | NF-κβ matrix | Children r = 0.504; p = 0.010 | Adult r = 0.473; p = 0.017 |
| TIMP-2 matrix | HβD-2 matrix | Children r = 0.505; p = 0.010 | Adult r = 0.416; p = 0.038 |
| TIMP-2 perimatrix | IL-1α matrix | Children r = 0.457; p = 0.022 | Adult r = 0.505; p = 0.010 |
| TIMP-2 perimatrix | IL-10 matrix | Children r = 0.423; p = 0.035 | Adult r = 0.630; p = 0.001 |
| TIMP-4 matrix | TIMP-4 perimatrix | Children r = 0.841; p = 0.000 | Adult r = 0.431; p = 0.031 |
| TIMP-4 matrix | SHH matrix | Children r = 0.681; p = 0.000 | Adult r = 0.436; p = 0.029 |
| TIMP-4 matrix | NF-κβ matrix | Children r = 0.738; p = 0.000 | Adult r = 0.540; p = 0.005 |
| TIMP-4 perimatrix | IL-1α matrix | Children r = 0.641; p = 0.001 | Adult r = 0.457; p = 0.022 |
| TIMP-4 perimatrix | IL-1α perimatrix | Children r = 0.663; p = 0.000 | Adult r = 0.637; p = 0.001 |
| TIMP-4 perimatrix | IL-10 matrix | Children r = 0.638; p = 0.001 | Adult r = 0.425; p = 0.034 |
| TIMP-4 perimatrix | IL-10 perimatrix | Children r = 0.721; p = 0.000 | Adult r = 0.638; p = 0.001 |
| TIMP-4 perimatrix | NF-κβ matrix | Children r = 0.654; p = 0.000 | Adult r = 0.422; p = 0.036 |
| TIMP-4 perimatrix | NF-κβ perimatrix | Children r = 0.457; p = 0.022 | Adult r = 0.749; p = 0.000 |
| TIMP-4 perimatrix | Ki-67 matrix | Children r = 0.481; p = 0.015 | Adult r = 0.402; p = 0.046 |
| TIMP-4 perimatrix | Ki-67 perimatrix | Children r = 0.414; p = 0.040 | Adult r = 0.651; p = 0.000 |
| TIMP-4 perimatrix | HβD-2 matrix | Children r = 0.568; p = 0.003 | Adult r = 0.404; p = 0.045 |
| TIMP-4 perimatrix | HβD-2 perimatrix | Children r = 0.397; p = 0.049 | Adult r = 0.545; p = 0.005 |
| SHH matrix | NF-κβ matrix | Children r = 0.753; p = 0.000 | Adult r = 0.549; p = 0.005 |
| SHH perimatrix | Ki-67 matrix | Children r = 0.746; p = 0.000 | Adult r = 0.646; p = 0.000 |
| SHH perimatrix | HβD-2 perimatrix | Children r = 0.428; p = 0.033 | Adult r = 0.527; p = 0.007 |
| IL-1α matrix | IL-1α perimatrix | Children r = 0.716; p = 0.000 | Adult r = 0.557; p = 0.004 |
| IL-1α matrix | IL-10 matrix | Children r = 0.709; p = 0.000 | Adult r = 0.813; p = 0.000 |
| IL-1α matrix | IL-10 perimatrix | Children r = 0.720; p = 0.000 | Adult r = 0.762; p = 0.000 |
| IL-1α matrix | NF-κβ perimatrix | Children r = 0.406; p = 0.044 | Adult r = 0.519; p = 0.008 |
| IL-1α matrix | HβD-2 matrix | Children r = 0.700; p = 0.000 | Adult r = 0.827; p = 0.000 |
| IL-1α perimatrix | IL-10 perimatrix | Children r = 0.694; p = 0.000 | Adult r = 0.640; p = 0.001 |
| IL-1α perimatrix | NF-κβ perimatrix | Children r = 0.510; p = 0.009 | Adult r = 0.692; p = 0.000 |
| IL-1α perimatrix | Ki-67 perimatrix | Children r = 0.441; p = 0.027 | Adult r = 0.583; p = 0.002 |
| IL-1α perimatrix | HβD-2 matrix | Children r = 0.630; p = 0.001 | Adult r = 0.499; p = 0.011 |
| IL-10 matrix | IL-10 perimatrix | Children r = 0.668; p = 0.000 | Adult r = 0.801; p = 0.000 |
| IL-10 matrix | NF-κβ perimatrix | Children r = 0.536; p = 0.006 | Adult r = 0.554; p = 0.004 |
| IL-10 matrix | VEGF matrix | Children r = 0.559; p = 0.004 | Adult r = 0.611; p = 0.001 |
| IL-10 matrix | HβD-2 matrix | Children r = 0.828; p = 0.000 | Adult r = 0.841; p = 0.000 |
| IL-10 matrix | HβD-2 perimatrix | Children r = 0.677; p = 0.000 | Adult r = 0.462; p = 0.020 |
| IL-10 perimatrix | NF-κβ matrix | Children r = 0.602; p = 0.001 | Adult r = 0.396; p = 0.050 |
| IL-10 perimatrix | VEGF matrix | Children r = 0.696; p = 0.000 | Adult r = 0.687; p = 0.000 |
| IL-10 perimatrix | HβD-2 matrix | Children r = 0.592; p = 0.002 | Adult r = 0.686; p = 0.000 |
| IL-10 perimatrix | HβD-2 perimatrix | Children r = 0.516; p = 0.008 | Adult r = 0.687; p = 0.000 |
| NF-κβ matrix | VEGF matrix | Children r = 0.595; p = 0.002 | Adult r = 0.414; p = 0.039 |
| NF-κβ matrix | HβD-2 matrix | Children r = 0.750; p = 0.000 | Adult r = 0.418; p = 0.038 |
| NF-κβ perimatrix | Ki-67 perimatrix | Children r = 0.494; p = 0.012 | Adult r = 0.571; p = 0.003 |
| NF-κβ perimatrix | VEGF matrix | Children r = 0.637; p = 0.001 | Adult r = 0.474; p = 0.017 |
| NF-κβ perimatrix | HβD-2 matrix | Children r = 0.621; p = 0.001 | Adult r = 0.526; p = 0.007 |
| NF-κβ perimatrix | HβD-2 perimatrix | Children r = 0.635; p = 0.001 | Adult r = 0.748; p = 0.000 |
| Ki-67 matrix | HβD-2 perimatrix | Children r = 0.520; p = 0.008 | Adult r = 0.498; p = 0.011 |
| VEGF matrix | VEGF perimatrix | Children r = 0.745; p = 0.000 | Adult r = 0.429; p = 0.032 |
| VEGF matrix | HβD-2 perimatrix | Children r = 0.509; p = 0.009 | Adult r = 0.413; p = 0.040 |
| HβD-2 matrix | HβD-2 perimatrix | Children r = 0.748; p = 0.000 | Adult r = 0.477; p = 0.016 |
4. Discussion
4.1. Human Beta Defensins
4.2. Pro- and Anti-Inflammatory Cytokines
4.3. Proliferation Marker Ki-67
4.4. Transcription Factor NF-κβ
4.5. Angiogenetic Factor
4.6. Sonic Hedgehog
4.7. Remodeling Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhutta, M.F.; Williamson, I.G.; Sudhoff, H.H. Cholesteatoma. BMJ 2011, 342, d1088. [Google Scholar] [CrossRef]
- Kuo, C.L.; Shiao, A.S.; Yung, M.; Sakagami, M.; Sudhoff, H.; Wang, C.H.; Hsu, C.H.; Lien, C.F. Updates and knowledge gaps in cholesteatoma research. Biomed. Res. Int. 2015, 2015, 854024. [Google Scholar] [CrossRef]
- Britze, A.; Møller, M.L.; Ovesen, T. Incidence, 10-year recidivism rate and prognostic factors for cholesteatoma. J. Laryngol. Otol. 2017, 131, 319–328. [Google Scholar] [CrossRef]
- Olszewska, E.; Wagner, M.; Bernal-Sprekelsen, M.; Ebmeyer, J.; Dazert, S.; Hildmann, H.; Sudhoff, H. Etiopathogenesis of cholesteatoma. Eur. Arch. Otorhinolaryngol. 2004, 261, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Harder, J.; Bartels, J.; Christophers, E.; Schröder, J.M. A peptide antibiotic from human skin. Nature 1997, 387, 861. [Google Scholar] [CrossRef] [PubMed]
- Schröder, J.M.; Harder, J. Human beta-defensin-2. Int. J. Biochem. Cell Biol. 1999, 31, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Cieślik, M.; Bagińska, N.; Górski, A.; Jończyk-Matysiak, E. Human β-Defensin 2 and Its Postulated Role in Modulation of the Immune Response. Cells 2021, 10, 2991. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Abiko, Y.; Kusano, K.; Yamazaki, M.; Saitoh, M.; Mizoguchi, I.; Jinbu, Y.; Noguchi, T.; Kaku, T. Localization of human beta-defensin 3 mRNA in normal oral epithelium, leukoplakia, and lichen planus: An in situ hybridization study. Med. Electron. Microsc. 2003, 36, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Song, J.J.; Chae, S.W.; Woo, J.S.; Lee, H.M.; Jung, H.H.; Hwang, S.J. Differential expression of human beta defensin 2 and human beta defensin 3 in human middle ear cholesteatoma. Ann. Otol. Rhinol. Laryngol. 2007, 116, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Moon, S.K.; Choung, Y.H.; Choi, H.S. Expression of beta-defensins in human middle ear cholesteatoma. Acta Otolaryngol. 2003, 123, 236–240. [Google Scholar] [CrossRef] [PubMed]
- García, J.R.; Krause, A.; Schulz, S.; Rodríguez-Jiménez, F.J.; Klüver, E.; Adermann, K.; Forssmann, U.; Frimpong-Boateng, A.; Bals, R.; Forssmann, W.G. Human beta-defensin 4: A novel inducible peptide with a specific salt-sensitive spectrum of antimicrobial activity. FASEB J. 2001, 15, 1819–1821. [Google Scholar] [CrossRef]
- Dambergs, K.; Sumeraga, G.; Pilmane, M. Complex Evaluation of Tissue Factors in Pediatric Cholesteatoma. Children 2021, 8, 926. [Google Scholar] [CrossRef]
- Dambergs, K.; Sumeraga, G.; Pilmane, M. Morphopathogenesis of Adult Acquired Cholesteatoma. Medicina 2023, 59, 306. [Google Scholar] [CrossRef]
- Schürmann, M.; Goon, P.; Sudhoff, H. Review of potential medical treatments for middle ear cholesteatoma. Cell Commun. Signal. 2022, 20, 148. [Google Scholar] [CrossRef]
- Dinarello, C.A. The interleukin-1 family: 10 years of discovery. FASEB J. 1994, 8, 1314–1325. [Google Scholar] [CrossRef]
- Bujía, J.; Kim, C.; Ostos, P.; Sudhoff, H.; Kastenbauer, E.; Hültner, L. Interleukin 1 (IL-1) and IL-1-receptor antagonist (IL-1-RA) in middle ear cholesteatoma: An analysis of protein production and biological activity. Eur. Arch. Otorhinolaryngol. 1996, 253, 252–255. [Google Scholar] [CrossRef]
- Lee, Y.M.; Fujikado, N.; Manaka, H.; Yasuda, H.; Iwakura, Y. IL-1 plays an important role in the bone metabolism under physiological conditions. Int. Immunol. 2010, 22, 805–816. [Google Scholar] [CrossRef]
- Mosser, D.M.; Zhang, X. Interleukin-10: New perspectives on an old cytokine. Immunol. Rev. 2008, 226, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Sabat, R.; Grütz, G.; Warszawska, K.; Kirsch, S.; Witte, E.; Wolk, K.; Geginat, J. Biology of interleukin-10. Cytokine Growth Factor Rev. 2010, 21, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Sabat, R.; Krätzschmar, J.; Seidel, H.; Wolk, K.; Schönbein, C.; Schütt, S.; Friedrich, M.; Döcke, W.D.; Asadullah, K.; et al. Expression profiling of IL-10-regulated genes in human monocytes and peripheral blood mononuclear cells from psoriatic patients during IL-10 therapy. Eur. J. Immunol. 2004, 34, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Kuczkowski, J.; Sakowicz-Burkiewicz, M.; Iżycka-Świeszewska, E.; Mikaszewski, B.; Pawełczyk, T. Expression of tumor necrosis factor-α, interleukin-1α, interleukin-6 and interleukin-10 in chronic otitis media with bone osteolysis. ORL J. Otorhinolaryngol. Relat. Spec. 2011, 73, 93–99. [Google Scholar] [CrossRef]
- Yeşilova, M.; Görür, K.; Ismi, O.; Özcan, C.; Büyükafşar, K. The Role of Rho/Rho-Kinase Pathway in the Pathogenesis of Cholesteatoma. Otol. Neurotol. 2017, 38, 516–520. [Google Scholar] [CrossRef]
- Araz Server, E.; Kalaycık Ertugay, Ç.; Baykal Koca, S.; Longur, E.S.; Yiğit, Ö.; Demirhan, H.; Çakır, Y. Predictive Role of Ki-67 and Proliferative-Cell Nuclear Antigen (PCNA) in Recurrent Cholesteatoma. J. Int. Adv. Otol. 2019, 15, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell Physiol. 2000, 182, 311–322. [Google Scholar] [CrossRef]
- Hamed, M.A.; Nakata, S.; Shiogama, K.; Suzuki, K.; Sayed, R.H.; Nishimura, Y.; Iwata, N.; Sakurai, K.; Badawy, B.S.; Inada, K.I.; et al. Cytokeratin 13, Cytokeratin 17, and Ki-67 Expression in Human Acquired Cholesteatoma and Their Correlation with Its Destructive Capacity. Clin. Exp. Otorhinolaryngol. 2017, 10, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-κB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, C.; Bucci, I.; Napolitano, G. The Role of the Transcription Factor Nuclear Factor-kappa B in Thyroid Autoimmunity and Cancer. Front. Endocrinol. 2018, 9, 471. [Google Scholar] [CrossRef]
- Byun, J.Y.; Yune, T.Y.; Lee, J.Y.; Yeo, S.G.; Park, M.S. Expression of CYLD and NF-kappaB in human cholesteatoma epithelium. Mediat. Inflamm. 2010, 2010, 796315. [Google Scholar] [CrossRef] [PubMed]
- Fukudome, S.; Wang, C.; Hamajima, Y.; Ye, S.; Zheng, Y.; Narita, N.; Sunaga, H.; Fujieda, S.; Hu, X.; Feng, L.; et al. Regulation of the angiogenesis of acquired middle ear cholesteatomas by inhibitor of DNA binding transcription factor. JAMA Otolaryngol. Head Neck Surg. 2013, 139, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Duffy, A.M.; Bouchier-Hayes, D.J.; Harmey, J.H. Vascular Endothelial Growth Factor (VEGF) and Its Role in Non-Endothelial Cells: Autocrine Signalling by VEGF. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Ankamreddy, H.; Bok, J.; Groves, A.K. Uncovering the secreted signals and transcription factors regulating the development of mammalian middle ear ossicles. Dev. Dyn. 2020, 249, 1410–1424. [Google Scholar] [CrossRef]
- Wright, C.G. Development of the human external ear. J. Am. Acad. Audiol. 1997, 8, 379–382. [Google Scholar]
- Brito, J.M.; Teillet, M.A.; Le Douarin, N.M. Induction of mirror-image supernumerary jaws in chicken mandibular mesenchyme by Sonic Hedgehog-producing cells. Development 2008, 135, 2311–2319. [Google Scholar] [CrossRef]
- Chiang, C.; Litingtung, Y.; Lee, E.; Young, K.E.; Corden, J.L.; Westphal, H.; Beachy, P.A. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 1996, 383, 407–413. [Google Scholar] [CrossRef]
- Chole, R.A. The molecular biology of bone resorption due to chronic otitis media. Ann. N. Y. Acad. Sci. 1997, 830, 95–109. [Google Scholar] [CrossRef]
- Morales, D.S.; Penido Nde, O.; da Silva, I.D.; Stávale, J.N.; Guilherme, A.; Fukuda, Y. Matrix metalloproteinase 2: An important genetic marker for cholesteatomas. Braz. J. Otorhinolaryngol. 2007, 73, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Juhász, A.; Sziklai, I.; Rákosy, Z.; Ecsedi, S.; Adány, R.; Balázs, M. Elevated level of tenascin and matrix metalloproteinase 9 correlates with the bone destruction capacity of cholesteatomas. Otol. Neurotol. 2009, 30, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Galis, Z.S.; Khatri, J.J. Matrix metalloproteinases in vascular remodeling and atherogenesis: The good, the bad, and the ugly. Circ. Res. 2002, 90, 251–262. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Patel, K.D. Matrix metalloproteinase-2 (MMP-2) and MMP-9 in pulmonary pathology. Exp. Lung Res. 2005, 31, 599–621. [Google Scholar] [CrossRef] [PubMed]
- Nikolov, A.; Popovski, N. Role of Gelatinases MMP-2 and MMP-9 in Healthy and Complicated Pregnancy and Their Future Potential as Preeclampsia Biomarkers. Diagnostics 2021, 11, 480. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G.; Brekken, R.; McMahon, G.; Vu, T.H.; Itoh, T.; Tamaki, K.; Tanzawa, K.; Thorpe, P.; Itohara, S.; Werb, Z.; et al. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat. Cell Biol. 2000, 2, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Ezhilarasan, R.; Jadhav, U.; Mohanam, I.; Rao, J.S.; Gujrati, M.; Mohanam, S. The hemopexin domain of MMP-9 inhibits angiogenesis and retards the growth of intracranial glioblastoma xenograft in nude mice. Int. J. Cancer 2009, 124, 306–315. [Google Scholar] [CrossRef]
- Givvimani, S.; Tyagi, N.; Sen, U.; Mishra, P.K.; Qipshidze, N.; Munjal, C.; Vacek, J.C.; Abe, O.A.; Tyagi, S.C. MMP-2/TIMP-2/TIMP-4 versus MMP-9/TIMP-3 in transition from compensatory hypertrophy and angiogenesis to decompensatory heart failure. Arch. Physiol. Biochem. 2010, 116, 63–72. [Google Scholar] [CrossRef]
- Schönermark, M.; Mester, B.; Kempf, H.G.; Bläser, J.; Tschesche, H.; Lenarz, T. Expression of matrix-metalloproteinases and their inhibitors in human cholesteatomas. Acta Oto-Laryngol. 1996, 116, 451–456. [Google Scholar] [CrossRef]
- Suchozebrska-Jesionek, D.; Szymański, M.; Kurzepa, J.; Gołabek, W.; Stryjecka-Zimmer, M. Gelatinolytic activity of matrix metalloproteinases 2 and 9 in middle ear cholesteatoma. J. Otolaryngol. Head Neck Surg. 2008, 37, 628–632. [Google Scholar]
- Pilmane, M.; Shine, J.; Iismaa, T.P. Distribution of galanin immunoreactivity in the bronchi of humans with tuberculosis. Ann. N. Y. Acad. Sci. 1998, 863, 445–449. [Google Scholar] [CrossRef]
- Moon, S.K.; Lee, H.Y.; Li, J.D.; Nagura, M.; Kang, S.H.; Chun, Y.M.; Linthicum, F.H.; Ganz, T.; Andalibi, A.; Lim, D.J. Activation of a Src-dependent Raf-MEK1/2-ERK signaling pathway is required for IL-1alpha-induced upregulation of beta-defensin 2 in human middle ear epithelial cells. Biochim. Biophys. Acta 2002, 1590, 41–51. [Google Scholar] [CrossRef]
- Wehkamp, K.; Schwichtenberg, L.; Schröder, J.M.; Harder, J. Pseudomonas aeruginosa- and IL-1beta-mediated induction of human beta-defensin-2 in keratinocytes is controlled by NF-kappaB and AP-1. J. Investig. Dermatol. 2006, 126, 121–127. [Google Scholar] [CrossRef]
- Kanda, N.; Kamata, M.; Tada, Y.; Ishikawa, T.; Sato, S.; Watanabe, S. Human β-defensin-2 enhances IFN-γ and IL-10 production and suppresses IL-17 production in T cells. J. Leukoc. Biol. 2011, 89, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Yetiser, S.; Satar, B.; Aydin, N. Expression of epidermal growth factor, tumor necrosis factor-alpha, and interleukin-1alpha in chronic otitis media with or without cholesteatoma. Otol. Neurotol. 2002, 23, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Artono; Surarto, B.; Purnami, N.; Hutahaen, F.; Mahardhika, M.R. The Association of IL-1 Alpha Level and TNF Alpha Expressions on Bone Destruction in Chronic Suppurative Otitis Media and Cholesteatoma. Indian. J. Otolaryngol. Head Neck Surg. 2020, 72, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kusano, K.; Miyaura, C.; Inada, M.; Tamura, T.; Ito, A.; Nagase, H.; Kamoi, K.; Suda, T. Regulation of matrix metalloproteinases (MMP-2, -3, -9, and -13) by interleukin-1 and interleukin-6 in mouse calvaria: Association of MMP induction with bone resorption. Endocrinology 1998, 139, 1338–1345. [Google Scholar] [CrossRef]
- Mertz, P.M.; DeWitt, D.L.; Stetler-Stevenson, W.G.; Wahl, L.M. Interleukin 10 suppression of monocyte prostaglandin H synthase-2. Mechanism of inhibition of prostaglandin-dependent matrix metalloproteinase production. J. Biol. Chem. 1994, 269, 21322–21329. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, H.S. The anti-inflammatory role of tissue inhibitor of metalloproteinase-2 in lipopolysaccharide-stimulated microglia. J. Neuroinflamm. 2014, 11, 116. [Google Scholar] [CrossRef]
- Sikka, K.; Sharma, S.C.; Thakar, A.; Dattagupta, S. Evaluation of epithelial proliferation in paediatric and adult cholesteatomas using the Ki-67 proliferation marker. J. Laryngol. Otol. 2012, 126, 460–463. [Google Scholar] [CrossRef]
- Bujía, J.; Holly, A.; Antolí-Candela, F.; Tapia, M.G.; Kastenbauer, E. Immunobiological peculiarities of cholesteatoma in children: Quantification of epithelial proliferation by MIB1. Laryngoscope 1996, 106, 865–868. [Google Scholar] [CrossRef]
- Chung, J.H.; Lee, S.H.; Park, C.W.; Kim, K.R.; Tae, K.; Kang, S.H.; Oh, Y.H.; Pyo, J.Y. Expression of Apoptotic vs Antiapoptotic Proteins in Middle Ear Cholesteatoma. Otolaryngol. Head Neck Surg. 2015, 153, 1024–1030. [Google Scholar] [CrossRef]
- Akdogan, V.; Yilmaz, I.; Canpolat, T.; Ozluoglu, L.N. Role of Langerhans cells, Ki-67 protein and apoptosis in acquired cholesteatoma: Prospective clinical study. J. Laryngol. Otol. 2013, 127, 252–259. [Google Scholar] [CrossRef]
- Hamajima, Y.; Komori, M.; Preciado, D.A.; Choo, D.I.; Moribe, K.; Murakami, S.; Ondrey, F.G.; Lin, J. The role of inhibitor of DNA-binding (Id1) in hyperproliferation of keratinocytes: The pathological basis for middle ear cholesteatoma from chronic otitis media. Cell Prolif. 2010, 43, 457–463. [Google Scholar] [CrossRef]
- Shuman Moss, L.A.; Jensen-Taubman, S.; Stetler-Stevenson, W.G. Matrix metalloproteinases: Changing roles in tumor progression and metastasis. Am. J. Pathol. 2012, 181, 1895–1899. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, X.; Ai, L.; Yuan, R.; Ye, J. Clinicohistopathological implications of MMP/VEGF expression in retinoblastoma: A combined meta-analysis and bioinformatics analysis. J. Transl. Med. 2019, 17, 226. [Google Scholar] [CrossRef] [PubMed]
- Olszewska, E.; Chodynicki, S.; Chyczewski, L. Znaczenie angiogenezy w patogenezie perlaka ucha środkowego u dorosłych [Role of angiogenesis in the pathogenesis of cholesteatoma in adults]. Otolaryngol. Pol. 2004, 58, 559–563. [Google Scholar]
- Viac, J.; Palacio, S.; Schmitt, D.; Claudy, A. Expression of vascular endothelial growth factor in normal epidermis, epithelial tumors and cultured keratinocytes. Arch. Dermatol. Res. 1997, 289, 158–163. [Google Scholar] [CrossRef]
- Ma, J.; Tian, L.; Cheng, J.; Chen, Z.; Xu, B.; Wang, L.; Li, C.; Huang, Q. Sonic hedgehog signaling pathway supports cancer cell growth during cancer radiotherapy. PLoS ONE 2013, 8, e65032. [Google Scholar] [CrossRef]
- de Carvalho Dornelles, C.; da Costa, S.S.; Meurer, L.; Rosito, L.P.; da Silva, A.R.; Alves, S.L. Comparison of acquired cholesteatoma between pediatric and adult patients. Eur. Arch. Otorhinolaryngol. 2009, 266, 1553–1561. [Google Scholar] [CrossRef]
- Banerjee, A.R.; James, R.; Narula, A.A. Matrix metalloproteinase-2 and matrix metalloproteinase-9 in cholesteatoma and deep meatal skin. Clin. Otolaryngol. Allied Sci. 1998, 23, 345–347. [Google Scholar] [CrossRef] [PubMed]
- Rezende, C.E.; Souto, R.P.; Rapoport, P.B.; Campos, L.D.; Generato, M.B. Cholesteatoma gene expression of matrix metalloproteinases and their inhibitors by RT-PCR. Braz. J. Otorhinolaryngol. 2012, 78, 116–121. [Google Scholar] [CrossRef]
- Kaya, İ.; Avcı, Ç.B.; Şahin, F.F.; Özateş, N.P.; Sezgin, B.; Kurt, C.Ç.; Bilgen, C.; Kirazlı, T. Evaluation of significant gene expression changes in congenital and acquired cholesteatoma. Mol. Biol. Rep. 2020, 47, 6127–6133. [Google Scholar] [CrossRef] [PubMed]
- Sun, J. Matrix metalloproteinases and tissue inhibitor of metalloproteinases are essential for the inflammatory response in cancer cells. J. Signal Transduct. 2010, 2010, 985132. [Google Scholar] [CrossRef]
- Preciado, D.A. Biology of cholesteatoma: Special considerations in pediatric patients. Int. J. Pediatr. Otorhinolaryngol. 2012, 76, 319–321. [Google Scholar] [CrossRef]









| Grading Scale | Explanation of Grading Scale | Percentage of Factor-Positive Cells in the Visual Field |
|---|---|---|
| 0 | No positive structures | 0% |
| 0/+ | Occasional positive structures | 12.5% |
| + | Few positive structures | 25% |
| +/++ | Few-to-moderate number of positive structures | 37.5% |
| ++ | Moderate number of positive structures | 50% |
| ++/+++ | Moderate-to-numerous positive structures | 62.5% |
| +++ | Numerous positive structures | 75% |
| +++/++++ | Numerous-to-abundant structures | 87.5% |
| ++++ | An abundance of positive structures in the visual field | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dambergs, K.; Sumeraga, G.; Pilmane, M. Comparison of Tissue Factors in the Ontogenetic Aspects of Human Cholesteatoma. Diagnostics 2024, 14, 662. https://doi.org/10.3390/diagnostics14060662
Dambergs K, Sumeraga G, Pilmane M. Comparison of Tissue Factors in the Ontogenetic Aspects of Human Cholesteatoma. Diagnostics. 2024; 14(6):662. https://doi.org/10.3390/diagnostics14060662
Chicago/Turabian StyleDambergs, Kristaps, Gunta Sumeraga, and Māra Pilmane. 2024. "Comparison of Tissue Factors in the Ontogenetic Aspects of Human Cholesteatoma" Diagnostics 14, no. 6: 662. https://doi.org/10.3390/diagnostics14060662
APA StyleDambergs, K., Sumeraga, G., & Pilmane, M. (2024). Comparison of Tissue Factors in the Ontogenetic Aspects of Human Cholesteatoma. Diagnostics, 14(6), 662. https://doi.org/10.3390/diagnostics14060662